
Qu’y a-t-il de vital dans un organisme vivant ?
Université de Nice – ceperc (cnrs, Université de Provence)
Introduction
1) Une épistémologie non fondationnelle
1§ 1. Concernant l’usage philosophique de l’adjectif « vital », on s’attend à voir surgir une distinction entre le vivant et le vécu, comme si d’emblée nous pouvions et devions accepter que le vital soit aussi quelque chose d’éprouvé par la conscience, par opposition au vivant qui serait simplement observé et expliqué par la science. Pourquoi, dès lors, faudrait-il rechercher dans ou chez les êtres vivants quelque chose de vital ? Cela ne reviendrait-il pas à multiplier les entités et à transformer subrepticement le prédicat « vivant » en être ? Cela ne nous engagerait-t-il pas vers une substantialisation de la vie érigée ainsi en réalité métaphysique ?
2N’est-ce pas justement le défaut des vitalistes comme Bergson (1907) ou Driesch, de créer un tel processus de surdétermination (Mayr, 1961 ; Kim, 1993, 1998) par et à travers lequel, à un phénomène inconnu X devrait correspondre en même temps des causes observables X’, X’’ et une force vitale inobservable Y ? Ce processus introduirait ainsi une entité supranaturelle dans le cours des événements naturels et rendrait impossible toute forme de stabilité et de généralité pour la relation associant X à X’, X’’ et X’’’.
3§ 2. Nous allons pourtant partir d’une position épistémologique inédite. Il n’est pas question pour nous d’imposer d’emblée aux biologistes l’existence d’entités supranaturelles, mais il n’est pas question non plus de les refuser a priori. Nous allons renvoyer dos à dos les deux énoncés : « Il existe des entités supranaturelles » vs. « Il n’existe pas d’entités supranaturelles ». Ce sont en effet ce que nous nommerons des « énoncés mondains ». Nous rejetterons donc toute forme d’énoncé physicaliste susceptible de nous engager ontologiquement, comme par exemple celle du type : « Les faits physiques fixent tous les faits » (Kaplan, Rosenberg, 2005). Notre point de départ sera plus modeste. Nous soulignerons simplement d’abord, comme le fit Canguilhem, que « la philosophie est une réflexion ».
4a) Nous nous inscrirons ainsi dans une tradition française, pour laquelle la matière de la philosophie n’est précisément pas initialement philosophique. Ce n’est pas l’œil du philosophe qui décrit en premier les objets scientifiques, leurs propriétés et leurs relations. Ce sont les yeux intellectuels du technicien et du savant. Pour cette « réflexion », toute « matière étrangère » est bonne. Ses premiers énoncés ne sont donc pas mondains. Ils portent sur ce que nous nommerons abstraitement des objets (parmi lesquels les objets mathématiques) pour indiquer simplement que ce n’est pas le philosophe qui les exprime. Il n’est même pas question ici de conclure en affirmant que « le monde se dissout en faits ». Nous refusons jusqu’au principe d’une telle dissolution normative, et la philosophie que nous recherchons n’est pas une grammaire transcendantale.
5On pourrait s’interroger sur ce point de départ, arguant que nous opérons une sorte de « contre-réduction » à la fois au sens analytique et au sens phénoménologique du terme et finalement peut-être assez proche de celle à laquelle tente de procéder R. Barbaras (2003). Mais nous répondrons que notre point de départ est simplement le quotidien de la recherche, de l’expérimentation et de la formalisation scientifique, celui-là même que les phénoménologues appelaient autrefois « l’attitude naturelle » (Husserl, 1907). Plutôt que de nous en écarter, nous souhaitons au contraire nous y replonger.
6b) Il ne s’agit pourtant en rien non plus d’un point de départ naturaliste. Notre intention n’est pas de naturaliser l’épistémologie (Quine, 1969) et de la dompter avec l’aide de la psychologie cognitive. Nous n’avons rien a priori contre un tel projet de naturalisation, sinon tout simplement qu’il n’épuise pas la question. De même que nous pouvons bien dire que toute morale est d’essence biologique, sans réduire pourtant la morale à la biologie, nous pouvons sans doute ajouter, que « toute bonne matière doit être étrangère » à la philosophie. C’est la réflexion qui ajoute en effet ce devoir-être philosophique à l’être des énoncés et des expériences scientifiques. Il y a bien une norme de vérité qui est propre au philosophe des sciences. Ce que nous allons dire, nous allons le dire au sujet du monde à l’intérieur duquel des objets sont analysés par le technicien et le savant. Mais cette norme, qui peut d’ailleurs emprunter elle-même des outils aux disciplines scientifiques, se constitue en creux et rétrospectivement dans les travaux de la science. Elle ne saurait prétendre au statut et à la fausse dignité d’un transcendantal.
7c) Enfin, cette épistémologie non fondationnelle ne va pas rejeter a priori le physicalisme comme méthode. Mais elle va ouvrir au rejet de la neutralisation de l’ontologie par le physicalisme méthodologique. Le physicalisme en effet, nous engage toujours ontologiquement, quoiqu’il en ait, même quand il défend l’hypothèse de neutralisation de l’ontologie. Le physicalisme nous engage ontologiquement dans une direction qui est également toujours la même : il réintroduit des effets ou des phénomènes d’imprédicativité dans les objets qu’il sert à décrire d’une manière qu’il voudrait épurée et réduite. Ce qu’il chasse par la porte, il le rapporte toujours par la fenêtre.
8§ 3. Nous allons donc refuser de chasser la vie par la porte, comme voulut autrefois le faire F. Jacob (1970). Tel est le sens de la question que nous allons poser : qu’y a-t-il de vital dans un organisme vivant ? Et nous allons prendre appui pour ce faire sur rien moins que le quotidien de la recherche, de l’expérimentation et de la formalisation scientifiques contemporaines.
9La vie revient en effet au centre même du discours et des pratiques des scientifiques en biologie et en physique et nous allons tenter de comprendre pourquoi (Morange, 2003 ; Luigi Luisi, 2006). La question du « vital », dès lors, n’est plus une simple question anthropologique. Elle redevient un problème épistémologique et métaphysique.
2) Le phénomène de vie : ni réductionnisme, ni holisme
10§ 4. La vie revient au centre du discours des scientifiques comme un phénomène et non plus comme un être métaphysique, puisque se développent depuis plus de quarante ans en biologie des sciences des origines de la vie. C’est supposer ainsi que des conditions d’observation et d’expérimentation existent qui permettent de rendre compte du passage du non-vivant au vivant (Maynard-Smith, 2000 ; Morange, 2003). Mais de quel type de phénomène s’agit-il ?
11Dans un article récent et à paraître, Ch. Malaterre propose un schéma explicatif, à travers lequel l’émergence de la vie semble pouvoir être considérée comme le résultat d’une évolution en trois étapes. Il y a celle des briques prébiotiques avec la fabrication des premiers acides aminés et des premiers nucléotides comme l’adénine, puis la formation des macromolécules fonctionnelles, comme les ARN, et enfin les propriétés auto-organisationnelles comme la réplication. On pourrait alors, au terme de ces trois étapes, considérer la vie comme une disposition émergente, comme une propriété de second ordre qui déterminerait les changements de propriétés des éléments qui composent l’organisme qui la possède, et comme s’il y avait ainsi une fonction vitale que l’on pouvait parvenir à décrire.
A disposition is a property (such as solubility, fragility, elasticity) whose instantiation entails that the thing that has the property would change, or bring some change, under certain conditions (Love, 2003).
12Ce n’est pourtant pas ce que conclut Malaterre, puisque le modèle qu’il propose est plus complexe que ce langage fonctionnaliste sommaire. À chaque niveau d’analyse en effet, et depuis le départ, des conditions aux limites viennent s’ajouter aux propriétés caractérisant les objets scientifiques étudiés. Pour la formation de l’adénine par exemple, à partir de la polymérisation de l’acide cyanhydrique, elle est impossible sans que l’eau puisse se changer en glace, de telle sorte que l’acide se trouve piégé et concentré dans les cristaux de glace. Quant à la formation des molécules fonctionnelles — les premiers ribozymes — elle passe par un processus de sélection-amplification. Malaterre ne fait ainsi que prolonger l’idée initiale de G. Cairn Smith qui faisait de l’hérédité enfermée dans des molécules d’ADN ou d’ARN le résultat d’un processus démarrant à partir de gènes minéraux, de séquences apériodiques de cristaux d’argile empilés les uns sur les autres.
13Il n’en reste pas moins que cette structure en marchepied suit selon l’auteur un ordre à la fois logique et temporel. Telle est son hypothèse d’analyse. Elle laisse entendre qu’en remontant jusqu’aux briques élémentaires, il est possible ensuite de redescendre jusqu’aux propriétés auto-organisationnelles, comme la formation d’une cellule ainsi que la disposition ou aptitude qu’elle aurait de se répliquer. Cette structure en marchepied n’est donc rien d’autre que ce que Kim nomme un modèle stratifié (1999).
14§ 5. Or si l’on veut éviter les effets d’imprédicativité dus à l’introduction de tels modèles, il est prudent de partir plutôt de phénomènes de couplage (Morange, 2003).
15Traditionnellement, on définit la différence entre vivant et non-vivant par « la sélection naturelle » de « variations héréditaires favorables ». On retrouve cette définition chez Sober (1984), qui privilégie la « fitness », comme indicateur pour marquer la différence entre un système physique et un système biologique. Mais il faut que ces variations favorables puissent se multiplier, ce qui suppose autre chose qu’un simple processus de croissance. Comment donc l’hypothèse de sélection naturelle pourrait-elle servir à comprendre comment ce processus se met en place, si le concept de sélection naturelle présuppose préalablement l’existence de ce processus. Impossible de répondre, si on en reste au seul niveau de la définition classique et darwinienne du concept de sélection.
16De même, l’idée « d’un système chimique auto-entretenu » est loin d’être claire et suffisante pour définir la vie. La flamme d’un bec Bunsen n’en constitue-t-elle pas un exemple parfait ? Dans le même ordre d’idées, Gaia est bien un système autorégulé de ce genre. Mais ce n’est pas un être vivant. Pourtant cette formule ambiguë contient des indications importantes. Il n’y a pas de vie sans métabolisme entendu à la fois comme « capacité d’auto-renouvellement » des molécules d’un système vivant (la nutrition) et comme persistance ou encore perpétuation « d’une même architecture complexe » (l’organisation).
17En revanche, si nous plaçons ensemble, sur le plan du métabolisme, une structure en réseau de molécules rendant possible le passage de l’autocatalyse à la catalyse croisée, d’une part (Von Kiedrovski, 1994) et l’action d’un processus de sélection qui n’est pas encore la sélection naturelle de l’autre, n’est-il pas imaginable de comprendre comment, à partir de simples processus de croissance, les premiers schémas de réplication peuvent se mettre en place ? Il est vrai que pour accepter une telle vision des choses, il faut imaginer une forme première de sélection qui n’agit pas encore véritablement sur des variations héréditaires. Mais une telle idée, clairement non darwinienne, n’est pourtant pas absurde. Elle répond à ce que Spencer (1862, § 162, § 166) nommait déjà un principe de « ségrégation ».
18Mais que faut-il entendre par couplage ? Notre objectif n’est pas ici de donner une réponse complète à cette question. Nous allons défendre une position épistémologique plus large. Les balbutiements de la science des origines de la vie traduisent le fait que :
19(1) Le vivant vient du non-vivant.
20(2) Mais en même temps la vie est une propriété spécifique de certains systèmes complexes.
21Se pose alors la question : que faut-il entendre par « systèmes complexes » ? La réponse que nous proposons est la suivante :
22(3) L’explication d’un phénomène complexe de ce type ne saurait être ni réductionniste, ni holiste.
23Mais si nous proposons cette explication, c’est simplement parce que nous la trouvons régulièrement aujourd’hui dans la littérature scientifique, tant au niveau expérimental, qu’à celui de la modélisation théorique (Kauffman, 2000 ; Noble, 2006 ; Lesne, 2006 ; Soto, Sonnenschein, 2004).
24Chez Noble, cette formule a une valeur heuristique. Elle traduit le fait que l’explication scientifique ne doit partir ni d’en haut, ni d’en bas, mais du centre. Elle est un cercle dont le centre est partout et la circonférence nulle part (« middle out »). Cette métaphore désigne le fait que le problème physiologique complexe qu’il se pose (comment fonctionne le muscle cardiaque) requiert une simulation multi-échelles. Cette simulation tient compte de l’effet qu’entretiennent les uns sur les autres des processus qui se déroulent au niveau cellulaire et des processus qui apparaissent en même temps au niveau moléculaire. Mais elle a également une valeur ontologique, selon Noble. Si la modélisation du rythme cardiaque requiert une approche multi-niveaux du fait que le potentiel électrique créé dans la cellule agit sur le transport des molécules chargées électriquement dans les canaux ioniques, de même que le transport de ces molécules de calcium ou de potassium interagit avec la forme même du potentiel électrique, c’est parce que le biologiste a affaire à des effets de causalité réciproque complexes. Non seulement le système agit et modifie les propriétés de ses éléments en produisant ainsi un effet de retour ou de causalité descendante, mais les éléments sculptent et façonnent l’allure générale du système.
25Peut-on lui emboîter le pas et dire plus généralement que l’explication du phénomène de vie ne saurait être ni réductionniste, ni holiste, tant au niveau de l’individuation d’un organisme vivant qu’au niveau de l’évolution ? Prenons tout de suite une précaution, avant de répondre à cette question. Notons, avec Morange, que le développement, comme le vieillissement, sont des phénomènes évolutifs tardifs et qu’ils ne sauraient servir de modèles exclusifs pour comprendre la vie. Si le problème que nous posons a un sens fondamental, c’est au niveau évolutif que nous devons nous en apercevoir. Telle est en tout cas la démarche que nous proposons dans cet article. L’ontogénie récapitule peut-être la phylogénie, mais elle n’en fournit en rien une explication, même rétrospective.
1. Il n’y a pas de molécules de la vie
§ 6. Y a-t-il des primordia rerum en biologie ?
26Commençons par souligner les limites de l’approche réductionniste classique. Qu’entendre ici par réductionnisme, ou par approche du bas vers le haut ? On pourrait répondre qu’il s’agit d’identifier le tout à partir de la description de ses parties, de leurs propriétés et de leurs relations. On pourrait donc supposer qu’il y a des « éléments fondamentaux » définissant un système biologique et que ces éléments sont les gènes contenus dans les molécules d’ADN ou d’ARN. C’est exactement ainsi que D. Dennett s’exprime encore aujourd’hui :
27(4) x is biologically possible if and only if x is an instantiation of an accessible genome or a feature of its phenotypic products (Dennett, 1996, p. 118).
28C’est au niveau des gènes ou encore du génome, qu’il faudrait donc décrire l’espace des possibles en biologie et il y aurait ainsi un espace des possibles susceptible d’être décrit. L’évolution n’apparaîtrait alors que comme un portrait dans cet espace.
29Mais que faut-il entendre par « gène », ou encore « génome » ? S’agit-il d’une fonction ? Par exemple la fonction : « X code pour la synthèse d’une protéine » ? Ou s’agit-il du support matériel chimique ou encore ce que l’on pourrait nommer « le réaliseur » de cette fonction, c’est-à-dire par exemple un triplet de nucléotides dans le brin d’ADN ? On voit immédiatement ici une série de grandes difficultés surgir. Faut-il parler de molécules de la vie (l’ADN, par exemple) ? Ou faut-il parler des fonctions de base grâce auxquelles la vie prend naissance ?
§ 7. La survenance méréologique et l’émergence faible
30On pourrait imaginer pouvoir opérer une certaine simplification du problème réductionniste en biologie moléculaire.
31On pourrait supposer d’abord qu’il y a bien une différence irréductible entre la fonction (le gène) et le support matériel de la fonction (le triplet de nucléotides).
32On pourrait admettre également qu’une même fonction peut être accomplie avec différents supports (multi-réalisabilité), ce qui serait dire que la fonction est une propriété qui ne peut pas être « déduite » de la propriété de base, mais qui « survient » à partir de celle-ci :
B survient faiblement sur P, si et seulement si nécessairement, pour toute propriété F en P, si un objet x a F, il existe une propriété G en B, telle que x a G, et si tout y a F, il a aussi G. (Kim, 1993)
33On pourrait supposer, enfin, qu’en mettant en relation entre eux des éléments déjà dotés de ces fonctions élémentaires, un nouveau niveau de survenance pourrait émerger, la survenance méréologique :
Systems with an identical total microstructural property have all over properties in common. Equivalently, all properties of a physical system supervene on, or are determined by, its microstructural property (Kim, 1999).
34Si tel est le cas, à partir d’une approche « bottom-up » initiale, nous pourrions ensuite examiner par une approche « top-down », comment un système d’éléments envisagé comme un tout produit des effets physiologiques mesurables et observables.
§ 8. L’exemple de l’opéron lactose
35Le modèle de l’opéron lactose peut en fournir une illustration. Par lui, Monod explique comment la cellule bactérienne se nourrit. Il l’explique par le simple agencement approprié des gènes de régulation et des gènes de structure, comme s’il s’agissait là d’un algorithme d’ordinateur. L'ensemble O-Z-Y-A constitue ainsi une unité génétique à expression coordonnée ou opéron. Z-Y-A sont des gènes de structure. (R) est un gène régulateur qui code pour la synthèse d’une protéine allostérique répresseur qui se fixe sur le site opérateur et empêche la transcription des gènes Z-Y-A.
36En présence d'inducteur (le bétagalactoside), le répresseur (R) change de forme. Il est inactivé et n’est donc plus fixé sur l’opérateur. Le déplacement de l’ARN-polymérase est rendu possible. Il permet la synthèse des ARN messagers de la bétagalactosidase qui hydrolyse le bétagalactoside, et de la perméase qui fait pénétrer celui-ci dans la membrane bactérienne d’E. Coli.
37L’avantage de ce schéma est qu’on peut effectivement identifier ici un niveau de base, le niveau des gènes de régulation et de structure. Ils sont décrits, leur fonction est décrite ainsi que leurs relations. On peut en conclure d’une certaine manière que la fonction « nutrition de la bactérie » survient ici de manière méréologique à partir de ce niveau de base, conformément aux propositions de Kim.
§ 9. Où est le niveau de base ? De l’imprédicativité en biologie moléculaire
38La première difficulté, bien sûr c’est la multiplication des niveaux de base. S’agit-il d’un niveau de base structurel, par exemple les triplets de nucléotides ? En ce sens, le niveau de base, c’est celui des molécules de la vie. Mais le niveau de base peut aussi être fonctionnel, ce sont alors les gènes de régulation et de structure. Admettons cependant que tout cela s’empile merveilleusement. Il nous faut encore ici parler d’un niveau de base par analogie. De même qu’il y a des atomes en physique, il y aurait les molécules de la vie.
39Mais nous avons des problèmes avec le niveau de base en biologie. Monod l’évoquait lui-même. Il n’y a pas d’unité génétique définissable sans facteurs de transcription ou de réplication. Les gènes dépendent donc eux-mêmes des protéines qu’ils synthétisent. Le problème s’est largement compliqué depuis Monod, puisqu’il y a en fait un nombre considérable de protéines qui interviennent. La fonction de base « coder pour la synthèse d’une protéine » est définie par la fonction supérieure (« réguler l’action des gènes par les protéines »).
40On pourrait penser alors que nous n’avons pas trouvé le niveau fonctionnel pertinent. Plutôt que d’insister sur les gènes, regardons comment les gènes s’allument et s’éteignent. Nous voyons alors qu’il existe des groupements chimiques épigénétiques sur les queues des histones qui semblent jouer un rôle important en ce qui concerne le passage d’un état chromatinien à un autre. Notons que nous focalisons ainsi notre attention sur les eucaryotes, ce qui est un premier point. Mais le problème fondamental vient plutôt de ce que nous rencontrons exactement la même difficulté. Par exemple on peut relier l’état hétérochromatinien à la méthylation, tel semble être le cas en ce qui concerne la méthylation de H3K9 (la lysine 9 de l’histone 3). Le processus semble fonctionner à partir d’une méthyltransférase. Il induit une méthylation de l’ADN qui provoque une déacetylation des histones. Il semble aussi être réversible, comme la littérature l’indique aujourd’hui (Wang, 2004). Mais on a souligné, à juste titre, que l’ouverture et la fermeture de la chromatine dépendaient ainsi en dernière instance des facteurs de transcription à l’origine de la synthèse des déacétylases ou des méthyltransférases. Une nouvelle fois, donc, le serpent se mord la queue. Il faut des gènes actifs et des protéines synthétisées pour que les groupements épigénétiques puissent produire leurs effets. D’autant que la déacetylation de H3K9 est elle-même un préalable à la méthylation. La méthylation engendre la déacetylation car elle est elle-même rendue possible par la déacetylation.
41On pourrait remonter plus haut encore pour indiquer la difficulté qu’il y a à trouver les niveaux de base en biologie. Ainsi il faut une membrane basale pour que le stroma et les cellules épithéliales aient une architecture fonctionnelle normale. Et pourtant c’est à partir des cellules épithéliales et du stroma que la membrane basale est constituée. Ce problème n’est donc pas local, il est généralisé. Nous allons l’appeler le problème de l’enchevêtrement. Ce qui vient d’abord en biologie, ce n’est pas ce fameux niveau de base, sorte d’introuvable serpent de mer. Ce sont plutôt les enchevêtrements.
42On pourrait toujours arguer enfin que ce problème ne peut pas se résoudre au niveau des fonctions de l’ADN, mais qu’il peut trouver une solution si nous estimons que l’ADN lui-même est un produit de l’évolution et de l’action de la sélection naturelle. Mais le concept même de sélection naturelle présuppose l’usage de variations héréditaires, donc génétiques. Il faut remonter loin pour éviter les enchevêtrements et supposer que la sélection naturelle agit déjà sur des objets qui ne sont pas encore véritablement des gènes qui ne peuvent donc pas être le niveau le plus bas, le niveau de base.
2. L’énigme des invariants et la faillite du holisme
§ 10. De la difficulté de dégager des invariants fondamentaux en biologie
43On peut supposer que si nous ne connaissons pas tous les éléments d’un système, nous pouvons néanmoins en dégager les invariants et le définir ainsi en tant qu’il forme un tout, même si les propriétés de ce tout ne se réduisent pas à la somme de ses éléments.
44Cette supposition a sans doute un sens profond en physique. Elle est à l’origine du courant énergétiste. Elle est défendue par Duhem et Mach. L’invariance de l’énergie et la loi d’entropie peuvent suffire à prévoir comment un système se transforme, sans faire d’hypothèses sur les éléments ultimes qui le composent. Mais elle a également un sens en biologie et jusque dans le projet fondateur de la biologie moléculaire :
Nous réservons pour un chapitre ultérieur l’exposé et les justifications de l’hypothèse considérée comme seule acceptable aux yeux de la science moderne : à savoir que l’invariance précède nécessairement la téléonomie (Monod, 1970).
45Mais comment s’y prendre pour dégager des grands invariants en biologie ? Des invariants de quoi ? Les gènes ? Les combinaisons possibles d’ADN ou d’ARN ? Mais pourquoi ces acides aminés plutôt que certains autres ? On sait aujourd’hui que l’ADN a existé après l’ARN, ce qui suppose déjà que le code génétique actuel n’est pas ce qu’il était initialement. Mais pourquoi le code génétique ? Cairns Smith parle de gènes minéraux. Ce qui vient en premier n’est pas la notion d’invariant, mais plutôt celle de variation et de corrélation.
§ 11. Systèmes conservatifs et systèmes dissipatifs
46On peut néanmoins avancer sur un premier point important. Les systèmes biologiques ne sont pas conservatifs. Ils sont dissipatifs. Cela ne signifie pas que tous les systèmes dissipatifs soient déjà biologiques. On peut tabler sur cette hypothèse, en tout cas.
47On peut accepter l’idée que le niveau de base des phénomènes biologiques c’est la thermodynamique loin de l’équilibre. Mais qu’entend-on alors par niveau de base ? Qu’est-ce qu’une condition de ce type ?
48i) Prigogine a montré que les systèmes conservatifs avaient tous la même forme. Tel est le cas pour la conservation de l’énergie mécanique. C’est la forme qui permet de les décrire en remplaçant des équations différentielles du premier ordre par des équations du second ordre, avec pour la mécanique classique :
(5a) 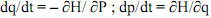
(5b) 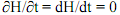
49On retrouverait le même type de substitution si on prenait en exemple l’équation de Schrödinger. L’idée est d’expliquer la forme du mouvement, ou d’une évolution de telle sorte que le temps n’agisse pas et qu’il puisse être éliminé des équations. C’est évidemment la solution la plus simple pour obtenir des invariants et pour dégager des symétries spatio-temporelles qui supposent en effet que l’on paramètre le temps de la même manière que l’espace, c’est-à-dire comme une variable indépendante.
50Maintenant prenons un pendule simple placé dans un champ de pesanteur et soumettons le point d’origine à un mouvement vertical alternatif. Le paramètre g va varier avec le temps, de sorte que l’équation initiale :
(5c) 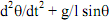
51Est transformée en :
(5d) 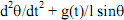
52(5d) n’est alors plus intégrable par une loi g(t) quelconque.
53ii) Mais il y a une seconde difficulté liée à l’usage de ce que Prigogine (1989) nomme « les contraintes externes ». Les équations des systèmes dissipatifs peuvent s’écrire dans le langage des mathématiques, sous forme d’équations différentielles. Mais elles ont toutes une forme de type
(5e) 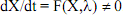
Avec 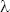 comme paramètre de contrôle ou contrainte externe. Puisque la dérivée n’est pas nulle, nous sommes hors équilibre.
comme paramètre de contrôle ou contrainte externe. Puisque la dérivée n’est pas nulle, nous sommes hors équilibre.
54Le principe c’est justement de ne jamais considérer un système en tant qu’il forme un tout, mais plutôt comme « un sous-système d’un système plus vaste qui agit sur lui ». Les paramètres de contrôle sont précisément l’expression de cette action. Le résultat de cette opération, c’est qu’il est toujours possible d’écrire comment le système en question évolue sous forme d’équation, mais l’explication fournie sera toujours limitée, contextuelle et teintée d’une hétérogénéité constitutive de ces systèmes.
§ 12. Le rejet du principe de complétude et de clôture causale du niveau de base
55On pourrait rétorquer que l’écart qui se creuse ici entre déterminisme et prévisibilité vient de cette structure en « système faisant en même temps partie d’un système plus vaste » et que cet écart n’existe que du point de vue de l’observateur qui dissocie les deux. Définissons au contraire un point de vue de la Nature, un point de vue de l’univers. L’écart n’existe plus si la nature est un système qui n’a pas de bords et qui est caractérisé par l’ensemble de ses contraintes internes. Telle est bien la position de J. Bricmont, héritée en droite ligne de Lloyd Morgan et de Laplace : « In a nutshell, determinism has to do with how the nature behaves, and predictability is related to what we human beings are able to observe, analyse and compute » (Bricmont, 1996).
56Cette position prend appui sur le double principe de complétude et de clôture causale du niveau de base (Kim, 1999 ; Rosenberg, 2005).
57Nous pouvons supposer en effet, dans le sillage de Lloyd Morgan, que si nous connaissons l’ensemble des contraintes de base qui caractérisent la nature comme système physique et donc les invariants fondamentaux à partir de quoi tout mouvement et tout changement sont expliqués, une parfaite prévision découlera de l’explication que nous fournirons.
58Si tel est le cas, le niveau de base, ou niveau physique apparaîtra aussi saturé causalement. Il nous faudra dire que tout phénomène biologique ou psychique est aussi un phénomène physique, et qu’en tant que phénomène biologique ou psychique qui est aussi un phénomène physique, il n’ajoute rien à l’explication qui le relie à d’autres phénomènes physiques, sous la forme de propriétés, de lois et d’invariants mesurables. Si nous le refusons en effet, nous allons encore mettre en relation un effet inconnu X, simultanément avec des causes X’ et X’’ et avec une cause Y agissant au niveau supérieur. Nous recréerons ainsi les conditions d’une « surdétermination causale ».
59Bien entendu, le « nous » capable d’énoncer cet ensemble de contraintes physiques caractérisant « la manière dont la nature se comporte » ne peut pas être un observateur humain. C’est bien au contraire le démon laplacien lui-même dont il est explicitement question dans le texte de Bricmont : « The best way to explain is to go back to Laplace » (Bricmont, 1999). Mais nous arrivons ainsi à la pire des contradictions : il faut un œil métaphysique pour réduire le monde à l’ensemble de ses contraintes physiques. Cette réduction réalise donc le contraire de ce qu’elle vise.
60Et l’hypothèse de la neutralité ontologique de la conjecture physicaliste ne nous aide en rien. Quand nous posons comme exigence simplement méthodologique que le monde se dissout en faits physiques et que les faits biologiques sont aussi des faits physiques et non l’inverse, nous posons une exigence non pas de notre point de vue d’observateur, mais d’un point de vue qui ne serait pas notre point de vue d’observateur. Nous sommes ainsi forcément engagés ontologiquement, puisque notre méthodologie nous oblige à introduire un être métaphysique au sein de la nature. Cela peut être une simple fiction utile, mais elle est nécessairement engageante. Nous allons refuser ce point de vue, de par notre approche non fondationnelle. Notre point de départ ne peut donc être aucune conjecture au sujet du monde, même pas qu’il se dissout en faits. Notre point de départ n’est rien d’autre que le travail du physicien et du chimiste. Et que vient de dire le physicien ? Que les systèmes clos ne sont que des cas particuliers et idéalisés. Les systèmes ouverts sont au contraire la règle. Cette règle est donc aussi la nôtre. C’est la règle de non-séparabilité.
§ 13. L’hypothèse de la non-séparabilité revue et corrigée
61Si donc on entend par niveau de base celui des propriétés d’un système dissipatif, on voit que nous parlerons bien là d’un niveau essentiellement incomplet à la base.
62a) C’est vrai que le pendule dont on perturbe le mouvement obéirait à l’équation précédemment décrite si on ne le perturbait pas (argument de Bricmont). Mais cet argument n’a aucun sens, car ce mouvement non perturbé est lui-même une idéalisation. Dans la nature, le pendule subirait les frottements de l’air et connaîtrait la dissipation d’une autre manière. Il en est de même, quoique d’une manière bien plus complexe quand on étudie les convections de Bénard. L’explication est toujours limitée par l’exigence d’avoir recours à des contraintes externes. Dans le cas des convections de Bénard, ce sera l’explication en termes statistiques du comportement d’une population de molécules d’eau qui n’est pas non plus ce qu’il devrait être.
63b) On voit qu’il faut également renoncer à la description complète des éléments qui constituent le niveau de base. Dans un système dissipatif perturbé artificiellement par une contrainte externe, que cela soit une population de poissons ou un nuage de molécules, l’action des paramètres de contrôle (pour certaines valeurs de ceux-ci) sur une perturbation infinitésimale peut avoir des conséquences finies. C’est l’effet de SCI (sensibilité aux conditions initiales). Il y a donc clairement violation de la complétude du niveau de base, mais pour des raisons internes au système que l’action de la contrainte externe met à jour.
64c) Pourtant nous sommes encore loin de notre problème et il n’y a pas véritablement de causalité complexe dans de tels systèmes. Le principe selon lequel le futur ne peut agir sur le présent n’est pas violé. Les lignes qui composent la trajectoire de phase de tels systèmes ne sauraient se recouper en un point. Il y a dilatation (SCI) et il peut y avoir contraction (Attracteurs), mais pas de recoupements. Donc pas de violation du principe de causalité au niveau de la dynamique de ces systèmes. Création d’information nouvelle sans violation du principe de causalité. Ce sont des systèmes déterministes, en ce sens.
3. Génétique moléculaire et sélection naturelle
§ 14. Stochasticité et sélection naturelle
65Nous sommes encore loin de la non-séparabilité au sens de ce que les biologistes appellent « couplage », ou « bricolage ». La nouvelle prémisse que nous allons ajouter ici est bien connue : les systèmes vivants ne sont pas entièrement déterministes, mais ils ne sont pas non plus entièrement stochastiques. L’idée vient de Darwin.
66Commençons par reprendre ce que nous disions plus tôt au sujet de l’imprédicativité des propriétés biologiques qui se prédiquent elles-mêmes, qui sont « autoréférentielles ». Tâchons de remonter jusqu’au niveau où nous étions restés alors, celui de la sélection naturelle. Peut-être peut-on déjà parler de sélection naturelle sans gènes, mais avec simplement une sélection de structures identiques qui se compliquent.
67Darwin a formulé un concept crucial. Il disait : « C’est la relation entre les variations héréditaires et l’action de la sélection naturelle qui est accidentelle », mais pas au sens où on le croit, pas au sens d’un ingénieur qui utiliserait des pièces qu’il n’a pas fabriquées lui-même pour construire une maison, pas au sens du fonctionnalisme. Non, Darwin disait que les pièces étaient mal taillées, au sens où elles n’étaient pas taillées dans le but de servir à quelque chose. On peut traduire cela par le fait que la sélection naturelle agit comme une contrainte externe, mais qu’elle agit en plus sur une structure qui se réplique et qui se modifie de manière aléatoire. Ce point vient de connaître une tentative de formalisation de la part du biophysicien chinois Ao (2005).
(6) 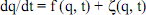
Le paramètre q est la fréquence de l’allèle d’un gène par rapport à tous les autres allèles. L’équation décrit comment q varie au cours du temps sous l’effet de la pression de sélection qui est décrite par la première fonction (f), mais s’y rajoute une seconde fonction à caractère stochastique (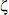 ), correspondant déjà à ce que Fisher (1930) appelait les effets de détérioration de l’environnement, c’est-à-dire toutes les autres formes de variances qui venaient interférer avec la variance additive, notamment les effets d’interaction épistatiques et alléliques. D’où l’idée de bricolage et de couplage.
), correspondant déjà à ce que Fisher (1930) appelait les effets de détérioration de l’environnement, c’est-à-dire toutes les autres formes de variances qui venaient interférer avec la variance additive, notamment les effets d’interaction épistatiques et alléliques. D’où l’idée de bricolage et de couplage.
§ 15. La protéine RecA et la boîte SOS
68Je vais montrer maintenant, dans un langage plus contemporain, que les trois niveaux des gènes (individus), des populations de gènes (espèces) et des relations entre espèces (milieux, niches) s’enchevêtrent et qu’il nous faut penser cette interaction. Je vais développer trois points en prenant comme exemple les bactéries, cette fois.
691) Le premier point est la boîte SOS de réparation de l’ADN chez le Colibacille ou la Salmonelle. En interaction avec l’environnement, un système est activé qui va induire une série de mutations ciblées et non ciblées sous l’action de protéines codées par des gènes mutateurs (Radman, 1999). Une protéine RecA se transforme en co-protéase. Elle clive une autre protéine LexA qui réprime la synthèse des gènes mutateurs en polymérases 4 et 5. Il est donc impossible de traduire ces mutations comme soumises aux seules lois du hasard. Il y a une dynamique de transformation des éléments sur lesquels la sélection naturelle agit. L’action du niveau supérieur implique bien une modification des éléments qui constituent le niveau de base.
702) Mais le second point consiste en ce que ces modifications ont une résonance évolutive, puisqu’elles fabriquent ce que Radman (1999) va nommer de « l’adaptabilité ». Ces mutations n’interviennent en effet qu’au cours d’une période critique où le stress bactérien empêche le processus de division cellulaire de s’effectuer normalement. Elles induisent de l’adaptabilité sans payer le prix d’un fardeau de mutations trop lourd. Cette modification induit l’émergence d’un processus populationnel nouveau (evolvability).
713) Il est impossible de dissocier complètement le niveau des réseaux de gènes, celui des individus d’une espèce, et le niveau interspécifique, puisque ce qui se produit ici pour une espèce a en même temps des répercussions pour une autre.
72En cas de stress UV, en effet, la protéine RecA activée clive le répresseur chez le Bactériophage Lambda (Brotcorne-Lannoye et al., 1986). Le génome du phage s’exprime et un cycle lytique est amorcé. C’est ainsi le même système qui permet à la bactérie de survivre (la proie) qui facilite aussi la survie de son prédateur (le phage). Ces modifications génétiques vont permettre l’émergence d’un processus écologique proie-prédateur.
73On a donc là typiquement un phénomène d’intrication entre des processus qui se développent à une échelle et d’autres qui se développent à une autre échelle, simultanément. C’est cela qui induit ce que Kim (1999) appelle un effet de « causalité descendante synchronique réflexive ». Il ne pose plus de problème, cette sorte d’imprédicativité essentielle (et non accidentelle) ne pose plus de problème, si nous renonçons à inclure notre système d’observation défaillant à l’intérieur d’un monde physique parfait qui l’englobe. Tout au contraire, un système, puisqu’il est en même temps un sous-système de système, a interminablement des bords qu’il nous faut dé-border. Cet énoncé si simple a des implications ontologiques essentielles (Barbaras, 2003).
§ 16. Émergence faible
74Derrière ces concepts de bricolage, de couplage, et maintenant de non-séparabilité et d’émergence diachronique, il nous semble qu’apparaît un vrai problème épistémologique qui prend une double forme. La première est l’action du temps et la seconde la non-séparabilité, comme problème ontologique et non pas seulement comme problème épistémologique.
75a) Le problème de la surdétermination causale perd toute valeur du fait de cette action. Il prend appui sur les deux postulats de complétude et de clôture causale du niveau de base, on pourrait aussi les nommer des postulats d’intemporalité. C’est supposer, comme le reconnaît volontiers Kim (2006, 34), qu’il y a une « relationnalité » « relatedness » (concept qui vient de Lloyd Morgan) du système qu’on analyse et qu’il est possible d’en dégager les contraintes internes et les invariants fondamentaux. Mais une telle vision des choses devient absurde dès que nous évoquons un système dynamique, avec conditions aux limites et paramètres de contrôle, ainsi que la propriété caractéristique d’un tel système de ne valoir que comme sous-système d’un système plus vaste et du point de vue de la pratique d’un expérimentateur (Stengers, 1997). Dès que nous acceptons ainsi que la définition des objets est contextuelle et qu’il y a dans notre explication du système lui-même un point de vue du système et un point de vue de l’observateur, il n’y a plus d’exigence de se référer à un principe de complète intelligibilité, ce qui est un premier point. C’est même la définition du système qu’il n’y ait plus de complète intelligibilité. Le concept d’invariant perd alors de sa force et de sa pertinence.
76b) Je pense de moins en moins que l’action du temps et la non-séparabilité soient deux problèmes directement liés l’un à l’autre. Mais je pense qu’il y a une sorte de couplage qui s’effectue avec le vivant.
77Imaginons en effet un système au sujet duquel nous avons une information complète, ou au sujet duquel il est imaginable que nous puissions avoir une information complète. Nous pouvons estimer ainsi par exemple qu’il est possible de dresser la liste de toutes les machines particulières dont le fonctionnement est susceptible d’être simulé par une machine universelle, pour reprendre l’exemple célèbre de Turing.
78Mais simplifions encore le modèle. Revenons aux automates reproducteurs de Von Neumann. On voit clairement apparaître un accroissement de complexité dû à une « récursivité » bizarre d’un système à l’intérieur duquel une description complète de celui-ci est fournie aux machines qui le composent pour effectuer un calcul. Nous voyons qu’ici la description des propriétés initiales et des objets qui composent la machine est complète. Pourtant on voit comment le modèle de cette machine s’oppose aux hypothèses d’Henri Atlan (1986). Atlan cherchait à montrer en effet que ce qui semble être une perte de « redondance » et une augmentation de « complexité » au niveau de l’observateur (apparition d’une fonction nouvelle) était en fait une réduction de l’espace des possibles au niveau de la voie du calcul, ce qui correspondait au fait que les automates aléatoires ne parcouraient pas l’ensemble de leurs configurations possibles. Mais ici, du fait que la description complète du système est à présent une information intégrée et fournie à certains éléments du système lui-même, l’espace des possibles va se transformer. Paradoxalement, il ne diminue pas. Ce n’est donc plus le même système, ou encore le même point de vue du système à l’instant To et à l’instant T1. Mais il faut concevoir pour cela la voie du calcul comme autre chose qu’une voie « déductive ».
79Voilà à présent pourquoi je soutiens une version forte de l’émergence diachronique. Elle a un pouvoir causal, parce que le temps agit sur le système, dès que le système est en situation d’agir sur lui-même. Autrement dit, le point de vue que je défends ne consiste pas à faire une mythologie de l’action du temps. Mais je pense que le temps agit, et qu’il agit dès que nous avons affaire à des systèmes en situation d’agir sur eux-mêmes, des systèmes où une information concernant l’ensemble du système est susceptible d’être rendue fonctionnelle à l’intérieur de ce système lui-même. Autrement dit, ce qu’il faut réfuter ici, si l’on refuse cet argument, ce n’est pas que le temps agisse, mais plutôt qu’il y ait en biologie des systèmes susceptible d’agir sur eux-mêmes. Cette réfutation nous semble mal partie, dès que nous évoquons par exemple les hyper-cycles et les structures de catalyse croisée. On voit en effet qu’avec de telles structures qui démarrent de l’auto-catalyse, le système en fonctionnant ajoute des propriétés systémiques à la liste initiale de ses propriétés. Il est à la fois capable de se reproduire et de se compliquer. D’où une définition de l’émergence faible, qui n’a plus de rapport direct avec le point de vue de l’observateur et le point de vue du système. On peut toujours dire néanmoins que cette différence temporelle n’a de sens que du point de vue de quelqu’un ou de quelque chose qui l’éprouve, il n’en reste pas moins qu’elle met en scène un système qui agit sur lui-même et qui n’est donc jamais rien. C’est un système sans essence et sans substance, un « événement de système » :
80(7) L’action du système sur lui-même en T0 ajoute des propriétés au système en T1.
81c) On voit d’autre part, qu’un tel système pourrait très bien n’être qu’un système formel. Le modèle de Von Neumann met en scène des automates reproducteurs, on pourrait sans doute montrer qu’ils ne font que se comporter de la même manière que des automates calculateurs. On voit aussi que c’est à ce type d’itération ou de récursivité que nous avons affaire, dans les modèles de simulation discrets dès que la fonction initialement caractérisée devient ensuite argument pour elle-même. Il n’y a aucun cercle vicieux là-dedans, pas plus que dans le modèle de Von Neumann, mais d’une certaine manière on met en scène un système initial qui agit sur lui-même et qui se complique au cours du déroulement temporel du calcul. Le problème ne vient donc pas d’un défaut dans le déterminisme ou dans le formalisme. Au contraire, d’une certaine manière, plutôt que de conclure à l’insuffisance des systèmes formels, nous pourrions alors en induire que les systèmes formels se comportent un peu comme les systèmes naturels. Ce sont des systèmes qui sont définis par le fait que leur action ne se résume pas à leur description. D’une certaine manière, ils vérifient complètement l’adage de Bergson et de Whitehead, selon lequel un système formel ne se contente pas de représenter le réel, mais qu’il agit lui-même réellement. Une manière de le dire consiste à signaler qu’il agit dans le temps.
§ 17. Émergence forte
82Par conséquent pour nous, même l’émergence faible a une valeur ontologique. Nous distinguerons cependant les émergences forte et faible. La seconde est sans aucun doute le produit de ce couplage entre non-séparabilité et la propriété qu’a un système d’agir sur lui-même. Dans le modèle de l’émergence faible, de nouvelles propriétés apparaissent, mais sans remettre en cause la syntaxe fondamentale du système, ce que l’on pourrait appeler sa « structure causale ». En termes philosophiques, nous pourrions dire que le principe de complétude est violé, mais pas le principe de causalité. La manière dont le système agit sur lui-même peut se superposer à l’action de ses éléments sans en modifier les propriétés fondamentales. Tel est évidemment le cas pour les systèmes dissipatifs que nous avons évoqués. Ils ont une trajectoire de phase qui ne peut pas se recouper en un point. C’est une idée au fond très simple qui ne correspond à rien d’autre que le fait que nous pouvons jouer aux échecs sans avoir à modifier les pièces. Ce qui est valable pour ces systèmes dissipatifs est aussi largement valable pour la modélisation et la simulation informatique ou numérique actuelles.
83Nous pensons que les systèmes vivants sont plus complexes. Ce que traduit l’exemple de la protéine RecA, c’est un processus à travers lequel un système complexe d’objets agit sur lui-même en modifiant la structure causale initiale qui le caractérise. Si nous nous plaçons en effet au niveau des gènes (individu), ainsi qu’à celui des dynamiques de populations de gènes (espèce), enfin au niveau des interactions extra-spécifiques (milieu), nous voyons que nous avons là l’exemple d’un processus complexe en un nouveau sens.
84Le « couplage déterminisme-stochasticité », permet déjà de comprendre (d’une façon très différente de celle évoquée par Von Neumann), comment un système dont les éléments de base sont changés de façon aléatoire peut engendrer de la complexité. On peut même imaginer qu’avec de nouvelles équations plus sophistiquées, il puisse intégrer des effets d’interaction liés par exemple à l’action de l’espace sur des populations de gènes, ou encore à plusieurs formes distinctes d’interactions (entre allèles, entre gènes, entre phénotypes et milieu, Ao, 2005). Mais cette modélisation ne suffit pas à prendre en compte ce qui se produit dans le modèle de Radman :
85(8) Les éléments de base qui composent le système sont transformés par la dynamique du système.
86Voilà ce que j’appellerai l’émergence forte. Il est à noter que ce que je viens d’en dire ne s’oppose pas aux descriptions engagées par Kim dans ses derniers articles. Le concept que je viens de décrire ici n’est en effet rien d’autre que ce qu’il nomme lui-même « la causalité descendante réflexive diachronique » (2005, 69). Il remarque avec raison que l’on peut alors penser une causalité descendante comme celle que l’on vient de décrire sans aucun effet de circularité, en raison du délai temporel introduit « entre la cause putative et l’effet ».
Bibliographie
Bibliographie
Achim, Stephan, « Varieties of Emergentism », Evolution and Cognition, Vol. 5, N°1, 49-59, 1999.
Achim, Stephan, « Emergentism, irreducibility, and downward causation », Grazer Philosophische Studien, 65, 77-93, 2002.
Ameisen, Jean-Claude, La Sculpture du vivant, Paris, Seuil, 2003.
Ao, Ping, « Laws in Darwinian Evolutionary Theory », Physics of life Reviews, 2, 117-156, 2005.
Atlan, Henri, et al., « Emergence of Classification Procedures in Automata Networks as a Model of Self Organization », Journal of Biological Theory, 120, 371-80, 1986.
Atlan, Henri, « Le gène programme ou données ? », in Théories de la complexité, Paris, Seuil, 1991.
Atlan, Henri, « Le rôle de la sophistication », in L’intentionnalité en question, Paris, Vrin, 1995.
Atlan, Henri, Koppel, Moshe, « Self-Organization and Computability », La Nuova Critica, 30/2, 5-17, 1997.
Atlan, Henri, La fin du tout génétique, Paris, Inra éd., 1999.
Barabasi, Albert-Laszlo, Albert, Reka, « Emergence of scaling in random networks », Science, 286, 509-512, 1999.
Barbaras, Renaud, Vie et intentionnalité, Paris, Vrin, 2003.
Barwise, Jon, Moss, Lawrence, Vicious Circles, On the Mathematics of Non Wellfounded Phenomena, USA, CSLI publications, 1996.
Bedeau, Mark A., « Weak emergence », Philosophical perspectives of mind causation and world, 375-99, 1997.
Bergson, Henri, L’évolution créatrice, Paris, PUF, 2008 [1907].
Bricmont, Jean, « Science of Chaos or Chaos in Science ? » in P.R. Gross, N. Levitt and M.W. Lewis (eds.), The Flight from Science and Reason, Ann. of the New York Academy of Sciences, 775, 131-175, New York Academy of Sciences, New York, 1996. [Also appeared in Physicalia Magazine, 17 (1995), 159-208].
Brotcorne-Lannoye, A., Maenhaut-Michel, G. « Role of RecA protein in untargeted UV mutagenesis of bacteriophage lambda : evidence for the requirement for the dinB gene », Proc. Natl. Acad. Sci. USA, Vol. 83, 3904-3908, 1986.
Burdon, R.H., « Control of Cell Proliferation by Reactive Oxygen Species », Biochemical Society Transactions, Vol. 24, 1028-1031, 1996.
Cairns-Smith, A.G., Genetic takeover and the mineral origin of life, Cambridge University Press, 1982.
Church, Alonso, « A Note on the Entscheidungsproblem », Journal of Symbolic Logic, 1, 40-41, 1936.
Darwin, Charles, On the origin of species by means of natural selection, London, John Murray, 1859 ; tr. fr. 6e éd., Paris, Reinwald, 1882.
Darwin, Charles, On the various contrivances by which British and foreign orchids are fertilized by insects, I & II, London, John Murray, 1862.
Darwin, Charles, The Variation of Animals and Plants under Domestication, J. Murray, London, 1868, 2 vol. La variation des animaux et des plantes sous l’effet de la domestication, trad. fr., Paris, Reinwald, 1868.
Dennett, Daniel, Darwin’s Dangerous Idea, NY, Touchstone ed., 1996.
Duboule, D. « The evolution of bricolage », TIG, Vol. 14, N°2, 57-67, 1998.
Fagot-Largeault, Anne, Andler, Daniel, Saint Sernin, Bertrand, Philosophie des sciences, Tome 2, Paris, Gallimard, Folio, 2002.
Eigen, M., Schuster, P., « The hypercycle. A principle of natural self-organization. Part A : Emergence of the hypercycle », Naturwissenschaften, 64 (November 1977), 541-565.
Eigen, M., Schuster, P., The Hypercycle : A Principle of Natural Self-Organization, Springer-Verlag, New York, 1979.
Falconer, D.S., Introduction to Quantitative Genetics, N.Y., Ronald Press, 1960.
Fisher, Ronald, The Genetical Theory of Natural Selection, Oxford, Clarendon Press, 1930.
Fox Keller, Evelyn, The Century of the Gene, Harvard University Press, 2000.
Fox Keller, Evelyn, Making Sense of Life, Harvard University Press, 2003.
Hénon, Michel, « A Two-dimensional Mapping with a strange attractor », Communications in Mathematical Physics, 50, 69-77, 1976.
Hesse, Mary B., Models and Analogies in Science, Notre Dame Un. Pr., 1966.
Hofstadter, Douglas, Gödel, Escher, Bach, N.Y., Basic Books, 1979.
Holton, Gerald, L’imagination scientifique, tr. fr. Paris, Gallimard, 1982.
Hume, David, An Inquiry Concerning Human Understanding, Oxford philosophical texts, Paperback, 1999.
Husserl, Edmund, Die Idee der Phänomenologie, La Haye, Martinus Nijhoff, 1973 [1907].
Jacob, François, La logique du vivant, Paris, Gallimard, 1970.
Jacquard, Albert, Concepts en génétique des populations, Masson, Paris, 1977.
Junien, Claudine, « L’empreinte parentale : de la guerre des sexes à la solidarité entre générations », Médecine Science, n°3, Vol. 16, 341-45, mars 2000.
Karlin, Stephane, « Two locus selection models », Theor. Populations Biology, 7, 364-98, 1977.
Kauffman, Stuart, « Metabolic stability and epigenesis in randomly constructed genetics nets », Journal of Biological Theory, 22, 437-67, 1969.
Kauffman, Stuart, « The large scale structure and dynamics of gene control circuits », Journal of Biological Theory, 44, 167-190, 1974.
Kauffman, Stuart, At Home in the Universe, Oxford Un. Pr., 1995.
Kauffman, Stuart, Investigations, Oxford Un. Pr., 2000.
Kauffman, Stuart, « Simulation of a Chemical Autonomous Agent », Zeitschrift für Physikalisches Chemie, 216, 1-9, 2002.
Kim, Jaegwon, Supervenience and Mind, Cambridge Un. Pr., 1993.
Kim, Jaegwon, Philosophy of Mind, Westview Pr., 1998.
Kim, Jaegwon, « Making sens of Emergence », Philosophical Studies 95, 3-36, 1999.
Laforge, B., Guez, D., Martinez, M. et Kupiec, J.J., « Modeling embryogenesis and cancer : an approach based on an equilibrium between the autostabilization of stochastic gene expression and the interdependence of cells for proliferation », Progress in Biophysics and Molecular Biology, Vol. 89 (1), 93-120, 2005.
Lesne, A., Victor, J.M., « Chromatin Fiber Functional Organisation : a Plausible Model », Eur. Phys. J., E 19, 279-290, 2006.
Levin, S.A., « Complex adaptative systems : exploring the known and the unknown and the unknowable », Bull. Am. Soc., 40 (1), 3-19, 2003.
Lloyd Morgan, C., Emergent Evolution, Henry Holt and Co., 1923.
Longo, Giuseppe, « Mathematical intelligence, infinity and machines : Beyond the Gödelitis », Journal of Consciousness Studies, special issue on Cognition, Vol. 6, 11-12, 1999.
Longo, Giuseppe, « The causal incompleteness of Programming Theory in Molecular Biology », Noesis, 14, 165-201, décembre 2008.
Love, Alan C. « Evolvability, Dispositions and Intrinsicality », Philosophy of Science, 70, 1015-1027, December 2003.
Lowe, Edward J., « Lois et dispositions » in La Structure du monde, éd. J.M. Monnoyer, Vrin, 2004.
Luigi Luisi, Pier, The Emergence of Life : From Chemical Origins to Synthetic Biology, Cambridge, Cambridge University Press, 2006.
Malaterre, Christophe, « Les origines de la vie : schèmes explicatifs et structure théorique », Noesis, 14, 89-113, décembre 2008.
Maurel, Marie-Christine, La Naissance de la vie, Paris, Dunod, 2003.
Maynard-Smith, John, Les origines de la vie, tr. fr. Paris, Masson Sciences, Dunod, 2000.
Mayr, Ernst, « Where are we ? », Cold Spring Harbor Symposia on Quantitative Biology, 24, 58-64, 1959.
Mayr, Ernst, « Cause and Effect in Biology », Nature, 134, 1501-1505, November 1961.
Miquel, Paul-Antoine, « Considérations épistémologiques sur les cercles vicieux épigénétiques du vieillissement », Journal de la Société de Biologie, 198(3), 2004.
Miquel, Paul-Antoine, « Complexité, sophistication, émergence en biologie », in Autour de Henri Atlan, éd. P. Bourgine, Paris, La découverte, 159-174, 2008.
Miquel, P.A., Soto, A., Sonnenschein, C., « Physicalism and downward causation in developmental and cancer biology », Acta Biotheoretica, 56, N° 4, 257-274, décembre 2008.
Monod, Jacques, Le hasard et la nécessité, Paris, Seuil, 1970.
Morange, Michel, La part des gènes, Paris, O. Jacob, 1998.
Morange, Michel, Histoire de la biologie moléculaire, Paris, La découverte, 2003.
Morange, Michel, La vie expliquée ?, Paris, O. Jacob, 2003.
Noble, Denis, The Music of Life. Biology Beyond the Genome, Oxford University Press, 2006.
Perovic, S., « The limitation of Kim’s reductive physicalism », Acta Biotheoretica, 55, 243-267, 2007.
Prigogine, Ilia, Stengers, Isabelle, La Nouvelle alliance, Paris, Gallimard, 1979.
Prigogine, Ilia, Nicolis, Grégoire, Exploring Complexity : An Introduction, KG Verlag, München, 1989.
Putnam, Hilary, Realism with a Human Face, Harvard Un. Pr., 1990.
Quine, W.V., Ontological Relativity and Other Essays, N.Y., Columbia Press, 1969.
Radman, Miroslav, « Enzymes of Evolutionary Change », Nature, 401, 866-69, 1999a.
Radman, Miroslav, et al., « Evolution of Evolvability », N.Y. Ac. Of Sc., 870, 146-155, 1999b.
Radman, Miroslav, « Entretien avec Miroslav Radman », in Marie-Christine Maurel et Paul-Antoine Miquel, Nouveaux débats sur le vivant, Paris, Kimé, 106-115, 2003.
Raup, David, Extinction : Bad Genes or Bad Luck ?, NY, WW Norton, 1991.
Rayssiguier, Charles, Thaler, D., Radman, Miroslav, « The barrier to recombination between E coli and Salomnella typhimurium is disrupted in mismatch-repair mutants », Nature, Vol. 342, 396-400, 23 nov. 1989.
Rivera, Maria, Lake, J.A., « The ring of life provides evidence for a genome fusion of eukariotes », Nature, 431, 152-55, September 2004.
Rosenberg, Alex, Kaplan, D.M., « How to reconcile Physicalism and Antireductionism about Biology », Philosophy of Science, 72, 43-68, January 2005.
Schrödinger, Erwin, What is life ?, Paperback, Cambridge University Press, 1992 [1944].
Sober, Elliott, The Nature of Selection : Evolutionary Theory in Philosophical Focus, Cambridge MA, MIT Press, 1984.
Soto, Ana, Sonnenschein, Carlos, The Society of Cells, Oxford, Bios Scientific Publishers, 1999.
Soto, Ana, Sonnenschein, Carlos, « Emergentism as a default : Cancer as a
problem of tissue organization », J. Biosci., 30, 103-118, 2004.
Spencer, Herbert, First Principles, London, William and Norgate, 1862.
Stengers, Isabelle, Cosmopolitiques VI : La vie et l’artifice : visages de l’émergence, Paris, La découverte, 1997.
Von Kiedrovski, Günter, Sievers, D., « Self-replication of complementary nucleotide-based oligomers », Nature, 369, 221-24, 1994.
Von Neumann, J., Théorie générale et logique des automates [1953], tr. fr. J.P. Auffrand, Paris, Champs Vallon, 1996.
Wang, Y., Wysocka, J., Sayegh, J., et al. « Human PAD4 regulates histone arginine
methylation levels via demethylimination », Science, 306, 279-83, 2004.
Watson, Jim, Crick, Francis, « General Implications of the Structure of Deoxyribonucleic Acid », Nature, 171, 964-67, may 1953.
Wright, Sewall, « The Roles of Mutation, Inbreeding, Crossbreeding, and Selection in Evolution », in D.F. Jones (ed.) Proceedings of the Sixth International Congress of Genetics, 356-366, Menasha, 1932.
Wright, Sewall, Evolution and the Genetics of Population, T3, Un. Chicago Press, 1977.