


- Accueil
- N° 203 (décembre 2020)
- Diversité floristique de la Steppe sud algéroise : cas de la région de Djelfa (Algérie)


Visualisation(s): 8889 (91 ULiège)
Téléchargement(s): 380 (24 ULiège)

Diversité floristique de la Steppe sud algéroise : cas de la région de Djelfa (Algérie)

Résumé
La zone sur laquelle porte notre contribution est localisée dans la steppe sud Algéroise (Algérie). Cette recherche est consacrée à l’analyse de la diversité floristique dans la région de Djelfa.
L'étude de la végétaion de notre zone d'étude, a permis de discriminer sept groupements végétaux distincts sur le plan floristique et physionomique.
L’analyse de la phytodiversité globale, a permis de recenser 170 taxons appartenant à 34 familles et 111 genres. Asteraceae, Poaceae, Fabaceae, Brassicaceae et Cistaceae sont les familles les plus représentées avec environ 59% des espèces. Le spectre biogéographique global indique la dominance des espèces méditerranéennes avec 80 espèces soit 47 % des taxons dénombrés. 27 espèces endémiques soit 16 % ont été identifiées, dont 16 espèces le sont d’Afrique du Nord. L'analyse des types biologiques a montré la dominance des thérophytes qui représentent 85 taxa soit 50 % par rapport aux autres formes de vie. L’analyse de la diversité floristique par l’utilisation de l’indice de Shannon-Weaver (H’) dans les unités de végétation (forêt, matorral, pelouse, steppe à alfa, steppe présaharienne), a montré une diversité moyenne à faible qui oscille entre 3,34, dans les formations végétales de la steppe à alfa, et 2,40 dans la steppe présaharienne dégradée. Ces résultats sont confirmés par l’indice de Pielou (E) avec respectivement 0,55 et 0,39 dans les deux formations.
Abstract
The area covered by our contribution is located on the southern Algerian steppe (Algeria). This research is dedicated to the analysis of the floristic diversity of the flora in the Djelfa region.
The study of the vegetation in our study area made it possible to discriminate seven distinct plant groupings on the floristic and physiognomic level.
The analysis of the global plant diversity made it possible to identify 170 taxa, belonging to 34 families and 111 genera. Asteraceae, Poaceae, Fabaceae, Brassicaceae and Cistaceae are the most represented families with about 59 % of species. The biogeographic spectrum indicates the dominance of Mediterranean species with 80 species or 47 % of the recorded taxa. 27 endemic species or 16 % have been identified, 16 of which are endemic to North Africa. The analysis of biological types showed the dominance of therophytes which represent 85 taxa or 50 % compared to other life forms. The analysis of floristic diversity with the Shannon-Weaver (H’) index in the vegetation units (forest, matorral, grassland, alfa steppe, pre-Saharan steppe), showed a medium to low diversity that oscillates between 3.34 in alfa steppe, and 2.40 in degraded pre-Saharan steppe. These results are confirmed by the Pielou index (E) with 0.55 and 0.39 respectively in both formations.
Table des matières
1. INTRODUCTION
1La steppe algérienne, qui occupait plus de 20 millions d'hectares, constitue les parcours de pâturage les plus répandus des pays Nord africains (DJEBAILI, 1984, HIRCHE et al., 2011). La formation végétale steppique en Algérie, malgré le rôle écologique et économique qu’elle assure sur une grande région géographique, est confrontée depuis plusieurs décennies à un processus de dégradation devenant de plus en plus irréversible (AIDOUD et al., 2006, NEDJRAOUI & BEDRANI, 2008, TAIBAOUI, 2008, HIRCHE et al., 2010, MOULAY et al., 2011).
2La région de Djelfa, à l’instar des autres wilayas steppiques de l’Algérie est marquée par le phénomène de désertification. L’étude sur la sensibilité à la désertification menée par l’agence spatiale algérienne ASAL en 2010, a mis en exergue le danger qui menace cette région ; environ 39.213,83 hectares de terres sont désertifiés et 48.148,50 hectares sont très sensibles à la désertification (CHAOUCHE KHOUANE et al., 2018, GHAFOUL et al., 2019).
3En outre, la gestion rationnelle des ressources de ces écosystèmes ne saurait être efficace sans une bonne connaissance de la structure et de la composition floristique de ces écosystèmes (ARONSON et al., 1993, CHAUVIN & VALLAURI, 2002).
4Par ailleurs, plusieurs recherches ont été effectuées sur l’écosystème steppique de la région de Djelfa telles que les travaux phytoécologiques et cartographiques de POUGET (1977, 1980), DJEBAILI (1978, 1984), MELZI (1986), MELZI et al. (1993), URBT (1987), NEDJRAOUI & BEDRANI (2008). Cependant peu d’études ont été consacrées à l’inventaire et l’actualisation de la diversité floristique de cette zone qui offre un patrimoine préforestier de 152.753 hectares (DGF, 2012).
5C’est dans cette optique que s’inscrit notre recherche qui porte comme objectif l’étude de la diversité floristique des formations végétales de la steppe Sud Algéroise de la région de Djelfa.
2. MATÉRIELS ET MÉTHODES
2.1 Zone d’étude
6La province de Djelfa est située dans la partie centrale de l'Algérie, au-delà de la limite sud de l’Atlas Tellien (HABIB et al., 2020). Notre zone d’étude qui couvre une superficie de 179.903 hectares, se situe sur le versant sud des Monts des Ouled Nail, qui constitue la partie centrale des massifs montagneux de l’Atlas Saharien. Elle touche plusieurs communes et s’inscrit entre les coordonnées suivantes (Fig.1) :
-
Latitude Nord : 34°05’ et 34° 30’
-
Longitude : Est : 3°10’ et 3°30’
La zone d’étude appartient à l’étage bioclimatique semi-aride à hivers froids avec des étés chauds et secs. Elle est caractérisée par une pluviométrie de l’ordre de 350 mm au Nord à moins de 243 mm au Sud. La zone enregistre 10 à 13 jours d’enneigement en moyenne et 20 à 30 jours de vents dominants d’origine désertique durant l’année (POUGET, 1980).
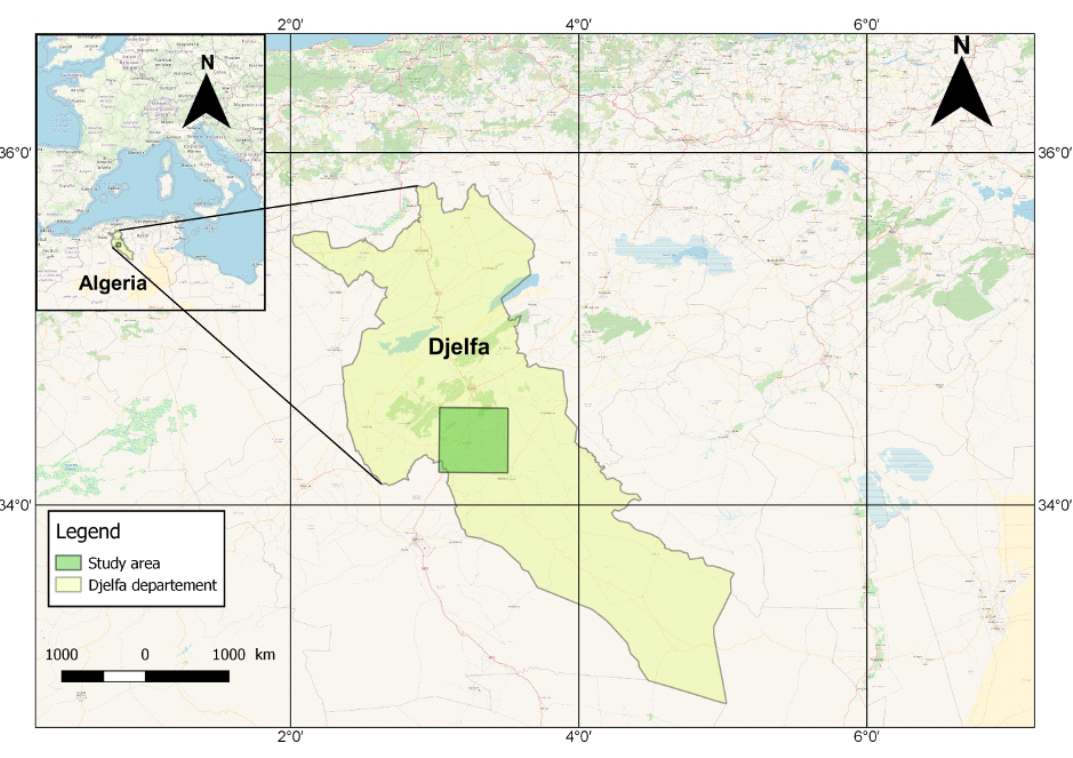
FIG. 1 - Situation géographique de la zone d’étude.
2.2 Méthodes
7Plusieurs habitats écologiques ont été étudiés: forêt claire, matorral, steppe à alfa et steppe présaharienne. L’étude de la végétation a été faite en tenant compte des travaux antérieurs, tels que LE HOUÉROU et al. (1977), POUGET (1977) et MELZI (1986).
850 relevés floristiques ont été effectués durant la période optimale de végétation suivant un échantillonnage mixte (starifié, subjectif) qui tient compte de la variabilité des formations végétales rencontrées. Selon la méthode sigmatiste, les relevés ont été réalisés sur des aires homogènes d’un point de vue écologique (GOUNOT, 1961, 1969, DAGET & POISSONET, 1971, LONG, 1974, LE HOUÉROU, 1995, OSS, 2008, RICHARD et al., 2010).
9Nous avons adopté, d’une part une aire minimale de 100 m2 préconisée par l’URBT (1978), DJEBAILI (1978, 1984) et BOUAZZA & BENABADJI (2002) pour les formations steppiques algériennes, et d’autre part une aire minimale de 400 m2 recommandée pour les groupements forestiers (GUINOCHET, 1955, 1973, GÉHU & RIVAS-MARTINEZ, 1981, OZENDA, 1982, DJEBAILI, 1984).
10L'identification et la nomenclature des espèces se base sur les ouvrages suivants : la nouvelle flore d'Algérie et des régions désertiques méridionales (QUÉZEL & SANTA, 1962), la flore de l'Afrique du Nord de René MAIRE (1952 - 1987), Flore et végétation du Sahara (OZENDA, 2004), l'index synonyme et bibliographique de la flore Nord-africaine (DOBIGNARD & CHÂTELAIN, 2010 - 2013).
11Les types biogéographiques des espèces ont été déterminés sur base des travaux de QUÉZEL & SANTA (1962), PIGNATTI (1982) et DOBIGNARD & CHÂTELAIN (2010 - 2013). Nous avons utilisé la classification de RAUNKIAER (1934) qui se base sur la position qu’occupent les méristèmes en dormance par rapport au niveau du sol durant la mauvaise saison et se subdivise ainsi en phanérophytes, chaméphytes, hémi-cryptophytes, thérophytes et géophytes.
12Afin d’évaluer la diversité spécifique de notre zone, nous avons utilisé l’indice de Shannon-Weaver pour mesurer la structure intra-biotope de chaque unité de végétation (RAMADE, 2003) et l’indice d’équitabilité ou indice de régularité de PIÉLOU qui permet de mesurer la répartition des individus au sein des espèces, indépendamment de la richesse spécifique. L’indice varie entre 0 (dominance d’une des espèces) et 1 (équirépartition des individus dans les espèces) (FRONTIER et al., 2008).
13Dans le but d’estimer la rareté des taxons, nous avons utilisé l’échelle proposée par MEDJAHDI et al. (2018). RR : l’espèce est considérée comme très rare si elle n’est présente que dans 1 à 2 relevés ; R : rare si elle est retrouvée de 3 à 5 fois ; C : commune si elle se rencontre de 6 à 10 fois ; AC : assez commune si elle se rencontre de 11 à 20 fois ; CC : très commune si on la rencontre plus de 20 fois.
2.3 Analyse des données
14L’analyse du tableau de relevés a été faite au moyen de l'analyse non-symétrique des correspondances NSCA (GIMARET-CARPENTIER et al., 1998). Cette technique part de l'observation de la manière dont les données sont récoltées sur le terrain. Les présences des espèces sont observées parmi des relevés. Les relevés (nombre, position, par exemple) sont choisis par l'observateur, pas les espèces. Il en résulte une dissymétrie fondamentale qui est explorée par cette technique. La différence entre l’analyse des correspondances classique (CA) et l’analyse non-symétrique concerne précisément la métrique. Les espèces ont un poids uniforme égal à l'unité dans NSCA alors que CA est basée sur la métrique qui implique un double "averaging" calculé à partir des totaux marginaux des lignes et colonnes des tableaux. Cette dernière technique est ainsi qualifiée de duale. Dans NSCA, une forte importance est accordée aux espèces abondantes et l’effet des espèces rares est fortement limité. Dans un tableau d’occurrences, un grand nombre de taxons rares ne sera donc pas un facteur limitant. Un poids très faible ramène automatiquement un taxon vers l’origine.
15Avant analyse, le tableau originel a été transformé en un tableau disjonctif, avec trois lignes potentielles par espèce, une pour les présences, une pour les abondances supérieures à 1 et une troisième pour les abondances supérieures à 3. Cette technique, utilisée à de nombreuses reprises, évite les écueils venant de l’analyse de données d’abondance brutes tout en les intégrant d’une autre manière (BOUXIN, 2016, 2020). Les analyses ont été faites avec des programmes en R. Un test par permutation permet d’identifier les valeurs propres et contributions relatives significatives des espèces et relevés, avec un risque de première espèce α = 0,05.
16La classification ascendante hiérarchique (CAH), est préconisée par ROUX & ROUX (1967) comme complément à des analyses multivariées (LERMAN, 1977, MEDDOUR, 2011). Elle est faite suivant la méthode de WARD jugée la plus pertinente par plusieurs chercheurs (VACHON et al., 2005, LABATTE, 2010).
3. Résultats
3.1 Phytodiversité des unités de végétation
17170 taxons appartenant à 34 familles et 111 genres ont été identifiés, ce qui représente 3,95 % de la flore totale algérienne estimée à 4300 espèces (DOBIGNARD & CHÂTELAIN, 2010 – 2013, BENAISSA et al., 2018).
18Sur l’ensemble des familles recensées (Fig. 2), cinq familles dominent la flore de la région d’étude, ce sont : Asteraceae avec 41 espèces (23,56 %), Poaceae avec 20 espèces (11,49 %), Fabaceae avec 17 espèces (9,77 %), Brassicaceae avec 13 espèces (7,47 %) et Cistaceae avec 9 espèces (5,17 %). Ces familles représentent à elles seules 59 % des espèces inventoriées.
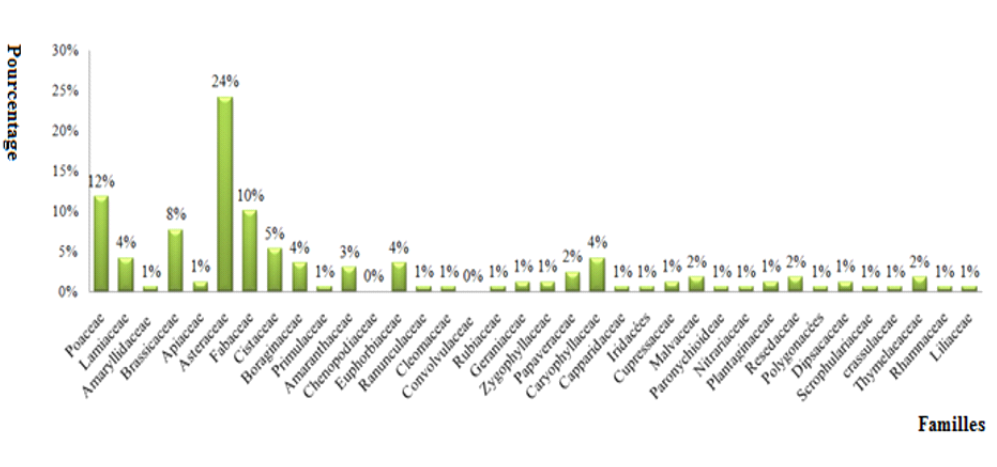 FIG. 2 - Richesse globale des familles botaniques recensées.
FIG. 2 - Richesse globale des familles botaniques recensées.
19D'autres familles comme les Lamiaceae, Caryophyllaceae, Euphorbiaceae, Boraginaceae, Amaranthaceae et Papaveraceae sont moyennement représentées avec respectivement 7, 6, 5 et 4 taxons. Les autres familles sont peu représentées et offrent seulement 1 à 3 espèces.
20Nous constatons ainsi la dominance des Asteraceae, Poaceae et Fabaceae. Ce constat est souligné par plusieurs chercheurs, notamment BENABADJI et al. (2007).
3.2 Types biologiques
21La répartition des types biologiques de l’ensemble de la flore de notre région d’étude est caractérisée par le schéma qui suit : Th > He > Ch > Ph> Géo (Fig. 3). Nous enregistrons une dominance des thérophytes qui constituent la moitié de l’ensemble floristique, avec un total de 85 espèces, soit 50 %. En deuxième rang, viennent les hémicryptophytes qui sont représentées par 44 taxons soit 26 %. Les chaméphytes occupent la troisième position avec 29 espèces, ce qui représente 17 %. Ces trois types biologiques (thérophytes, hémicryptophytes, chaméphytes) sont largement majoritaires par rapport aux autres types biologiques qui ne représentent que 7 %.
22Plusieurs auteurs (GRIME, 1977, DAGET, 1980, BARBERO et al., 1990, AIDOUD, 1984 et BARBERO et al., 2001) ont mis en exergue la relation qui existe entre le taux élevé de thérophytes et le gradient croissant d'aridité dans les zones arides méditerranéennes. Nous assistons à une thérophytisation à cause de l’aridité du climat d’une part et l’anthropisation d’autre part (SAUVAGE, 1961, GAUSSEN, 1963, NÈGRE, 1966, DAHMANI, 1997 et KADI-HANIFI, 1998).
23Cette thérophytisation est due aussi au phénomène de self-mulching, mécanisme se produisant dans les zones à forte évaporation et dans les sols limoneux en profondeur et couverts de sable en surface. Ce phénomène permet l’emmagasinement de l’eau et favorise ainsi le développement des annuelles (HARRANE-ZAOUI, 2012).
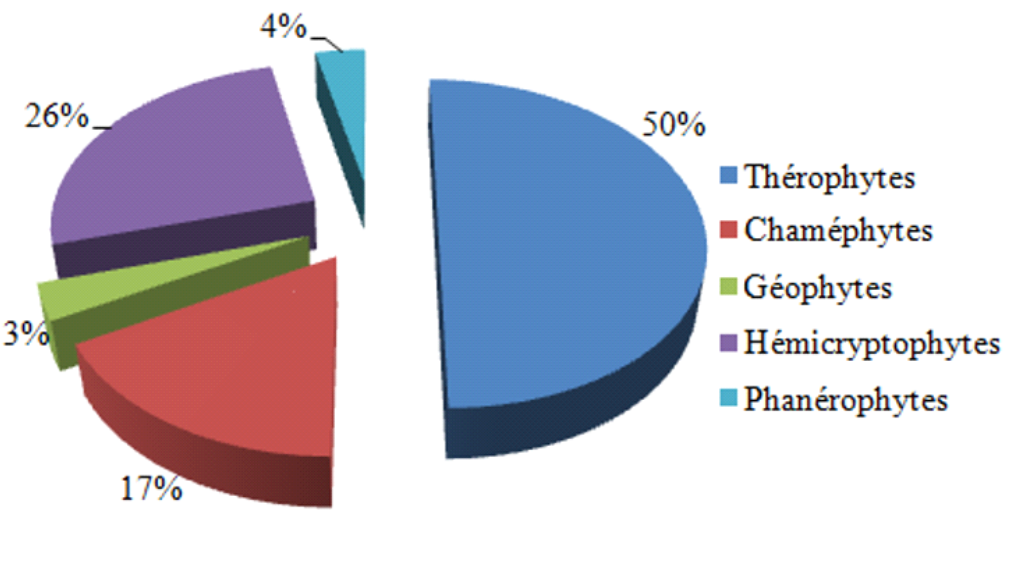
FIG. 3 - Spectre biologique global.
3.3 Types géographiques
24L’examen du spectre biogéographique des espèces recensées (Fig. 4) montre une dominance des espèces méditerranéennes avec 80 espèces, ce qui représente 47 % de l’ensemble de la flore étudiée, parmi ces espèces, citons : Pinus halepensis, Cistus libanotis, Juniperus oxycedrus, Juniperus phoenicea, Jurinea humilis, Helianthemum virgatum, Thymelaea tartonraira, Artemisia herba-alba, Artemisia campestris, Plantago albicans, Noaea mucronata, Peganum harmala, Ziziphus lotus…
25Le type géographique saharien est représenté par 33 espèces (25,56 %), dont 12 taxons sont ibéro-mauritanien. Pour ce type nous citons à titre d’exemple : Arthrophytum scoparium, Aristida pungens, Argyrolobium uniflorum, Atractylis serratuloides, Globularia alypum, Asteriscus pygmaeus, Cleome arabica, Salsola vermiculata, Herniaria fontanesii, Launaea acanthoclada, Stipa tenacissima…
26On dénombre 27 espèces endémiques (16 %), dont la majorité est nord-africaine (16 espèces) telles que : Ammoides atlantica, Anacyclus cyrtolepidioides, Arnebia decumbens, Astragalus armatus, Atractylis phaeolepis, Atractylis polycephala, Ceratocephalus falcatus, Echium trygorrhizum, Fagonia microphylla, Genista microcephala, Polycnemum fontanesii, Rosmarinus tournefortii, Thymelaea virgata et Thymus ciliatus.
27Enfin, l’ensemble géographique septentrional compte 11 taxons (5 %). Les espèces eurasiatiques sont les plus nombreuses avec 7 espèces telles que : Alyssum granatense, Androsace maxima, Genista cinerea, Micropus bombycinus, Poa bulbosa, Salvia verbenaca.
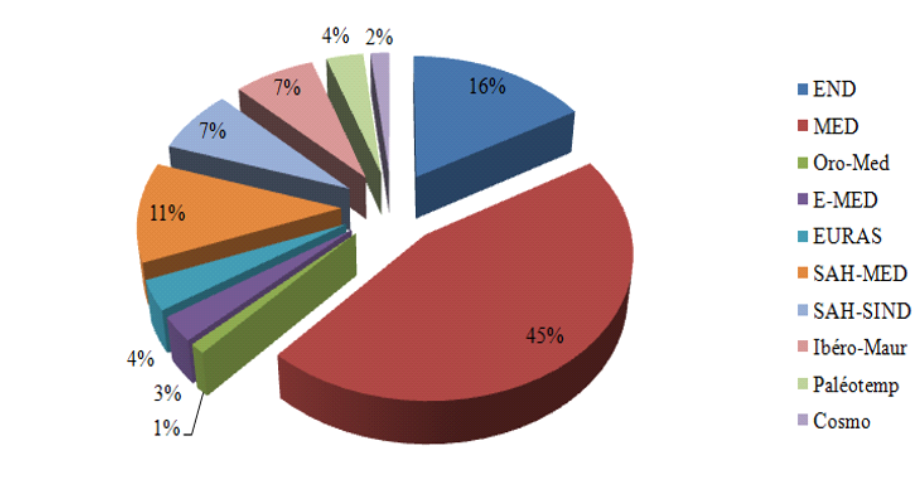
FIG. 4 - Spectre biogéographique des espèces inventoriées.
3.4 Analyse de la rareté des espèces
28Concernant le critère de rareté des espèces dénombrées dans notre zone d’étude, nous distinguons (Fig. 5) :
29- 34 espèces rares, soit 33 % de l’ensemble telles que : Alyssum scutigerum, Anacyclus cyrtolepidioides, Moricandia arvensis ;
30- 56 espèces communes (28 %) telles que : Allium cupani, Arnebia decumbens, Artemisia herba -alba ;
31- 48 espèces très rares (20 %) telles que : Alyssum alpestre, Ammoides atlantica, Euphorbia bupleuroides ;
32- 29 espèces assez communes (17 %) telles que: Arthrophytum scoparium, Astragalus armatus, Pinus halepensis, Atractylis serratuloides ;
33- 3 espèces très communes (2 %): Juniperus phoenicea, Stipa parviflora, Stipa tenacissima.
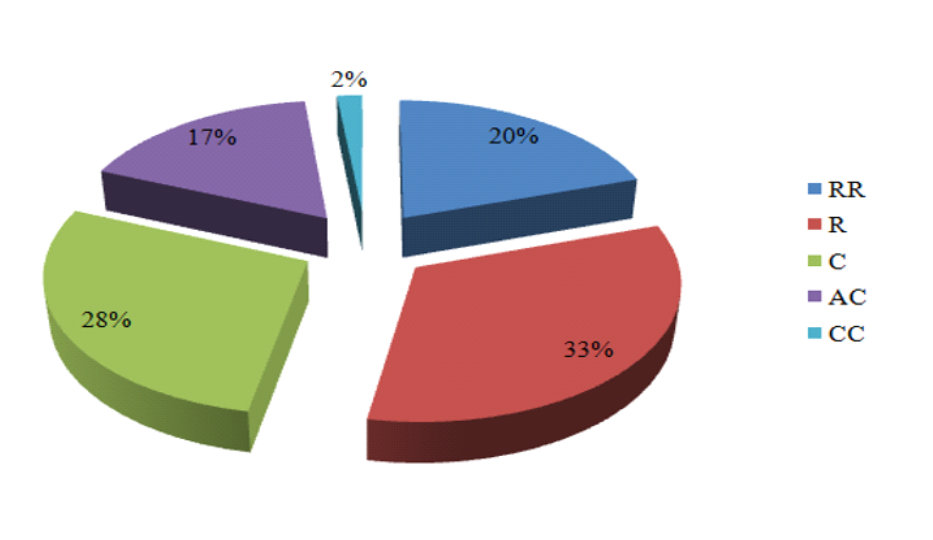
FIG. 5 - Fréquence des espèces inventoriées.
2.5 Analyse statistique
2.5.1. Interprétation de l’analyse non-symétrique des correspondances
34Sur base du test par permutations, les huit premières valeurs propres de l’analyse sont significatives. L’interprétation se base sur les coordonnées des espèces sur les axes et sur les contributions relatives. Seuls les résultats des trois premiers axes sont considérés, les autres ne produisant plus que des variations locales difficiles à interpréter.
35Le premier axe (tableaux 1 et 2 et figures 6 et 7) sépare les espèces et relevés en deux sous-ensembles. Le premier forestier, est dominé par Juniperus phoenicea, le second présaharien par Astragalus armatus.
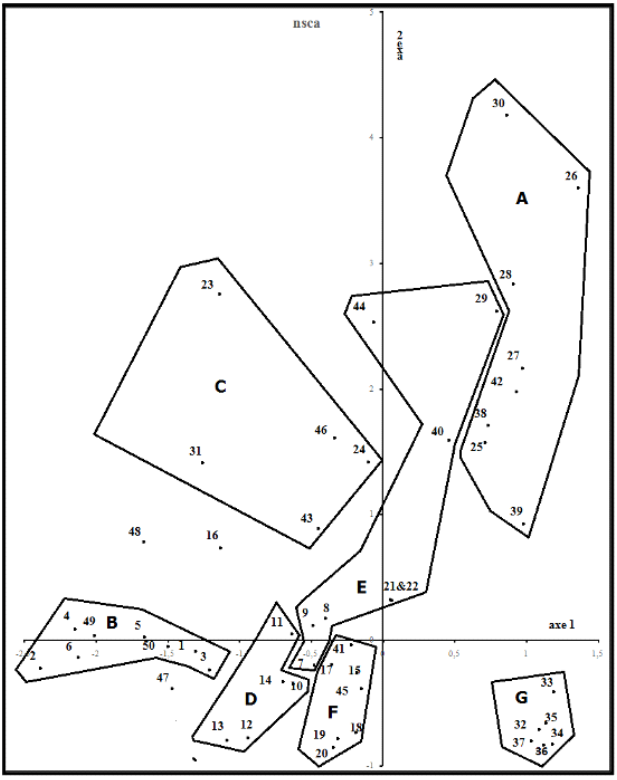
FIG. 6 - Répartition globale des relevés dans le plan des axes 1 et 2 de l’analyse non-symétrique des correspondances.

FIG. 7 - Répartition globale des espèces dans le plan des axes 1 et 2 de l’analyse non-symétrique des correspondances.
36Dans le premier groupe (tableau 2), on trouve les relevés R1, R2, R3, R4, R5, R6, R49 et R50. Les espèces qui ont les contributions relatives les plus importantes sont celles qui caractérisent la formation de forêt claire avec un faciès à Pinus halepensis, Stipa tenacissima et Juniperus phoenicea.
37Dans le deuxième groupe, on trouve les relevés R32, R33, R34, R35, R36, R37. Les espèces qui ont les contributions la plus importantes positives sont celles de formations de dégradation présahariennes, elles sont représentées par le faciès à Astragalus armatus et Bromus rubens.
38Cet axe traduit un facteur d’aridité climatique.
Tableau 1. Coordonnées et contribution relatives des espèces sur le premier axe.
|
Espèces |
coord 1 |
cr% 1 |
Espèces |
coord 1 |
C% 1 |
|
|
Astragalus armatus |
0,19 |
3,79 |
Juniperus phoenicea |
-0,30 |
9,27 |
|
|
Astragalus armatus>1 |
0,18 |
3,42 |
Jurinea humilis |
-0,23 |
5,53 |
|
|
Stripa tenacissima |
-0,22 |
4,89 |
||||
|
Pinus halepensis |
0,20 |
4,17 |
||||
|
Reseda alba |
0,20 |
3,86 |
||||
|
Stripa tenacissima>1 |
-0,19 |
3,61 |
||||
|
Astractylis humilis |
-0,19 |
3,53 |
>1 : espèces avec une abondance-dominace supérieure à 1 dans l'échelle sigmatiste. À gauche, les espèces avec les coordonnées positives et à droite, avec les coordonnées négatives.
Tableau 2. Coordonnées et contributions relatives des relevés sur le premier axe.
|
Relevés |
Coord 1 |
Cr % 1 |
Relevés |
Coord 1 |
Cr % 1 |
|
|
R35 |
1,14 |
6,77 |
R2 |
-2,38 |
7,82 |
|
|
R34 |
1,18 |
6 ,54 |
R49 |
-2,01 |
5,92 |
|
|
R36 |
1,12 |
6,16 |
R6 |
-2,12 |
4,95 |
|
|
R32 |
1,09 |
5,88 |
R4 |
-2,14 |
4,63 |
|
|
R37 |
1,04 |
5,63 |
R5 |
-1,66 |
4,29 |
|
|
R33 |
1,19 |
5,50 |
R1 |
-1,30 |
4,06 |
|
|
R50 |
-1,49 |
3,88 |
||||
|
R3 |
-1,21 |
3,47 |
À gauche, les relevés avec les coordonnées positives et à droite, avec les coordonnées négatives.
39Sur le deuxième axe (tableaux 3 et 4, figure 6 et 7), les relevés qui ont les contributions relatives les plus importantes et positives sont : R38, R42, R25, R39, R27, R28, R26, R30, ce qui représente l’ensemble floristique steppique présaharien à Astragalus armatus, Arthrophytum scoparium et Thymelaea microphylla.
40Ce deuxième axe est dominé essentiellement par un groupe d’espèces herbacées steppiques tels que Stipa tenacissima, par des espèces sahariennes tels que Pulicaria crispa et par des espèces indicatrices de dégradation tels que Peganum harmala.
41Les relevés avec des coordonnées négatives, soit R36 et R37, représentent un faciès steppique présaharien plus dégradé à Astragalus armatus et Bromus rubens. Cet axe représente donc beaucoup plus le facteur de dégradation anthropique.
Tableau 3. Coordonnées et contributions relatives des espèces sur le deuxième axe.
|
Espèces |
Coord 2 |
C% 2 |
|
Peganum harmala |
0,31 |
9,92 |
|
Pulicaria crispa |
0,29 |
8,69 |
|
Stipa tenacissima |
0,27 |
7,12 |
|
Astragalus armatus |
0,25 |
6,50 |
|
Astragalus cruciatus |
0,24 |
5,58 |
|
Astragalus armatus>1 |
0,23 |
5,18 |
|
Helianthemum hirtum |
0,18 |
3,42 |
|
Onopordum acanthium |
0,18 |
3,25 |
>1 : espèce avec une abondance-dominance supérieure à 1 dans l’échelle sigmatiste. Espèces avec des coordonnées positives.
Tableau 4. Coordonnées et contributions relatives des relevés sur le deuxième axe.
|
Relevés |
coord 2 |
cr% 1 |
Relevés |
coord 2 |
cr% 2 |
|
|
R30 |
4,18 |
14,44 |
R36 |
-0,84 |
3,40 |
|
|
R26 |
3,60 |
10,72 |
R37 |
-0,80 |
3,38 |
|
|
R29 |
2,62 |
6,94 |
||||
|
R28 |
2,84 |
5,91 |
||||
|
R44 |
2,53 |
5,31 |
||||
|
R27 |
2,17 |
4,75 |
||||
|
R42 |
1,98 |
4,32 |
||||
|
R23 |
2,75 |
4,18 |
||||
|
R25 |
1,57 |
3,62 |
||||
|
R38 |
1,71 |
3,48 |
À gauche, les relevés avec les coordonnées positives et à droite, avec les coordonnées négatives.
42Sur ce troisième axe (tableaux 5 et 6), sont mis en évidence deux ensembles végétaux steppiques, celui des relevés (R21, R22) qui correspond à la formation steppique à Stipa tenecissima et Stipa parviflora, représenté ici par les espèces Stipa parviflora et Helianthemum hirtum, qui se superpose localement à la formation steppique présaharienne à Astragalus armatus et Arthrophytum scoparium avec des espèces sahariennes telles que Pulicaria crispa et Reseda decursiva.
Tableau 5. Coordonnées et contribution des espèces de troisième axe.
|
Espèces |
Coord 3 |
C% 3 |
Espèces |
coord3 |
C% 3 |
|
|
Reseda decursiva |
0,29 |
8,16 |
Stipa parviflora |
-0,28 |
7;69 |
|
|
Jniperus phoenicea>1 |
0,23 |
5,46 |
Pulicaria crispa |
-0,21 |
4,29 |
|
|
Jupinera humilis |
0,23 |
5 ,06 |
Helianthuemum hirtum |
-0,17 |
3,c3 |
|
|
Plantago albicans |
0,21 |
4,44 |
>1 : espèce avec une abondance-dominance supérieure à 1 dans l’échelle sigmatique. À gauche, aspèces avec des coordonnées positives, à droite, avec des coordonnées négatives.
Tableau 6. Coordonnées et contributions relatives des relevés sur le troisième axe.
|
Relevés |
coord 3 |
cr% 3 |
Relevés |
coord 3 |
cr% 3 |
|
|
R47 |
3,28 |
7,89 |
R21 |
-1,87 |
9,34 |
|
|
R48 |
2,90 |
7,74 |
R22 |
-1,80 |
8,94 |
|
|
R42 |
2,19 |
5,30 |
R11 |
-1,68 |
5,98 |
|
|
R1 |
1,36 |
4,43 |
R10 |
-1,13 |
3,05 |
|
|
R38 |
1,65 |
3,24 |
À gauche, les relevés avec les coordonnées positives et à droite, avec les coordonnées négatives.
43La classification hiérarchique ascendante (figure 8) individualise sept groupements végétaux distincts sur les plans floristiques et physionomiques, il s’agit de :
44Le groupement A représente une formation présaharienne à Astragalus armatus, Arthrophytum scoparium et Thymelaea microphylla.
45Le groupement B correspond à une forêt claire à Pinus halepensus, Stipa tenacissipa et Juniperus phoenicea.
46Le groupement C correspond à une formation herbacée steppique à Stipa tenacissima, Stipa parviflora et Noaea mucronata.
47Le groupement D représente une formation post-forestière, c’est une pelouse à : Loeflingia hispanica, Noaea mucronata et Pulicaria crispa.
48Le groupement E correspond à une steppe à Stipa tenacissima et Stipa parviflora.
49Le groupement F correspond à une steppe à Astragalus armatus et Bromus rubens.
50Le groupement G correspond à une formation préforestière (matorral) à Juniperus phoenicea et Stipa tenacissima.
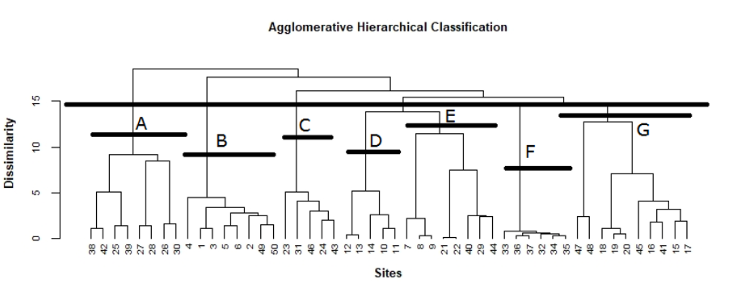 FIG. 8 – Dendrogramme obtenu par la classification hiérarchique ascendante des relevés.
FIG. 8 – Dendrogramme obtenu par la classification hiérarchique ascendante des relevés.
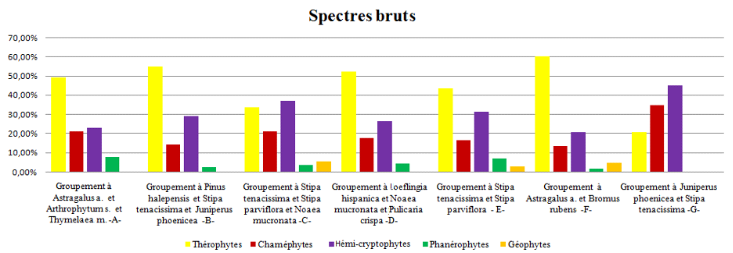 FIG. 9 - Spectres biologiques bruts de différents groupements végétaux.
FIG. 9 - Spectres biologiques bruts de différents groupements végétaux.
51Selon le spectre brut (figure 9), on a toujours la dominance des thérophytes suivie par les hemicryptophytes, puis les chaméphytes suivie par les phanérophytes et les géophytes selon le schéma suivant : Th > He > Ch > Ph > Géo, sauf dans les formations présahariennes de dégradation des groupements végétaux (A) et (E), dominées successivement par le groupement à Astragalus armatus, Arthrophytum scoparium et Thymelaea microphylla et le groupement à Astragalus armatus, Bromus rubens, qui sont représentés par le schéma suivant : Th > He > Ch > Géo > Ph, où les géophytes sont plus importants que les phanérophytes.
52En ce qui concerne les spectres réels (figure 10), on a les schémas suivants :
53Les formations forestières :
54Groupement B: Ph > He > Ch > Th > Géo
55Groupement G: Ph > He > Th > Ch > Géo
56Groupements herbacés, steppe à alfa :
57Groupement C: He > Ch > Th > Ph > Géo
58Groupement E: He > Th > Ch > Géo > Ph
59Pelouses post-forestières :
60Groupement D: Th > Ch > He > Ph > Géo
61Steppe présaharienne :
62Groupement A: Ch > He > Th > Ph > Géo
63Groupement F: Th > He > Ch > Géo > Ph
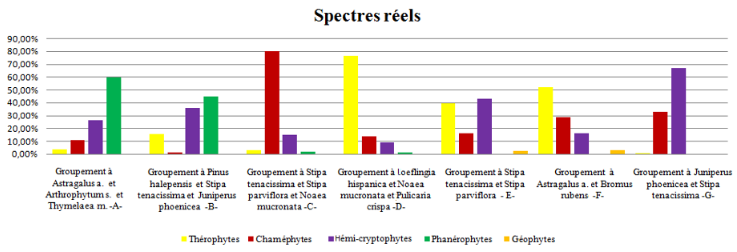 FIG. 10 – Spectres biologiques réels de différents groupements végétaux.
FIG. 10 – Spectres biologiques réels de différents groupements végétaux.
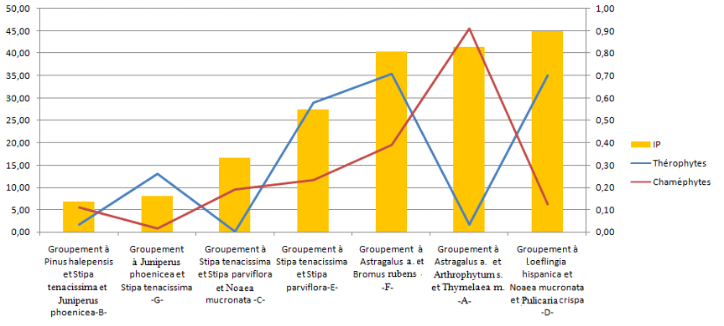 FIG. 11 - Indice de perturbation de différents groupements végétaux.
FIG. 11 - Indice de perturbation de différents groupements végétaux.
64Afin d’évaluer l’état de dégradation de chaque groupement, qui s’exprime par l’abondance des deux types biologiques, les thérophytes et des chaméphytes, et correspond aux deux phénomènes de dégradation (thérophytisation et chaméphytisation) des zones arides et semi-arides, on a utilisé l’indice de perturbation de HEBRARD et al. (1995) :
65Indice de perturbation IP = (chaméphytes + thérophytes)/nombre total d’espèces.
66Cette indice (figure 11) montre bien que les formations herbacées présahariennes, dominées par des espèces chaméphytiques à Astragalus armatus, Arthrophytum scoparium et Thymelaea microphylla et la pelouse post-forestière, riche en thérophytes, représentent les formations les plus dégradées, issues des formations steppiques à Stipa tenacissima et Stipa parviflora, qui ont subi à leur tour, ces dernières décennies, une forte pression anthropique.
67Ces groupements végétaux, qui correspondent aux diverses formations de dégradation, de notre zone d’études, représentent selon plusieurs auteurs (DJEBAILI et al., 1982 ; LE HOUÉROU, 1995 ; ACHOUR-KADI-HANIFI et al., 1997) les différents stades de dégradation des zones arides et semi-arides de la steppe algérienne.
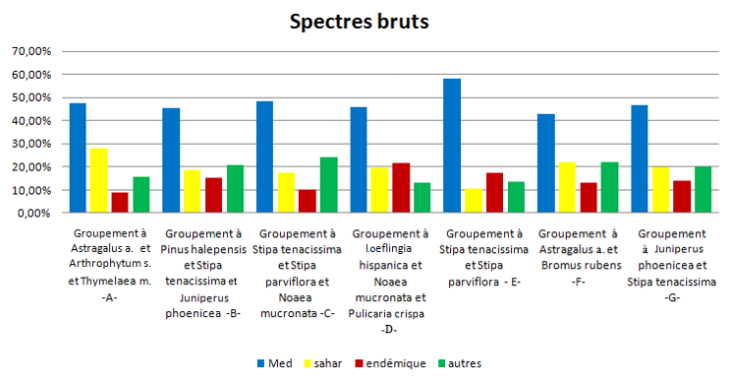 FIG. 12 - Spectres bruts phytogéographiques des sept groupements végétaux.
FIG. 12 - Spectres bruts phytogéographiques des sept groupements végétaux.
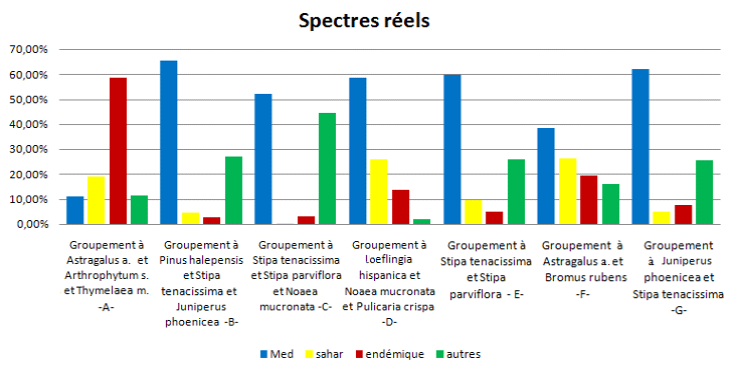 FIG. 13 - Spectres réels phytogéographiques des sept groupements végétaux.
FIG. 13 - Spectres réels phytogéographiques des sept groupements végétaux.
68En ce qui concerne la phytogéographie des groupements végétaux, on remarque bien que dans les deux spectres (figures 12 et 13), que ce soit brut ou pondéré, il y a la dominance de l’élément méditerranéen, qui confirme l’originalité floristique de notre zone d’étude.
69L’analyse phytogéographique du spectre réel de chaque groupement a donné les schémas suivants :
70Formations forestières :
71Groupement B : Med > Autres > Saharien > Endémique
72Groupement G : Med > Autres > Endémique > Saharien
73Groupement herbacé, steppe à alfa :
74Groupement C : Med > Autres > Endémique > Saharien
75Groupement E : Med > Autres > Saharien > Endémique
76Pelouse post-forestière :
77Groupement D : Med > Saharien > Endémique > Autres
78Steppe présaharienne :
79Groupement A : Endémique > Saharien > Autres > Med.
80Groupement F : Med > Saharien > Endémique > Autres
81Dans les sept groupements, l’élément méditerranéen domine tous les autres sauf dans le groupement présaharien à Astragalus armatus, Arthrophytum scoparium et Thymelaea microphylla, dominés par des espèces présahariennes endémiques de dégradation qui représente selon MELZI (1986, 1993) une formation de dégradation d’une steppe à Stipa tenacissima, qui à son tour représente une formation de dégradation d’une forêt claire du pin d’Alep et du chêne vert, (LE HOUÉROU, 1969, CELLES, 1975, DJEBAILI, 1978, AIDOUD-LOUNIS, 1984), à cause d’une aridité croissante du Nord vers le Sud, accentuée par l’action humaine.
2.6 Variation de la biodiversité par rapport à l’aridité du climat
82A partir de la figure 14, qui montre l’évolution de la diversité floristique des groupements végétaux de notre zone d’étude, représentée par la richesse floristique (S), l’indice de Shannon-Weaver (H’) et l’équitabilité (E), en fonction d’un gradient d’aridité, on a remarqué que : la richesse floristique ne suit pas toujours l’évolution de l’indice de Shannon-Weaver et la régularité, par ailleurs ces trois paramètres ne semblent pas bien corrélés avec l’aridité. Par contre, on remarque bien que ces deux derniers paramètres ont montré les valeurs les plus faibles dans le groupement à Astragalus armatus et Bromus rubens de steppe présaharienne dégradée, avec H’= 2,40 et E = 0,39 alors les valeurs les plus élevées ont été enregistrées au niveau de groupement steppique à Stipa tenacissima et Stipa parviflora avec H’= 3,34 et E = 0,55 ce qui fait que l’anthropisation affecte les valeurs de la biodiversité plus que l’aridité climatique dans notre zone d’étude.
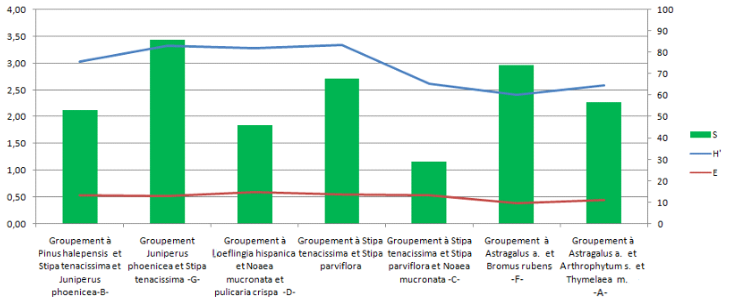 FIG. 14 - Variation des indices de diversités en fonction du gradient d’aridité.
FIG. 14 - Variation des indices de diversités en fonction du gradient d’aridité.
CONCLUSION
83L’analyse de l’état actuel de la flore des formations végétales dans la steppe sud algéroise met en évidence un état de dégradation plus ou moins avancé d’un point de vue floristique.
84A travers la présente recherche, nous avons identifié 170 taxons répartis en 34 familles et 111 genres. Ces chiffres sont plus au moins importants par rapport à d’autres études réalisées dans cette région telles que : Korichi (2016), Maamri (2016), Rahmoune (2019), Habib et al. (2020).
85Nous avons constaté dans notre zone d’étude une dominance des taxons appartenant à la famille des Asteraceae qui sont généralement des espèces à faible valeur énergétique et écologique ; c’est le cas du genre Atractylis.
86La dominance des espèces annuelles et chaméphytiques confirme les deux phénomènes de dégradation des formations steppiques, phénomène de chaméphytisation et phénomène de thérophytisation, qui caractérisent la flore des formations végétales des zones arides et semi-arides. C’est une stratégie d’adaptation des formations végétales vis-à-vis des pressions anthropiques et climatiques que subit cette région. L’analyse phytogéographique a montré l’affinité méditerranéenne de la flore de la steppe sud-algéroise, qui diminue en allant du Nord vers le Sud.
REMERCIEMENTS
87Nous remercions pour leur appui à ce travail le Dr Hadjaj Kouider, enseignant à l’Université de Djelfa-Algérie, et MM. Daoudi Belkacem et Hazerchi Ahmed du Centre National de Développement des Ressources Biologiques de Djelfa-Algérie.
Bibliographie
Aidoud-Lounis F., 1984. Contribution à la connaissance des groupements à sparte (Lygeum spartum L.) des Hauts Plateaux sud-oranais ; étude phytoécologique et syntaxonomique. Thèse 3e cycle, Univ. Sci. Technol. H. Boumediene, Alger, 253 p. + ann.
Aidoud A., Le Floc’h É., Le Houérou H. N., 2006. Les steppes arides du nord de l’Afrique. Science et changements planétaires/ Sécheresse 17 (1-2) : 19-30.
Ampapour S., 2003. Introduction à l’analyse des données, Document de Travail n° 08/2003, BAMSI, Brazzaville, p35.
Aronson J., Floret Ch., Le Floc’h É., Ovalle C., Pontannier R., 1993. Restoration and rehabilitation of degraded ecosystems in arid and semi-arid lands. I. A view from the south. Restoration Ecology 1 : 8-17.
ASAL, 2010. Agence spatiale algérienne, 2010, Finalisation de la carte nationale de sensibilité à la désertification par l’outil spatial. Algérie, ASAL. http://www.asal.dz/desertification.php.
Barbero M., Bonin G., Loisel R. & Quézel P., 1990. Changes and disturbances of forest ecosystems caused by human activities in the western part of the mediterranean bassin. Vegetatio 87 : 151-173.
Barbero M., Loisel R., Medail F. & Quézel P., 2001. Signification biogéographique et biodiversité des forêts du bassin méditerranéen. Bocconea 13 : 11-25.
Benabadji N. & Bouazza M., 2002. Contribution à l’étude du cortège floristique de la steppe au Sud d’El Aricha (Oranaie–Algérie), Rev. Sci. Techn. (Univ. Constantine) 17 : 11-19
Benabadji N., Benmansour D. & Bouazza M., 2007. La flore des monts d’Ain Fezza dans l’Ouest Algérien, biodiversité et dynamique. Sciences & Technologie 26 : 47–59.
Benaissa M., El Haitoum A., Hadjadj K., 2018. Floristic and medical diversity interest of Djebel Aissa national park (Ksour Montains, Algeria), Malaysian Journal of Fundamental and Applied Sciences 14 (2) : 303-306.
Bouxin G. 2016. Analyse statistique des tableaux de relevés de végétation/Recherche d’adéquation entre les données de végétation et les techniques statistiques, au moyen d’exemples. Éditions Universitaires Européennes, Saarbrücken, 440 p.
Bouxin G., 2020. Analyse statistique des données de végétation. 577 pp. Disponible sur Internet à l’adresse suivante : http://guy-bouxin.e-monsite.com/.
Celles J.C., 1975. Contribution à l'étude de la végétation des confins saharo- constantinois (Algérie). Thèse Doct., Univ. Nice.
Chaouche Khouane H., Belhamra M. & Aidoud A., 2018. Évolution des communautés végétales de la steppe d’Alfa (Stipa tenacissima L.) en Algérie. Courrier du Savoir 26 : 107-118.
Chauvin C. & Vallauri D., 2002. Indicateurs de restauration écologique de marnes dégradées dans les Alpes du Sud, 120 ans après reboisement. Revue Ecologique (Terre Vie), supplément 9 : 241-250.
CRBT, 1978. Centre de recherche sur les Ressources Biologiques Terrestres. Rapport phytoécologique et pastoral sur les hautes plaines steppiques de la wilaya de Saïda. CRBT, Alger, 256 pp. + Ann. & Cartes.
Daget Ph. & Poissonet J., 1971. Une méthode d’analyse phytosocioécologique des prairies. Ann. Agro 21 (1) : 5-41.
Daget Ph., 1980. Sur les types biologiques botaniques en tant que stratégie adaptative (cas des thérophytes). In : Barbault R., Blandin P., Meyer J. Actes du colloque d’écologie théorique : Recherches d'écologie théorique : les stratégies adaptatives. Maloine, Paris : 89-114.
Dagnelie P., Bonin G., Loisel R. & Quézel P., 1965. Quelques méthodes statistiques d'étude de l'homogénéité et de caractérisation de la végétation. 1er Colloque Intern. sur les « Ecosystèmes », 30 juillet 1965, Copenhague, 1-7.
Dahmani- Megrerouche M., 1997. Le chêne vert en Algérie, syntaxonomie, phytoécologie et dynamique des peuplements. Thèse. Doct. Etat, Univ. Houari Boumediene, Alger, 330 pp.
DGF (Direction Générale des Forêts), 2012. Analyse de la politique du secteur forestier et des secteurs connexes en Algérie, 81 pp. www.dgf.org.dz.
Djebaili S., 1978. Recherches phytosociologiques et phytoécologiques sur la végétation des Hautes plaines steppiques et de l'Atlas saharien. Thèse Doct., Univ. Sci. Tech. Languedoc, Montpellier, 229 pp.
Djebaili S., Achour H., Aidoud F. & Khelifi H., 1982. Groupes écologiques édaphiques dans les formations steppiques du sud-Oranies. Bulletin d’écologie terrestre. Biocénose 1 : 7-59.
Djebaili S., 1984. Steppe algérienne, phytosociologie et écologie. Office des publications universitaires (OPU), Alger, 182 pp.
Dobignard, A. & Chatelain, C., 2010-2013. Index synonymique flore d’Afrique du Nord, Conservatoire et jardin botaniques. Genève.
Floret Ch. & Pontanier R., 1982. L'aridité en Tunisie présaharienne : climat, sol, végétation et aménagement. Thèse Doc. Uni. Sci. Tech. Languedoc, Montpellier, 580 pp.
Frontier S., Pichod Viale D., Lepretre A., Davoult D. & Luczak C., 2008. Écosystèmes: structure, fonctionnement, évolution, 4eédition, Dunod, Paris, 558 pp.
Gaussen H., 1963. Ecologie et phytogéographie. In : Précis de Sciences biologiques, Botanique : 926-972.
Gimaret-Carpentier C, Chessel D. & Pascal J.-P., 1998. Non-symmetric correspondence analysis : an alternative for species occurences data. Plant Ecology 138 : 97-112.
Ghafoul M., Dellal A., Latreche A., Hadjadj K., 2019. The study of desertification in Algerian steppic rangelands. Case of the Djelfa region, Azarian J. Agric. 6 (5) : 129-138.
Gounot M. 1961. Les méthodes d’inventaire de la végétation. Bull. Serv. Carte Phytogéogr., Série B, 7 : 65-84.
Gounot M., 1969. Méthodes d’études quantitatives de la végétation. Ed. Masson, Paris, 314 pp.
Grime J.P., 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist 111 : 982, 1169-1194. https://doi.org/10.1086/283244.
Guinochet M., 1955. Logique et dynamique du peuplement végétal. Ed. Masson, Paris, 144 pp.
Guinochet M., 1973. La phytosociologie. Collection d'écologie I. Ed. Masson, Paris, 227 pp.
Habib N., Regagba Z., Djamel Miara M., Ait Hammou M. & Snorek J., 2020. Floristic diversity of steppe vegetation in the region of Djelfa, North-West Algeria, Acta Botanica Malacitana 45 : 1-10.
Hamada S., Dakki M., Ibn Tattou M., Ouyahya A. & Fennane M., 2004. Analyse de la biodiversité floristique des zones humides du Maroc, Flore rare, menacée et halophile. Acta Botanica Malacitana 29 : 43-66.
Harrane-Zaoui A., 2012. Écologie et Systématique du genre Rosmarisnus L. dans la région du sud algérois (Wilaya de Djelfa), Thèse Magister, Écologie et Environnement, Univ. Houari Boumediene, Alger, 144 p.
Hebrard J.P., Loisel R., Roux C., Goumila H., & Bonin G., 1995. Incidence of clearing on phanerogamic and cryptogamic vegetation in South Eastern France: disturbance indexes. In Bellan, D., Bonin, G. et Emig, G. coord, Functioning and dynamics of natural and perturbated ecosystems. Ed. Lavoisier, Intercept Ltd. 747-758.
Hirche A., Salamani M., Abdellaoui A., Benhouhou S., Valderrama J. M.., 2011. Landscape changes of desertification in arid areas: the case of southwest Algeria. Environ. Monit. Assess. 179 : 403-420.
Kadi Hanifi_Achour H. & Loisel R., 1997. Caractéristiques édaphiques des formations à Stipa tenacissima L. de l’Algérie en relation avec la dynamique de la végétation. Ecol. Medit. 23 : 33-43.
Kadi Hanifi_Achour H., 1998. L’alfa en Algérie. Syntaxonomie, relation milieu végétation, dynamique et perspective d’avenir. Thèse Doct. Etat. Univ. Alger, 228 pp. & Ann.
Korichi A., 2016. Etude floristique et ethnobotanique de Chouchet Tobdji (forêt de Sénalba Chergui), Région de Djelfa, Thèse Magister, Univ Djelfa, 78 pp.
Labatte J. M., 2010. Biostatistiques. Rappels des cours et travaux dirigés. Analyse des données M2. Université Anger. http://www.math.univangers.fr.pdf.
Le Houérou H.N., 1969. La végétation de la Tunisie steppique. Ann. Inst. Nat. Rech. Agron. Tunisie 42 : 624 pp.
Le Houérou H.N., 1977. Etude bioclimatique des steppes algériennes., Bull. Soc. Hist. nat. Afrique Nord, 68 (3/4) : 33-74.
Le Houérou H.N., 1995. Bioclimatologie et Biogéographie des steppes arides du Nord de l’Afrique, Diversité biologique, développement durable et désertisation. Options méditerranéennes, sér. B : recherche et études 10 : 396 pp.
Lerman I.C., 1977. Reconnaissance et Classification des structures finies en Analyse des données, rapport 70, I.R.I.S.A., Université de Rennes.
Long N., 1974. Diagnostic phytoécologique et aménagement du territoire, Vol.1. Ed. Masson, Paris, 252 pp.
Maamri F., 2016. Les espèces pastorales et fourragères originaires de la steppe Algérienne : distribution, diversité et voie d’utilisation. ENSA, Alger, 220 pp.
Madon O. & Médail F., 1996. The ecological significance of annuals on a Mediterranean grassland (Mt Ventoux, France). Plant Ecol. 129 : 189-199.
Maire R. 1952-1987. Flore de l’Afrique du Nord, (Maroc, Algérie, Tripolitaine, Cyrénaïque et Sahara). Paris, Paul Lechevalier, coll. « Encyclopédie biologique » (ISSN 03014274).
Meddour R., 2011. La méthode phytosociologique sigmatiste ou Braun-Blanqueto- Tuxenienne, Université Mouloud Mammeri, Dept. Sc. Agronom., Tizi Ouzou, 40 pp.
Medjahdi B., Letreuch-Belarouci A., Maazouz & Taïbi K., 2018. Diversité floristique des subéraies des monts de Tlemcen (Nord-Ouest Algérien) Fl. Medit. 28 : 66- 77.
Melzi S., 1986. Approche phytoécologique du processus de la désertification dans un secteur présaharien. Messaad Djelfa., thèse Magister. Univ. Sci. Technnol. H. Boumediene. Alger, 133 pp.
Melzi S., 1993. Evolution de la végétation et du milieu dans la région présaharienne des steppes algériennes. Sécheresse 4 : 113-116.
Moulay A., Benabdeli K. et Morsli A., 2011. Contribution a l’identification des principaux facteurs de dégradation des steppes à Stipa tenacissima du sud- ouest Algérien. Mediterranea, Ser. II, n° 22 : 149-188.
Morey L. C., Blashfield, R. K. & Skinner H.A., 1983. A comparison of cluster analysis techniques within a sequential validation framework. Multivar. Behav. Res. 18 : 309-329.
Nedjraoui D. & Bedrani S., 2008. La désertification dans les steppes algériennes : causes, impacts et actions de lutte. Vertigo 8 (1) : 53-75. http://vertigo.revues.org/.
Nègre R., 1966. Les thérophytes. Bull. Soc. Bot. France 113, suppl. 2 : 92-108.
Ozenda P., 1982. Les végétaux dans la biosphère. Ed. Doin, Paris, 431 pp.
Ozenda P., 2004. Flore du Sahara, 3èm ed. CNRS. Paris, 622 pp.
Pignatti S., 1982. Flora d’Italia, 1. Ed. Edagricole, Bologna.
Pouget M., 1971. Etude agro-pédologique du bassin du Zahrez Gharbi (Feuille Rocher de Sel), Echelle 1/10 000. Étude DEMRH. Alger, 160 pp. & 5 cartes couleurs.
Pouget M., 1977. Région de Messaad Ain El Ibel. Notice explicative n° 67, cartographie des zones arides. Géomorphologie, pédologie, groupement végétal, aptitude du milieu pour mise en valeur. ORSTOM., Paris, 69 pp.
Pouget M., 1980. Les relations sol-végétation dans les steppes Sud Algeroise. Trav. Docum. ORSTOM (Paris) 116 : 8-556.
Quézel P., & Santa S., 1962-1963. Nouvelle flore de l’Algérie et des régions désertiques méridionales. CNRS, Paris.
Ramade F., 2003. Ecologie fondamentale. 3éme éd. Ed. Dunod. Paris. 690 pp.
Raunkiær C.,1934. Biological types with reference to the adaption of plants to survive the unfavourable season, Ch. 1, p. 1 du recueil de travaux traduits en anglais, Oxford.
Richard C., Grégoire G. & Frédéric A., 2010. Relevés floristiques pour le suivi de la biodiversité végétale des écosystèmes forestiers : éléments de réflexion pour faire les bons choix. Rev. For. Fr. LXII (2) : 141-154.
ROSEL/OSS, 2008. Contribution Technique n° 1, CT n° 1 : Guide méthodologique pour l'étude et le suivi de la flore et de la végétation.
Roux G. & Roux M., 1967. A propos de quelques méthodes de classification en phytosociologie. Rev. Stat. Appl., 15 (2) : 9-72.
Sauvage Ch., 1961. Recherches botaniques sur les subéraies marocaines. Trav. Inst. Sci. Cherifien, Bot. 2 : 1-462.
Taibaoui B., 2008. Etude phytoécologique et diachronique de la végétation d’une steppe à Armoise blanche (Artemisia herba-alba) de la coopérative Yahiaoui (région d’Aïn Oussera, Wilaya de Djelfa). Mémoire de Magister. Univ. Sci. Technol. H. Boumedienne, Alger, 83pp. + ann.
URBT (Unité de recherche biologiques et terrestres), 1987 Rapport Phytoécologique de la Wilaya de Djelfa. URBT, Alger, 159 pp.
Vachon M., Beaulieu-Prévost D., Ouellette A. & Achille M., 2005. Analyse de classification hiérarchique et qualité de vie. Tutorials in Quantitative Methods for Psychology 1 : 25-30.
