
Les papillons de jour (Lepidoptera, Papilionoidea) et les orthoptères (Orthoptera) des bassins ardoisiers du Parc national de la Vallée de la Semois : bilan des inventaires réalisés entre 2023 et 2025
Résumé
Ce travail présente le bilan des inventaires des papillons de jour (Rhopalocères) et des Orthoptères réalisés dans les bassins ardoisiers du Parc national de la Vallée de la Semois. Entre 2023 et 2025, 57 sites ardoisiers répartis dans les régions d’Alle – Rochehaut, Fays-les-Veneurs et Herbeumont – Bertrix ont été visités. Au total, 41 espèces distinctes de papillons de jour ont été observées au sein de 30 sites et 19 espèces d’orthoptères ont été détectées au sein de 26 sites. Parmi ces 60 espèces, 9 espèces patrimoniales ont été identifiées, dont 2 espèces d’orthoptères (Calliptamus italicus et Oedipoda caerulescens) et 7 espèces de Rhopalocères (Callophrys rubi, Fabriciana adippe, Favonius quercus, Pyrgus malvae, Satyrium ilicis, Speyeria aglaja et Thymelicus lineola). Les résultats sont notamment discutés à la lumière du premier inventaire général des sites ardoisiers publié il y a près de 20 ans. Les trois sites présentant actuellement le plus grand intérêt lépidoptérique et/ou orthoptérique sont situés dans la vallée d’Aise (Herbeumont) : Goutelle Husson, Prigeai et Petit Babinay.
Abstract
This work presents the results of butterfly and orthopteran inventories carried out in the slate mining regions of the National Park of the Semois Valley. Between 2023 and 2025, 57 slate quarries distributed across the Alle – Rochehaut, Fays-les-Veneurs, and Herbeumont – Bertrix regions were visited. In total 41 distinct butterfly species were observed within 30 sites, and 19 Orthoptera species were detected within 26 sites. Among these 60 species, 9 heritage species have been identified including 2 Orthoptera species (Calliptamus italicus and Oedipoda caerulescens) and 7 butterfly species (Callophrys rubi, Fabriciana adippe, Favonius quercus, Pyrgus malvae, Satyrium ilicis, Speyeria aglaja, and Thymelicus lineola). The results are discussed, in particular in light of the first general inventory of slate quarries published almost 20 years ago. The three sites currently presenting the greatest interest regarding butterflies and/or orthopterans are located in the Aise valley (Herbeumont): Goutelle Husson, Prigeai, and Petit Babinay.
Article reçu le 29 janvier 2026, accepté le 26 mai et mis en ligne le 26 juin 2026.
Cet article est distribué suivant les termes et les conditions de la licence CC-BY (http://creativecommons.org/licenses/by/4.0/deed.fr)
INTRODUCTION
1L’exploitation des matières premières minérales est une activité ancienne qui s’est développée dans chacune des régions naturelles de Wallonie. Actuellement, les carrières wallonnes exploitent essentiellement les roches carbonatées qui sont à la base de l’activité de l’industrie extractive régionale (Gosselin, 2006 ; Castiau & Godart, 2017). En dehors de ces sites actifs, plus de 5000 carrières abandonnées ont été répertoriées par Remacle (2009), dont environ 30 % situées en Ardenne. C’est notamment dans cette région que les phyllades et schistes ardoisiers ont été massivement exploités et que la production industrielle d’ardoises s’est nettement développée à partir du 19ème siècle. Au 20ème siècle, le déclin multifactoriel de l’industrie ardoisière wallonne a conduit à la fermeture de la quasi-totalité des ardoisières (Remacle, 2007).
2Les anciennes carrières jouent un rôle majeur en termes de maintien de la biodiversité car elles abritent et voient se développer des milieux et des taxons à forte valeur patrimoniale (biotopes rares, habitats d’intérêt communautaire et taxons rares, menacés et/ou protégés) (Remacle, 2006). En Wallonie, dans les années 1990 et 2000, l’inventaire des anciennes carrières a été réalisé dans le cadre des conventions « Les sablières en Région wallonne » et « Les carrières en Région wallonne ». Cet inventaire s’inscrivait dans l’inventaire plus large des Sites de Grand Intérêt biologique (SGIB), dont le but était de recenser les espaces (semi-)naturels remarquables (Bisteau et al., 2009). Avec notamment le massif de Philippeville, les bassins ardoisiers ardennais abritent une concentration de carrières reconnues de grand intérêt biologique (Remacle, 2009). À cet égard, l’intérêt biologique des parties en surface des sites ardoisiers a fait l’objet d’un travail publié par Remacle (2007). Ces sites se répartissent dans huit bassins (régions) ardoisiers : L’Escaillère – Cul-des-Sarts, Oignies, Alle – Rochehaut, Fays-les-Veneurs, Herbeumont – Bertrix, Neufchâteau, Martelange et Vielsalm. Dans son travail, Remacle en décrit l’intérêt faunistique et floristique de manière détaillée, en se concentrant sur certains groupes ciblés dans le cadre de la convention « Les Carrières en Région wallonne », notamment les papillons de jour et les orthoptères recensés en raison de leur qualité de bioindicateurs (Remacle, 2009). Dans les milieux ouverts, ces deux groupes sont utiles en tant qu’indicateurs écologiques ou pour le suivi des effets des mesures de gestion ou des travaux de restauration (Gerlach et al., 2013).
3Dans le cadre du Plan de Relance de la Wallonie et plus particulièrement de son projet 98 dédié à la création de deux parcs nationaux, les Parcs nationaux de l’Entre-Sambre-et-Meuse (PNESEM) et de la Vallée de la Semois (PNVS) ont été reconnus en décembre 2022. Au sein du premier Plan opérationnel du PNVS, la fiche-action n° 13 (FA 13) « Restauration et préservation des sites ardoisiers » est mise en œuvre par l’association Ardenne & Gaume. Elle s’appuie sur le travail de Remacle (2007) et se concentre sur les bassins ardoisiers d’Alle – Rochehaut, de Fays-les-Veneurs et d’Herbeumont – Bertrix, totalisant 68 sites (18 étant situés totalement ou partiellement en dehors du périmètre du PNVS). Certains sites ardoisiers abritent des biotopes remarquables tels que des éboulis (WalEUNIS H2.3) et des pelouses sèches (WalEUNIS E1) qui constituent des habitats pour de nombreuses espèces animales et végétales patrimoniales qui nécessitent une conservation. C’est bien l’enjeu de l’action n° 13 qui vise :
-
la mise à jour des connaissances biologiques résultant du travail de Remacle (2007) ;
-
la restauration et la préservation de la biodiversité particulière des sites ardoisiers dans au moins trois sites ;
-
la protection réglementaire des sites restaurés.
4Dans ce contexte, cet article présente les résultats des inventaires des papillons de jour (Lepidoptera, Papilionoidea) et des Orthoptères (Orthoptera) réalisés durant la période 2023-2025.
MATÉRIEL ET MÉTHODES
Sites étudiés
5Les sites concernés par la présente étude sont les 68 sites ardoisiers (Figure 1 ; Tableau 1) renseignés et décrits par Remacle (2007) dans les bassins d’Alle – Rochehaut (C.1 à C.23), Fays-les-Veneurs (D.1 à D.15) et Herbeumont – Bertrix (E.1 à E.30). Aucune donnée n’a été récoltée dans 11 sites, et ce pour diverses raisons (propriété privée, autorisation d’accès non accordée, site inaccessible, etc.). Au total, des inventaires ont donc été réalisés dans 57 sites et des données ont été récoltées entre mai 2023 et septembre 2025. Les périmètres des sites prospectés sont basés sur ceux présentés par Remacle (2007), avec d’éventuelles adaptations selon la réalité de terrain. La superficie des 68 sites varie de 0,001 à 6,2 hectares. Notons qu’entre 2007 et 2023, parmi les sites inventoriés, les sites C.2, C.15, C.18, E.10, E.16, E.18 et E.20 ont fait l’objet d’un déboisement notable, partiel ou total.
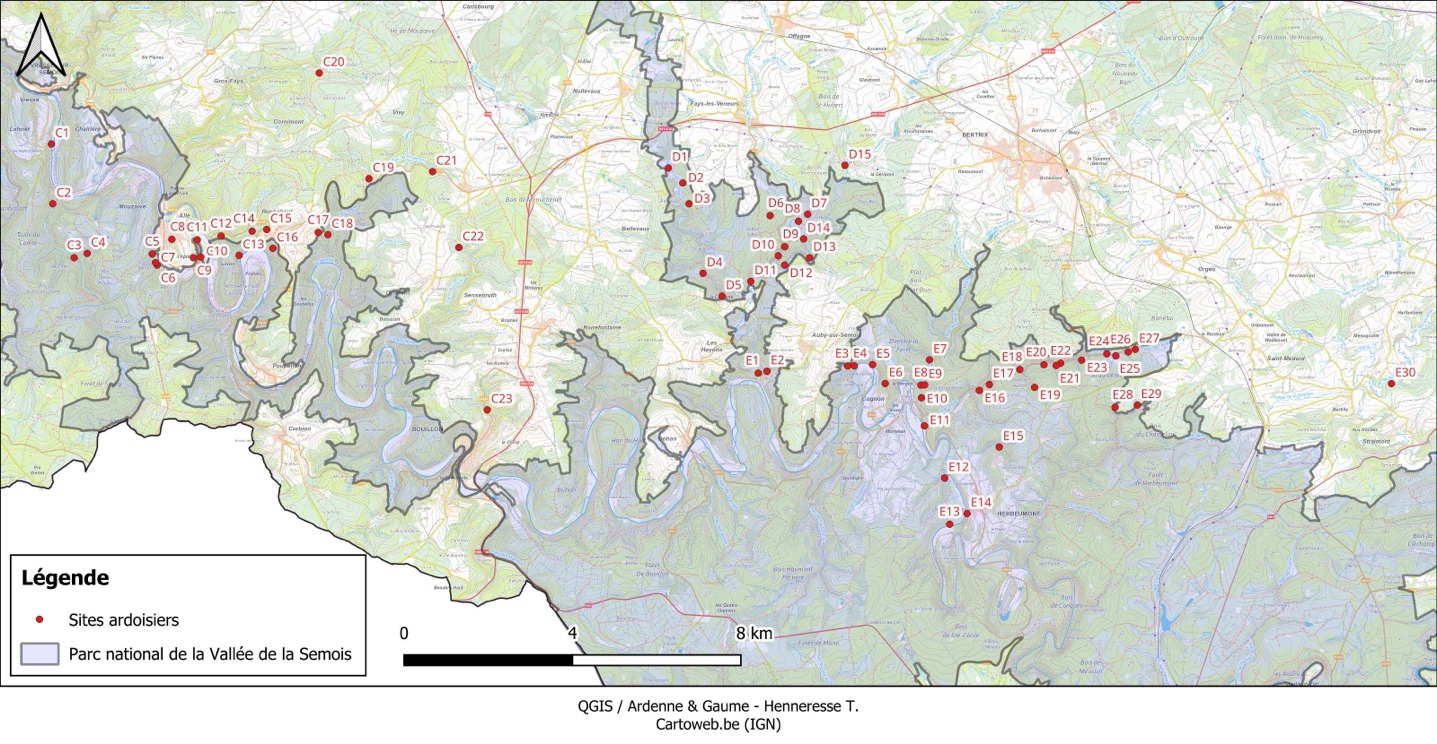
Figure 1 : Localisation des sites ardoisiers étudiés.
Tableau 1 : Sites concernés par la FA 13 du Plan opérationnel 2023-2026 du PNVS. Sauf mention contraire indiquée par un astérisque, les informations présentées sont tirées de la contribution de Remacle (2007).
|
Bassin ardoisier |
Site |
N° |
Commune(s) |
Superficie (ha) |
X (LB72) |
Y (LB72) |
Site inventorié (FA 13) |
|
Alle – Rochehaut |
Cavité le long de la Semois à Laforêt |
C.1 |
Vresse-sur-Semois |
0,01 |
190530 |
61190 |
Oui |
|
Alle – Rochehaut |
Ardoisière de Laforêt |
C.2 |
Vresse-sur-Semois |
0,4* |
190550 |
59740 |
Oui |
|
Alle – Rochehaut |
Recherche ardoisière de la Virée des Malheurs |
C.3 |
Vresse-sur-Semois |
0,015 |
191063 |
58459 |
Oui |
|
Alle – Rochehaut |
Trou Hoffman |
C.4 |
Vresse-sur-Semois |
0,005 |
191380 |
58580 |
Oui |
|
Alle – Rochehaut |
Ardoisière du Reposseau |
C.5 |
Vresse-sur-Semois |
0,85 (excavation exclue) |
192890 |
58580 |
Oui |
|
Alle – Rochehaut |
Recherche ardoisière de Nancrette |
C.6 |
Vresse-sur-Semois |
0,05 |
193040 |
58320 |
Oui |
|
Alle – Rochehaut |
Recherches ardoisières secondaires de Nancrette |
C.7 |
Vresse-sur-Semois |
0,2* |
193000 |
58400 |
Oui |
|
Alle – Rochehaut |
Trou des Moines |
C.8 |
Vresse-sur-Semois |
0,1* |
193390 |
58915 |
Non |
|
Alle – Rochehaut |
Ardoisière Laplet (siège occidental) |
C.9 |
Vresse-sur-Semois |
> 1,5 |
193950 |
58480 |
Non |
|
Alle – Rochehaut |
Ardoisière Laplet (siège oriental) |
C.10 |
Vresse-sur-Semois |
1,5* |
194080 |
58500 |
Oui |
|
Alle – Rochehaut |
Ardoisière Sainte-Barbe |
C.11 |
Vresse-sur-Semois |
1 |
193960 |
58890 |
Oui |
|
Alle – Rochehaut |
Ardoisière de Hoûr |
C.12 |
Bouillon |
0,6 |
194570 |
58980 |
Non |
|
Alle – Rochehaut |
Ardoisière Laviot |
C.13 |
Bouillon |
1,3* |
195000 |
58500 |
Oui |
|
Alle – Rochehaut |
Ardoisière du Moulin de Bochet |
C.14 |
Bouillon |
0,15 |
195280 |
59070 |
Oui |
|
Alle – Rochehaut |
Ardoisière La Faligeotte |
C.15 |
Bouillon |
0,3* |
195650 |
59140 |
Oui |
|
Alle – Rochehaut |
Ardoisière Notre-Dame |
C.16 |
Bouillon |
0,15 |
195790 |
58710 |
Oui |
|
Alle – Rochehaut |
Recherche ardoisière voisine de Gérardfosse |
C.17 |
Bouillon |
< 0,1 |
196870 |
59080 |
Non |
|
Alle – Rochehaut |
Ardoisière Gérardfosse |
C.18 |
Bouillon |
0,45* |
197080 |
59050 |
Oui |
|
Alle – Rochehaut |
Ardoisière de Lîresse |
C.19 |
Bouillon |
0,2 |
198060 |
60360 |
Oui |
|
Alle – Rochehaut |
Trou de Tailfer |
C.20 |
Bièvre |
> 1 |
196920 |
62865 |
Oui |
|
Alle – Rochehaut |
Recherche ardoisière de Mogimont |
C.21 |
Bouillon |
0,06 |
199590 |
60525 |
Oui |
|
Alle – Rochehaut |
Trou du Voile |
C.22 |
Bouillon |
0,05* |
200185 |
58730 |
Oui |
|
Alle – Rochehaut |
Trou de la Mère Pelée |
C.23 |
Bouillon |
0,002 |
200895 |
54840 |
Oui |
|
Fays-les-Veneurs |
Ardoisière de Pont le Prêtre |
D.1 |
Paliseul |
0,02* |
205180 |
60600 |
Oui |
|
Fays-les-Veneurs |
Ardoisière des Boules de Launoy |
D.2 |
Bouillon* & Paliseul |
0,3* |
205545* |
60260* |
Oui |
|
Fays-les-Veneurs |
Ardoisière du Péreu |
D.3 |
Bouillon & Paliseul |
1,6* |
205610 |
59770 |
Oui |
|
Fays-les-Veneurs |
Scaïre du Billard |
D.4 |
Bouillon |
0,3* |
206010 |
58080 |
Oui |
|
Fays-les-Veneurs |
Ardoisière des Badats |
D.5 |
Bouillon |
0,1 |
206450 |
57580 |
Oui |
|
Fays-les-Veneurs |
Site du Bois à Ban |
D.6 |
Paliseul |
0,8* |
207600* |
59480* |
Oui |
|
Fays-les-Veneurs |
Ardoisière de Rougebeau |
D.7 |
Paliseul |
1,7* |
208490 |
59500 |
Oui |
|
Fays-les-Veneurs |
Petite Ardoisière de Rougebeau |
D.8 |
Paliseul |
0,1* |
208275* |
59345* |
Oui |
|
Fays-les-Veneurs |
Site de la Côte de Châtillon (?) |
D.9 |
Paliseul |
0,02* |
207970 |
58750 |
Oui |
|
Fays-les-Veneurs |
Ardoisière des Laid Joué |
D.10 |
Paliseul |
0,15* |
207785 |
58520 |
Oui |
|
Fays-les-Veneurs |
Ardoisière de la Cornette |
D.11 |
Paliseul |
0,2* |
207135 |
57920 |
Oui |
|
Fays-les-Veneurs |
Ardoisière Sainte-Adèle |
D.12 |
Bertrix* & Paliseul |
0,2 |
207950 |
58310 |
Oui |
|
Fays-les-Veneurs |
Ardoisière des Séhan |
D.13 |
Bertrix |
0,8* |
208550 |
58470 |
Oui |
|
Fays-les-Veneurs |
Ardoisière de la Côte de Châtillon |
D.14 |
Bertrix & Paliseul |
1,3* |
208390 |
58910 |
Oui |
|
Fays-les-Veneurs |
Ardoisière de la Géripont |
D.15 |
Bertrix |
0,5 |
209380 |
60660 |
Non |
|
Herbeumont – Bertrix |
Trou du Maka des Aleines |
E.1 |
Bouillon |
? |
207265 - 207345 |
55665 - 55755 |
Non |
|
Herbeumont – Bertrix |
Recherche ardoisière au lieu-dit Dessous le Village |
E.2 |
Bertrix |
0,04* |
207540 |
55780 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière du Pé (ouest) |
E.3 |
Bertrix |
0,15* |
209445 |
55880 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière du Pé |
E.4 |
Bertrix |
< 0,02 |
209600 |
55910 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière du Pé (est) |
E.5 |
Bertrix |
< 0,01 |
210035 |
55935 |
Non |
|
Herbeumont – Bertrix |
Ardoisière Latour (?) |
E.6 |
Bertrix |
0,06* |
210335* |
55485* |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière de la Goutelle de Derrière la Forêt |
E.7 |
Bertrix |
0,03 |
211395 |
56040 |
Oui |
|
Herbeumont – Bertrix |
Cavité du Cul du Mont |
E.8 |
Bertrix |
< 0,01 |
211185 |
55440 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière du Cul du Mont |
E.9 |
Bertrix |
0,07* |
211280 |
55450 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière de Linglé |
E.10 |
Bertrix |
5,8* |
211150 |
55150 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière La Fortelle |
E.11 |
Herbeumont |
0,9* |
211275 |
54475 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière du Bois Petite Dansau |
E.12 |
Bertrix |
0,05 |
211745 |
53240 |
Non |
|
Herbeumont – Bertrix |
Recherche ardoisière du Bois Grande Dansau |
E.13 |
Bertrix |
0,02 |
211865 |
52140 |
Non |
|
Herbeumont – Bertrix |
Cavité au pied de la Forteresse |
E.14 |
Herbeumont |
< 0,01 |
212270 |
52390 |
Non |
|
Herbeumont – Bertrix |
Recherche ardoisière des Burzais |
E.15 |
Herbeumont |
0,05* |
213035 |
53955 |
Oui |
|
Herbeumont – Bertrix |
Ardoisières de Wilbauroche |
E.16 |
Bertrix & Herbeumont |
2,9* |
212550 |
55330 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière de la Goutelle Husson |
E.17 |
Herbeumont |
1,2* |
212810 |
55470 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière de la Maljoyeuse |
E.18 |
Bertrix |
1,7* |
213535 |
55815 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière des Collard |
E.19 |
Herbeumont |
1,4* |
213880 |
55400 |
Oui |
|
Herbeumont – Bertrix |
Ardoisières des Anciennes Carrières et du Prigeai |
E.20 |
Bertrix & Herbeumont |
6,2* |
214100 |
55930 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière à l'est du Prigeai |
E.21 |
Herbeumont |
0,02 |
214405 |
55900 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière à l'est du Prigeai |
E.22 |
Herbeumont |
0,015 |
214500 |
55965 |
Non |
|
Herbeumont – Bertrix |
Ardoisière de la Morépire |
E.23 |
Bertrix |
3,8* |
215000 |
56050 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière du Grand Babinay |
E.24 |
Bertrix |
0,04* |
215590 |
56190 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière du Grand Babinay |
E.25 |
Bertrix & Herbeumont |
2,9 |
215800 |
56120 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière La Renaissance |
E.26 |
Herbeumont |
0,06* |
216115 |
56235 |
Oui |
|
Herbeumont – Bertrix |
Ardoisière du Petit Babinay |
E.27 |
Bertrix & Herbeumont |
2,4* |
216280 |
56290 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière de Bochaban |
E.28 |
Herbeumont |
0,04* |
215800 |
54915 |
Oui |
|
Herbeumont – Bertrix |
Recherche ardoisière de Bochaban (est) |
E.29 |
Herbeumont |
0,08* |
216315 |
54980 |
Oui |
|
Herbeumont – Bertrix |
Trou le long du ruisseau de Neufchâteau à Martilly |
E.30 |
Herbeumont |
0,001 |
222350 |
55480 |
Oui |
Récolte des données
6Les contributions de Sevilleja et al. (2019), Lacoeuilhe et al. (2020), Freitas et al. (2021), Sperber et al. (2021) et Barkmann et al. (2023) ont été consultées pour élaborer les protocoles d’inventaire des papillons de jour et des orthoptères.
7Les prospections ciblant les papillons de jour ont été effectuées entre le 1er mai et le 31 août et celles ciblant les orthoptères entre le 1er juillet et le 30 septembre. Elles ont été réalisées entre 10h00 et 17h00, en l’absence de précipitations ou de vents trop forts (vitesse supérieure à 38 km/h) et à des températures supérieures à 13 °C. Entre 13 et 17 °C, la couverture nuageuse maximale était de 25 %.
8Dans les 16 sites pourvus de milieux (semi-)ouverts propices à la présence d’un cortège diversifié de papillons de jour (Tableau 2 ; Figure 2) ou d’orthoptères (Tableau 3 ; Figure 3), ces deux groupes ont été recherchés activement selon les protocoles décrits ci-après. Le caractère « propice » des sites a été évalué subjectivement par l’auteur selon différents critères, essentiellement l’abondance des fleurs nectarifères, le degré de boisement et la superficie des milieux ouverts.
9Le suivi minimal consistait à réaliser, lors d’une seule des 3 années d’étude, 2 (orthoptères) ou 3 passages (papillons de jour) séparés d’au moins 4 semaines et idéalement réalisés à des moments différents de la journée au fil des passages. Bien qu’habituellement utilisé comme méthode d’inventaire des papillons de jour, le transect n’a pas été retenu dans le cadre de la présente étude. Au lieu de recourir à un parcours échantillon fixe, les papillons et les orthoptères ont été recensés en parcourant, dans la mesure du possible, tous les milieux ouverts d’un site à vitesse réduite. D’après Barkmann et al. (2023), cette méthode est davantage adaptée pour évaluer la richesse et la composition spécifiques des papillons de jour, ce que visait en premier lieu l’étude. De plus, le transect se prêtait peu à la structure des sites, fortement hétérogènes en termes de types de végétation et souvent de taille réduite.
10La durée d’inventaire n'a été en aucun cas inférieure à 10 min par passage et par groupe. Les imagos de papillons de jour ont été identifiés à l’œil nu, à l’aide d’une paire de jumelles macro ou après capture au filet à papillons. Pour chaque taxon, le nombre maximal d’imagos observés simultanément et les plantes vasculaires éventuellement visitées ont été notés. La détection et l’identification des orthoptères reposait essentiellement sur l’écoute des chants audibles. Ponctuellement, la diffusion du chant de certaines espèces, initialement non détectées mais dont la présence était possible, a été opérée afin de faciliter leur détection. L’observation à l’œil nu ou à l’aide d’une paire de jumelles macro et la capture des individus au filet à papillons ou au filet fauchoir suivie d’un examen visuel ont permis de compléter les informations sur les espèces présentes. Concernant les orthoptères arboricoles, l’inventaire par battage des branches d’arbres et d’arbustes et récolte des individus qui en chutaient à l’aide d’un parapluie japonais a initialement été pratiqué mais a été abandonné à partir de 2024 vu le très faible nombre d’insectes capturés.
11De manière générale, en dehors du cadre de cet effort d’échantillonnage minimal (lors de passages additionnels – ne ciblant pas les papillons de jour ou les orthoptères – réalisés entre 2023 et 2025 et éventuellement en dehors de la période de prospection), les chenilles et imagos détectés ont également été notés. Le nombre total de passages réalisés par site est indiqué aux Tableaux 2 et 3.




Figure 2 : Milieux (semi-)ouverts propices à la présence d’un cortège diversifié de papillons de jour (© Thomas Henneresse).
(a) Ardoisière de la Goutelle Husson (E.17 – mai 2025). (b) Ardoisière du Petit Babinay (E.27 – juillet 2025). (c) Ardoisières des Anciennes Carrières et du Prigeai (E.20 – juillet 2023). (d) Ardoisière de la Maljoyeuse (E.18 – juin 2024).
Tableau 2 : Nombre de passages dans les sites où les papillons de jour ont été recherchés activement. Les chiffres romains indiquent le nombre de passages réalisés selon le protocole défini et les chiffres arabes indiquent le nombre de passages additionnels ne respectant pas le protocole mais lors desquels au moins une observation a été réalisée.
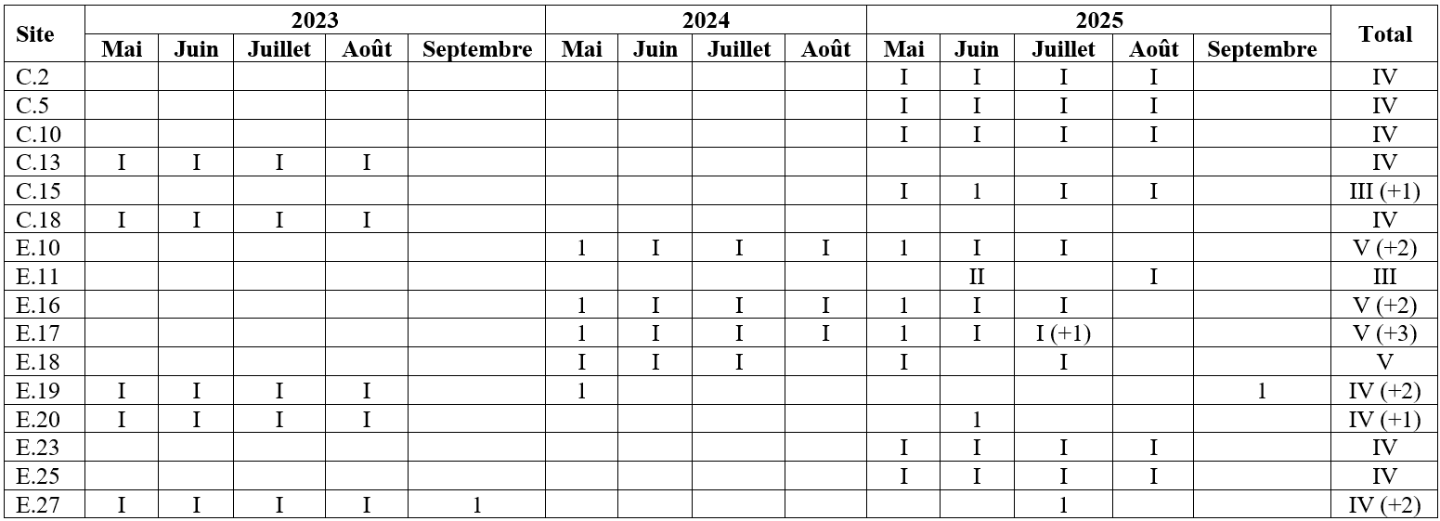
Tableau 3 : Nombre de passages dans les sites où les orthoptères ont été recherchés activement. Les chiffres romains indiquent le nombre de passages réalisés selon le protocole défini et les chiffres arabes indiquent le nombre de passages additionnels ne respectant pas le protocole mais lors desquels au moins une observation a été réalisée.
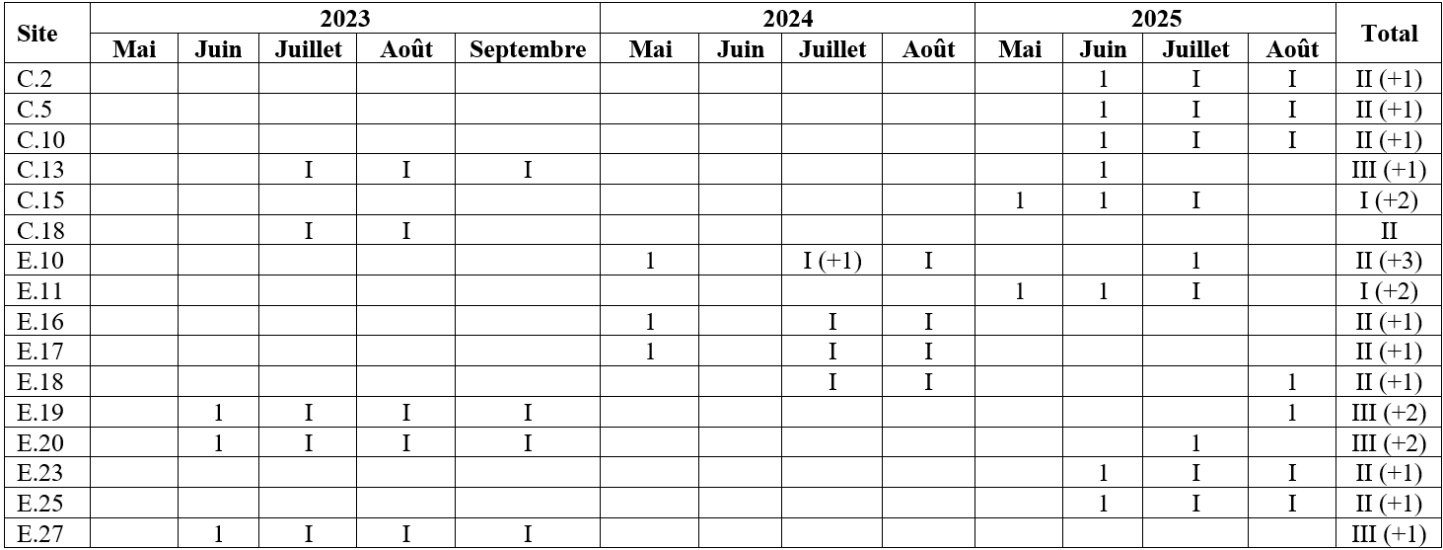




Figure 3 : Milieux (semi-)ouverts propices à la présence d’un cortège diversifié d’orthoptères (© Thomas Henneresse).
(a) Ardoisière La Faligeotte (C.15 – mai 2025). (b) Ardoisières de Wilbauroche (E.16 – mai 2024). (c) Ardoisière Laplet (siège oriental) (C.10 – juin 2025). (d) Ardoisière de Laforêt (C.2 – mai 2025).
12Les sites dépourvus ou très faiblement pourvus de milieux (semi-)ouverts propices à la présence d’un cortège diversifié de papillons de jour ou d’orthoptères ont fait l’objet d’au moins un passage (en pratique, généralement un seul) lors duquel étaient ciblés la flore vasculaire et l’herpétofaune ; les papillons de jour et les orthoptères n’y ont pas été recherchés activement mais les individus détectés ont été identifiés. Ces sites n’ont pas été inclus dans l’analyse des données par ordination et classification (voir ci-après).
13Dans site donné, dans le but de se focaliser sur un seul groupe à la fois, les papillons de jour et les orthoptères n’étaient pas étudiés simultanément mais il arrivait ponctuellement qu’une observation d’un orthoptère soit réalisée durant l’inventaire des papillons, et vice versa. En outre, les inventaires réalisés ne consistant pas en un suivi biologique mais visant plutôt à dresser une liste d’espèces, une espèce détectée dans un site lors d’un passage n’a généralement pas été renotée lors des passages suivants.
14Dans le cadre de cette étude, une dérogation aux mesures de protection des espèces a été demandée et obtenue pour chacune des trois années d’inventaires, avec autorisation de perturber, capturer et détenir temporairement (individus directement relâchés après identification), sous certaines conditions, différentes espèces animales protégées par la Loi du 12 juillet 1973 sur la Conservation de la Nature.
15Toutes les données présentées dans la section « Résultats » ont été récoltées par l’auteur et encodées sur le site web Observations.be. La localisation de certains taxons, qualifiés de patrimoniaux, a été obscurcie en raison de leur rareté en Ardenne belge, de leur statut de conservation défavorable et/ou de leur protection en Wallonie.
Analyse des données
16Toutes les analyses ont été réalisées à l’aide de la version 4.5.1 du logiciel R (R Core Team, 2025) et des packages ade4 (Dray et al., 2025), cluster (Maechler et al., 2025), indicspecies (De Cáceres & Legendre, 2009) et vegan (Oksanen et al., 2025). Les données utilisées sont de type présence/absence et sont issues du cumul de tous les relevés réalisés dans un site donné.
17Afin de réaliser le positionnement multidimensionnel non-métrique (non-metric multidimensional scaling – NMDS), une analyse d’ordination permettant la réduction des données et la visualisation graphique des distances entre sites (facilitant ainsi l’interprétation des résultats), les données des taxons non identifiés jusqu’à l’espèce ont tout d’abord été écartées. Ensuite, seuls les 16 sites pourvus de milieux (semi-)ouverts propices à la présence d’un cortège diversifié de papillons de jour ou d’orthoptères (voir ci-dessus) ont été considérés, à l’exception des sites C.10 (papillons de jour et orthoptères) et C.18 (orthoptères uniquement) abritant un nombre anormalement faible de taxons et donc considérés comme des outliers. L’analyse en coordonnées principales (PCoA) constituait une alternative valable au NMDS pour analyser les données. Le choix de cette deuxième méthode est lié au fait que l’accent était mis sur le rang des distances entre les points (cas du NMDS – Bakker, 2024) plutôt que sur leurs valeurs absolues (cas de la PCoA) et que l’interprétation des axes (et leur contribution à la variance totale) n’était pas visée.
18Pour les deux groupes biologiques, à partir de la matrice binaire (présence/absence), l’indice de dissimilarité de Jaccard a été calculé (fonction vegdist) pour chaque paire de sites afin de générer une matrice de dissimilarité sur laquelle portait le NMDS (fonction metaMDS, 2 dimensions). En outre, partant de la matrice de dissimilarités, une classification ascendante hiérarchique (CAH) des sites a été réalisée (fonction hclust, méthode d’agrégation de Ward « ward.D » choisie car elle minimise l’inertie intra-classe et tend à créer des groupes bien séparés et uniformes) et le dendrogramme généré (non représenté ici) a été découpé (fonction cutree) en deux groupes sur base du coefficient de silhouette (fonction silhouette), ce coefficient donnant une indication sur la qualité de la partition. Enfin, une analyse des espèces indicatrices a été réalisée (fonction multipatt, indice « IndVal.g », nombre de permutations fixé à 9999) afin de déterminer si certaines étaient indicatrices d’un des deux groupes et donc de caractériser davantage ceux-ci en termes écologiques.
Référentiels
19La nomenclature scientifique des plantes vasculaires est conforme à Verloove & Van Rossum (2024). La nomenclature scientifique et la classification des autres groupes biologiques sont conformes aux informations présentées sur le site web Observations.be.
20Les espèces de papillons de jour et d’orthoptères reprises dans la liste établie par Desmet et al. (2025) sont considérées comme exotiques en Wallonie, en particulier Cacyreus marshalli (Butler 1898), Locusta migratoria (Linnaeus 1758), Meconema meridionale (Costa 1860) et Pieris mannii (Mayer 1851). Bien que n’y figurant pas, Acanthacris ruficornis (Fabricius 1787) est également considéré comme exotique (Claerebout, 2012).
21En considérant les évaluations les plus récentes, le statut wallon de conservation a été déterminé par Fichefet et al. (2008) pour les papillons de jour et par Lock et al. (2011) pour les orthoptères.
22Sur base de la contribution de Dupont (2015), le degré de spécialisation, la propension à la dispersion à l’échelle d’un écocomplexe et l’habitat optimal (pour la chenille en France) ont été attribués à chaque espèce de papillon, en sélectionnant les conditions suivantes (correspondant au mieux au contexte des sites étudiés) : région Champagne-Ardenne, département des Ardennes et altitude comprise entre 200 et 400 m. Le degré de spécialisation (sténoécie) se décline en 4 listes (Dupont, 2015) :
-
Liste 1 : « espèces généralistes dont les chenilles se développent dans de nombreux types d’habitat » ;
-
Liste 2 : « espèces moyennement généralistes dont les chenilles se développent principalement dans l’habitat associé. L’espèce peut se maintenir au niveau de l’habitat même dans le cas où ce dernier subit une dégradation. Ces espèces sont généralement communes » ;
-
Liste 3 : « espèces spécialistes dont les chenilles se développent majoritairement dans l’habitat associé. Ces espèces ont généralement une répartition étroitement liée à la répartition de l’habitat. Le bon état de conservation de l’habitat est un facteur clef pour la dynamique des populations de l’espèce » ;
-
Liste 4 : « espèces spécialistes ayant une répartition très localisée dans le département. Cette répartition peut être liée à une spécialisation importante de l’espèce vis à vis d’une composante de l’habitat et/ou à une adaptation chorologique moindre de l’espèce dans le département concerné ».
23La propension à la dispersion se décline en trois niveaux (Dupont, 2015) :
-
Dispersion faible : réalisation de la majorité des déplacements dans la parcelle ou une parcelle adjacente ;
-
Dispersion moyenne : réalisation de la majorité des déplacements à l'échelle de l'écocomplexe d'habitats ;
-
Dispersion forte : « les déplacements des individus permettent la visite de plusieurs écocomplexes avec des habitats favorables ».
24Sur base de la contribution de Logghe et al. (2025b), la capacité de dispersion a été attribuée à chaque espèce d’orthoptères ; elle varie entre 0,1 et 1 et exprime le potentiel d’une espèce à se disperser / à se déplacer à une certaine distance de son lieu de naissance (Logghe et al., 2025a). Le caractère spécialiste (espèces xérophiles ou hygrophiles) ou généraliste (espèces mésophiles) des orthoptères a été attribué selon Reinhardt et al. (2005). L’appartenance à un groupe écologique a également été indiquée et déterminée sur base des communautés mentionnées par Claerebout (2013), celles-ci étant éventuellement modifiées suivant un avis d’expert (Devriese H., comm. pers.).
RÉSULTATS
25Papillons de jour (Rhopalocères)
26Les sites ont fait l’objet de trois à cinq passages selon le protocole défini, avec éventuellement un à trois passages additionnels ne respectant pas le protocole (Tableau 2). Concernant les sites E.10 et E.16 à E.18 visités une première fois en 2024, deux passages supplémentaires selon le protocole ont été réalisés en 2025 afin de compenser l’abondance et la diversité anormalement faibles qui avaient été constatées. En 2024, en Wallonie et dans des régions voisines, les conditions météorologiques du printemps et du début de l’été ont en effet affecté très négativement les espèces printanières de papillons de jour et celles développant des générations printanières et estivales, menant à une forte diminution des populations (https://papillons.natagora.be/resultats-de-loperation).
27Au total, 334 données ont été encodées sur Observations.be ; elles représentaient en majorité des occurrences « taxon × site » uniques (peu d’encodage de taxons plus d’une fois par site). Un total de 42 taxons (40 indigènes identifiés jusqu’à l’espèce + Leptidea sp. + Pieris sp.) appartenant à 5 familles a été répertorié dans 30 des 57 sites inventoriés (Tableau 4). La famille, le statut de conservation wallon, le degré de spécialisation, la propension à la dispersion, et l’habitat optimal des espèces sont présentés au Tableau 5. Les 30 sites abritaient entre 1 et 29 taxons (Figure 4). En tenant uniquement compte des 16 sites où les papillons ont été cherchés activement, entre 4 et 29 taxons ont été observés (Figure 5), avec en moyenne 13,5 (± 6,5) taxons par site. Notons que le nombre de passages réalisés selon le protocole (Tableau 2) par site n’est pas corrélé significativement au nombre de taxons observés (coefficient de corrélation de Spearman ρ = 0,26, p-valeur = 0,34). Les sites présentant une diversité supérieure à 20 taxons étaient le Petit Babinay (E.27) et le Prigeai (E.20) avec respectivement 29 et 26 taxons. Le bassin d’Herbeumont – Bertrix était le plus riche avec 41 taxons, contre 17 dans celui d’Alle – Rochehaut et 8 dans celui de Fays-les-Veneurs.
Tableau 4 : Papillons de jour recensés dans les sites étudiés (Source : données personnelles, collectées à l’aide d’Observations.be), avec le nombre maximal d’imagos (sauf pour P. machaon en E.23 et P. c-album en E.25 où il s’agit de chenilles) observés simultanément (les estimations sont mises en évidence en italique). NSO = nombre de sites occupés.
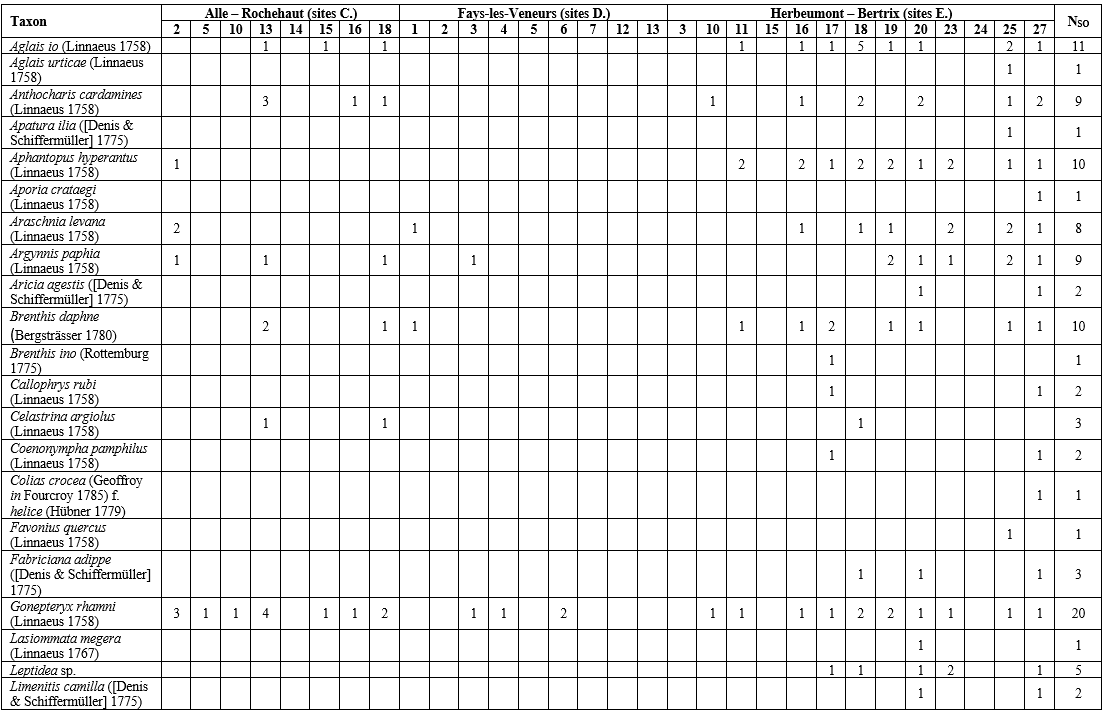
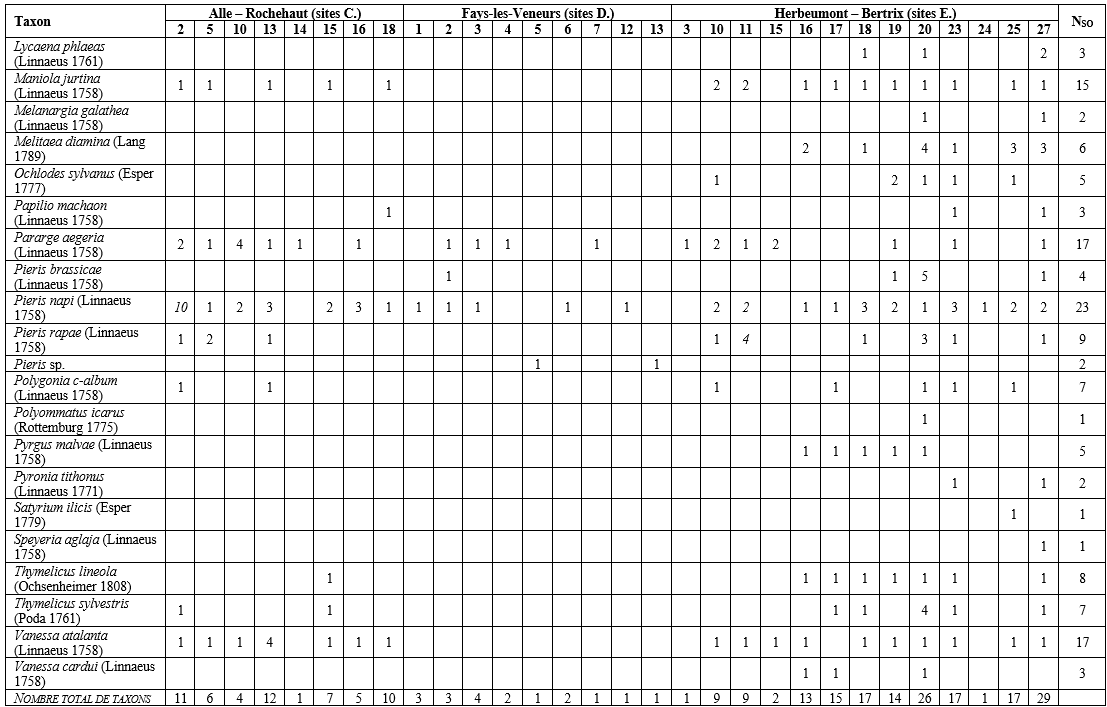
Tableau 5 : Papillons de jour : famille, statut de conservation wallon, degré de spécialisation, propension à la dispersion, habitat optimal et taxons végétaux visités pour leurs fleurs dans les sites ardoisiers étudiés. Disp. = dispersion ; Spéc. = spécialisation ; Stat. cons. = statut de conservation.
|
Taxon |
Famille |
Stat. cons. |
Spéc. |
Disp. |
Habitat optimal |
Taxon(s) végétal(aux) visité(s) dans les sites ardoisiers |
|
Aglais io |
Nymphalidae |
LC |
Liste 1 |
Forte |
Lisières et prairies humides ou mouilleuses à grandes herbes et à fougères |
Buddleja davidii Franch. (E.25), Centaurea gr. jacea (E.17), Cirsium vulgare (Savi) Ten. (E.18), Eupatorium cannabinum L. (C.13), Origanum vulgare L. (E.20), Rubus sp. (E.25), Senecio ovatus (G.Gaertn., B.Mey & Scherb.) Willd. (C.18) & Trifolium pratense L. (C.15) |
|
Aglais urticae |
Nymphalidae |
LC |
Liste 1 |
Forte |
Lisières et prairies humides ou mouilleuses à grandes herbes et à fougères |
Hieracium lachenalii C.C.Gmel. (E.25) |
|
Anthocharis cardamines |
Pieridae |
LC |
Liste 2 |
Moyenne |
Lisières et prairies humides ou mouilleuses à grandes herbes et à fougères |
Hesperis matronalis L. (E.16) |
|
Apatura ilia |
Nymphalidae |
LC |
Liste 2 |
Moyenne |
Forêts riveraines et forêts galeries, avec dominance de Populus, d'Alnus ou de Salix |
/ |
|
Aphantopus hyperantus |
Nymphalidae |
LC |
Liste 2 |
Moyenne |
Ourlets forestiers thermophiles |
/ |
|
Aporia crataegi |
Pieridae |
LC |
Liste 3 |
Moyenne |
Fourrés tempérés |
/ |
|
Araschnia levana |
Nymphalidae |
LC |
Liste 2 |
Moyenne |
Lisières et prairies humides ou mouilleuses à grandes herbes et à fougères |
Cirsium arvense (L.) Scop. (E.25), Eupatorium cannabinum (E.23), Euphorbia amygdaloides Lam. (E.18), Heracleum sphondylium L. (E.23 & E.25), Rubus sp. (C.2 & E.25), Sonchus arvensis L. (E.18 & E.23), Torilis japonica (Houtt.) DC. (C.2) & Valeriana officinalis L. (E.25) |
|
Argynnis paphia |
Nymphalidae |
LC |
Liste 2 |
Moyenne |
Forêts mésophiles ou eutrophiles avec Quercus, Carpinus, Fraxinus, Acer, Tilia ou Ulmus |
Eupatorium cannabinum (E.23), Origanum vulgare (E.20) & Rubus sp. (E.19 & E.25) |
|
Aricia agestis |
Lycaenidae |
LC |
Liste 1 |
Moyenne |
Friches, jachères ou terres arables récemment abandonnées |
/ |
|
Brenthis daphne |
Nymphalidae |
NE |
Liste 1 |
Forte |
Fourrés tempérés |
Centaurea gr. jacea (E.27), Leontodon hispidus L. (E.16), Origanum vulgare (E.20) & Rubus sp. (D.1, E.11, E.16, E.19 & E.25) |
|
Brenthis ino |
Nymphalidae |
LC |
Liste 3 |
Moyenne |
Lisières et prairies humides ou mouilleuses à grandes herbes et à fougères |
Centaurea gr. jacea (E.17) |
|
Callophrys rubi |
Lycaenidae |
NT |
Liste 1 |
Faible |
Fourrés tempérés |
Leucanthemum ircutianum DC. (E.27) & Polygala vulgaris L. (E.17) |
|
Celastrina argiolus |
Lycaenidae |
LC |
Liste 1 |
Moyenne |
Fourrés tempérés |
/ |
|
Coenonympha pamphilus |
Nymphalidae |
LC |
Liste 1 |
Forte |
Prairies de fauche de basse et moyenne altitude |
/ |
|
Colias crocea f. helice |
Pieridae |
LC |
/ |
/ |
/ |
/ |
|
Favonius quercus |
Lycaenidae |
NT |
Liste 2 |
Faible |
Forêts mésophiles ou eutrophiles avec Quercus, Carpinus, Fraxinus, Acer, Tilia ou Ulmus |
/ |
|
Fabriciana adippe |
Nymphalidae |
VU |
Liste 3 |
Moyenne |
Ourlets forestiers thermophiles |
Centaurea gr. jacea (E.20) & Cirsium vulgare (E.18) |
|
Gonepteryx rhamni |
Pieridae |
LC |
Liste 2 |
Forte |
Fourrés tempérés |
Buddleja davidii (E.25), Centaurea gr. jacea (E.16 & E.27), Cirsium vulgare (E.18), Dianthus armeria L. (E.20), Epilobium angustifolium L. (E.25), Eupatorium cannabinum (C.5), Galeopsis tetrahit L. (C.2 & C.18), Geranium robertianum L. (C.2), Leontodon hispidus (E.10), Lotus pedunculatus Cav. (C.5), Origanum vulgare (E.20), Phedimus spurius (M. Bieb.) ‘t Hart (C.5), Rubus sp. (C.13, D.6 & E.25), Scorzoneroides autumnalis (L.) Moench (C.5) & Silene dioica (L.) Clairv. (E.18) |
|
Lasiommata megera |
Nymphalidae |
LC |
Liste 3 |
Forte |
Pelouses calcicoles vivaces et steppes riches en bases |
/ |
|
Leptidea sp. |
Pieridae |
/ |
/ |
Moyenne |
/ |
Leucanthemum ircutianum (E.17) & Trifolium pratense (E.23) |
|
Limenitis camilla |
Nymphalidae |
LC |
Liste 3 |
Moyenne |
Fourrés tempérés |
/ |
|
Lycaena phlaeas |
Lycaenidae |
LC |
Liste 1 |
Forte |
Prairies de fauche de basse et moyenne altitude |
Eupatorium cannabinum (E.20) |
|
Maniola jurtina |
Nymphalidae |
LC |
Liste 1 |
Forte |
Prairies de fauche de basse et moyenne altitude |
Centaurea gr. jacea (E.16, E.17, E.18, E.23 & E.27), Eupatorium cannabinum (C.5), Galeopsis tetrahit (C.18), Origanum vulgare (E.20), Rubus sp. (C.13 & E.25) & Valeriana officinalis (E.10) |
|
Melanargia galathea |
Nymphalidae |
LC |
Liste 2 |
Moyenne |
Prairies de fauche de basse et moyenne altitude |
Centaurea gr. jacea (E.27) |
|
Melitaea diamina |
Nymphalidae |
LC |
Liste 4 |
Faible |
Prairies oligotrophes humides ou mouilleuses |
Pilosella officinarum Vaill. (E.16) & Rubus sp. (E.25) |
|
Ochlodes sylvanus |
Hesperidae |
LC |
Liste 1 |
Moyenne |
Lisières et prairies humides ou mouilleuses à grandes herbes et à fougères |
Origanum vulgare (E.20) |
|
Papilio machaon |
Papilionidae |
LC |
Liste 1 |
Moyenne |
Végétations herbacées rudérales |
Centaurea gr. jacea (E.27) & Echium vulgare L. (E.27) |
|
Pararge aegeria |
Nymphalidae |
LC |
Liste 1 |
Forte |
Ourlets forestiers thermophiles |
/ |
|
Pieris brassicae |
Pieridae |
LC |
Liste 1 |
Forte |
Friches, jachères ou terres arables récemment abandonnées |
Origanum vulgare (E.20) & Rubus sp. (D.2 & E.19) |
|
Pieris napi |
Pieridae |
LC |
Liste 1 |
Forte |
Prairies de fauche de basse et moyenne altitude |
Alliaria petiolata (M.Bieb) Cavara & Grande (C.2), Allium ursinum L. (C.16), Buddleja davidii (E.25), Cardamine amara L. (E.25), Centaurea gr. jacea (E.18), Euphorbia amygdaloides (E.17), Geranium columbinum L. (E.10), Geranium robertianum (C.2, C.10 & E.10), Hieracium laevigatum Willd. (E.23), Lapsana communis L. (C.2), Leontodon hispidus (C.5), Lythrum salicaria L. (E.16), Prunella vulgaris L. (C.2), Rubus sp. (C.10, E.10, E.11, E.25), Thymus pulegioides L. (E.17), Trifolium pratense (C.5), Valeriana officinalis (E.25) & Vinca major L. (C.10) |
|
Pieris rapae |
Pieridae |
LC |
Liste 1 |
Forte |
Friches, jachères ou terres arables récemment abandonnées |
Centaurea gr. jacea (E.23), Hieracium lachenalii (C.5), Leucanthemum ircutianum (C.5), Origanum vulgare (E.20), Rubus sp. (E.11) & Scorzoneroides autumnalis (C.5) |
|
Pieris sp. |
Pieridae |
/ |
/ |
Forte |
/ |
/ |
|
Polygonia c-album |
Nymphalidae |
LC |
Liste 2 |
Forte |
Fourrés tempérés |
/ |
|
Polyommatus icarus |
Lycaenidae |
LC |
Liste 1 |
Forte |
Prairies de fauche de basse et moyenne altitude |
/ |
|
Pyrgus malvae |
Hesperidae |
VU |
Liste 1 |
Faible |
Pelouses calcicoles vivaces et steppes riches en bases |
/ |
|
Pyronia tithonus |
Nymphalidae |
LC |
Liste 2 |
Moyenne |
Ourlets forestiers thermophiles |
/ |
|
Satyrium ilicis |
Lycaenidae |
CR |
Liste 3 |
Faible |
Forêts caducifoliées thermophiles |
Rubus sp. (E.25) |
|
Speyeria aglaja |
Nymphalidae |
EN |
Liste 3 |
Moyenne |
Ourlets forestiers thermophiles |
Centaurea gr. jacea (E.27) |
|
Thymelicus lineola |
Hesperidae |
NT |
/ |
/ |
/ |
Campanula rotundifolia L. (E.19) & Lotus corniculatus L. (E.23) |
|
Thymelicus sylvestris |
Hesperidae |
LC |
Liste 1 |
Moyenne |
Prairies de fauche de basse et moyenne altitude |
Centaurea gr. jacea (E.23) & Hypericum perforatum L. (E.20) |
|
Vanessa atalanta |
Nymphalidae |
NE |
Liste 1 |
Forte |
Végétations herbacées rudérales |
Centaurea gr. jacea (E.27) & Rubus sp. (C.13) |
|
Vanessa cardui |
Nymphalidae |
NE |
Liste 1 |
Forte |
Végétations herbacées rudérales |
Pilosella flagellaris (Willd.) Arv.-Touv. (E.20) & Pilosella officinarum (E.17) |
Figure 4 : Nombre total de taxons de papillons de jour et abondance cumulée des imagos par site.
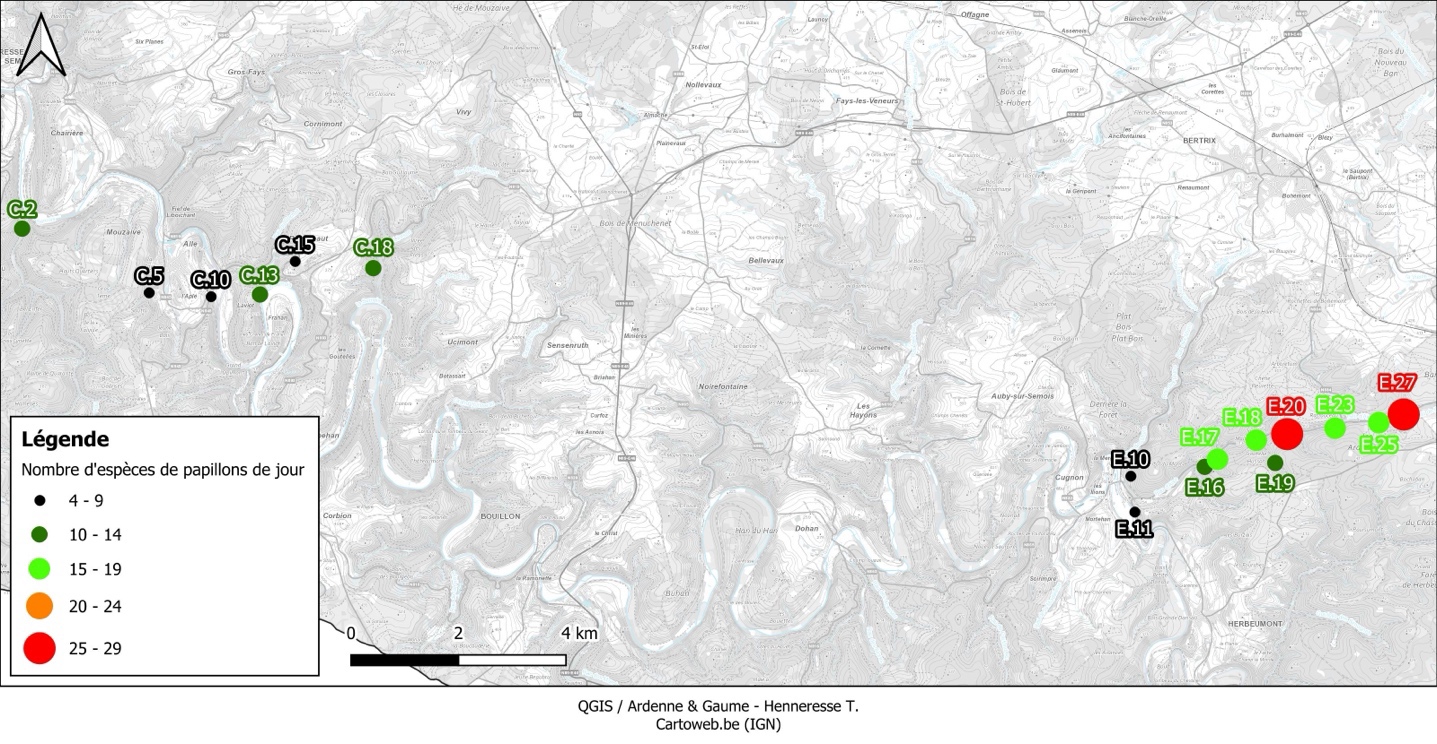
Figure 5 : Localisation des 16 sites où les papillons de jour ont été recherchés activement, avec le nombre d’espèces détectées (par catégorie).
28Concernant l’ordination par NMDS, la valeur de l’indicateur « stress » (Bakker, 2024) de 0,16 et le diagramme de Shepard (non représenté ici – ajustement non-métrique, R² = 0,973) indiquent une solution finale et une qualité d’ajustement correctes. Les deux groupes de sites formés à la suite de la CAH sont représentés sur le diagramme d’ordination (Figure 6) où ils se distinguent relativement bien l’un de l’autre des côtés gauche et droit du plan de l’ordination. Le groupe I comprend 8 sites (C.15, E.16, E.17, E.18, E.19, E.20, E.25 et E.27), quasiment tous situés dans le bassin d’Herbeumont – Bertrix et plus précisément dans la vallée du ruisseau d’Aise (Herbeumont) ; le groupe II inclut 7 sites (C.2, C.5, C.13, C.18, E.10, E.11, E.23) répartis entre les bassins d’Alle – Rochehaut et d’Herbeumont – Bertrix. Notons que la valeur moyenne du coefficient de silhouette s’élève à 0,15 en formant 2 groupes, contre 0,12 en formant 3 groupes et 0,14 en formant 4 groupes. Concernant les deux groupes formés, notons également que la valeur moyenne du coefficient de silhouette du groupe I s’élève à 0,09 contre 0,22 pour le groupe II. Dans le groupe I, aucune valeur du coefficient n’est supérieure à 0,2 et 3 sites (C.15, E.19 et E.25) possèdent une valeur proche de 0, ce qui indique qu’ils se situeraient sur ou à proximité directe de la limite de décision entre les deux clusters. Dans le groupe II, 2 sites (C.18 et E.23) possèdent une valeur du coefficient positive inférieure à 0,1, ce qui indique également qu’ils se situeraient sur ou à proximité directe de la limite de décision. Les espèces détectées dans les sites du groupe II représentaient strictement un sous-ensemble des espèces détectées dans les sites du groupe I (celui-ci accueillant l’ensemble des espèces détectées). L’analyse des espèces indicatrices a révélé qu’une seule espèce était significativement (p-valeur < 0,05) indicatrice du groupe II (Pararge aegeria (Linnaeus 1758), valeur indicatrice = 0,815) et que deux autres étaient significativement indicatrices du groupe I (Pyrgus malvae (Linnaeus 1758), val. ind. = 0,867 ; Thymelicus lineola (Ochsenheimer 1808), val. ind. = 0,791).
29Au total, 244 occurrences « taxon × site » (par exemple, Aglais io (Linnaeus 1758) en C.13) ont été notées (Tableau 4), 242 correspondant à l’observation, au même moment, d’un ou de plusieurs imagos et 2 correspondant à l’observation d’une chenille. Concernant les imagos, 187 des occurrences (77 %) correspondaient à l’observation d’un seul imago. Les 55 autres occurrences concernaient l’observation simultanée de 2 à 10 imagos. Les espèces les plus abondantes étaient Pieris napi (Linnaeus 1758) en C.2 (10 individus – estimation), Aglais io en E.18 (5 individus) et Pieris brassicae (Linnaeus 1758) en E.20 (5 individus). Pour 24 des 42 taxons, le nombre maximal d’imagos observés simultanément était un seul individu. Les sites présentant plus de 30 imagos (selon la somme des valeurs des occurrences « taxon × site ») étaient E.20 (39 individus toutes espèces confondues) et E.27 (34 individus) (Figure 4).
30Au total, 43 taxons de plantes vasculaires ont été visités pour leur nectar dans les sites étudiés (Tableau 5). De manière générale, les taxons attirant le plus grand nombre d’espèces distinctes de papillons de jour étaient Centaurea gr. jacea (13 espèces dans 6 sites), Rubus sp. (12 dans 11 sites), Origanum vulgare L. (8, toutes en E.20) et Eupatorium cannabinum L. (6 dans 4 sites), les autres en attirant entre 1 et 3. Ces mêmes taxons attiraient également le plus grand nombre de familles distinctes (4 pour C. gr. jacea et 3 pour les 3 autres taxons ; 1-2 pour les 39 autres taxons).
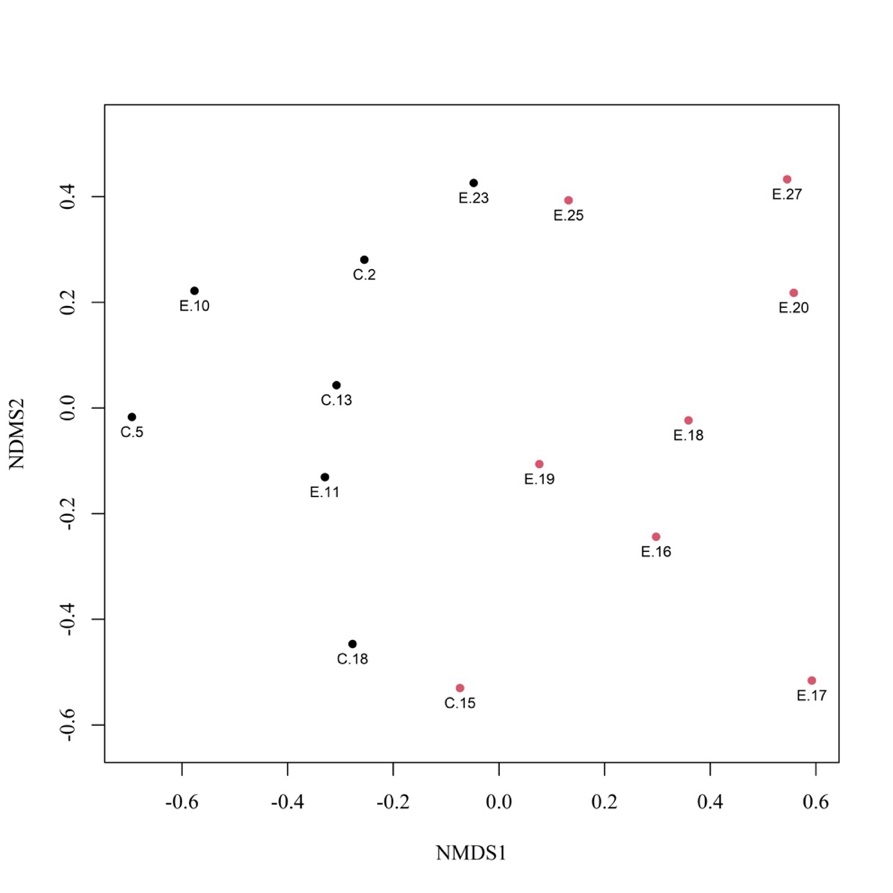
Figure 6 : Diagramme bidimensionnel d’ordination des sites réalisé par NMDS à partir des données de présence/absence des papillons de jour. Les 2 groupes formés à la suite de la CAH sont représentés en rouge et noir.
31Orthoptères
32Les sites ont fait l’objet d’un à trois passages selon le protocole défini, avec éventuellement un à trois passages additionnels ne respectant pas le protocole (Tableau 3). Concernant les sites C.15 et E.11, bien qu’un seul passage ait été réalisé en juillet selon le protocole défini, le passage additionnel de juin le complétait adéquatement en raison du printemps particulièrement chaud et sec responsable d’une précocité apparente des orthoptères en 2025.
33Au total, 148 données ont été encodées sur Observations.be ; elles représentaient en majorité des occurrences « taxon × site » uniques. Un total de 19 espèces (indigènes) appartenant à 4 familles a été répertorié dans 26 des 57 sites inventoriés (Tableau 6). La famille, le statut de conservation wallon, la capacité de dispersion, le caractère spécialiste/généraliste et le groupe écologique des espèces sont présentés au Tableau 7. Les 26 sites abritaient entre 1 et 13 espèces (Figure 7). En tenant uniquement compte des 16 sites où les orthoptères ont été cherchés activement, entre 3 et 13 espèces ont été observées (Figure 8), avec en moyenne 8,0 (± 3,1) espèces par site. Notons que le nombre de passages réalisés selon le protocole (Tableau 3) par site n’est pas corrélé significativement au nombre de taxons observés (coefficient de corrélation de Spearman ρ = 0,15, p-valeur = 0,58). Les sites présentant une diversité supérieure à 10 espèces étaient le Prigeai (E.20), La Faligeotte (C.15), la Goutelle Husson (E.17) et le Petit Babinay (E.27) avec respectivement 13, 12, 12 et 11 espèces. Le bassin d’Herbeumont – Bertrix était le plus riche avec 18 espèces, contre 13 dans celui d’Alle – Rochehaut et 6 dans celui de Fays-les-Veneurs.
Tableau 6 : Orthoptères recensés dans les sites étudiés (Source : données personnelles, collectées à l’aide d’Observations.be). NSO = nombre de sites occupés.
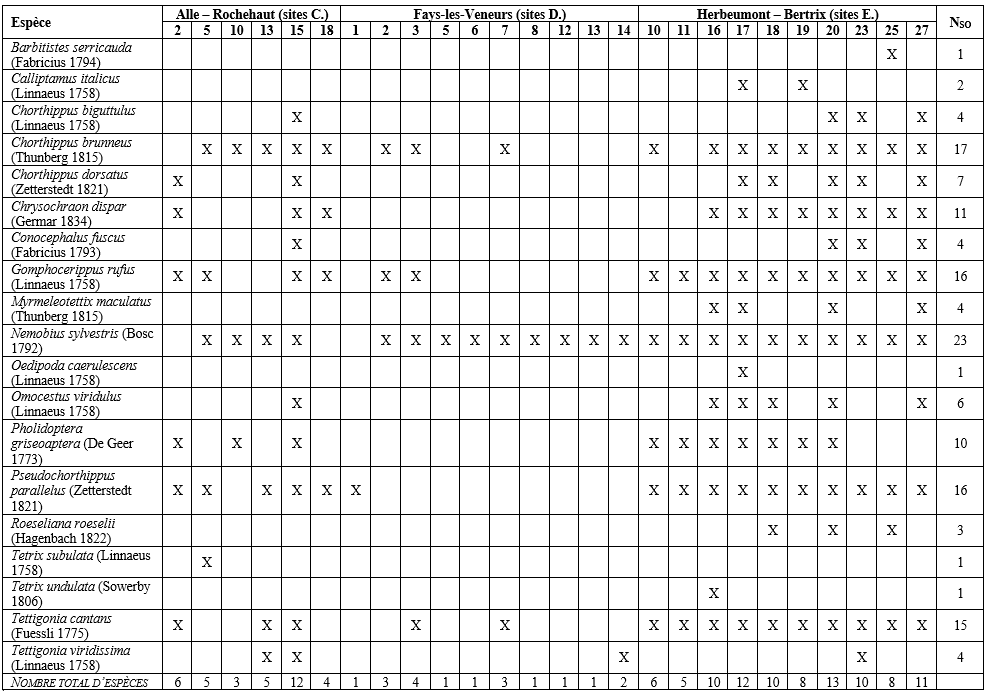
34Concernant l’ordination par NMDS, la valeur de l’indicateur « stress » de 0,10 et le diagramme de Shepard (non représenté ici – ajustement non-métrique, R² = 0,989) indiquent une solution finale et une qualité d’ajustement correctes. Les deux groupes de sites formés à la suite de la CAH sont représentés sur le diagramme d’ordination (Figure 9) où ils se distinguent bien l’un de l’autre des côtés gauche et droit du plan de l’ordination. Le groupe I comprend 7 sites (C.15, E.16, E.17, E.18, E.20, E.23 et E.27), quasiment tous situés dans la vallée du ruisseau d’Aise (Herbeumont) ; le groupe II inclut également 7 sites (C.2, C.5, C.13, E.10, E.11, E.19 et E.25) répartis entre les bassins d’Alle – Rochehaut et d’Herbeumont – Bertrix. Notons que la valeur moyenne du coefficient de silhouette s’élève à 0,23 en formant 2 groupes, contre 0,23 en formant 3 groupes (avec une valeur moyenne de - 0,01 pour un des 3 groupes) et 0,20 en formant 4 groupes. Concernant les deux groupes formés, notons également que la valeur moyenne du coefficient de silhouette du groupe I s’élève à 0,38 contre 0,08 pour le groupe II. Dans le groupe I, aucune valeur du coefficient n’est inférieure à 0,2. Dans le groupe II, 2 sites (E.19 et E.25) possèdent une valeur du coefficient proche de 0, ce qui indique qu’ils se situeraient sur ou à proximité directe de la limite de décision entre les deux clusters. Toujours dans le groupe II, le site C.2 possède une valeur de - 0,12, indiquant une légère tendance à être classé erronément. L’analyse des espèces indicatrices a révélé que deux espèces étaient significativement (p-valeur < 0,05) indicatrices du groupe I (Omocestus viridulus (Linnaeus 1758), valeur indicatrice = 0,926 ; Chorthippus dorsatus (Zetterstedt 1821), val. ind. = 0,857) et qu’aucune n’était significativement indicatrice du groupe II.
Tableau 7 : Orthoptères : famille, statut de conservation wallon, capacité de dispersion, caractère spécialiste (« Oui ») ou généraliste (« Non ») de l’espèce et groupe écologique. Concernant la capacité de dispersion, notons que Chrysochraon dispar et Roeseliana roeselii présentent un dimorphisme alaire avec des individus macroptères (Kleukers et al., 2004).
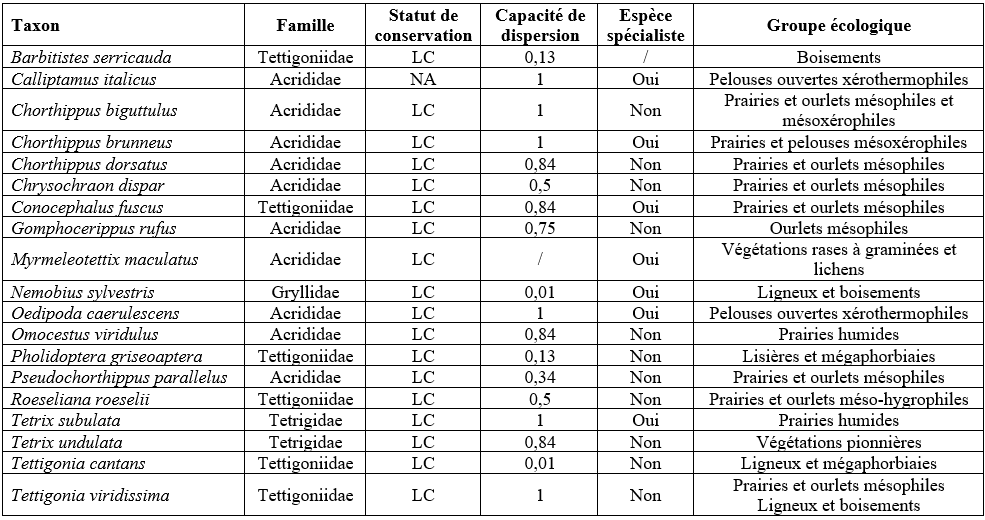
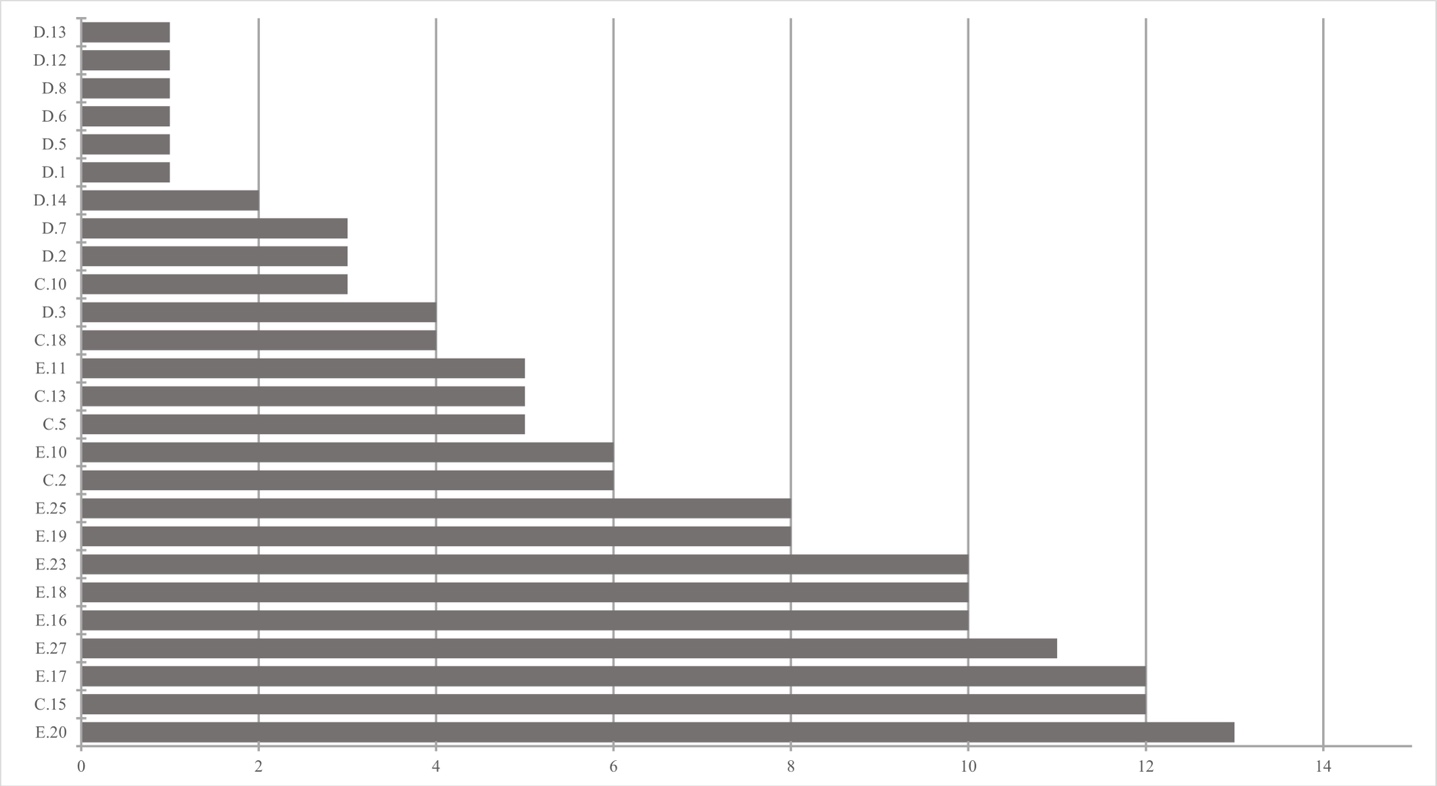
Figure 7 : Nombre total d’espèces d’orthoptères par site.
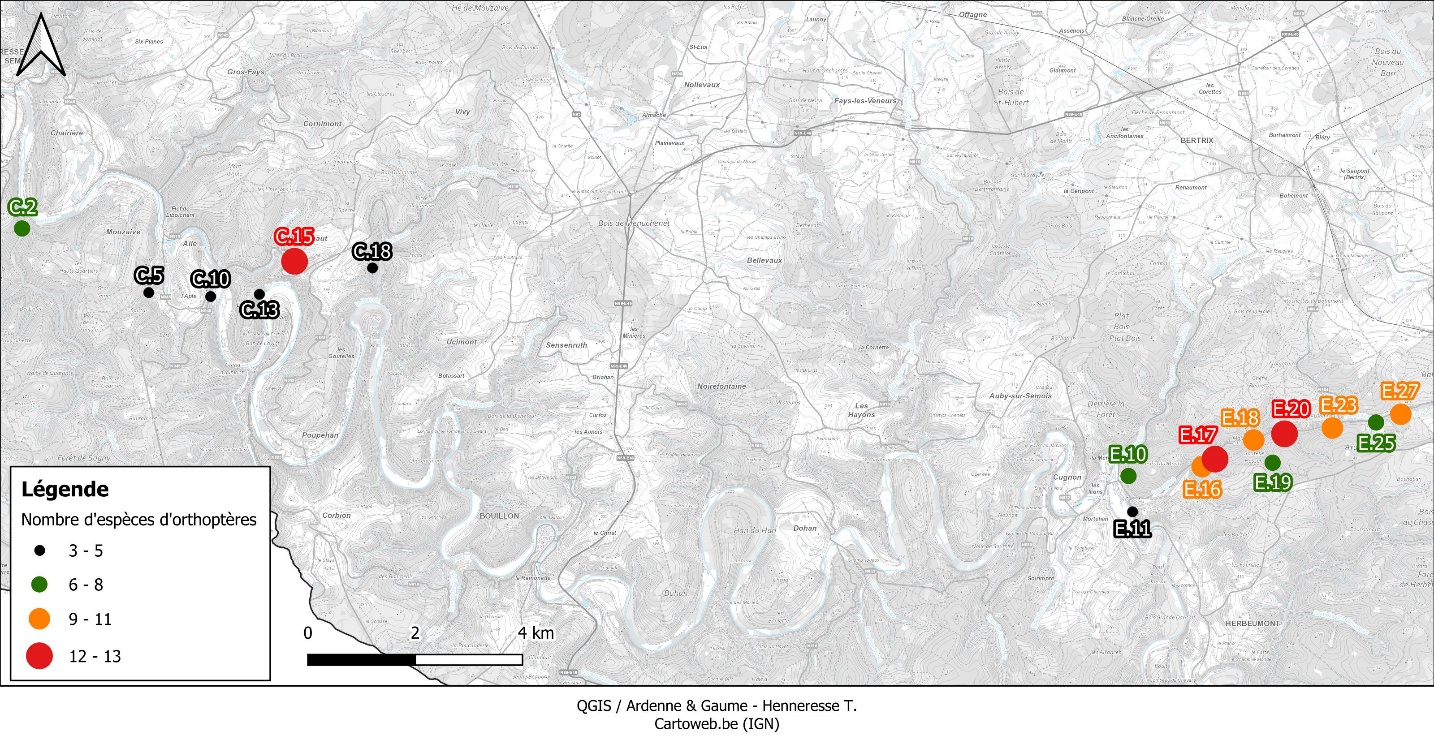
Figure 8 : Localisation des 16 sites où les orthoptères ont été recherchés activement, avec le nombre d’espèces détectées (par catégorie).
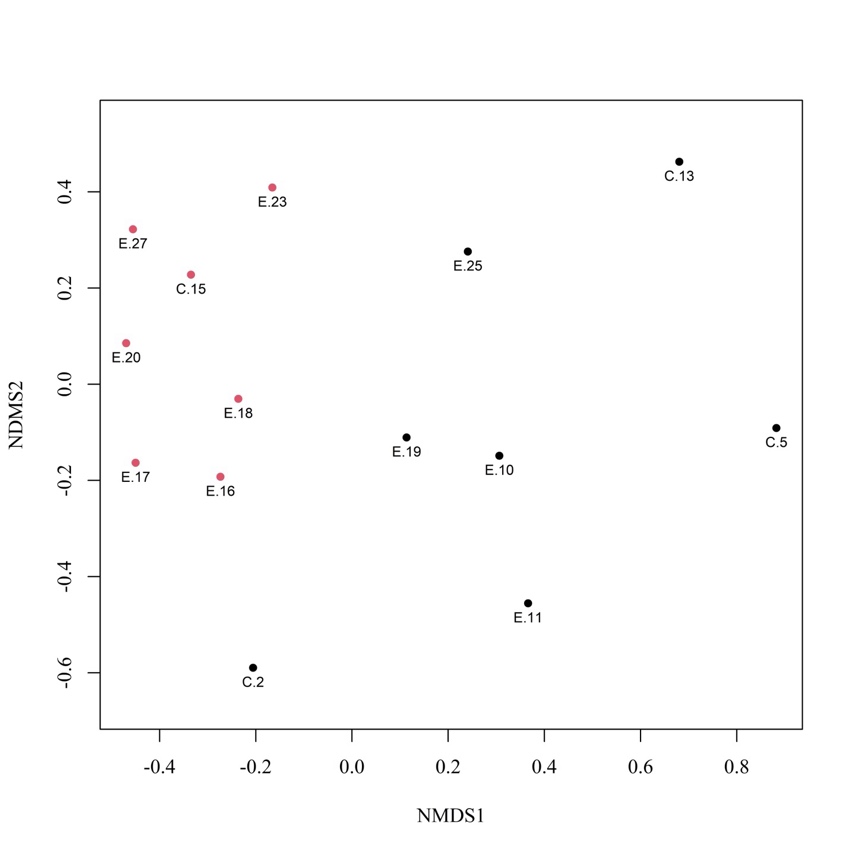
Figure 9 : Diagramme bidimensionnel d’ordination des sites réalisé par NMDS à partir des données de présence/absence des orthoptères. Les 2 groupes formés à la suite de la CAH sont représentés en rouge et noir.
DISCUSSION
35Richesse spécifique des sites
36Dans la présente étude, 41 espèces distinctes de papillons de jour ont été observées, soit 35 % de la diversité wallonne (115 espèces indigènes – Fichefet et al., 2008 + 1 espèce indigène supplémentaire après 2010 – SPW ARNE, 2010-2025). Quant aux 19 espèces d’orthoptères détectées, elles représentent 40 % de la diversité wallonne (46 espèces indigènes – Lock et al., 2011 + 1 espèce supplémentaire après 2020 – Horney, 2023). À l’échelle des huit régions ardoisières wallonnes, le chiffre atteint 49 % (23 espèces d’orthoptères) en considérant les données de Remacle (2007). En comparaison, à l’échelle du bassin minier wallon (formé des terrils issus de l’exploitation du charbon), on retrouve 57 % des espèces indigènes d’orthoptères (28 espèces – Preud’homme, 2020).
37La majorité des papillons de jour étaient rares à exceptionnels dans les sites étudiés, 52 % n’occupant qu’1 à 3 sites (Tableau 4). À l’inverse, 6 espèces y étaient assez communes à communes : Aglais io, Gonepteryx rhamni (Linnaeus 1758), Maniola jurtina (Linnaeus 1758), Pararge aegeria, Pieris napi et Vanessa atalanta (Linnaeus 1758). Il s’agit d’espèces généralistes très mobiles, très communes et non menacées en Wallonie. De telles espèces avaient été négligées par Remacle (2007) qui s’était surtout concentrée sur les espèces (quasi) menacées selon la liste rouge de l’époque (mise à jour en 2008 par Fichefet et al.). Concernant les sites où les papillons de jour ont été recherchés activement, on constate que ce sont les sites E.20 et E.27 dans la vallée du ruisseau d’Aise qui déterminent fortement la richesse spécifique à l’échelle du bassin d’Herbeumont – Bertrix ; les sites E.17, E.18, E.23 et E.25 y contribuent également, mais dans une moindre mesure (Figure 5). La superficie estimée des milieux (semi-)ouverts propices à la présence d’un cortège diversifié de papillons de jour n’est ici pas corrélée significativement à la richesse spécifique des sites (coefficient de corrélation de Spearman ρ = 0,41, p-valeur = 0,11). Pourtant, selon Öckinger & Smith (2006), la taille des prairies semi-naturelles est l’un des facteurs, avec la richesse en habitats favorables dans le paysage environnant, influençant positivement la richesse en papillons de jour des prairies semi-naturelles. Plusieurs autres variables, éventuellement en interaction, sont certainement responsables des différences entre sites, dont la situation microclimatique et l’abondance des fleurs nectarifères.
38Près d’un tiers des espèces d’orthoptères étaient rares à exceptionnelles dans les sites étudiés et n’occupaient qu’1 à 3 sites (Tableau 6). À l’inverse, 6 espèces y étaient assez communes à communes : Chorthippus brunneus (Thunberg 1815), Chrysochraon dispar (Germar 1834), Gomphocerippus rufus (Linnaeus 1758), Nemobius sylvestris (Bosc 1792), Pseudochorthippus parallelus (Zetterstedt 1821) et Tettigonia cantans (Fuessli 1775). Il s’agit d’espèces assez communes à très communes en Ardenne belge. Concernant les sites où les orthoptères ont été recherchés activement, le site C.15 et 6 sites de la vallée du ruisseau d’Aise (E.16, E.17, E.18, E.20, E.23 et E.27) déterminent fortement la richesse spécifique à l’échelle de leur bassin respectif (Figure 8). La superficie estimée des milieux (semi-)ouverts propices à la présence d’un cortège diversifié d’orthoptères n’est ici pas corrélée significativement à la richesse spécifique des sites (coefficient de corrélation de Spearman ρ = 0,41, p-valeur = 0,11). Plusieurs autres variables, éventuellement en interaction, sont certainement responsables des différences entre sites, dont l’hétérogénéité structurale de la végétation, la situation microclimatique et le type de gestion des milieux ouverts (Weiss et al., 2013).
39Richesse spécifique des bassins
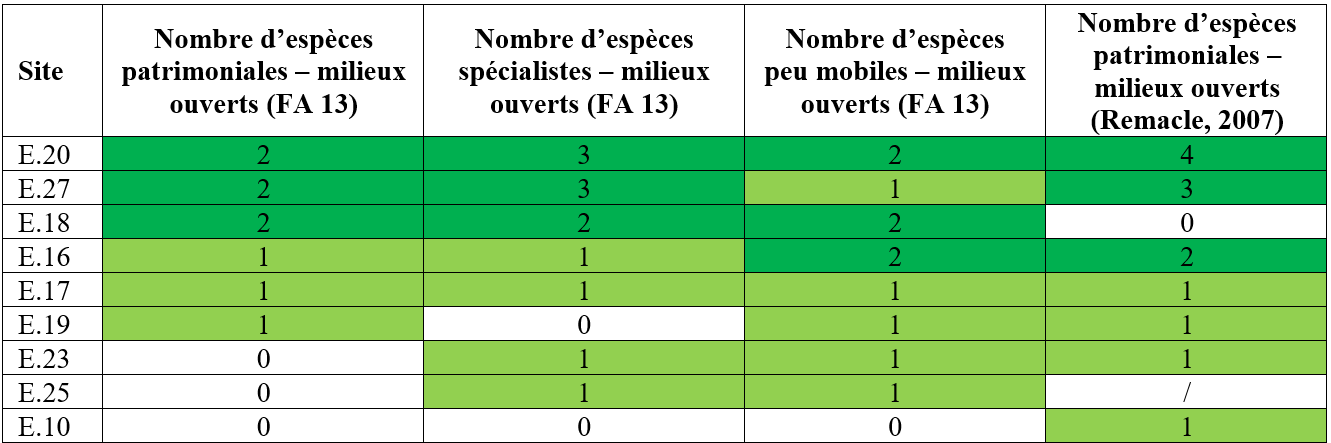
40Le bassin d’Herbeumont – Bertrix est le plus riche en papillons de jour avec 41 espèces, ceux d’Alle – Rochehaut et de Fays-les-Veneurs étant loin derrière avec respectivement 17 et 7 espèces distinctes. Ces différences peuvent en partie s’expliquer par le fait que les papillons de jour n’ont pas été recherchés activement dans le bassin de Fays-les-Veneurs. De plus, à l’échelle des carrés U.T.M. de 25 km², en considérant les données de 2001 à 2007 (Fichefet et al., 2008 – anciennes mais tout de même indicatives), on peut remarquer que le nombre d’espèces de papillons de jour est bien moins important au niveau des carrés abritant la quasi-totalité des sites des régions d’Alle – Rochehaut et de Fays-les-Veneurs (6 à 15 espèces) qu’au niveau de ceux abritant la majorité des sites du bassin d’Herbeumont - Bertrix (1 carré avec 16 à 30 espèces et deux carrés avec 31 à 50 espèces). Il est probable que de multiples facteurs déterminent ces différences de richesse entre bassins, notamment via des effets à l’échelle des sites individuels (p. ex., la composition du paysage et en particulier la proportion d’habitats favorables ou encore l’abondance des fleurs nectarifères – Öckinger & Smith, 2006 ; Milberg et al., 2016).
41Araschnia levana (Linnaeus 1758), Argynnis paphia (Linnaeus 1758), Brenthis daphne (Bergsträsser 1780), G. rhamni et P. aegeria et P. napi sont les seules espèces de papillons de jour à avoir été détectées dans les trois bassins étudiés. Dans son travail, Remacle (2007) s’était concentrée sur les espèces (quasi) menacées et sur quelques espèces non menacées mais dont la présence en Ardenne belge semblait à l’époque plus ou moins localisée. Elle listait 25 papillons de jour pris en compte et inventoriés, dont 19 détectés au sein des trois bassins étudiés. Sur ces 19 espèces, 11 (58 %) ont été à nouveau observées par l’auteur, plus précisément 11/11 dans le bassin d’Herbeumont – Bertrix, 2/11 dans celui d’Alle – Rochehaut et 1/11 dans celui de Fays-les-Veneurs. Parmi les espèces non retrouvées, la majorité sont (quasi) menacées et font l’objet d’un commentaire ci-après.
42Concernant les orthoptères, les résultats de l’auteur sont équivalents à ceux fournis par Remacle (2007), avec respectivement 19 et 20 espèces dans les trois bassins étudiés. Considérés individuellement, c’est également le cas au niveau des bassins d’Alle – Rochehaut (13 espèces pour chacune des études) et d’Herbeumont – Bertrix (18 vs 20 espèces). Concernant celui de Fays-les-Veneurs, seules 6 espèces ont été détectées par l’auteur contre 11 par Remacle (2007), cette différence pouvant en partie s’expliquer par le fait que les orthoptères n’y ont pas été recherchés activement. Il est probable que de multiples facteurs déterminent la richesse plus importante dans le bassin d’Herbeumont – Bertrix et assez faible dans celui de Fays-les-Veneurs. À l’échelle des carrés U.T.M. de 25 km², on remarque que le nombre d’espèces d’orthoptères (données de 2000 à 2018 – Lock et al., 2020 ; Adriaens et al., 2025) est bien moins important au niveau des 2 carrés abritant la région de Fays-les-Veneurs (3 et 7 espèces) qu’au niveau de ceux abritant la majorité des sites des 2 autres bassins (> 10 espèces). L’hétérogénéité du paysage (Schouten et al., 2009) pourrait ici être déterminante pour expliquer la variation spatiale de la richesse spécifique.
43Chorthippus brunneus, Gomphocerippus rufus, Nemobius sylvestris, Pseudochorthippus parallelus, Tettigonia cantans et Tettigonia viridissima (Linnaeus 1758) sont les seules espèces à avoir été détectées dans les trois bassins étudiés. Cette liste d’espèces est assez similaire à celle, établie par Remacle (2007), des espèces trouvées dans le plus grand nombre de sites à l’échelle des huit bassins (C. brunneus, C. dispar, G. rufus, N. sylvestris, Omocestus viridulus, P. parallelus et Tetrix undulata (Sowerby 1806)). Il s’agit d’espèces assez communes à très communes en Ardenne belge dont la présence n’a rien d’inattendu. Remacle (2007) identifie C. brunneus comme l’espèce la mieux représentée sur les déblais ardoisiers aux premiers stades de la dynamique de végétation. La majorité des sites où il n'a pas été détecté par l’auteur n’étaient a priori pas favorables à la présence d’un cortège diversifié d’orthoptères.
44Similarité et composition spécifique des sites
45L’ordination et la classification des sites révèle que le site C.15 et la majorité des sites de la vallée du ruisseau d’Aise (E.16 à E.20, E.25 et E.27) peuvent être regroupés (groupe I) en ce qui concerne la composition spécifique en papillons de jour (Figure 6). Dans ces 8 sites, les formations herbeuses maigres (prairies mésophiles, pelouses sèches, etc.) sont généralement bien représentées et tendent à abriter davantage d’espèces spécialistes et/ou peu mobiles telles que M. diamina et P. malvae (espèce indicatrice du groupe I). Ce n’est pas le cas du site C.15 qui abrite toutefois deux espèces du genre Thymelicus (Hübner 1819) presqu’uniquement détecté dans les sites E.16 à E.20, E.23 et E.27 (T. lineola étant une espèce indicatrice du groupe I). Rappelons que le site C.15 se situerait à la limite de décision entre les deux clusters, sur base du coefficient de silhouette. Notons que les sites composant le groupe I ne forment pas un cluster compact. Ce cluster semble cependant offrir une description simplifiée cohérente de la réalité et est assez clairement séparé de celui formant le groupe II. Malgré des similitudes avec les sites du groupe I, le site E.23 est classé par l’analyse dans le groupe II mais pourrait se trouver à la limite de décision entre les deux clusters. Comparé à la plupart des sites du groupe I, le site E.23 abrite en effet à la fois sensiblement moins d’espèces spécialistes (6 %) et peu mobiles (6 %). Quant au groupe II, il ne forme pas un ensemble cohérent sur le plan de l’ordination (Figure 6) et ses sites pourraient plutôt rassemblés à la suite de la CAH sur base de leur dissimilarité vis-à-vis des sites du groupe I. On remarque toutefois que la majorité des espèces retrouvées dans les sites du groupe II sont assez généralistes et mobiles, à l’instar de P. aegeria, espèce indicatrice de ce groupe. On remarque également que toutes les espèces du groupe II se retrouvent également dans le groupe I (incluant toutes les espèces détectées), ce qui suggère que la composition spécifique du groupe II constitue une version « appauvrie » de celle du groupe I. Plusieurs facteurs pourraient expliquer les différences de composition spécifique entre groupes, notamment la connectivité des sites, les conditions microclimatiques, l’abondance des ressources florales et l’affectation ainsi que la gestion des sites.
46Selon l’analyse des données, les sites C.15, E.16 à E.18, E.20, E.23 et E.27 sont proches en termes de composition spécifique en orthoptères (Figure 9). Dans ces sites formant un cluster compact (groupe I), les formations herbeuses maigres sont bien représentées et abritent des espèces typiques telles que Chorthippus biguttulus (Linnaeus 1758) et C. dorsatus, espèce indicatrice de ce groupe. L’autre espèce indicatrice du groupe I, Omocestus viridulus, est plutôt typique des prairies humides (Tableau 7) mais peut être retrouvée dans des habitats variés pourvus d’une végétation herbacée suffisamment dense et ni trop haute ou basse (Couvreur & Godeau, 2000). Les sites du groupe II (caractérisé par l’absence d’espèce indicatrice) ne forment pas un ensemble cohérent sur le plan de l’ordination (Figure 9) et seraient plutôt rassemblés à la suite de la CAH sur base de leur dissimilarité vis-à-vis des sites du groupe I. En outre, sur base du coefficient de silhouette, les sites E.19 et E.25 se situeraient à la limite de décision entre les deux clusters et le site C.2 pourrait être classé erronément dans le groupe II. Plusieurs facteurs pourraient expliquer les différences de composition spécifique entre les groupes, notamment la connectivité des sites, les conditions microclimatiques (influence généralement plus forte du milieu forestier environnant dans les sites du groupe II) et l’affectation (fréquentation, infrastructures, etc.) ainsi que la gestion des sites.
47Espèces patrimoniales
48Au total, 9 espèces patrimoniales ont été détectées par l’auteur. Il s’agit de 2 espèces d’orthoptères (Calliptamus italicus et Oedipoda caerulescens) et de 7 espèces de Rhopalocères (Callophrys rubi (Linnaeus 1758), Fabriciana adippe ([Denis & Schiffermüller] 1775), Favonius quercus (Linnaeus 1758), Pyrgus malvae, Satyrium ilicis (Esper 1779), Speyeria aglaja (Linnaeus 1758) et Thymelicus lineola). Certaines d’entre elles font l’objet d’un commentaire ci-dessous.
49Sur les 11 signalées par Remacle (2007) dans les trois bassins étudiés, seules 3 ont été observées par l’auteur :
-
Callophrys rubi : signalé dans 4 sites par Remacle (2007) et dans 2 sites par l’auteur (1 site commun). Il est probable qu’il soit actuellement toujours présent dans davantage de sites. Ce papillon considéré comme rare en Ardenne belge par Fichefet et al. (2008) y a été détecté beaucoup plus largement entre 2010 et 2022 (SPW ARNE, 2010-2025), ce commentaire étant valable pour d’autres espèces (M. diaminea, P. machaon, P. malvae, etc.). Une évolution de l’échantillonnage général via une augmentation du nombre d’observateurs et d’observations encodées sur Observations.be pourraient en partie expliquer cette « expansion » (entre 2008 et 2019, 15 fois plus de bénévoles qui encodent des données de papillons de jour – Dufrêne et al., 2019) ;
-
Pyrgus malvae : signalé dans 7 sites par Remacle (2007) et dans 5 sites par l’auteur (4 sites communs). Aux Pays-Bas, de Schaetzen et al. (2018) mettent en évidence un possible effet bénéfique des sangliers sur cette espèce. Par leur activité de fouissage, les sangliers créent et permettent le développement de microhabitats favorables aux chenilles de P. malvae. Ils pourraient donc contribuer à la conservation de l’espèce (à condition que l’effet positif du fouissage surpasse son impact négatif lié à la destruction de l’habitat), en complément du pâturage extensif de son habitat assuré par le bétail ou le gibier ;
-
Satyrium ilicis : uniquement signalé en E.20 par Remacle (2007) et en E.25 (non inventorié par Remacle) par l’auteur. Spécialiste et peu mobile, son habitat optimal est représenté par les forêts caducifoliées thermophiles. Des mesures telles la création et le maintien de lisières étagées et la création en rotation de clairières via la coupe d’essences exotiques ou l’application du régime de taillis lui seraient bénéfiques (Maes et al., 2014).
50Parmi les 8 autres espèces, compte tenu des données récentes (2020-2025) disponibles sur le site web Observations.be (Source : Observations.be, Natagora, Natuurpunt et Fondation « Observation International »1), 4 pourraient encore être présentes dans certains des sites étudiés (ou dans la région, avec alors la possibilité d’y établir des populations viables vu leur capacité de dispersion moyenne et le maintien ou la restauration d’habitats adéquats) : Boloria selene ([Denis & Schiffermüller] 1775), Cyaniris semiargus (Rottemburg 1775), Lycaena hippothoe (Linnaeus 1760) et Lysandra coridon (Poda 1761). Il s’agit d’espèces spécialistes (sauf C. semiargus qui est moyennement généraliste) associées aux prairies humides (B. selene et L. hippothoe), aux prairies mésophiles (C. semiargus) et aux pelouses sèches (L. coridon).
51Concernant les 4 dernières espèces, le pronostic est assez négatif :
-
Lycaena virgaureae (Linnaeus 1758) : la persistance actuelle de populations viables dans les sites étudiés est incertaine voire improbable. D’après Remacle (2007), la région forestière d’Herbeumont-Bertrix abritait un noyau de population de L. virgaureae. Une partie de cette population était présente dans la vallée du ruisseau d’Aise, au niveau des anciennes ardoisières pourvues de milieux ouverts. Des prospections ciblées organisées en 2025 dans les régions de Fays-les-Veneurs et d’Herbeumont – Bertrix par l’équipe du PNVS n’ont pas permis de confirmer la présence actuelle de l’espèce (Tinel C., comm. pers.). Plus largement, depuis 2019, seules 2 observations ont été encodées sur Observations.be, une en 2020 et l’autre en 2023 (Source : Observations.be et al.), ce qui témoigne de la régression extrême de L. virgaureae ;
-
Nymphalis antiopa (Linnaeus 1758) : la donnée signalée par Remacle (2007) en E.27 pourrait correspondre à celle d’un individu issu d’un afflux d’immigrants, ces afflux n’aboutissant que très rarement à une implantation durable dans les sites atteints (Fichefet et al., 2008). D’après les données disponibles, aucune population n’existerait actuellement dans les trois bassins étudiés ou à proximité (Source : Observations.be et al.) ;
-
Omocestus rufipes (Zetterstedt 1821) : selon les données récentes, l’espèce n’est plus présente dans les régions ardoisières étudiées (Source : Observations.be et al.) et il est donc plutôt improbable que l’espèce, assez rare en Ardenne belge, soit retrouvée en E.10 (obs. Jeunes & Nature in Remacle, 2007) ou découverte dans un des autres sites ardoisiers dans un futur proche. Assez mobile, son apparition sporadique à la faveur de certaines conditions (climatiques et démographiques) évoquées par Remacle (2007) pourrait toutefois se reproduire ;
-
Platycleis albopunctata (Goeze 1778) : le constat réalisé pour O. rufipes s’applique également à cette espèce anciennement signalée en E.20 (obs. Jeunes & Nature in Remacle, 2007). Decleer et al. (2000) la qualifient de rarissime en Ardenne belge, ce constat semblant être toujours valable actuellement (Source : Observations.be et al.). Cette situation contraste avec celle d’autres espèces xérothermophiles qui, elles, ont significativement étendu leur aire de répartition en l’espace de quelques décennies en Europe occidentale et notamment en Allemagne (Poniatowski et al., 2018).
52Aucune des 2 espèces patrimoniales d’orthoptères détectées par l’auteur n’étaient signalées par Remacle (2007) dans les bassins étudiés :
-
Calliptamus italicus : l’espèce avait disparu en Wallonie dans la première moitié du 20ème siècle (Decleer et al., 2000) pour y réapparaître en 2019 (Gerend et al., 2020). Xérothermophile, C. italicus aurait bénéficié de la succession d’étés chauds et secs lors des années précédentes, ces évènements favorisant sa reproduction et permettant à davantage d’individus de coloniser de nouveaux sites à partir du nord de la Lorraine française, l’espèce étant très mobile (Gerend et al., 2020). Actuellement, elle semble encore assez rare en Ardenne belge. En E.19, un seul individu a été observé par l’auteur en 2023 et une population ne s’y est pas encore développé. En E.17 par contre, plusieurs individus ont été observés en 2024 et la présence de milieux ouverts chauds et secs (Figure 10) couvrant plusieurs ares aurait permis à une population de s’y établir. À l’avenir, davantage de sites du bassin d’Herbeumont – Bertrix devraient être occupés par l’espèce, notamment E.20, E.25 et E.27. Notons que malgré l’existence, en E.16, d’habitats favorables situés à moins de 100 m de ceux présents en E.17, C. italicus n’y a curieusement pas été détecté ;
-
Oedipoda caerulescens (Linnaeus 1758) : en 2007, cette espèce était signalée comme apparemment absente du bassin d’Herbeumont – Bertrix par Remacle (2007). Couvreur & Godeau (2000) mentionnaient toutefois la vallée ardennaise de la Semois comme l’une des régions où se concentrent les observations de l’espèce. Entre 2000 et 2010, selon les données de Lock et al. (2020) et d’Adriaens et al. (2025), O. caerulescens était très rare en Ardenne belge et notamment présent à l’Ardoisière Donner dans le bassin de Martelange (Remacle, 2007). Ces dernières années, l’espèce s’est grandement répandue en Ardenne belge (Source : Observations.be et al.) où elle semble actuellement assez commune. Elle pourrait avoir profité des voies ferrées pour coloniser de nouvelles zones (Devriese H., comm. pers.) En Allemagne, O. caerulescens fait partie, avec notamment C. italicus et P. albopunctata, des espèces spécialistes xérothermophiles ayant fortement bénéficié de la hausse des températures moyennes annuelle et estivale, avec une expansion significative de son aire de répartition en l’espace de 20 à 30 ans (Poniatowski et al., 2018). Observé par l’auteur en 2024 en E.17, ce criquet y trouve, comme C. italicus, des milieux chauds, secs et à végétation éparse correspondant à ses préférences écologiques (Remacle, 2007). Le fait qu’O. caerulescens n’ait pas été détecté dans des sites (notamment E.20 et E.25) abritant des surfaces importantes de milieux favorables à végétation clairsemée et à sol filtrant (Couvreur & Godeau, 2000) est étonnant mais une présence effective passée inaperçue n’est pas à exclure.


Figure 10 : Milieux ouverts xérothermophiles favorables à C. italicus et O. caerulescens à l’Ardoisière de la Goutelle Husson (E.17 – mai 2025 – © Thomas Henneresse).
53Relations plantes-insectes
54Les observations (non considérées comme exhaustives) réalisées dans le cadre de cette étude indiquent que seuls quelques taxons végétaux étaient particulièrement attractifs en termes de nombre d’espèces associées (> 5 espèces : Centaurea gr. jacea, Rubus sp., Origanum vulgare et Eupatorium cannabinum), de familles associées (> 3 familles : les 4 mêmes taxons) ou d’espèces et/ou d’imagos attirés simultanément :
-
En E.18, un plant de Cirsium vulgare (Savi) Ten. attirait simultanément 3 espèces de papillons (A. io, F. adippe et G. rhamni) pour un total 8 imagos ;
-
En E.20, une plage de quelques mètres carrés d’O. vulgare attirait simultanément 4 espèces de papillons (A. paphia, B. daphne, P. brassicae et Pieris rapae (Linnaeus 1758)) pour un total de 10 imagos ;
-
En C.13, une plage de quelques dizaines de mètres carrés de Rubus sp. en bord de Semois attirait simultanément 2 imagos de B. daphne et 4 de V. atalanta.
55La forte attractivité de plusieurs de ces taxons envers les papillons de jour est notamment documentée par Haaland & Bersier (2011) et Nijland (2025). Ajoutons que parmi les plantes typiquement visitées par les papillons de jour figurent celles mentionnées ici : les Astéracées (E. cannabinum, Centaurea spp., Cirsium spp., etc.) pourvues de grandes inflorescences jouant le rôle de « pistes d’atterrissage », les Lamiacées à petites fleurs dont la corolle est dépourvue d’une lèvre supérieure très convexe (Ajuga spp., Mentha spp., etc. – ici, O. vulgare) et les ronces (Rubus spp.) qui fournissent une grande quantité de nectar (Willmer, 2011). Quelques interactions avec des espèces exotiques ont observées, notamment avec l’espèce invasive Buddleja davidii Franch. qui était visitée en E.25 par trois des espèces les plus fréquemment observées en 2023-2025.
56Sites d’intérêt lépidoptérique et orthoptérique
57Sur base du nombre d’espèces patrimoniales, d’espèces spécialistes et d’espèces peu mobiles dont l’habitat optimal est représenté par les prairies, pelouses, ourlets ou mégaphorbiaies (Tableau 8), les 3 sites présentant actuellement le plus grand intérêt lépidoptérique sont les sites E.20 (Prigeai), E.27 (Petit Babinay) et E.18 (Maljoyeuse), suivis par 5 sites à l’intérêt moindre mais non négligeable (E.16, E.17, E.19, E.23 et E.25) ; tous sont situés dans le bassin d’Herbeumont – Bertrix et plus précisément dans la vallée du ruisseau d’Aise. Dans les années 2000, les données présentées par Remacle (2007) indiquaient que les sites E.20 et E.27 abritaient déjà le plus grand nombre d’espèces patrimoniales typiques des milieux ouverts. Les travaux de déboisement réalisés en 2025 dans le cadre de la FA 13 et la future gestion assurée par le Département de la Nature et des Forêts (DNF) dans les sites E.20, E.27 et E.19 (Collard) devraient permettre de pérenniser ou de renforcer leur intérêt lépidoptérique. Celui du site E.10 (Linglé) apparaissait en 2024-2025 comme étant très limité voire inexistant et le déboisement de 2025 et la future gestion par le DNF devraient sensiblement le développer dans les années à venir. Notons que le site E.18 a fait l’objet d’un déboisement (hors FA 13) il y a moins de cinq ans en bord de route, augmentant ainsi son intérêt lépidoptérique. En outre, en tant que carrière active, le site E.25 (Grand Babinay) devrait, à l’avenir, bénéficier d’un plan de gestion écologique incluant notamment le maintien de milieux ouverts, tel qu’indiqué dans l’avis adopté en 2022 par le Pôle Environnement du Conseil économique, social et environnemental de Wallonie et concernant la révision du plan de secteur et l’extension de la carrière.
Tableau 8 : Nombre d’espèces patrimoniales (FA 13 ; Remacle, 2007) et nombre d’espèces spécialistes et d’espèces peu mobiles (FA 13) par site. Les couleurs vert foncé et vert clair indiquent les chiffres les plus élevés par variable.
58Le site présentant le plus grand intérêt orthoptérique est le site E.17 (Goutelle Husson), notamment de par sa richesse spécifique (au minimum 26 % de la diversité wallonne) et la présence de deux espèces patrimoniales. Notons que l’essentiel des milieux (semi-)ouverts de ce site bénéficient d’un statut fort de protection depuis 2020, au sein de la réserve naturelle « La Vallée d’Aise et Suchamps » gérée par le DNF. Le site E.17 est suivi de près par les sites E.20 (Prigeai) et E.27 (Petit Babinay) qui abritent chacun 4 espèces spécialistes et une faune orthoptérique diversifiée (28 % de la diversité wallonne en E.20). Le site C.15 (Faligeotte) est également intéressant vu sa richesse spécifique, surtout à l’échelle du bassin d’Alle – Rochehaut (Figure 8). Occupé par un taillis en 2005 (Remacle, 2007), il a subi un déboisement il y a une dizaine d’années et sa partie orientale fait actuellement l’objet de dépôts de terres, de déchets inertes et de déchets verts. Le replat (Figure 11), accessible par un chemin partant de la rue de la Faligeotte, est partiellement occupé par une végétation herbacée plus ou moins basse et discontinue dans le prolongement du chemin et plus ou moins haute et dense voire colonisée par des ligneux bas en périphérie.


Figure 11 : Végétation herbacée sur le replat de l’Ardoisière de la Faligeotte (C.15 – mai et juillet 2025 – © Thomas Henneresse).
59Limites de l’étude
60De manière générale, les inventaires sont très rarement exhaustifs, d’autant plus si un seul observateur les met en œuvre, comme dans la présente étude. En l’occurrence, durant la période 2023-2025, deux espèces non détectées par l’auteur ont été signalées par d’autres membres de l’équipe du PNVS : Phaneroptera falcata (Poda 1761) en E.20 (obs. Ferrais L.) et Ruspolia nitidula (Scopoli 1786) en E.17 (obs. Casalanguida G.) (Source : Observations.be et al.). De plus, le pourcentage d’espèces présentes dans un site mais manquées n’est pas quantifiable. La principale difficulté posée par les inventaires de terrain est la détection des espèces. Celle-ci est très variable et dépend de multiples facteurs tels que les conditions météorologiques, l’activité des animaux ou encore l’observateur (expérience, état de fatigue, etc.). Pour y remédier, les prospections peuvent être standardisées à plusieurs niveaux (période de l’année fixe, toujours le même observateur, etc.), dans le but de maximiser le taux de détection et de réduire sa variabilité entre sites. Une autre limite posée par les inventaires est liée à leur nature qualitative : par exemple, la présence d’une espèce dans un site à plusieurs années d’intervalle ne fournit aucune information sur l’évolution de sa population. Ces deux limites ne remettent pas pour autant en cause la pertinence des inventaires (plus intéressants pour mettre en évidence la présence d’espèces patrimoniales que pour comparer des sites dans l’espace ou dans le temps) mais rappellent surtout que ceux-ci doivent être interprétés avec précaution (Besnard & Salles, 2010).
61Commentaires additionnels
62Quelques taxons non patrimoniaux mais intéressants à noter font ici l’objet d’un commentaire spécifique :
-
Barbitistes serricauda : surtout active la nuit et ne descendant que rarement au sol (Couvreur & Godeau, 2000), cette sauterelle arboricole n’a été observée qu’à une seule reprise par l’auteur. Dans le site E.25, dans des hautes herbes en bord de route, un individu a été capturé en 2025 lors d’une séance de capture au filet fauchoir. Une seule donnée en E.20 datant de 1997 est signalée par Remacle (2007). L’utilisation d’un détecteur à ultrasons est la meilleure façon de détecter l’espèce (Couvreur & Godeau, 2000) dont le caractère exceptionnel dans les sites étudiés n’est certainement qu’apparent. Elle devrait être présente dans davantage de sites de la vallée du ruisseau d’Aise et dans au moins certains sites du bassin d’Alle – Rochehaut ;
-
Chorthippus dorsatus : entre 1990 et 2000, il n’était pas connu en Ardenne belge et n’était signalé avec certitude qu’en Lorraine belge (Decleer et al., 2000). Remacle (2007) ne mentionne pas l’espèce, celle-ci n’étant signalée en Ardenne belge qu’après 2010 (Lock et al., 2020 ; Adriaens et al., 2025). Ces dernières années, l’espèce s’y est grandement répandue et en particulier dans sa partie méridionale (Source : Observations.be et al.). Actuellement, malgré sa progression récente, elle semble encore assez peu commune en Ardenne belge ainsi que dans les sites étudiés ;
-
Roeseliana roeselii (Hagenbach 1822) : entre 1990 et 2000, selon la carte présentée par Decleer et al. (2000), l’espèce était exceptionnelle en Ardenne belge. Elle n’est signalée dans la partie ardennaise de la vallée de la Semois qu’après 2010 (Lock et al., 2020 ; Adriaens et al., 2025), ce qui explique l’absence de mention par Remacle (2007). Ces dernières années, R. roeselii s’est énormément répandu presque partout en Ardenne belge (Source : Observations.be et al.) et peut actuellement y être considéré comme commun. Au niveau des sites étudiés, l’espèce reste par contre (curieusement) rare. En Europe occidentale, R. roeselii a connu durant les dernières décennies une forte expansion de son aire de répartition. En Allemagne, cette expansion a débuté à la fin des années 1980 et les individus macroptères, plus fréquents (ils représentent jusqu’à 20 % des individus de certaines populations, notamment celles en limite d’aire) actuellement qu’au milieu du 20ème siècle, en seraient en grande partie responsables, sous l’effet d’une forte augmentation des températures moyennes annuelle et estivale (Poniatowski et al., 2018) ;
-
Tettigonia cantans : avant les années 1980, l’espèce n’était pas connue dans le massif ardennais et la vallée de la Semois (Couvreur 1998). Plus précisément, aucune publication sur les orthoptères en Belgique n’a été produite entre 1953 et 1980 et la date du début de la colonisation de la vallée de la Semois est inconnue (Devriese H., comm. pers.). Par la suite, elle se serait répandue en Ardenne belge via cette vallée en remplaçant T. viridissima dans les sites au microclimat froid et humide qui lui étaient les plus propices. Un comportement d’évitement ou un phénomène de compétition interspécifique en faveur de T. cantans pourrait être à l’œuvre et expliquer l’absence de T. viridissima dans des sites qui lui conviennent particulièrement (Couvreur 1998). Ces hypothèses pourraient s’appliquer aux bassins étudiés : sur les 15 sites où T. cantans a été détecté par l’auteur, seuls 3 accueillaient également T. viridissima (observé dans un total de 4 sites). En outre, sur les 14 sites où la première espèce avait été détectée par Remacle (2007), seuls 4 accueillaient également la deuxième. Notons que T. viridissima n’a été retrouvé dans aucun des sites où Remacle (2007) l’avait signalé ; inversement, aucun des sites où l’espèce a été détectée par l’auteur n’était signalé comme occupé par Remacle (2007). Quant à T. cantans, les données des deux études sont peu similaires, ce qui pourrait en partie s’expliquer par des différences d’effort ou de période d’échantillonnage.
CONCLUSION
63Au sein des trois bassins ardoisiers ciblés, cette étude a permis de mettre à jour une part importante des connaissances entomologiques résultant du travail de Remacle (2007). Globalement, ces régions concentrent une part notable de la diversité indigène wallonne, avec 35 % des papillons de jour et 40 % des orthoptères. Une dizaine de sites ardoisiers présentent actuellement un certain intérêt lépidoptérique et/ou orthoptérique, les sites E.17 (Goutelle Husson), E.20 (Prigeai) et E.27 (Petit Babinay) étant les plus intéressants pour les deux groupes biologiques pris ensemble. Plusieurs espèces patrimoniales s’y développent, qu’il s’agisse d’espèces en expansion (Calliptamus italicus) ou au statut de conservation préoccupant (en particulier Satyrium ilicis et Speyeria aglaja).
64Les milieux ouverts des sites d’intérêt bénéficient déjà (Goutelle Husson) ou devraient bénéficier dans le futur (Prigeai et Petit Babinay) d’un statut fort de protection (assurée ici par le DNF). Bien qu’indispensable, le statut de réserve naturelle n’est pas suffisant et doit absolument être associé à une gestion appropriée des habitats ouverts et des écotones. Une gestion adéquate des sites est en effet indispensable afin notamment de maintenir une diversité et une abondance florales élevées (notamment Centaurea gr. jacea et Origanum vulgare évoqués plus haut) et une hétérogénéité dans la structure du tapis végétal et donc dans les conditions microclimatiques. Le pâturage extensif à faible charge de bétail associé au contrôle des rejets ligneux par débroussaillage apparaît comme une méthode de gestion appropriée et soutenable (financièrement) à long terme pour maintenir les habitats ouverts de ces sites. Pour garantir et améliorer la connectivité des sites gérés, la gestion extensive et l’augmentation des superficies d’habitats linéaires reliant les sites comme les lisières et bords de routes/chemins ne doit pas être négligée.
65En plus de leur gestion, les habitats ouverts méritent une surveillance continue afin d’étudier l’évolution des populations de papillons de jour et d’orthoptères et d’estimer les tendances dans les sites les plus intéressants. En outre, afin d’évaluer les effets engendrés par la gestion des réserves et de pouvoir éventuellement l’adapter si elle ne permet pas d’atteindre les objectifs de conservation fixés, un suivi biologique à long terme se doit d’être mis en place. Dans ce cadre, il est de la responsabilité des pouvoirs publics de fournir les moyens humains et financiers nécessaires à la bonne exécution de ce suivi (et de la gestion) par le DNF ou les autres gestionnaires agréés.
REMERCIEMENTS
Je tiens à remercier les cantonnements de Bièvre, de Bouillon et de Neufchâteau du Département de la Nature et des Forêts ainsi que plusieurs propriétaires privés pour les autorisations d’accès ou renseignements fournis ainsi que pour les visites de terrain, en particulier Stéphan Adant, Vincent Brichart, Yves Crul, Ann De Neve, Benjamin de Potter, Pierre Gigounon, Marie-France Jacques, Stéphane Michaux, Benjamin Nollevaux et Benoît Pierlot. Je remercie également Hendrik Devriese Marc Dufrêne pour leur relecture et commentaires du manuscrit et pour les discussions visant à l’améliorer. Enfin, je remercie Arnaud Segers pour l’évaluation du manuscrit et les propositions d’améliorations apportées.
Bibliographie
Adriaens T., Decleer K., Devriese H., Lock K., Lambrechts J., San Martin y Gomez G., Piesschaert F., Maes D., Brosens D. & Desmet P., 2025. Saltabel - Orthoptera in Belgium. Version 5.9. Research Institute for Nature and Forest (INBO). Occurrence dataset https://doi.org/10.15468/1rcpsq, (30/10/2025).
Bakker J.D., 2024. Applied Multivariate Statistics in R. Seattle: University of Washington. https://uw.pressbooks.pub/appliedmultivariatestatistics/, (24/10/2025).
Barkmann F., Huemer P., Tappeiner U., Tasser E. & Rüdisser J., 2023. Standardized butterfly surveys: comparing transect counts and area-time counts in insect monitoring. Biodiversity and Conservation, 32(3), 987-1004.
Besnard A. & Salles J.-M., 2010. Suivi scientifique d’espèces animales. Aspects méthodologiques essentiels pour l'élaboration de protocoles de suivis. Note méthodologique à l'usage des gestionnaires de sites Natura 2000. DREAL PACA – Pôle Natura 2000.
Bisteau E., Baugnée J.-Y., Dufrêne M. & Mahy G., 2009. L’inventaire des Sites de Grand Intérêt Biologique en Région wallonne. Forêt Wallonne, 103, 31-40.
Castiau E. & Godart M.-F., 2017. L’exploitation du sous-sol en Wallonie. Notes de recherche CPDT n° 73. Conférence permanente du Développement territorial.
Claerebout S., 2012. Observation surprenante d’Acanthacris ruficornis citrina (Serville) en Belgique (Orthoptera, Caelifera, Acrididae). L’Érable, 36(4), 13-16.
Claerebout S., 2013. Clé de détermination photographique des criquets, sauterelles, grillons et tétrix de Belgique. Vierves-sur-Viroin : Cercles des Naturalistes de Belgique.
Couvreur J.-M., 1998. Tettigonia cantans (Fuessly, 1775) : avec ou sans Tettigonia viridissima Linné, 1758 ? Saltabel, 17, 11-20.
Couvreur J.-M. & Godeau J.-F., 2000. Atlas des Orthoptères de la Famenne (Criquets, sauterelles et grillons). Gembloux : Centre de Recherche de la Nature, des Forêts et du Bois, Jeunes & Nature a.s.b.l. et Groupe d’étude des Orthoptères du Benelux – SALTABEL.
De Cáceres M. & Legendre P., 2009. Associations between species and groups of sites: indices and statistical inference. Ecology, 90(12), 3566-3574.
Decleer K., Devriese H., Hofmans K., Lock K., Barenbrug B. & Maes D., 2000. Voorlopige atlas en "rode lijst" van de sprinkhanen en krekels van België (Insecta, Orthoptera)/Atlas et « liste rouge » provisoire des sauterelles, grillons et criquets de Belgique (Insecta, Orthoptera). Rapport Instituut voor Natuurbehoud/Rapport Instituut voor Natuurbehoud 2000/10. Brussel/Bruxelles : Werkgroep Saltabel i.s.m. I.N. en K.B.I.N./Groupe de travail Saltabel e.c.a. I.N. et I.R.S.N.B.
de Schaetzen F., van Langevelde F. & WallisDeVries M.F., 2018. The influence of wild boar (Sus scrofa) on microhabitat quality for the endangered butterfly Pyrgus malvae in the Netherlands. Journal of Insect Conservation, 22(1), 51-59.
Desmet P., Reyserhove L., Oldoni D., Groom Q., Adriaens T., Vanderhoeven S. & Pagad S., 2025. Global Register of Introduced and Invasive Species - Belgium. Version 1.15. Invasive Species Specialist Group ISSG. Checklist dataset https://doi.org/10.15468/xoidmd, (09/01/2026).
Dray S., Dufour A.-B., Thioulouse J., Chessel D., Jombart T., Pavoine S., Lobry J.R., Ollier S., Borcard D., Legendre P., Bougeard S. & Siberchicot A., 2025. ade4: Analysis of Ecological Data: Exploratory and Euclidean Methods in Environmental Sciences. R package version 1.7-23. https://cran.r-project.org/web/packages/ade4.
Dufrêne M., Barbier Y., Paquet J.-Y., Goffart P., Motte G., Fichefet V. & Cors R., 2019. Le suivi des papillons de jour et des odonates en Wallonie – Naturalistes bénévoles et professionnels : des apports complémentaires. Carnets des Espaces Naturels, 2, 22-24.
Dupont P., 2015. Base de données de connaissance sur les Lépidoptères Rhopalocères. Version 01. MNHN-SPN, http://inpn.mnhn.fr/telechargement/documentation/natura2000/evaluation.
Fichefet V., Barbier Y., Baugnée J.-Y., Dufrêne M., Goffart P., Maes D. & Van Dyck H., 2008. Papillons de jour de Wallonie (1985 – 2007). Série « Faune – Flore – Habitat » n° 4. Gembloux : Groupe de Travail Lépidoptères Lycaena et DEMNA (SPW-DGARNE).
Freitas A.V.L., Santos J.P., Rosa A.H.B., Iserhard C.A., Richter A., Siewert R.R., Gueratto P.E., Carreira J.Y.O. & Lourenço G.M., 2021. Sampling Methods for Butterflies (Lepidoptera). In: Santos J.C. & Fernandes G.W., eds. Measuring Arthropod Biodiversity – A Handbook of Sampling Methods. Cham: Springer Nature Switzerland AG, 101-123.
Gerend R., Birlenbach F., Clemens M. & Proess R., 2020. The Italian locust Calliptamus italicus (Linnaeus, 1758), a recent addition to the Orthopteran fauna of Luxembourg (Insecta: Orthoptera, Saltatoria, Acrididae). Bulletin de la Société des Naturalistes luxembourgeois, 122, 207-210.
Gerlach J., Samways M. & Pryke J., 2013. Terrestrial invertebrates as bioindicators: an overview of available taxonomic groups. Journal of Insect Conservation, 17(4), 831-850.
Gosselin G., 2006. L’utilisation des roches en Région wallonne. Dossier scientifique réalisé dans le cadre de l’élaboration du Rapport analytique 2006-2007 sur l’État de l’Environnement wallon. Incitec SPRL.
Haaland C. & Bersier L.-F., 2011. What can sown wildflower strips contribute to butterfly conservation?: an example from a Swiss lowland agricultural landscape. Journal of Insect Conservation, 15(1-2), 301-309.
Horney G., 2023. Naturoscope. Clin d’Œil Nature, 30, 27-31.
Kleukers R.M.J.C., van Nieukerken E.J., Odé B., Willemse L.P.M. & van Wingerden W.K.R.E., 2004. De sprinkhanen en krekels van Nederland (Orthoptera). Nederlandse Fauna 1. Tweede druk. Leiden: Nationaal Natuurhistorisch Museum, knnv Uitgeverij & European Invertebrate Survey – Nederland.
Lacoeuilhe A., Roquinarc’h O., Thierry C. & Latour M., 2020. Protocole de suivi des Orthoptères pour l’étude des milieux prairiaux – Application à deux cas d’étude. UMS PatriNat – OFB/CNRS/MNHN.
Lock K., Adriaens T., Devriese H., San Martin G. & Decleer K., 2011. Updated red lists of the grasshoppers and crickets (Orthoptera) in Flanders, Brussels and Wallonia. Bulletin de la Société royale belge d’Entomologie/van de Koninklijke Belgische Vereniging voor Entomologie, 147(3), 211-225.
Lock K., Foquet B., Foquet R., San Martin G., Arijs G., Cortens J., Deflorenne P., Laudelout A., Paquet J.-Y., Desmet P. & Bronne L., 2020. Observations.be - Orthoptera occurrences in Wallonia, Belgium. Version 1.1. Natagora. Occurrence dataset https://doi.org/10.15468/r763pb, (30/10/2025).
Logghe G., Batsleer F., Maes D., Permentier T., Berg M.P., Brosens D., Cooleman S., De Smedt P., Hagge J., Lambrechts J., Pollet M., Verheyde F. & Bonte D., 2025a. An in-depth dataset of northwestern European arthropod life histories and ecological traits. Biodiversity Data Journal, 13, e146785. doi: 10.3897/BDJ.13.e146785.
Logghe G., Batsleer F., Maes D., Permentier T., Berg M.P., Brosens D., Cooleman S., De Smedt P., Hagge J., Lambrechts J., Pollet M., Verheyde F. & Bonte D., 2025b. Northwestern European arthropod life histories and ecological traits. Version 1.1. Ghent University. Checklist dataset https://doi.org/10.15468/75g5z9, (09/10/2025).
Maechler M., Rousseeuw P., Struyf A., Hubert M. & Hornik K., 2025. cluster: Cluster Analysis Basics and Extensions. R package version 2.1.8.1. https://CRAN.R-project.org/package=cluster.
Maes D., Jacobs I., Segers N., Vanreusel W., Van Daele T., Laurijssens G. & Van Dyck H., 2014. A resource-based conservation approach for an endangered ecotone species: the Ilex Hairstreak (Satyrium ilicis) in Flanders (north Belgium). Journal of Insect Conservation, 18(5), 939-950.
Milberg P., Bergman K.-O., Cronvall E., Eriksson Å.I., Glimskär A., Islamovic A., Jonason D., Löfqvist Z. & Westerberg L., 2016. Flower abundance and vegetation height as predictors for nectar-feeding insect occurrence in Swedish semi-natural grasslands. Agriculture, Ecosystems & Environment, 230, 47-54.
Nijland F., 2025. Differences in local flower visitation rates between nectaring butterfly species. Entomologische Berichten, 85(1), 19-29.
Öckinger E. & Smith H.G., 2006. Landscape composition and habitat area affects butterfly species richness in semi-natural grasslands. Oecologia, 149(3), 526-534.
Oksanen J., Simpson G.L., Blanchet F.G., Kindt R., Legendre P., Minchin P.R., O'Hara R.B., Solymos P., Stevens M.H.H., Szoecs E., Wagner H., Barbour M., Bedward M., Bolker B., Borcard D., Borman T., Carvalho G., Chirico M., De Cáceres M., Durand S., Evangelista H.B.A., FitzJohn R., Friendly M., Furneaux B., Hannigan G., Hill M.O., Lahti L., Martino C., McGlinn D., Ouellette M.-H., Ribeiro Cunha E., Smith T., Stier A., Ter Braak C.J.F. & Weedon J., 2025. vegan: Community Ecology Package. R package version 2.7-1. https://cran.r-project.org/package=vegan.
Poniatowski D., Münsch T., Helbing F. & Fartmann T., 2018. Arealveränderungen mitteleuropäischer Heuschrecken als Folge des Klimawandels. Natur und Landschaft, 93(12), 553-561.
Preud’homme J., 2020. Une biodiversité singulière – La nature à l’assaut des terrils. Carnets des Espaces Naturels, 5, 8-11.
R Core Team, 2025. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing, https://www.R-project.org/.
Reinhardt K., Köhler G., Maas S. & Detzel P., 2005. Low dispersal ability and habitat specificity promote extinctions in rare but not in widespread species: the Orthoptera of Germany. Ecography, 28(5), 593-602.
Remacle A., 2006. Les potentialités d’accueil de la vie sauvage en milieux rocheux et souterrain. Partim « Anciennes carrières ». Dossier scientifique réalisé dans le cadre de l’élaboration du Rapport analytique 2006-2007 sur l’état de l’environnement wallon. Faculté universitaire des Sciences agronomiques de Gembloux.
Remacle A., 2007. Les ardoisières de l’Ardenne belge. Intérêt biologique et état des lieux des sites en surface. Travaux n° 30. Région wallonne, Direction générale des Ressources naturelles et de l’Environnement, Division de la Nature et des Forêts.
Remacle A., 2009. Contribution des anciennes carrières à la biodiversité wallonne. Convention « Les carrières en Région wallonne : inventaire et intérêt biologique » (C45). Rapport non publié. Service public de Wallonie, Direction générale opérationnelle de l'Agriculture, des Ressources naturelles et de l'Environnement, Direction de la Nature.
Schouten M.A., Verweij P.A., Barendregt A., Kleukers R.M.J.C., Kalkman V.J. & de Ruiter P.C., 2009. Determinants of species richness patterns in the Netherlands across multiple taxonomic groups. Biodiversity and Conservation, 18(1), 203-217.
Sevilleja C.G., van Swaay C.A.M., Bourn N., Collins S., Settele J., Warren M.S., Wynhoff I. & Roy D.B., 2019. Butterfly Transect Counts: Manual to monitor butterflies. Report VS2019.016. Wageningen: Butterfly Conservation Europe & De Vlinderstichting/Dutch Butterfly Conservation.
Sperber C.F., Zefa E., de Oliveira E.C., de Campos L.D., Bolfarini M.P., Fianco M., Lhano M.G., Vicente N., Szinwelski N., de Souza Dias P.G.B., Acosta R.C. & Prasniewski V.M., 2021. Measuring Orthoptera Diversity. In: Santos J.C. & Fernandes G.W., eds. Measuring Arthropod Biodiversity – A Handbook of Sampling Methods. Cham: Springer Nature Switzerland AG, 257-287.
SPW ARNE (Agriculture, Ressources naturelles et Environnement) (éditeur), 2010-2025. Portail de la Biodiversité en Wallonie. https://biodiversite-old.wallonie.be, (27/10/2025).
Verloove F. & Van Rossum F. (& coll.), 2024. Nouvelle Flore de la Belgique, du G.-D. de Luxembourg, du Nord de la France et des Régions voisines (Ptéridophytes et Spermatophytes). Septième édition. Meise : Jardin botanique de Meise.
Weiss N., Zucchi H. & Hochkirch A., 2013. The effects of grassland management and aspect on Orthoptera diversity and abundance: site conditions are as important as management. Biodiversity and Conservation, 22(10), 2167-2178.
Willmer P., 2011. Pollination and Floral Ecology. Princeton: Princeton University Press.
(50 Réf.)
Notes
1 Simplifié en « Observations.be et al. » dans la suite de l’article. Site consulté le 14 novembre 2025.
Pour citer cet article
