


- Portada
- N° 192 (mai 2014)
- les nardaies montagnardes À meum athamanticum du plateau d’ELSENBORN (EST DE LA Belgique). Évolution spontanÉe depuis l’arrÊt des pratiques agropastorales. Impact de la gestion conservatoire par la fauche ou la mise À feu contrÔlÉe


Vista(s): 1291 (19 ULiège)
Descargar(s): 135 (3 ULiège)

les nardaies montagnardes À meum athamanticum du plateau d’ELSENBORN (EST DE LA Belgique). Évolution spontanÉe depuis l’arrÊt des pratiques agropastorales. Impact de la gestion conservatoire par la fauche ou la mise À feu contrÔlÉe

Documento adjunto(s)
Version PDF originaleRésumé
Les nardaies montagnardes à Meum athamanticum du plateau d’Elsenborn (Est de la Belgique). Évolution spontanée depuis l’arrêt des pratiques agropastorales. Impact de la gestion conservatoire par la fauche ou la mise à feu contrôlée.−De vastes surfaces de nardaies montagnardes apparaissaient sur le plateau d’Elsenborn. Cet habitat, riche en espèces rares, résulte de plusieurs siècles de pratiques agropastorales par fauchage et pâturage occasionnel du regain dans les bas de versant des vallées. Abandonnées depuis la fin des années 1940, de grandes surfaces de cet habitat ont été converties en prairies intensives ou en plantations d’épicéa. Les nardaies reliques ont quant à elles évolué. Leur richesse botanique a diminué et elles sont aujourd’hui dominées par des graminées. Cette note a pour but de faire la synthèse de leur évolution depuis leur abandon, d’en décrire les modifications floristiques et d’estimer l’impact des mesures de gestion par fauchage ou par mises à feu contrôlées qui leur sont appliquées. Tous les relevés phytosociologiques publiés décrivant l’habitat sur le plateau d’Elsenborn, auxquels nous avons ajouté 165 relevés inédits, ont été analysés et soumis à des analyses statistiques multivariées. Les résultats montrent la régression progressive des espèces caractéristiques de l’habitat et l’enrichissement du milieu en azote, en relation avec l’ancienneté de l’abandon des parcelles.La gestion par fauchage ou par mise à feu contrôlée permet d’enrailler la dégradation de l’habitat et favorise le retour des espèces caractéristiques. Des différences apparaissent toutefois dans la composition floristique des parcelles gérées, en fonction du mode de gestion appliqué.
Abstract
Nardus grasslands from the Elsenborn plateau (East Belgium). Spontaneous evolution since the stop of traditional agriculture. Effects of management by mowing or controlled burning. − Large surfaces of montain Nardus grassland occured in the Elsenborn plateau. This habitat, rich in rare species, was previously maintained by traditional agriculture practices (annual mowing and occasional grazing of the aftermath). Abandoned since the end of the years 1940, large areas of this habitat have been converted into intensive meadows or spruce plantations. The remaining Nardus grasslands had evolved. Their botanic richness had decreased and nowadays they are dominated by coarse grasses. This paper aims to follow vegetation dynamic of the habitat since it was neglected, to describe its floristic alteration and to assess the impact of its restoration and management by mowing or controlled burning. All the published Nardus grasslands phytosociological data from the Elsenborn region and 165 new unpublished data from the same area (using the Braun-Blanquet method) have been analyzed and submitted to multivariate analysis using the « R package ». Results show a gradual decrease of the Nardus grasslands typical species and an increase in nitrogen availability of their soil, in relation with the oldness of their abandon. Management by mowing or burning promotes the restoration of the original habitat and increases species richness, but some differences appear in the floristic composition of the restored grasslands, according to the management techniques used.
Zusammenfassung
Die montanen bärwurzreichen Borstgrasflächen der Elsenborner Hochebene (Ostbelgien). Spontane Entwicklung seit der Aufgabe der agropastoralen Bewirtschaftung. Einfluss der Pflege durch Mahd oder kontrolliertes Abbrennen. − Auf der Hochebene Elsenborns befinden sich großflächige montane bärwurzreiche Bostgrasflächen. Dieser Lebensraum, sehr reich an seltenen Arten, ist das Ergebnis jahrzehntelanger agropastoraler Nutzung durch Mähen und gelegentlicher Beweidung des Grummet in den unteren Hangbereichen der Täler. Ungenutzt seit Ende der Jahre 1940 wurden viele dieser Flächen in intensiv genutzte Wiesen oder in Fichten-anpflanzungen umgewandelt. Die noch verbleibenden Bostgrasrasen haben sich verändert. Ihre botanische Vielfalt hat abgenommen und sie sind heute dominiert durch Gräser. Dieser Bericht beabsichtigt eine Synthese der Entwicklung dieses Lebensraums seit der Aufgabe der Nutzung zu erstellen, die floristische Veränderung zu beschreiben und den Einfluss der Bewirtschaftungsmaßnahmen durch Mahd oder kontrolliertes Abbrennen zu beschreiben. Alle veröffentlichten phytosoziologischen Erhebungen welche die Lebensräume auf der Elsenborner Hochebene beschreiben, ergänzt durch 165 nicht veröffentlichte Erhebungen, wurden analysiert und einer statistischen multivariaten Analyse unterworfen. Die Resultate zeigen den progressiven Rückgang der charakteristischen Arten dieses Lebensraumes und die Stickstoffanreicherung des Milieus, in Verbindung mit der Dauer des Brachliegens der Fläche. Die Bewirtschaftung durch Mahd oder kontrolliertes Abbrennen ermöglicht den Verfall des Lebensraumes zu beenden und favorisiert die Rückkehr der charakteristischen Arten. In Abhängigkeit der angewandten Bewirtschaftungs-methode erscheinen aber Unterschiede in der floristischen Zusammen- setzung der gepflegten Parzellen.
11. INTRODUCTION
2Les nardaies montagnardes à Meum athamanticum sont des pelouses acidophiles dominées par des plantes frugales, localisées dans les bas de versants des vallées de la haute Ardenne, principalement dans sa partie la plus septentrionale. Elles trouvent leur optimum sur les terrains du Dévonien infé-rieur (Eodévonien) du plateau d’Elsenborn, caractérisés par des roches siliceuses des étages Praguien et Emsien particulièrement pauvres en carbonates de calcium et de magnésium. Ces formations ont été utilisées depuis le XVème siècle comme prés maigres de fauche, non ou peu amendés, dans lesquels on pratiquait l’abîssage.
3Il s’agit d’un groupement d’une grande richesse floristique et qui comporte un grand nombre d’espèces rares, dont beaucoup à caractère montagnard plus ou moins accusé. Décrit pour la première fois par Schwickerath en 1944, il a ensuite été étudié en Allemagne par Klapp (1951) dans le nord-ouest de l’Eifel, à proximité de la frontière belge, Baur (1967) dans la Forêt Noire, Reichert (1972) dans le Hunsrück et Matzke (1985) dans l’Eifel et le Hunsrück, et en Belgique par Schumacker (1975), Ros (1980), BodarwÉ (1991) et Frankard & al. (1998) sur le plateau d’Elsenborn et par Sougnez (1977) en Ardenne septentrionale.
4Schwickerath a décrit ce groupement sous la dénomination d’Arnicetum montanae. Klapp (1951) le renomme Meo athamantici-Centaureetum nigrae, en raison de la combinaison quasi constante de Meum athamanticum et de Centaurea jacea subsp. nigra dans la végétation. Cette dénomination a par la suite été reprise par Reichert (1972), Schumacker (1975) et Sougnez (Centaureo nigrae-Meetum athamantici 1977). Stieperaere (1990) et Oberdorfer (1993) estiment que le groupement doit être rangé dans le Festuco-Genistetum sagittalis, appartenant à l’alliance du Violion caninae, dont les sous-associations et variantes restent à définir.
5Quoi qu’il en soit, les pelouses à Meum athamanticum relèvent incontestablement des Nardo-Callunetea Prsg. 1949, par la présence presque constante de Luzula multiflora, Danthonia decumbens, Potentilla erecta et Carex pilulifera. Ce sont les espèces des Nardetalia et du Violion caninae qui dominent en nombre et en recouvrement. Cependant, dans son expression optimale, ce groupement s’enrichit de nombreuses espèces des Molinietalia et des Arrhenatheretalia (Schumacker, 1975).
6Schumacker (1975) a distingué trois sous-associations dans les nardaies à Meum athamanticum du plateau d’Elsenborn :
7- une sous-association type, présente sur les colluvions limono-caillouteuses des bas de versants et parfois au contact de la plaine alluviale (où le groupement est souvent riche en Narcissus pseudonarcissus);
8- une sous-association hygrophile des sols plus frais, présente souvent sur les alluvions humides des replats des plaines alluviales, caractérisée par l’abondance et la fréquence de Deschampsia cespitosa et de Persicaria bistorta;
9- une sous-association thermophile des sols plus secs, exposés en plein sud sur des bas de pentes abrités ou sur des talus en lisières résiduelles de la chênaie de versant, caractérisée par l’abondance et la fréquence de Genistella sagittalis et d’Helianthemum nummularium subsp. obscurum.
10Les nardaies montagnardes ont couvert de vastes superficies sur le plateau d'Elsenborn, mais elles y ont considérablement régressé ces 70 dernières années. Aujourd'hui, elles ne sont plus abondantes que dans le champ de tir du camp militaire d’Elsenborn (où il en subsiste environ 500 ha, dont beaucoup font l’objet d’une gestion récurrente principalement par mises à feu contrôlées) et dans les vallées périphériques du camp militaire, où elles sont progressivement classées en réserves naturelles depuis les années 1990 et où elles font aujourd’hui l’objet de mesures de restauration et d’entretien par fauchage avec exportation du foin. Ailleurs, il n'en subsiste que de rares lambeaux ayant échappé à l'enrésinement et à l'agriculture intensive.
11En dehors du camp militaire, ces nardaies reliques étaient pour la plupart abandonnées depuis la fin de la seconde guerre mondiale. Suite au manque d’entretien, leur végétation a évolué et leur richesse botanique a diminué. Dès 1974, Schumacker & Duvigneaud mettaient en évidence la régression des espèces en rosettes liées aux pelouses rases et l’extension de graminées de grande taille, d’espèces indicatrices d’un certain enrichissement en azote du substrat ainsi que d’espèces des lisières forestières dans les nar-daies montagnardes thermophiles (Schumacker & Duvigneaud, 1974). Cette évolution, imputable à l’abandon des anciennes pratiques agro-pastorales, s’est poursuivie jusqu’à aujourd’hui. Elle semble toutefois bien moins prononcée dans la variante thermophile du groupement que dans les variantes types et hygrophiles (Ros, 1980 ; Frankard & al., 1998). Ces der-nières ont en effet subi d’importants changements depuis les années 1970, avec notamment la forte progression de graminées croissant en touradons, comme Deschampsia cespitosa, Molinia caerulea, Poa chaixii, au point qu'il est souvent difficile aujourd'hui de faire la distinction entre ces deux variantes (Frankard & al., 1998).
12Le but de cette note est de faire la synthèse de l’évolution des nardaies montagnardes à Meum athamanticum du plateau d'Elsenborn depuis leur abandon vers 1945, d’en décrire les modifications floristiques et d’estimer l’impact des mesures conservatoires de gestion qui leur sont appliquées, soit par fauchage depuis 1993 dans les réserves naturelles domaniales des vallées de la Schwalm et de l’Olef, soit par mises à feu contrôlées réalisées à intervalles plus ou moins réguliers dans le champ de tir du camp militaire d’Elsenborn.
132. CARACTÉRISTIQUES DE LA RÉGION ÉTUDIÉE
142.1. Caractéristiques physiques
15Le plateau d’Elsenborn est situé en Ardenne nord-orientale à la frontière germano-belge, au sud-est du plateau des Hautes-Fagnes et au nord-ouest de l’Eifel. Il est compris entre 50° 22' et 50° 30' de latitude Nord et entre 6° 10' et 6° 20' de longitude Est. L’altitude varie entre 520 et 689 mètres. Les sommets du plateau d’Elsenborn culminent à 689 m à Weisserstein, 650 m à Mürringen, 640 m à Rocherath et 630 m à Elsenborn. Ce plateau est découpé par des vallées profondes, comprenant de nombreux ruisseaux et rivières qui alimentent les bassins de la Roer au nord, de l’Amblève à l’ouest et de l’Our au sud.
16Le climat y est froid (température moyenne annuelle 6,5° C) et pluvieux (1300 mm de précipitations annuelles moyennes). Le mésoclimat des fonds de vallées est particulièrement rigoureux, les températures moyennes mensuelles y étant toute l’année inférieure à celle des plateaux, avec une différence plus marquée en été (± 2° C) qu’en hiver (± 0,5° C) (Deflandre, 1977). Cette différence s’explique entre autres par l’existence de vents froids catabatiques qui s’écoulent lentement le long des versants pour s’accumuler momentanément dans les fonds de vallées. Dans ceux-ci, des gelées nocturnes peuvent être enregistrées au sol toute l’année.
17Le plateau d’Elsenborn est situé sur des roches primaires de l’Eodévonien du synclinal de l’Eifel, en particulier sur les assises du Praguien et de l’Emsien. Il s’agit de roches siliceuses (schistes phylladeux, grès) particulièrement pauvres en carbonates de calcium et de magnésium, l’Emsien en étant toutefois mieux pourvu que le Praguien. Les sols sont principalement de type limono-caillouteux bien drainés, à charge schisto-phylladeuse sur le Praguien, à charge schisto-gréseuse sur l’Emsien (Frankard & al., 1998).
182.2. Evolution du territoire depuis le Moyen Age jusqu’à nos jours
19Avant que l’homme ne s’y installe, la région d’Elsenborn était couverte de forêts, essentiellement de la hêtraie à luzule blanche (Luzulo albidae-Fagetum), les sols étant naturellement bien drainés.
20Il est possible que les premières actions anthropiques (pratiques agropastorales extensives) sur le plateau d’Elsenborn datent du XIIIème siècle (Ros, 1980). Entre les XIIIème et XVème siècles, les fonds de vallées (plaines alluviales et bas de versants) ont été défrichés, puis ont été utilisés comme prés de fauche ou pâturés. Ces maigres herbages ont été progressivement améliorés à partir du XVème siècle par la pratique de l’abîssage. Cette ancienne technique culturale permettait de fertiliser légèrement les prés peu productifs de bas de versants par alluvionnement, via l’épandage d’eau captée des ruisseaux ou de sources durant la morte saison (de novembre à mai), au moyen de canaux de dérivation creusés le long des versants, parallèlement aux courbes de niveau (Fontaine & al., 1981). Ces prés servaient à produire du foin pour le bétail en stabulation pendant l’hiver.
21Les cartes de Ferraris (1780) montrent qu’à la fin du XVIIIème siècle tous les fonds de vallées étaient défrichés, tandis que les sommets et les versants étaient couverts de forêts feuillues ou de landes à callune. Par la suite, la limite entre le domaine forestier et le domaine rural a très peu changé, l’actuelle étant sensiblement pareille à l’ancienne, la grande différence étant le remplacement des feuillus par des monocultures de résineux en maints endroits et la transformation des milieux semi-naturels ouverts (landes et pelouses) en prairies artificielles ou pâtures permanentes.
22Jusqu’à la première guerre mondiale, les nardaies montagnardes ne reçurent aucun amendement, l’abîssage étant le seul moyen mis en œuvre pour améliorer les sols. Ensuite, entre la première et la seconde guerre mondiales, l’utilisation des prés de fauche fut progressivement abandonnée, l’agriculture se concentrant dans les parcelles les plus proches des exploitations, où l’emploi intensif d’engrais s’est généralisé.
23Aujourd’hui, les nardaies de bas de versants à Meum athamanticum ont considérablement régressé, beaucoup ayant été détruites, transformées le plus souvent en plantations de résineux ou en prairies intensives. Elles sont surtout encore présentes dans des sites qui ont été classés en réserves naturelles domaniales (vallées de la Schwalm, de l’Olef et de la Rur) ou privées (vallée de la Holzwarche), ainsi que dans les fonds de vallées du camp militaire d’Elsenborn, où le paysage est resté tel qu’il était lors de la création du camp en 1893.
243. MÉTHODOLOGIE
25Afin de caractériser au mieux l’évolution des nardaies à Meum athamanticum du plateau d'Elsenborn, nous avons repris pour les analyses l’ensemble des relevés phytosociologiques décrivant l’habitat, publiés par Schwickerath (15 relevés, 1944), Klapp (10 relevés, 1951), Schumacker (12 relevés, 1975), Ros (58 relevés, 1980) et Frankard & al. (42 relevés, 1998). A ces données, ont été ajoutés 165 relevés inédits réalisés : 1. dans des zones n’ayant plus fait l’objet de gestion depuis longtemps (43 relevés) ; 2. dans des zones gérées par fauchage depuis une dizaine d’années dans la réserve naturelle domaniale de la vallée de la Schwalm (24 relevés) ; 3. dans des zones gérées par mises à feu plus ou moins régulières dans le camp militaire d’Elsenborn (98 relevés). Un total de 302 relevés ont par conséquent été soumis à l’analyse, afin de décrire l’évolution spontanée des nardaies à Meum athamanticum et d’évaluer l’impact des mesures de gestion conservatoire qui leur sont appliquées.
26Pour faciliter le travail de confrontation des divers relevés, ceux-ci ont été rassemblés par époque ou par type de gestion. Les relevés ont été regroupés comme suit :
-
les données anciennes de Schwickerath (1944) et Klapp (1951) récoltées entre 1933 et 1938 pour Schwickerath et fin des années 1940 pour Klapp. Ces données peuvent servir de référence, car elles ont été récoltées à une époque où s’exerçaient encore les activités agropastorales (l’abandon de celles-ci datant d’après la seconde guerre mondiale);
-
les données des années 1970 de zones abandonnées (Schumacker, 1975 et Ros, 1980);
-
les données actuelles de zones abandonnées (Frankard & al., 1998 et inédits);
-
les données actuelles des zones gérées par fauchage avec enlèvement du foin;
-
les données actuelles des zones soumises à des mises à feu plus ou moins régulières (en général 2-10 ans).
27Le degré de rareté de la flore des nardaies à Meum athamanticum a été estimé en faisant appel à l'échelle de rareté de Stieperaere & Fransen (1982) établie pour le sud de la Belgique, sur base des données de l'atlas floristique de Van Rompaey & Delvosalle (1979). Cette analyse a été réalisée sur l'ensemble des relevés, puis sur les différents groupes de relevés définis.
28Les relevés phytosociologiques ont été traduits de façon synthétique, d’une part par le degré de présence de chacune des espèces recensées et d’autre part par leur coefficient de recouvrement. Ce dernier mode de représentation synthétique donne une image plus élaborée de la végétation, puisqu’il chiffre l’importance relative de chacune des espèces présentes dans le groupement végétal (Vanden Berghen, 1982). Seules les espèces les plus constantes ou les plus représentatives de l’association sont inscrites dans les tableaux. Les espèces sporadiques ont été éliminées pour plus de lisibilité.
29Des spectres écologiques pondérés, exprimés par les plantes, sur base des indices d’Ellenberg (Ellenberg & al., 1992) de teneur azotée du sol (N), d’acidité du sol (R) et d’ouverture du milieu (L) ont été dressés pour les données floristiques de chaque époque. Pour établir ces spectres pondérés, il a été nécessaire de convertir les coefficients d’abondance-dominance en quantités moyennes selon la méthode exposée par Vanden Berghen (1982). Ce type de représentation est plus raffiné qu’un spectre brut, car il tient compte non seulement de la présence des différentes espèces mais aussi de leur importance dans le couvert. Tous les relevés phytosociologiques ont ensuite été soumis à des analyses statistiques multivariées : à une méthode de groupement (indice de similarité de Steinhaus, mesure de la distance de Bray-Curtis et classification hiérarchique agglomérative par la méthode Ward) après transformation des coefficients d’abondance-dominance selon l’échelle de van der Maarel (1979), puis à une ordination par analyse en composantes principales (ACP). Le principe de cette méthode de groupement est de rassembler les relevés qui ont un degré de similarité suffisant pour être réunis dans le même ensemble. Ensuite, ces mêmes groupes et la hiérarchie produite ont été soumis à la méthode Indval pour déterminer les espèces indicatrices des différents groupes identifiés par la méthode Ward (DufrÊne & Legendre 1997). Le principe de la méthode Indval repose sur la définition du caractère indicateur d’une espèce : une espèce est considérée comme indicatrice si elle est typique d’un groupe de relevés (spécificité) et si elle est présente dans tous les relevés de ce groupe (fidélité). La valeur indicatrice de l’espèce pour un niveau de la classification des relevés en différents groupes est la plus large valeur d’Indval observée pour un des groupes. La valeur d’Indval est maximale (100%) quand la spécificité et la fidélité sont maximales. Comme cet indice peut être calculé pour les différents niveaux de la procédure de groupement, il est possible d’identifier le niveau où l’espèce est la plus indicatrice. Il est donc possible d’identifier rapidement les espèces géné-ralistes et les espèces spécialisées. La signification de la valeur indicatrice des espèces pour les différents niveaux est contrôlée par une procédure de permutation aléatoire (1000 permutations, P = 0,05). Le seuil de la valeur indicatrice de l’indice Indval est de 25%. Ce niveau correspond à une valeur minimale de 50% pour la spécificité et de 50% pour la fidélité.
30Les différentes analyses multivariées ont été menées à l’aide du logiciel R (version 2.15.2). La nomenclature adoptée suit celle de la "Nouvelle Flore de la Belgique, du Grand-Duché de Luxembourg, du Nord de la France et des Régions voisines (Ptéridophytes et Spermatophytes)" (Lambinon & al., 2012).
314. RESULTATS
324.1. Composition floristique des nardaies montagnardes à Meum athamanticum
33131 espèces ont été comptabilisées dans les relevés. Sur base des indices de Stieperaere & Fransen (1982) pour le sud du sillon Sambre-et-Meuse (tableau 1), il apparaît qu'un tiers de la flore totale des nardaies à Meum athamanticum relève des degrés de rareté 0 à 3 (RR-AR) et que 31 espèces (24 % du total des espèces) sont considérées comme franchement rares à très rares (l'une d'entre-elles, Pseudorchis albida, était même con-sidérée comme disparue du territoire, mais a été retrouvée en 2003) : Antennaria dioica*, Arnica montana*, Botrychium lunaria*, Calamagrostis arundinacea, Carex pulicaris*,Centaurea montana, Euphrasia nemorosa, Euphrasia officinalis subsp. rostkoviana, Festuca filiformis, Festuca rubra subsp. commutata, Galium pumilum, Genista anglica, Genista pilosa, Genistella sagittalis, Geranium sylvaticum, Gymnadenia conopsea*, Hieracium maculatum, Hypochaeris maculata*, Linaria repens, Meum athamanticum, Narcissus pseudonarcissus, Pedicularis sylvatica, Platanthera bifolia*, Pseudorchis albida*, Ranunculus serpens subsp. polyanthemoides*, Sanguisorba officinalis, Senecio hercynicus, Serratula tinctoria*, Thesium pyrenaicum*, Vaccinium vitis-idaea, Vicia orobus*.
|
Degré de rareté |
Belgique méridionale |
||
|
Nombre absolu |
% |
||
|
0 |
RR |
1 |
0,8 |
|
1 |
RR-R |
20 |
15,4 |
|
2 |
R |
10 |
7,7 |
|
3 |
R-AR |
12 |
9,2 |
|
4 |
AR |
8 |
6,2 |
|
5 |
AR-AC |
9 |
6,9 |
|
6 |
AC |
8 |
6,2 |
|
7 |
AC-C |
9 |
6,9 |
|
8 |
C |
15 |
11,5 |
|
9 |
C-CC |
13 |
10 |
|
10 |
CC |
25 |
19,2 |
34Tableau 1. Richesse floristique des nardaies à Meum athamanticum (d'après les données de Stieperaere & Fransen, 1982)
35Parmi ces espèces rares à très rares, on note 12 espèces inscrites dans la liste des plantes vasculaires protégées de Wallonie (notées * dans la liste qui précède). 14 espèces sont aussi reprises dans la liste rouge wallonne (Saintenoy-Simon & coll., 2006): 2 espèces vulnérables (Centaurea montana, Gymnadenia conopsea); 6 espèces en danger (Arnica montana, Carex pulicaris, Platanthera bifolia, Ranunculus serpens subsp. polyanthemoides, Sanguisorba officinalis, Serratula tinctoria) et 6 espèces menacées d'extinction (Antennaria dioica, Botrychium lunaria, Hypochaeris maculata, Pseudorchis albida, Thesium pyrenaicum, Vicia orobus). A l'autre extrémité du tableau, 41,5 %, des espèces sont communes à très communes.
36Le graphique 1 montre l'évolution du degré de rareté de la flore des nardaies à Meum athamanticum, en fonction des époques et de l'abandon ou de la gestion des parcelles. Il indique clairement que le nombre d'espèces très rares a fortement régressé depuis les années 1930-40, mais augmente quelque peu dans les milieux gérés (les plus rares apparaissant surtout dans les milieux incendiés, tandis qu'à l'autre extrémité du graphique les espèces ubiquistes progressent au fur et à mesure de l'ancienneté de l'abandon des parcelles, phénomène qui est enraillé par la gestion.
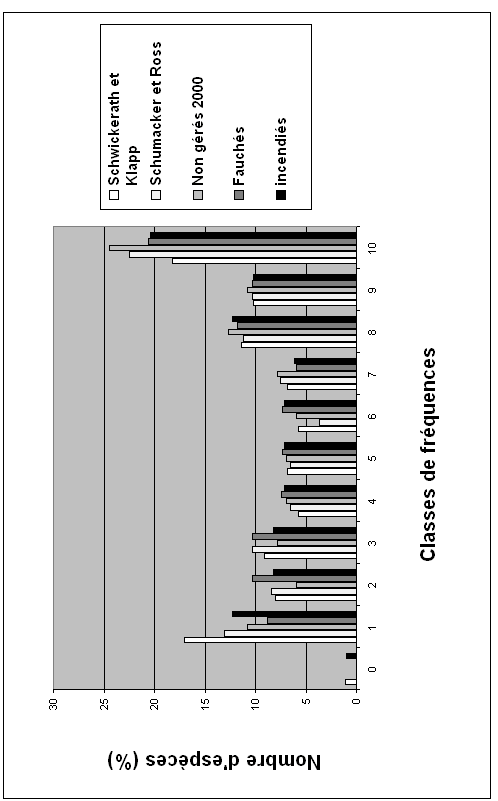
37Graphique 1. Evolution de la richesse en espèces rares des nardaies à Meum athamanticum en fonction de l’abandon plus ou moins ancien des parcelles et des mesures de gestion appliquées
384.2. Evolution de la végétation des nardaies à Meum athamanticum
39A la lecture du tableau 2, synthétisant l’ensemble des relevés disponibles, il apparaît que les nardaies à Meum athamanticum ont fortement évolué depuis l’abandon des pratiques agropastorales, leur composition floristique ayant subi d’importantes modifications.
|
Schwickerath & Klapp 1944-1951 |
Schumacker & Ross 1972-1981 |
Frankard et al. 1998 & Frankard inédits 2003-2004 |
Camp militaire géré par brûlage |
Schwalm géré par fauchage |
|||
|
1. Nardo-Callunetea |
|||||||
|
Meum athamanticum |
V (1840) |
V (3693) |
V (2906) |
V (4082) |
V (3698) |
||
|
Festuca rubra subsp. commutata |
V (169) |
IV (776) |
V (2309) |
V (1607) |
V (2958) |
||
|
Centaurea jacea subsp. nigra |
V (91) |
V (473) |
IV (205) |
V (386) |
IV (144) |
||
|
Potentilla erecta |
V (259) |
V (697) |
V (192) |
V (516) |
V (202) |
||
|
Lathyrus linifolius var. montanus |
V (90) |
V (372) |
V (182) |
V (99) |
V (146) |
||
|
Galium saxatile |
IV (139) |
IV (291) |
V (1021) |
II (71) |
V (742) |
||
|
Narcissus pseudonarcissus |
II (92) |
III (135) |
I (147) |
I (9) |
V (430) |
||
|
Carex pallescens |
I (2) |
II (22) |
I (5) |
I (1) |
I (2) |
||
|
Pimpinella saxifraga |
I (2) |
III (30) |
I (3) |
I (18) |
I (3) |
||
|
Thesium pyrenaicum |
I (3) |
I (156) |
I (6) |
I (9) |
. |
||
|
Carex caryophyllea |
I (1) |
II (24) |
I (1) |
I (1) |
. |
||
|
Cytisus scoparius |
I (121) |
I (11) |
I (1) |
I (21) |
. |
||
|
Helianthemum nummularium subsp. obscurum |
I (72) |
I (219) |
I (154) |
. |
. |
||
|
Vicia orobus |
I (2) |
I (11) |
I (3) |
||||
|
Nardus stricta |
V (493) |
II (315) |
II (192) |
V (404) |
V (156) |
||
|
Festuca filiformis |
V (674) |
IV (1282) |
II (123) |
V (427) |
V (155) |
||
|
Arnica montana |
V (646) |
III (154) |
I (1) |
IV (345) |
I (12) |
||
|
Danthonia decumbens |
V (248) |
III (141) |
I (49) |
IV (115) |
III (285) |
||
|
Hieracium pilosella |
V (160) |
II (261) |
I (19) |
I (7) |
II (35) |
||
|
Hypericum maculatum |
IV (104) |
IV (302) |
II (128) |
V (166) |
II (26) |
||
|
Luzula multiflora |
IV (60) |
IV (89) |
II (11) |
IV (83) |
IV (71) |
||
|
Calluna vulgaris |
IV (829) |
III (193) |
II (20) |
III (72) |
II (35) |
||
|
Polygala serpyllifolia |
IV (34) |
III (77) |
II (10) |
II (13) |
V (55) |
||
|
Ranunculus serpens |
IV (51) |
II (16) |
. |
III (9) |
. |
||
|
Genistella sagittalis |
III (195) |
II (194) |
II (149) |
I (14) |
III (56) |
||
|
Carex pilulifera |
III (20) |
II (25) |
I (2) |
III (73) |
IV (41) |
||
|
Thymus pulegioides |
II (16) |
II (162) |
I (71) |
I (3) |
I (1) |
||
|
Viola canina |
IV (12) |
III (85) |
I (4) |
II (7) |
I (3) |
||
|
Veronica officinalis |
II (13) |
II (22) |
I (1) |
. |
I (1) |
||
|
Vaccinium vitis-idaea |
II (16) |
I (122) |
I (1) |
I (18) |
. |
||
|
Genista anglica |
II (172) |
I (1) |
I (1) |
II (47) |
I (3) |
||
|
Genista pilosa |
II (82) |
I (8) |
. |
. |
. |
||
|
Linum catharticum |
II (4) |
I (<1) |
. |
I (1) |
I (2) |
||
|
Orchis mascula |
II (2) |
I (1) |
. |
I (1) |
. |
||
|
Hypochaeris maculata |
I (2) |
I (5) |
. |
I (8) |
. |
||
|
Botrychium lunaria |
I (2) |
. |
. |
. |
. |
||
|
Antennaria dioica |
I (2) |
. |
. |
. |
. |
||
|
Pseudorchis albida |
I (1) |
. |
. |
I (3) |
. |
||
|
Hieracium umbellatum |
I (3) |
. |
I (6) |
V (479) |
III (20) |
||
|
2. Molinio-Arrhenatheretea |
|||||||
|
Stachys officinalis |
IV (175) |
IV (274) |
V (388) |
IV (103) |
V (411) |
||
|
Sanguisorba officinalis |
IV (56) |
IV (221) |
IV (38) |
V (86) |
V (45) |
||
|
Rumex acetosa |
III (9) |
III (65) |
III (37) |
III (9) |
IV (21) |
||
|
Ranunculus acris |
III (26) |
II (58) |
II (15) |
I (1) |
II (27) |
||
|
Cirsium palustre |
II (5) |
III (26) |
II (32) |
I (3) |
II (4) |
||
|
Trifolium pratense |
II (144) |
I (10) |
I (93) |
I (1) |
. |
||
|
Centaurea jacea subsp. grandiflora |
II (97) |
I (<1) |
I (4) |
I (25) |
. |
||
|
Lotus corniculatus |
I (3) |
II (37) |
I (22) |
III (25) |
. |
||
|
Vicia cracca |
I (1) |
I (6) |
I (4) |
III (19) |
. |
||
|
Serratula tinctoria |
I (1) |
I (11) |
I (4) |
III (19) |
I (2) |
||
|
Heracleum sphondylium var. sphondylium |
I (1) |
I (9) |
I (1) |
I (1) |
. |
||
|
Leontodon autumnalis |
I (20) |
. |
I (3) |
I (6) |
I (2) |
||
|
Hypochaeris radicata |
I (21) |
. |
I (1) |
I (1) |
. |
||
|
Succisa pratensis |
V (168) |
IV (104) |
III (48) |
V (155) |
IV (205) |
||
|
Plantago lanceolata |
IV (147) |
II (12) |
I (21) |
I (9) |
I (1) |
||
|
Leucanthemum vulgare |
IV (40) |
III (96) |
I (1) |
I (2) |
II (5) |
||
|
Rhinanthus minor subsp. minor |
IV (60) |
II (52) |
I (33) |
III (42) |
IV (81) |
||
|
Phyteuma nigrum |
III (29) |
III (31) |
I (7) |
IV (48) |
III (10) |
||
|
Euphrasia officinalis subsp. rostkoviana |
II (13) |
I (4) |
I (3) |
. |
. |
||
|
Dactylorhiza maculata |
II (6) |
I (23) |
. |
I (1) |
II (14) |
||
|
Leontodon hispidus |
II (4) |
. |
I (1) |
I (44) |
. |
||
|
Pedicularis sylvatica |
I (2) |
I (<1) |
. |
. |
. |
||
|
Molinia caerulea |
I (12) |
II (169) |
III (729) |
IV (325) |
IV (430) |
||
|
Poa chaixii |
I (2) |
II (400) |
III (514) |
I (1) |
II (25) |
||
|
Deschampsia cespitosa |
. |
II (211) |
IV (554) |
III (101) |
III (49) |
||
|
Holcus lanatus |
II (14) |
IV (189) |
IV (261) |
I (1) |
III (39) |
||
|
Arrhenatherum elatius |
I (1) |
II (171) |
III (290) |
I (9) |
I (3) |
||
|
Dactylis glomerata |
. |
III (224) |
I (12) |
I (8) |
I (1) |
||
|
Persicaria bistorta |
II (93) |
III (582) |
IV (362) |
III (186) |
II (65) |
||
|
Achillea millefolium |
I (1) |
IV (209) |
II (56) |
V (26) |
II (6) |
||
|
Geranium sylvaticum |
I (1) |
II (450) |
II (30) |
I (22) |
I (2) |
||
|
Veronica chamaedrys |
I (1) |
II (97) |
I (17) |
I (1) |
I (11) |
||
|
Angelica sylvestris |
. |
I (5) |
I (1) |
I (1) |
. |
||
|
Crepis paludosa |
. |
I (35) |
I (1) |
. |
. |
||
|
Avenula pubescens |
. |
I (11) |
I (47) |
. |
II (15) |
||
|
Linaria vulgaris |
. |
I (<1) |
I (1) |
I (1) |
. |
||
|
Lotus pedunculatus |
. |
I (26) |
I (28) |
. |
. |
||
|
Filipendula ulmaria |
. |
I (2) |
I (18) |
. |
. |
||
|
Achillea ptarmica |
. |
I (<1) |
I (3) |
I (1) |
. |
||
|
Carex panicea |
. |
I (1) |
I (8) |
I (16) |
II (26) |
||
|
Knautia arvensis |
. |
IV (245) |
I (36) |
II (10) |
I (3) |
||
|
Galium mollugo (s. l.) |
. |
I (30) |
I (3) |
. |
. |
||
|
Stellaria graminea |
. |
II (30) |
I (6) |
II (49) |
. |
||
|
Platanthera bifolia |
. |
. |
. |
II (10) |
I (1) |
||
|
3. Lisières |
|||||||
|
Agrostis capillaris |
IV (466) |
III (258) |
V (1791) |
V (837) |
V (1615) |
||
|
Deschampsia flexuosa |
III (483) |
V (1438) |
V (959) |
IV (517) |
V (628) |
||
|
Holcus mollis |
I (1) |
I (34) |
II (243) |
III (138) |
II (34) |
||
|
Briza media |
IV (159) |
V (371) |
IV (217) |
V (143) |
V (65) |
||
|
Anemone nemorosa |
III (17) |
III (88) |
II (83) |
I (28) |
V (368) |
||
|
Teucrium scorodonia |
I (1) |
I (2) |
I (21) |
. |
. |
||
|
Convallaria majalis |
I (2) |
I (7) |
I (10) |
. |
. |
||
|
Hieracium laevigatum |
I (2) |
I (<1) |
I (1) |
. |
I (1) |
||
|
Centaurea montana |
I (1) |
I (30) |
I (1) |
. |
. |
||
|
Anthoxanthum odoratum |
V (395) |
III (72) |
III (97) |
IV (84) |
III (88) |
||
|
Campanula rotundifolia |
V (40) |
III (153) |
III (25) |
V (81) |
IV (44) |
||
|
Vaccinium myrtillus |
II (4) |
I (9) |
I (9) |
I (6) |
. |
||
|
Stellaria holostea |
. |
I (<1) |
II (32) |
. |
I (1) |
||
|
Senecio ovatus |
. |
I (6) |
I (4) |
I (1) |
I (1) |
||
|
Solidago virgaurea |
. |
I (<1) |
I (1) |
I (3) |
. |
||
|
Rubus idaeus |
. |
I (2) |
I (71) |
I (3) |
. |
||
|
Galeopsis tetrahit |
. |
I(<1) |
II (15) |
I (4) |
. |
||
|
Senecio hercynicus |
. |
I (<1) |
I (14) |
. |
I (2) |
||
|
Epilobium angustifolium |
I (1) |
. |
I (1) |
II (12) |
. |
||
40Tableau 2. Evolution des prairies à Meum athamanticum. Représentation synthétique des relevés phytosociologiques (degrés de présence et coefficients de recouvrement)
41Parmi les espèces caractéristiques de la classe des Nardo-Callunetea, certaines restent aussi fréquentes dans le groupement qu’il y a 60 ans, leur taux de recouvrement augmentant (Meum athamanticum, Centaurea jacea subsp. nigra, Festuca rubra subsp. commutata, Lathyrus linifolius var. montanus, Galium saxatile, Helianthemum nummularium subsp. obscurum), restant plus ou moins constant (Thesium pyrenaicum, Carex pallescens, Pimpinella saxifraga, Carex caryophyllea) ou diminuant (Potentilla erecta, Cytisus scoparius). Par contre, 24 espèces typiques de cette classe ont fortement régressé (16 espèces), ou ont aujourd’hui disparu dans les nardaies abandonnées (8 espèces, dont 7 considérées comme rares à très rares dans la flore du sud du sillon Sambre-et-Meuse). Il s’agit surtout d’espèces liées à des pelouses rases ou des landes herbeuses à végétation basse, dont de petites graminées (Nardus stricta, Danthonia decumbens, Festuca filiformis), des espèces à rosettes basilaires (Arnica montana, Hieracium pilosella, Hypochaeris maculata), deux petites orchidées(Pseudorchis albida, Orchis mascula), une thérophyte (Linum catharticum), ainsi que des espèces reliques des landes (Calluna vulgaris, Vaccinium vitis-idaea, Genista anglica, G. pilosa, Botrychiumlunaria, Antennaria dioica, Carex pilulifera). Deux espèces des Nardo-Callunetea ont un recouvrement moyen nettement plus important aujourd’hui, une graminée : Festuca rubra subsp. commutata et une espèce des landes sèches : Galium saxatile.
42Une grande partie des espèces qui ont régressé ou disparu depuis l'abandon de l'utilisation traditionnelle des nardaies réapparaissent ou progressent suite aux actes de gestion. Beaucoup de ces espèces sont plus fréquentes ou plus abondantes dans les parcelles traitées de longue date par mise à feu (Nardus stricta, Festuca filiformis, Arnica montana, Hypericum maculatum, Calluna vulgaris, Ranunculus serpens subsp. polyanthemoides, Genista anglica), voire ne réapparaissent que là (Orchis mascula, Hypochaeris maculata, Pseudorchis albida). Narcissus pseudonarcissus, Genistella sagittalis et Polygala serpyllifolia, par contre, sont manifestement plus favorisés par la fauche. Hieracium umbellatum, espèce qui était très peu présente auparavant dans l’habitat, est très favorisé par les deux techniques de gestion; tandis que Galium saxatile régresse nettement suite aux mises à feu. Certaines espèces devenues rares dans le groupement ne progressent pas malgré la gestion (Hieracium pilosella, Thymus pulegioides, Viola canina).
43En ce qui concerne les espèces des Molinio-Arrhenatheretea, certaines régressent également suite à l’abandon des parcelles, notamment des plantes de petites tailles (Phyteuma nigrum, Pedicularis sylvatica, Rhinanthus minor, Dactylorhiza maculata) ou à rosettes basilaires (Succisa pratensis, Leontodon hispidus, Plantago lanceolata). Par contre, quelques espèces ont progressé fortement depuis les années 1940, surtout des graminées de grande taille croissant en nappes (Holcus lanatus, Arrhenatherum elatius, Dactylis glomerata) ou en touradons (Molinia caerulea, Deschampsia cespitosa, Poa chaixii). Certaines dicotylédones typiques des prés frais à humides ont aussi progressé (Persicaria bistorta, Achillea millefolium, Geranium sylvaticum, Veronica chamaedrys) ou sont apparues (Angelica sylvestris, Crepis paludosa, Lotus pedunculatus, Filipendula ulmaria, Achillea ptarmica) dans le groupement. Plusieurs des espèces en extension sont indicatrices d’un certain enrichissement du substrat en azote (Arrhenatherum elatius, Holcus lanatus, Dactylis glomerata, Poa chaixii, Geranium sylvaticum, Heracleum sphondylium var. sphondylium, Achillea millefolium).
44L'impact principal de la gestion sur les espèces des Molinio-Arrhenatheretea concerne les graminées sociales qui, à l'exception de Molinia caerulea, régressent très fortement (Poa chaixii, Deschampsia cespitosa, Holcus lanatus, Arrhenatherum elatius). Molinia caerulea est nettement plus fréquente aujourd'hui que dans les relevés anciens. Si cette espèce semble actuellement plus fréquente dans les zones gérées que dans les zones non gérées, son recouvrement moyen y est toutefois moins important. La gestion favorise aussi l'extension ou le retour de quelques petites plantes typiques du groupement (Succisa pratensis, Rhinanthus minor, Phyteuma nigrum, Dactylorhiza maculata). La mise à feu favorise fortement Achillea millefolium, Lotus corniculatus, Vicia cracca, Serratula tinctoria. Par contre, deux espèces communes (Ranunculus acris et Cirsium palustre) semblent régresser dans les zones incendiées. Une orchidée (Platanthera bifolia) non décelée à l’époque des pratiques agropastorales et absente des zones abandonnées apparaît dans le groupement suite à la gestion, surtout par mise à feu contrôlée.
45Trois graminées des lisières forestières sont en nette extension suite à l’absence d’entretien de l’habitat (Agrostis capillaris, Deschampsia flexuosa et Holcus mollis), tandis que trois autres espèces régressent (Anthoxanthum odoratum, Campanula rotundifolia, Vaccinium myrtillus). Quelques autres espèces des lisières font leur apparition dans les zones abandonnées mais avec un taux de recouvrement qui reste assez faible (Stellaria holostea, Senecio ovatus, S. hercynicus, Solidago virgaurea, Rubus idaeus, Galeopsis tetrahit). Campanula rotundifolia répond très favorablement quel que soit le mode de gestion. Par contre, Holcus mollis et Epilobium angustifolium sont favorisés par les mises à feu et Anemone nemorosa est fortement favorisé par la fauche. Deux graminées (Agrostis capillaris et Deschampsia flexuosa) restent très abondantes malgré la gestion.
464.3. Spectres écologiques
47Le spectre de teneur azotée du sol exprimée par les plantes (graphique 2a) indique que depuis les années 1940 les espèces indicatrices des sols très pauvres en azote ont régressé, au profit d’espèces des sols pauvres à moyennement riches en azote. On note également l’apparition, mais avec une faible abondance, d’espèces nitrophiles. La gestion, surtout par mise à feu contrôlée, favorise le retour des espèces typiques des sols pauvres en azote. Dans les zones incendiées, les espèces nitrophiles sont quasiment absentes.
48Le spectre d’acidité du sol (graphique 2b) montre la régression des espèces des sols très acides à acides au profit surtout d’espèces des sols basiques dans les parcelles abandonnées. La gestion permet de faire régresser ces espèces au profit d'espèces acidiphiles.
49Le spectre d’ouverture du milieu (graphique 2c) indique que les espèces de pleine lumière régressent au profit d’espèces des milieux fermés à moyennement fermés dans les parcelles abandonnées. La gestion, surtout par la fauche, favorise le retour des espèces héliophiles.
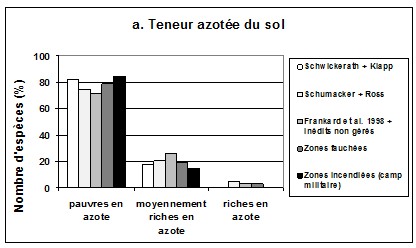
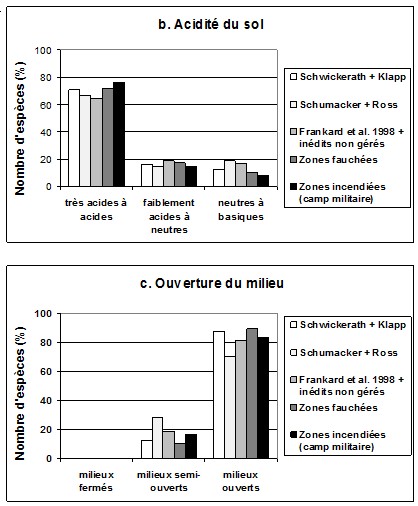
50Graphique 2. Spectres écologiques des espèces (d'après Ellenberg & al., 1992). a. Teneur azotée du sol; b. Acidité du sol; c. Ouverture du milieu
514.4. Analyses statistiques multivariées
524.4.1. Groupement des relevés
53La classification hiérarchique ascendante (graphique 3) montre une très forte opposition en deux groupes principaux et on propose une partition en 7 groupes.
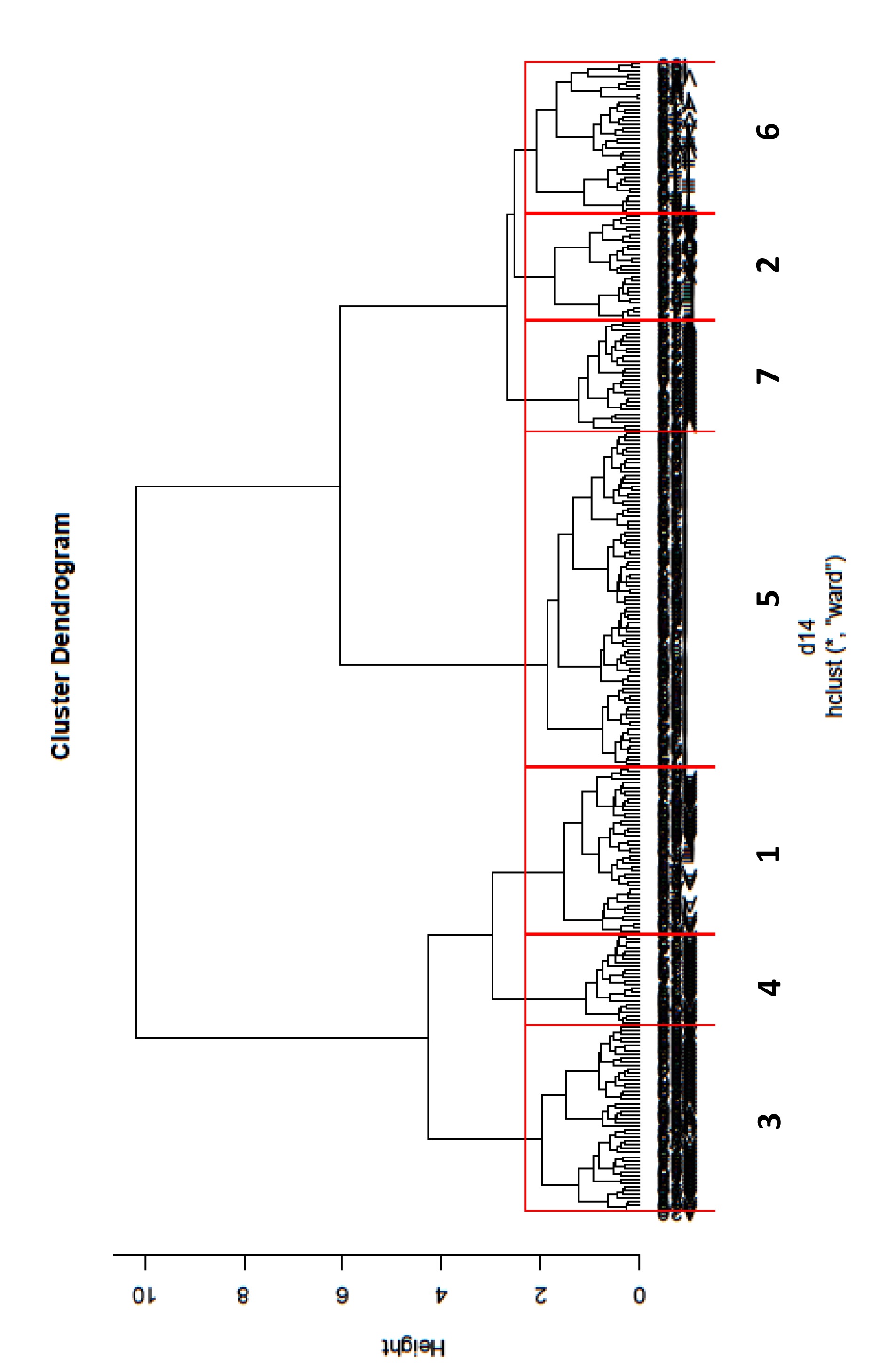
54Graphique 3. Classification hiérarchique ascendante des relevés (méthode Ward)
55Le premier groupe principal (filière 1) se divise en 3 groupes (groupes 1, 3, 4) comportant essentiellement des relevés de zones non gérées, ainsi que la moitié des relevés des zones gérées par fauchage (tableau 3).
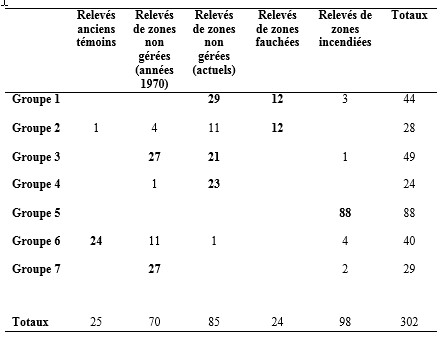
56Tableau 3. Répartition des relevés dans les 7 groupes définis par la classifi-cation hiérarchique ascendante
57En particulier, 86% des relevés récents de zones non gérées se retrouvent dans ces groupes (tableau 4). Un seul relevé issu d’une zone thermophile se retrouve dans cette filière.
|
Relevés anciens témoins |
Relevés zones non gérées (années 1970) |
Relevés zones non gérées (actuels) |
Relevés zones fauchées |
Relevés zones incendiées |
|
|
Filière 1 (groupes 1, 3, 4) |
0 % |
40% |
86% |
50% |
4% |
|
Filière 2 (groupes 2, 5, 6, 7) |
100% |
60% |
14% |
50% |
96% |
58Tableau 4. Répartition des relevés (en pourcentage) dans les deux filières principales générées par la classification hiérarchique ascendante
59 Le deuxième groupe principal (filière 2) se divise en 4 groupes (groupes 2, 5, 6, 7), comportant les relevés anciens témoins (groupe 6), la quasi-totalité des relevés des zones gérées par mise à feu (groupe 5), la moitié des relevés des zones gérées par fauchage (groupe 2) et des relevés de zones non gérées, datant surtout des années 1970 (groupes 6 et 7), parmi lesquels quasiment tous les relevés issus de zones thermophiles. On constate qu’une partie des relevés des années 1970 de zones abandonnées et n’ayant plus été gérées depuis une trentaine d’années sont encore très similaires aux relevés anciens témoins, puisqu’ils se retrouvent dans le même groupe. Seul un relevé réalisé récemment dans un secteur non géré est encore très similaire aux relevés anciens. Il est issu d’une zone thermophile. Les autres relevés récents de zones non gérées intégrés dans la filière 2 se retrouvent dans le groupe 2 avec les relevés des zones fauchées. 72% de ces relevés récents de zones non gérées correspondent à des relevés de zones thermophiles.
604.4.2. Espèces indicatrices du groupement (tableau 5)
61L'analyse des valeurs indicatrices avec la méthode Indval indique que 11 taxons sont indicateurs de la filière 1 (surtout des relevés de zones non gérées, en particulier des relevés récents, et des relevés de zones fauchées), tandis que 23 le sont de la filière 2 (surtout des relevés anciens témoins, des relevés de zones gérées et de zones thermophiles non gérées), leur valeur Indval étant largement significative (≥ 25 %; P ≤ 0.05).
62Les espèces indicatrices de la filière 1 (groupes 1, 3, 4) sont des graminées croissant en touradon : Deschampsia cespitosa (valeur Indval: 59,68 %), Poa chaixii (47,75 %) ; des graminées sociales des prés de fauche : Arrhenatherum elatius (36,83 %) ou des lisières forestières : Agrostis
capillaris (48,65 %), Deschampsia flexuosa (42,72 %) ; des espèces prairiales des prairies fraîches à humides : Persicaria bistorta (54,76 %), Holcus lanatus (52,15 %), Cirsium palustre (33,83 %) et Rumex acetosa (31,05 %) et une espèce des landes sèches particulièrement abondante dans les facies d’abandon : Galium saxatile (54,66 %).
|
Groupe |
Espèces indicatrices |
Indval ≥ 25% |
Pval ≤ 0,05 |
|
1 |
Galium saxatile |
0,25 |
0,001 |
|
2 |
Genistella sagittalis |
0,33 |
0,001 |
|
3 |
Holcus lanatus |
0,30 |
0,001 |
|
3 |
Cirsium palustre |
0,30 |
0,001 |
|
3 |
Crepis paludosa |
0,26 |
0,001 |
|
3 |
Angelica sylvestris |
0,25 |
0,001 |
|
4 |
Stellaria holostea |
0,61 |
0,001 |
|
4 |
Arrhenatherum elatius |
0,44 |
0,001 |
|
4 |
Poa chaixii |
0,40 |
0,001 |
|
4 |
Holcus mollis |
0,39 |
0,001 |
|
4 |
Galeopsis tetrahit |
0,30 |
0,001 |
|
4 |
Persicaria bistorta |
0,29 |
0,001 |
|
4 |
Deschampsia cespitosa |
0,28 |
0,001 |
|
4 |
Rubus idaeus |
0,26 |
0,001 |
|
5 |
Hieracium umbellatum |
0,57 |
0,001 |
|
5 |
Arnica montana |
0,26 |
0,001 |
|
7 |
Pimpinella saxifraga |
0,41 |
0,001 |
|
7 |
Thymus pulegioides |
0,35 |
0,001 |
|
7 |
Knautia arvensis |
0,31 |
0,001 |
|
7 |
Hieracium pilosella |
0,30 |
0,001 |
|
7 |
Viola riviniana |
0,29 |
0,001 |
|
7 |
Luzula multiflora |
0,28 |
0,001 |
|
7 |
Galium pumilum |
0,27 |
0,001 |
|
7 |
Achillea millefolium |
0,26 |
0,001 |
|
7 |
Dactylis glomerata |
0,26 |
0,001 |
|
7 |
Veronica officinalis |
0,25 |
0,001 |
|
7 |
Ajuga reptans |
0,25 |
0,001 |
63Tableau 5. Espèces indicatrices déterminées par la méthode Indval pour le niveau « 7 groupes »
64Au sein de cette filière, Galium saxatile (25 %) est indicateur pour le groupe 1 (relevés récents de zones non gérées ou relevés de zones fauchées). Le groupe 3 (relevés de zones non gérées des années 1970 ou actuelles) comporte des espèces prairiales de prairies fraîches à humides comme espèces indicatrices : Holcus lanatus (29,99 %), Cirsium palustre (29,97 %), Crepis paludosa (25,98 %), Angelica sylvestris (25 %) ; tandis que des graminées croissant en touradons ou en nappes : Arrhenatherum elatius (43,79 %), Poa chaixii (40,33 %), Holcus mollis (39,01 %), Deschampsia cespitosa (28,49 %) et des espèces des lisières forestières : Stellaria holostea (60,85 %), Galeopsis tetrahit (30,27 %), Rubus idaeus (26,20 %) sont indicatrices du groupe 4 (relevés récents de zones non gérées).
65Arnica montana (valeur Indval: 62,84), Nardus stricta (57,05 %), Succisa pratensis (57 %), Potentilla erecta (55,42 %), Centaurea jacea subsp. nigra (54,32 %), Festuca filiformis (53,89 %), Meum athamanticum (51,45 %), Danthonia decumbens (56,69 %), Calluna vulgaris (47,15 %), Briza media (46,23 %), Luzula multiflora (46,12 %), Hieracium umbellatum (45,22 %), Hypericum maculatum (44,51 %), Anthoxanthum odoratum (43,63 %), Campanula rotundifolia (41,77 %), Achillea millefolium (41,32 %), Carex pilulifera (38,42 %), Polygala serpyllifolia (38,15 %), Ranunculus serpens subsp. polyanthemoides (37,23 %), Rhinanthus minor subsp. minor (35,60 %), Phyteuma nigrum(32,78 %), … sont largement indicateurs de la filière 2 (groupes 2 et 5 à 7). Il s’agit essentiellement d’espèces caractéristiques ou compagnes de l’habitat.
66Au sein de cette filière, Genistella sagittalis (32,96 %) est indicateur pour le groupe 2 (zones non gérées actuelles, mais principalement thermophiles, ou zones fauchées) ; Hieracium umbellatum (56,62 %) et Arnica montana (25,87 %) sont indicateurs pour le groupe 5 exclusivement composé de relevés issus de zones gérées par mise à feu. Il n’y a pas d’espèces indi-catrices pour le groupe 6 (relevés anciens témoins) et diverses espèces, pour la plupart caractéristiques de l’habitat, sont indicatrices du groupe 7 (essentiellement des relevés de zones non gérées datant des années 1970), parmi lesquelles des thermophiles (beaucoup des relevés concernés relevant de cette variante du groupement) : Pimpinella saxifraga (41,16 %), Thymus pulegioides (35,12 %), Galium pumilum (27,66 %).
674.4.3 Ordination (ACP) (graphique 4)
68Les quatre premiers axes expliquent 44 % de la variance totale (respectivement 18; 11; 8; 7) (graphique 5). L'axe 1 paraît correspondre à un gradient croissant de naturalité ou de taux de conservation de l’habitat. Il oppose (côté -) des relevés des années 1970 de zones non gérées (groupes 3 et 4), des relevés récents de zones non gérées (groupes 1 et 3) et des relevés récents de zones fauchées (groupe 1) à des relevés ancestraux témoins (groupe 6), des relevés des années 1970 de zones non gérées (groupes 2 et 7) et des relevés de zones gérées par fauche (groupe 2) ou mise à feu (groupe 5) (côté+).
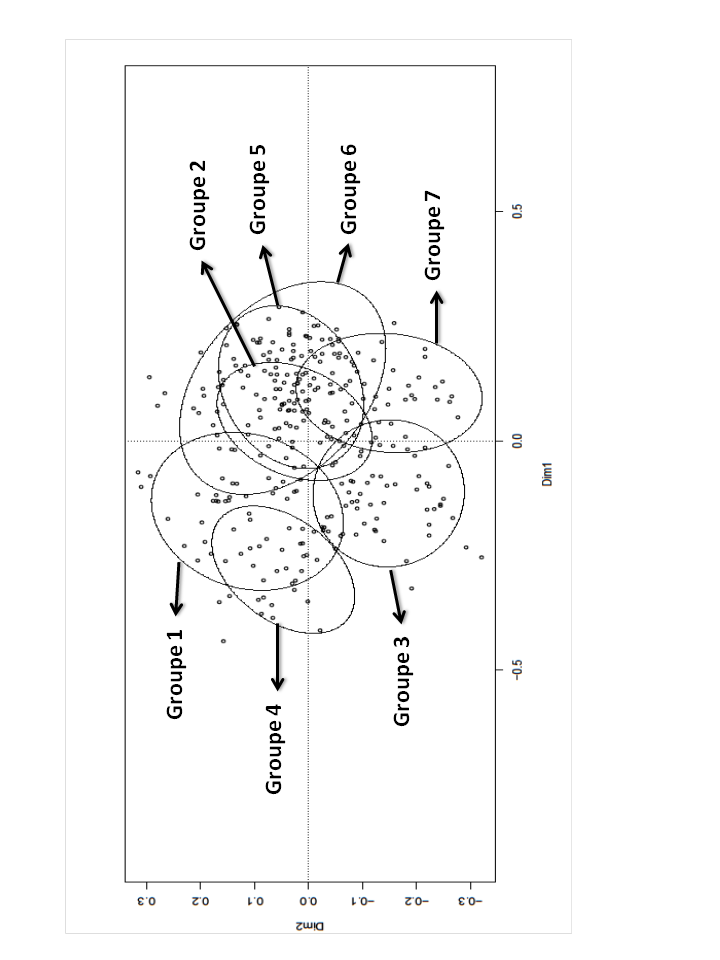
69Graphique 4. Ordination des relevés. Plan factoriel 1-2 de l’ACP
70On retrouve comme espèces majeures du côté négatif de l’axe 1 des graminées (Poa chaixii, Deschampsia cespitosa, Holcus lanatus, Arrhenatherum elatius), des dicotylédones de prairies fraîches à humides (Persicaria bistorta, Cardamine pratensis, Veronica chamaedrys, Cirsium palustre, Lotus pedunculatus, Angelica sylvestris),des espèces nitrophiles (Galium mollugo s. l., Geranium sylvaticum), des espèces des lisières forestières (Rubus idaeus, Galeopsis tetrahit, Stellaria holostea, Senecio hercynicus, S. ovatus). Ces espèces s'opposent (côté positif de l’axe 1) à des espèces typiques des landes et des nardaies. L’axe 2 oppose des relevés de zones non gérées (récents ou datant des années 1970) (côté -) à des relevés témoins, des relevés de zones gérées et des relevés récents de zones non gérées (côté +).

71Graphique 5. Analyse de la variance. L’axe de l’ordonnée donne le % de variance expliquée par chaque axe factoriel (1,2,…, de l’ACP).
725. DISCUSSION
73Les nardaies montagnardes sont des habitats très riches en espèces rares. L’abandon de ces milieux après la seconde guerre mondiale a conduit à une régression progressive manifeste de celles-ci (principalement des espèces caractéristiques des landes et des pelouses maigres) au profit d’espèces de haute taille, souvent hygrophiles, dont diverses graminées sociales, des espèces des lisières forestières et des espèces prairiales ubiquistes. Ce sont les espèces basses du groupement, moins compétitrices, qui ont le plus régressé, étouffées par des plantes hautes. Le fauchage et le pâturage extensif pratiqué jadis favorisaient les espèces de petite taille ou à feuilles en rosettes basilaires au dépend des plantes hautes, tandis que l’abandon des parcelles leur est défavorable.
74L’abandon des pratiques agropastorales a aussi entraîné l’accumulation progressive, chaque hiver, d’une couche de débris végétaux, ce qui a provoqué à la longue un certain enrichissement du milieu en azote. Diverses espèces en extension sont d’ailleurs indicatrices d’un enrichissement du substrat (Molinia caerulea, Arrhenatherum elatius, Dactylis glomerata, Poa chaixii, …). Cet enrichissement azoté accentue les effets négatifs liés à l’abandon en augmentant l’impact de la compétition interspécifique, en provoquant la régression des espèces les plus oligotrophes et en favorisant certaines espèces des Molinio-Arrhenatheretea de grande taille. Il est notamment établi que les espèces du Violion caninae ont une faible capacité à la compétition pour les nutriments et la lumière (Pegtel, 1994), ces deux causes combinées pouvant conduire à la dépression des populations des espèces caractéristiques de l’alliance.
75La tendance au reboisement spontané des nardaies abandonnées est par contre très faible (phénomène localisé surtout dans les formes les plus sèches de l’habitat). L’impact des dégâts de gel sur les jeunes plants forestiers et l’abondance des cervidés expliquent sans doute cette quasi absence de colonisation forestière.
76Les analyses statistiques confirment les résultats des tableaux phyto-sociologiques synthétiques. Elles montrent une forte opposition en 2 groupes principaux qui séparent d’une part les relevés anciens témoins, les relevés de zones thermophiles, 60% des relevés des années 1970 et une grande partie des relevés des zones gérées; et d’autre part les relevés récents de zones non gérées ou gérées très irrégulièrement par mise à feu ou seulement depuis quelques années par fauchage.
77La dégradation des nardaies a tout d’abord été assez lente, puisqu’une partie d’entre elles était toujours très similaire aux milieux témoins ou encore en relativement bon état de conservation 30-40 ans après leur abandon. Le phénomène s’est manifestement fortement accéléré par la suite, au point qu’au-jourd’hui, 70 ans après l’arrêt des pratiques agropastorales, la majorité des nardaies montagnardes abandonnées sont très dégradées.
78La variante thermophile de l’habitat évolue moins rapidement que les variantes types et hygrophiles. Dans celle-ci, les petites espèces liées aux pelouses rases ont moins régressé, ce qui explique que dans les analyses multivariées, la quasi totalité des relevés de zones non gérées issus de zones thermophiles, des années 1970 ou récents, soit toujours groupée avec les relevés ancestraux témoins ou présente dans la filière les englobant. Les analyses confirment donc que la dégradation des prairies thermophiles est moins prononcée que celle des prairies types et hygrophiles, ce que pressentait une étude antérieure (Frankard & al., 1998). On constate aussi que deux espèces caractéristiques de la variante thermophile (Helianthemum nummularium subsp. obscurum, Thymus pulegioides) ont un recouvrement nettement plus élevé dans les zones abandonnées que dans les zones témoins ou gérées. Ces 2 espèces sont des chaméphytes qui résistent relativement mal aux fauchages et aux mises à feu régulières et qui sont donc favorisés par l’abandon.
79Les changements floristiques observés dans les nardaies à Meum athamanticum : raréfaction ou disparition de diverses herbacées de petite taille typiques du groupement, en particulier des espèces des Nardo-Calluneteaet extension des graminées sociales ainsi que d’espèces indicatrices d’un enrichissement du sol en azote et d’espèces sciaphiles, témoignent de l’abandon déjà ancien des anciennes pratiques agropastorales (fauchage et pâturage extensif). La protection de ce type de milieux et sa préservation en bon état de conservation passent donc nécessairement par des actes de restauration/gestion adaptés, visant à remplacer la gestion traditionnelle. Deux techniques sont mises en œuvre sur le plateau d’Elsenborn : la mise à feu contrôlée, pratiquée de longue date dans les nardaies du camp militaire et le fauchage avec exportation du foin, entamé il y a une vingtaine d’années dans les réserves naturelles domaniales des vallées de la Schwalm et de l’Olef, mais qui s’est fortement intensifié depuis une dizaine d’années seulement.
80Les résultats montrent que les deux techniques de gestion sont efficaces et permettent le retour de diverses espèces caractéristiques du groupement. L’impact de la mise à feu semble plus important que celui de la fauche, mais cette technique de gestion est pratiquée depuis nettement plus longtemps. Les résultats obtenus par fauchage vont dans le bon sens, puisque 50 % des relevés des zones fauchées se retrouvent dans la filière comportant les relevés témoins. L’application de cette méthode de gestion est relativement récente dans certaines parcelles et les résultats seront probablement meilleurs d’ici quelques années.
81La composition floristique des zones gérées diffère toutefois quelque peu de celle des zones témoins et selon la méthode de gestion employée, comme en témoignent les analyses statistiques qui isolent les relevés de ces zones gérées dans des groupes distincts. Cela s’explique par la présence de quelques espèces différentielles très favorisées par l’un ou l’autre traitement : Arnica montana, Serratula tinctoria, Hypericum maculatum, Hieracium umbellatum, Genista anglica, Achillea millefolium, Platanthera bifolia, Lotus corniculatus, Vicia cracca… pour les mises à feu et Narcissus pseudonarcissus, Anemone nemorosa, Genistella sagittalis, Polygala serpyllifolia pour le fauchage.
82Plusieurs des espèces largement favorisées par les mises à feu sont réputées apprécier ce type de gestion (Hansen, 1964). Mallik & Gimingham (1983) ont notamment montré que les géophytes sont favorisées dans les premiers stades post-incendie. Arnica montana, en particulier,est une espèce qui réagit très positivement au feu (Hansen, 1964). Après incendie, cette astéracée se régénère en effet très vigoureusement et abondamment, tant par semis que par multiplication végétative En particulier, les épais rhizomesd’Arnica montana forment rapidement de nouvelles rosettes après le passage du feu, qui fleurissent en masse la seconde saison de végétation. Dans les zones incendiées, Genista anglica se régénère principalement par un rapide et vigoureux développement de jeunes pousses issues de la base des individus ayant survécu au passage du feu. Mais la chaleur atteinte dans les horizons superficiels du sol favorise aussi la germination des graines. L’élévation temporaire de la température du sol superficiel suite à un incendie est en effet un phénomène réputé favoriser la germination de diverses espèces de fabacées à graines dures (Baskin & Baskin, 1989 ; Probert, 1993). Cela explique probablement aussi l’abondance de Lotus corniculatus et Vicia cracca dans les zones régulièrement incendiées. De même, Serratula tinctoria est une composée dont le taux de germination est réputé être augmenté par l’apport de chaleur (Grime & al., 1981). Platanthera bifolia et Achillea millefolium sont capables de se régénérer rapidement et abondamment après le passage du feu, par tubercules souterrains pour la première (Hansen, 1964) et par rhizomes pour la seconde (Grime & al., 1988). Certaines espèces rares ne réapparaissent sporadiquement que dans les zones incendiées, notamment des orchidées à tubercules souterrains (Pseudorchis albida et Orchis mascula) et des astéracées à rosettes basilaires (Hypochaeris maculata, Leontodon hispidus).
83Par contre deux géophytes à bulbe (Narcissus pseudonarcissus) et à rhizome (Anemone nemorosa) ne semblent réagir très favorablement qu’à la fauche. Elles sont en effet très peu fréquentes et abondantes dans les zones incendiées, alors que l’on pourrait penser que leur mode de reproduction principalement végétatif à partir d’organes souterrains les y aurait favorisés. Il s’agit cependant d’espèces affectionnant les sols frais et les milieux très ouverts générés par la mise à feu sont peut-être trop secs pour ces deux espèces mésophiles. Quant à Genistella sagittalis et Polygala serpyllifolia il s’agit de deux espèces rampantes, émettant des rejets à partir d’une souche plus ou moins ligneuse, ce qui explique que le régime de la fauche leur soit plus favorable que la mise à feu qui détruit les organes aériens.
84La progression importante de Genistella sagittalis dans les zones fauchées explique pourquoi la moitié des relevés réalisés dans les zones fauchées (ceux qui marquent le mieux l’impact positif de la gestion) se regroupent avec des relevés de zones non gérées, mais thermophiles, cette espèce étant aussi indicatrice des zones les plus sèches.
85Les deux techniques de gestion permettent la régression des nitrophiles par l’exportation de nutriments. L’impact est plus marqué dans les zones incendiées, mais il est bien connu que cette technique de gestion fait diminuer considérablement la teneur en azote dans les horizons de surface et moyen (Allen, 1964 ; Evans & Allen, 1971 ; Rozé & Forgeard, 1982 ; Dolman & Land, 1995) et elle est appliquée depuis plus longtemps que la fauche.
866. CONCLUSION
87L’abandon déjà ancien des pratiques agropastorales est responsable d’un appauvrissement important de la richesse floristique des nardaies à Meum athamanticum. Les principaux impacts observés sont la régression de diverses espèces typiques du groupement au profit notamment de graminées sociales de grande taille, la fermeture du milieu et son léger enrichissement en azote. La sauvegarde et la restauration de ces pelouses maigres passent nécessairement par des actes de gestion adaptés. La mise à feu contrôlée et la fauche régulière avec enlèvement du foin semblent être de bonnes méthodes, puisqu’elles contrecarrent l’appauvrissement de la richesse de ces anciens prés de fauche et rétablissent les espèces qui avaient le plus tendance à régresser voire disparaître. Le choix de l’une ou l’autre technique de gestion dépend des opportunités locales. La mise à feu contrôlée implique nécessairement d’avoir des surfaces importantes de l’habitat à traiter, plutôt que de petites parcelles isolées et doit donc être réservée aux grands sites protégés. Là où c’est possible, un mélange des deux techniques est probablement une bonne option pour que s’exprime au mieux toute la diversité végétale de l’habitat.
88REMERCIEMENTS
89Je tiens à remercier tout particulièrement le Professeur honoraire Jacques Lambinon (ULg) pour sa relecture critique du manuscrit et ses précieux conseils. Un tout grand merci également au Professeur Marc Dufrêne (ULg) qui m’a initié au logiciel R et sans qui je n’aurais pu réaliser les analyses statistiques, ainsi qu’à René Dahmen (Chef du cantonnement forestier d’Elsenborn) qui a traduit le résumé en allemand.
90BIBLIOGRAPHIe
91Allen S. E., 1964. − Chemical aspects of heather burning. J. Appl. Ecol. 1: 347-367.
92Baskin J. M. & Baskin C. C., 1989. − Physiology of dormancy and germination in relation to seed bank ecology. In « Ecology of soil seed banks », Leck M. A., Parker V. T. & Simpson R. L., eds. Academic Press, San Diego: 53-66.
93Baur K., 1967. − Ökologische und soziologische Beobachtungen bei Meum athamanticum Jacq. Jh. Ver. Vaterl. Naturkde Württemberg 122 : 122-125.
94BodarwÉ F, 1991. − La vallée de la Holzwarche (Büllingen, Belgique) : études floristique, phytosociologique et écologique. Propositions d’un plan de gestion écologique. Univ. Liège. Mém. Lic. Sci. Bot., 71 p. + ann.
95Deflandre A., 1977. − Climatologie et végétation dans la vallée de la Schwalm moyenne.Univ. Liège, Mém. Lic. Sci. Géogr., 211 p. + ann.
96Dolman P. M. & Land R., 1995. − Lowland heathland. In « Managing habitats for conservation », Sutherland W. & Hill D. A., eds. Cambridge University Press, Cambridge.
97Dufrêne M. & Legendre P., 1997. − Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67: 345-366.
98Ellenberg H., Weber H. E., Düll R., Wirth V., Werner W. & Paulien D., 1992. − Zeigerwerte von Pflanzen in Mitteleuropas. Scripta Geobot. 18 : 258 p.
99Evans C. C. & Allen S. E., 1971. − Nutrient losses in smoke produced during heather burning. Oikos 22: 149-154.
100Fontaine S., (coll. Schumacker R., Ros G. & Fabri R.), 1981. − L’abîssage des prés dans les vallons de haute Ardenne nord-orientale. Hautes Fagnes 47 : 117-140 + 14 fig. h. t.
101Frankard Ph., Ghiette P. & Schumacker R., 1998. − Les réserves naturelles domaniales des vallées de la Schwalm et de l’Olef.Région Wallonne, Dir. Gén. des Ressources natur. et de l’Environnement,Conserv. de la nature, Travaux 19 : 80 p.
102Grime J. P., Mason G., Curtis A. V., Rodman J., Band S. R., Mowforth M. A. G., Neal A. M. & Shaw S., 1981. − A comparative study of germination characteristics in a local flora. J. Ecol. 69 : 1017-1059.
103Grime J. P., Hodgson J. G. & Hunt R., 1988. − Comparative plant ecology. A functional approach to common British species. Unwin Hyman Ltd, London, 742 p.
104Hansen K., 1964. − Studies on the regeneration of heath vegetation after burning-off. Bot. Tidsskrift 60: 1-41.
105Klapp E., 1951. − Borstgrasheiden des Mittelgebirge. Entstehung, Standort, Wert und Verbesserung. Zeitschr. F. Acker-u. Pflanzenbau 93 : 400-444.
106Lambinon J., Verloove F. (& coll.), 2012. − Nouvelle Flore de la Belgique, du Grand-Duché de Luxembourg, du Nord de la France et des régions voisines (Ptéridophytes et Spermatophytes). 6ème édition, Meise. Ed. Patrimoine du Jardin botanique national de Belgique, CXXXIX + 1195 p.
107Mallik A. & Gimingham C. H., 1983. − Regeneration of heathland plants following burning. Vegetatio 53: 45-58.
108Matzke G. 1985. − Zur Verbreitung, Sociologie und Ökologie des gelben Narzisse (Narcissus pseudonarcissus) in Eifel und Hunsrück. Bonn, Rheinisch Fried.-Wilhelm Univ., 82 p.
109Oberdorfer E., 1993. − Süddeutsche Pflanzengesellschaften. Teil 2: Sand- und Trockenrasen, Heide- und Borstgrasgesellschaften, alpine Magerrasen, Saum-Gesellschaften, Schlag- und Hochstauden-Fluren. Jena. Gustav Fischer Verlag, 355 p
110Pegtel D. M., 1994. − Habitat characteristics and the effect of various nutrient solutions on growth and mineral nutrition of Arnica montana L. grown on natural soil. Vegetatio 114 : 109-121.
111Probert R. J., 1993. − The role of temperature in germination ecophysiology. In « Seeds. The ecology of regeneration in plant communities », Fenner M., ed. Cab International, Wallingford: 285-325.
112Reichert H., 1972. − Verbreitung und Soziologie des Bärwurz, Meum athamanticum Jacq., im Hunsrück. Decheniana 125 (1-2): 15-22 + 1 ill. h. t.
113Ros G., 1980. − Etude floristique, phytosociologique et écologique des vallées de l’Olef et de la Holzwarche. Univ. Liège, Mém. Lic. Sci. Géogr., 138 p. + ann.
114Rozé F. & Forgeard F., 1982. − Evolution de la minéralisation de l’azote dans des sols de landes incendiées et non incendiées de la région de Paimpont (Bretagne, France). Acta Oecol., Oecol. Plant. 3 (3) : 249-268.
115Schumacker R., 1975. − Les landes, pelouses et prairies semi-naturelles des plateaux des Hautes-Fagnes et d’Elsenborn (Prov. de Liège, Belgique). 1. Aspects floristiques, phytosociologiques et phytogéographiques. Coll. Phytosoc. 2 : 13-36 + 2 tabl. h. t.
116Schumacker R. & Duvigneaud J., 1974. − Une espèce nouvelle pour la flore belge : Vicia orobus DC. dans la vallée de la Schwalm à Elsenborn (prov. de Liège, Belgique). Bull. Soc. Roy. Bot. Belg. 107 : 41-52.
117Schwickerath M., 1944. − Das Hohe Venn und seine Randgebiete. Pflanzensoziol. 6 : 278 p.
118Sougnez N., 1977. − Les associations de la nardaie en Belgique (Nardetalia Prsg. 49). Comm. Centre Ecol. forest. et rur. N.S. 15 : 1-90 + ann.
119Stieperaere H., 1990. − De heischrale grasslanden (Nardetea) van atlantisch Europa. Univ. Gent, Th. Doct. Sci. Bot., 303 p.
120Stieperaere H. & Fransen K., 1982. − Standaardlijst van de belgische vaatplanten,
met aanduiding van hun zeldzaamheid en socio-oecologische groep. Dumortiera 22: 1-42.
121Vanden Berghen C., 1982. − Initiation à l’étude de la végétation. Meise, Jardin Botanique National de Belgique, 263 p.
122van der Maarel E., 1979. − Transformation of cover-abundance values in phyto-sociology and its effects on community similarity. Vegetatio 39 : 97-114.
123Van Rompaey E. & Delvosalle L., 1979. − Atlas de la flore belge et luxembourgeoise. Ptéridophytes et spermatophytes. 2ème éd., Meise. Jard. Bot. Nat. Belgique, 293 p., 1542 cartes.
Notes
1 Direction de la Nature et de l’Eau ; Département de l’Etude du Milieu Naturel et Agricole (SPW), Avenue Maréchal Juin, 23, B-5030 Gembloux. E-mail : Philippe.Frankard@spw.wallonie.be
