


- Portada
- Volume 71 (2018)
- Tritomegas rotundipennis (Dohrn, 1862), une nouvelle espèce pour la faune belge passée inaperçue pendant plus d’un siècle et répartition des trois espèces de Tritomegas en Belgique (Hemiptera: Heteroptera: Cydnidae)


Vista(s): 2496 (32 ULiège)
Descargar(s): 413 (16 ULiège)

Tritomegas rotundipennis (Dohrn, 1862), une nouvelle espèce pour la faune belge passée inaperçue pendant plus d’un siècle et répartition des trois espèces de Tritomegas en Belgique (Hemiptera: Heteroptera: Cydnidae)

Notes de la rédaction
Reçu le 3 avril 2017, accepté le 5 mai 2017
Résumé
Tritomegas rotundipennis (Dohrn, 1862) (Hemiptera: Heteroptera: Cydnidae), une nouvelle espèce pour la Belgique. Nous résumons les données provenant de la littérature entomologique, de spécimens de collections, d’observations de terrain et des ressources internet pour examiner l'état des connaissances concernant les trois espèces de Tritomegas spp. renseignées de Belgique. Leur biologie générale et leur répartition géographique sont discutées.
Abstract
Tritomegas rotundipennis (Dohrn, 1862) (Hemiptera: Heteroptera: Cydnidae), a new species to the Belgian fauna. We summarize data from published entomological literature, collected specimens, field observations and biodiversity web resources to review the state of knowledge regarding the three species of Tritomegas from Belgium. Their general biology and geographic distribution are discussed.
Tabla de contenidos
Introduction
1Les Cydnidae (Hémiptères Hétéroptères) sont proches des Pentatomidae (super-famille des Pentatomoidea) et comptent 89 genres et 680 espèces dans le monde (Lis, 2006). Une quinzaine d’espèces seulement peuvent parfois occasionner de sérieux dégâts aux cultures (Lis et al., 2000). Aucune d’elles ne se rencontre en Belgique.
2Depuis la découverte chez nous de Geotomus petiti Wagner, 1954 (Aukema et al., 2014), la Belgique compte 14 espèces (Baugnée et al., 2003). Jusqu’il y a peu, seules deux espèces appartenant au genre Tritomegas avaient été recensées : T. bicolor (Linnaeus, 1758) et T. sexmaculatus (Rambur, 1839). Passée inaperçue pendant plus de cent ans, la présence de T.rotundipennis (Dohrn, 1862) est signalée pour la première fois, portant ainsi à 15 le nombre de Cydnidae belges.
3Deux sous-familles sont représentées dans notre faune : les Cydninae (plutôt fouisseurs) et les Sehirinae (davantage floricoles). Au sein des Sehirinae, le genre Tritomegas forme un groupe homogène dont les imagos sont assez faciles à identifier sur base de leur seule apparence externe, à l’œil nu, voire même sur base photographique, sans recourir obligatoirement à l’étude des genitalia. Six espèces de ce genre sont renseignées de l’Ancien Monde et sont de taille relativement faible avec une envergure oscillant entre 7,5 et 14,0 mm (Froeschner, 1960). Ce genre se distingue de tous les autres grâce à la combinaison des critères suivants pris ensemble : (1) clypéus toujours libre, jamais enserré par les jugas ; (2) bord latéral du pronotum présentant une large tache blanche se réduisant vers l’arrière ; (3) bords latéro-antérieur et postérieur des cories avec une tache blanche développée ; et (4) pattes noires dont la base du tibia est maculée de blanc (Charlot & Claerebout, 2016). L’identification des espèces au stade imaginal peut se faire à l’aide des clés de Lupoli & Dusoulier (2015), de Charlot & Claerebout (2016) ou de Stichel (1961). Les juvéniles au stade V sont colorés très différemment des adultes et permettent une identification spécifique plus aisée, grâce aux clés de Lupoli et al. (2014). Les photos des planches I et II permettent déjà néanmoins de distinguer les adultes (Planche I, Figures 1 - 6) et les juvéniles (Planche II, Figures 7 - 9) des trois espèces présentes dans notre pays.
4La structure des armatures mâles permet également de séparer nettement les différentes espèces du genre Tritomegas. C’est Vidal (1949) qui, pour la première fois, figure les paramères de T.bicolor et de T. sexmaculatus. Ensuite, la confusion règne dans les résultats des travaux de différents auteurs, notamment dans celui de Wagner (1963), excellent par ailleurs à maints égards. C’est à Kammerschen (1986a et 1986b) que revient le mérite d’avoir éclairci la situation, en décrivant en détail les genitalia mâles, en les comparant dans un tableau synthétique et en les illustrant convenablement, pour chacune de ces trois espèces. En outre, les spermathèques de nombreux genres de Cydnidae ont été magnifiquement illustrées dans une étude sur leur morphologie, mais ne comprenant que celle de T. sexmaculatus (Pluot-Sigwalt & Lis, 2008). Les genitalia externes femelles ont été illustrés par Wagner (1963), mais ne présentent pas de caractéristiques utiles. On notera simplement, que chez T. rotundipennis et T. bicolor, la longueur des plaques est peu élevée, alors qu’elle est très grande chez T. sexmaculatus (Wagner, 1963).
5Depuis les publications de Leston (1954) et Jordan (1958), la stridulation émise par les représentants de la famille des Cydnidae est connue. L’appareil stridulatoire est formé par la base de l’aile membraneuse finement denticulée, principalement au niveau de la bifurcation de la nervure cubitale, venant frotter sur le bord postérieurdes premiers tergites abdominaux (Jordan, 1958). Dans le cas des trois espèces apparentées et sympatriques de Tritomegas, différentes émissions sonores ont été enregistrées et analysées dans une étude menée par Gogala & Hočevar (1990). Cette étude bioacoustique comparative montre une nette distinction entre les trois espèces à différents niveaux, même si leurs émissions sonores montrent des similitudes dans leur structure. Les chants prénuptiaux, les chants nuptiaux, en particulier la partie consacrée au chant de cours des mâles, et les chants de rivalité émis par les mâles vis-à-vis d’autres mâles sont tous clairement spécifiques. Par contre, il n’existe pas de différence entre les trois espèces lorsqu’il s’agit de signaux de détresse et de perturbation émis tant par les mâles que par les femelles, ainsi que lors du chant d’acceptation émis par les femelles vierges et sexuellement matures, en réponse aux parades nuptiales des mâles conspécifiques.
6Suite à la découverte d’une troisième espèce du genre Tritomegas en Belgique, il nous paraît intéressant de faire le point sur l’état de nos connaissances.
Matériel et Méthodes
7Nos données proviennent de plusieurs sources : la littérature, l’analyse des collections privées et publiques, dont celles de l’Institut royal des Sciences naturelles de Belgique, de Gembloux Agro-Bio Tech (anciennement Faculté des Sciences agronomiques de Gembloux), de la Haute École de la Province de Liège (conservées dans le conservatoire de Gembloux), de la Haute École Provinciale de Hainaut - Condorcet et de l’Université de Mons. Les mentions figurant sur le site Observations.be et dans la banque de données de l’Observatoire Faune - Flore - Habitats du Service Public de Wallonie ont aussi été prises en compte.
8Si l’identification correcte de spécimens issus de collections ne pose plus aucun problème, il n’en va pas de même pour celle d’individus mentionnés sur internet. En effet, la validation nécessite quelques précautions particulières (Claerebout, 2011) car ces données ne sont pas toujours fiables.
9Dans le cadre du présent travail, nous ne prendrons en compte que les données certifiées par les auteurs ou par les validateurs reconnus par les responsables des portails d’encodage. Nous considérerons comme valide toute donnée soit basée sur un spécimen disponible en collection, privée ou publique, soit basée sur une mention figurant sur internet et accompagnée d’une photographie montrant les critères diagnostiques. Toute mention figurant sur internet non accompagnée d’une photographie même provenant d’un observateur ayant déjà publié une telle mention avec une photographie ne sera pas prise en compte.
10Abréviations utilisées
11Institutions
12CEL : Cercle des Entomologistes Liégeois, Liège, Belgique
13GxABT : Gembloux Agro-Bio Tech (ULg), Liège, Belgique
14HEPH : Haute École Provinciale de Hainaut - Condorcet, Ath, Belgique
15IRSNB : Institut royal des Sciences naturelles de Belgique, Bruxelles, Belgique
16UMONS : Université de Mons, Mons, Belgique
17Portails d’encodage en ligne
18OBSBE : observations en provenance de Waarnemingen.be, le site Web de Natuurpunt et Stichting Natuurinformatie, ainsi que de Observations.be, le site Web de Natagora (http://observations.be/index.php). Ces données ne peuvent être reproduites sans autorisation.
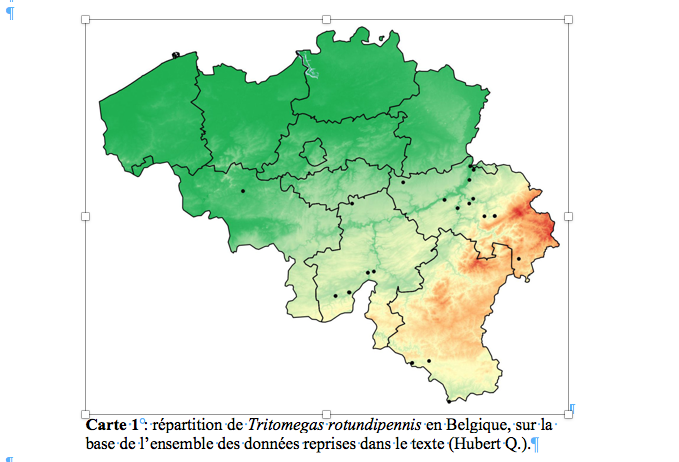
19OFFH : site Internet de l’Observatoire Faune – Flore – Habitats du Service Public de Wallonie (http://observatoire.biodiversite.wallonie.be/encodage/). L’origine de l’information provient du SPW-DGO3-DEMNA.
20L’ensemble des données provenant de ces deux portails ont été fournies à titre gracieux.
21Autres
22CEBE : site Internet de la CEBE-MOB (Commission de l'Environnement de Bruxelles et Environs asbl - Milieu Commissie Brussel en Omgeving vzw)
23PENTABEL : site Internet du forum de discussion ayant trait aux punaises de Belgique, géré par les CNB (Cercles des Naturalistes de Belgique)
24Stade des individus mentionnés
25ad. : imago ; juv. : juvénile ; ind. : exemplaire de stade indéterminé
Résultats
26Nous ne détaillerons ici que les données concernant T. rotundipennis, nouvelle espèce pour notre faune, et T. sexmaculatus, découverte chez nous assez récemment. Les données sur T. bicolor, beaucoup trop nombreuses, peuvent être consultées auprès des auteurs.
27Tritomegas rotundipennis (Dohrn, 1862)
28Région wallonne
29Hainaut : Ath, v.1985, 1 femelle, leg. inconnu (HEPH), 7.iv.2011, 1 mâle, Gillet Genin C. leg. (HEPH).
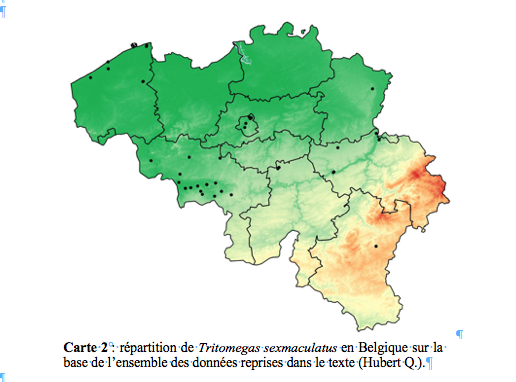
30Liège : Cheratte (Visé), iv.1904, 1 mâle (IRSNB) ; Crombach (Saint-Vith), 19.vii.1972, 1 femelle, Detry R. leg. (IRSNB) ; Esneux, 1.v.1997, 1 femelle, Gauthier leg., 28.vi.1999, 1 mâle, Hatert leg. (GxABT) ; Forêt (Trooz), 19.v.1918, 1 femelle, Lays P. det. (CEL) ; Ivoz-Ramet (Flémalle), 3.iv.2016, 2 ad., 15.iv.2016, 1 ad., Farinelle C. leg. (OBSBE) ; La Reid (Theux), 25.iv.1994, 1 mâle, Compère leg. (GxABT) ; Lixhe (Visé), 11.vi.1922, 1 femelle (IRSNB) ; Montagne Saint-Pierre (Liège), 7.v.1995, 1 mâle, Geerts leg. (GxABT) ; Péry (Trooz), 12.iv.1915, 1 mâle, Lays P. det. (CEL) ; Poucet (Hannut), 21.v.1995, 1 mâle, Évrard M. leg. (GxABT) ; Spa, 01.iv.2007, 2 ad., 31.iii.2008, 3 ad., 23.iii.2010, 5 ad., 21.iii.2011, 2 ad., 08.vi.2011, 1 juv. stade v, 19.iii.2012, 3 ad., 27.vi.2012, 1 juv. stade III, 02.vii.2012, 1 juv. stade V, 13.iv.2013, 6 ad., 26.iii.2016, 2 ad., 02.iv.2016, 1 ad., 03.iv.2016, 2 ad., Devillers C. leg. (PENTABEL).
31Luxembourg : Lacuisine (Florenville), 28.iv.1994, 1 mâle, Piron A. leg. (GxABT) ; Muno (Florenville), 27.iv.1994, 1 femelle, 29.iv.1994, 1 femelle, Warlomont leg., 28.iv.1994, 1 femelle, Collard leg. (GxABT) ; Torgny (Rouvroy), 8.iv.1943, 1 femelle, Mayné R. leg., vi.1947, 1 femelle, Paul B. leg. (IRSNB).
32Namur : Gembloux, 14.v.1995, 1 femelle, Jardin leg. (GxABT) ; Hermeton-sur-Meuse (Hastière), 20.v.1996, 1 femelle, Gardin leg. (GxABT), 25.iv.2014, 3 mâles, Limbourg P. leg. (IRSNB) ; Nismes (Viroinval), 14.iv.2013, 1 mâle, 15.iv.2016, 1 mâle, Claerebout S. leg. & coll. ; Treignes (Viroinval), 2.vi.2009, 1 femelle, 10.vi.2009, 1 femelle, 12.v.2016, 3 mâles et 2 femelles, Claerebout S. leg. & coll. ; Waulsort (Hastière), 15.iv.2010, 1 femelle, Huart leg. (GxABT).
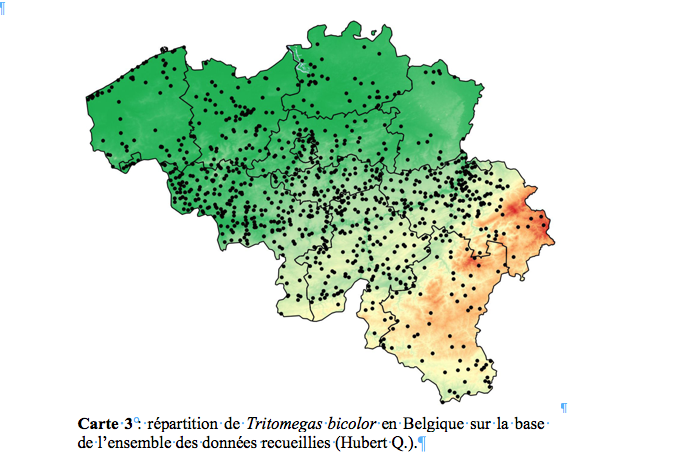
33La carte 1 montre la répartition de Tritomegas rotundipennis en Belgique sur base de l’ensemble des données reprises dans le texte ci-dessus. Dans l'état actuel de nos connaissances, l'espèce semble cantonnée dans le sud-est du pays.
34Tritomegas sexmaculatus (Rambur, 1839)
35Région Bruxelles-capitale
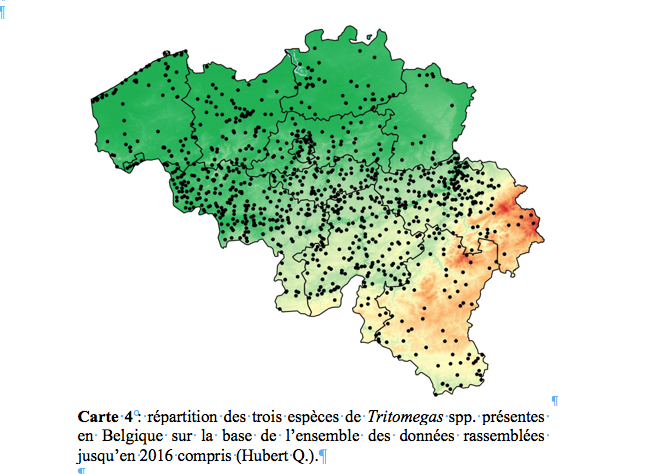
36Bruxelles, 16.viii.1994, 1 femelle, Dumortier leg. (GxABT) ; Evere-Moeraske, 17.viii.2006, 1 ind., 2.ix.2006, 1 ind., 24.vii.2007, 1 juv., 19.vii.2008, 3 juv., 26.vii.2009, 3 juv., 19.viii.2009, 2 ad., 11.ix.2010, 10 ad., 10.vii.2011, 1 ad., 20.v.2012, 1 ad., 2.vi.2013, 1 ad., 11.ix.2013, 1 ad., Hanssens B. leg. (OBSBE & CEBE) ; Forest (Uccle), 29.v.2014, 1 ad., Van den Neucker T. leg. (OBSBE) ; Neder-Over-Heembeek, 8.v.2016, 1 ad., Van Assche S. leg. (OBSBE).
37Région flamande
38Flandre occidentale : Beernem, 14.viii.2008, 2 ad., Libeer R. leg. (OBSBE), 15.vii.2015, 15 ad., Claerebout S. leg. (OBSBE) ; Knokke (Knokke-Heist), 27.v.2015, 1 ad., Wallays H. leg., 27.ix.2016, 1 ad., De Blauwe H. leg. (OBSBE) ; Nieuwpoort, 10.ix.2013, 1 ad., Lenaers C. leg. (OBSBE) ; Oostende, 24.v.2012, 1 ad., Decock W. leg. (OBSBE) ; Zandvoorde (Oostende), 24.ix.2013, 1 ad., Vanhoutte J. leg. (OBSBE).
39Limbourg : Opoeteren (Maaseik), 7.ix.2014, 1 ad., Paes J. leg. (OBSBE).
40Région wallonne
41Hainaut : Ath, 4.v.1995, 1 femelle, Runy P. leg. (HEPH) ; Boussu (Mons), 7.iv.1991, 1 femelle, Cheval B. leg. (IRSNB) ; Buvrinnes (Binche), 20.iv.1985 (HEPH) ; Estinnes-au-Val, 25.iv.1998, 1 femelle, Fromont leg. (GxABT) ; Frameries, 21.iv.2000, 1 mâle, inconnu leg. (GxABT), 24.iv.2002, 1 mâle, Benini leg. (GxABT) ; Genly (Quévy), 27.v.1995, 1 mâle, Cordier leg. (GxABT) ; Harchies (Bernissart), 11.x.2015, 1 ad., Lemaire M. & Lorin É. leg. (OBSBE) ; Havré (Mons), 10.v.2005, 1 femelle, Schoonbroodt leg. (GxABT) ; Mons, x.1984, 1 femelle (HEPH), 18.viii.1992, 1 mâle, 4.v.1992, 1 mâle, Gulino I. leg. (IRSNB) ; Petit-Roeulx-lez-Braine (Braine-le-Comte), 6.v.2000, 1 femelle (GxABT) ; Quaregnon, 23.v.1994, 1 femelle, Brohet A. leg. (UMONS) ; Quevaucamps, 4.vi.2015, 1 ad., Lemaire M. & Lorin É. leg. (OBSBE) ; Thulin (Hensies), 13.v.2001, 1 femelle, De Bie leg. (GxABT) ; Tournai, 18.v.1996, 1 mâle, Fontaine G. leg. (GxABT).
42Liège : Bleret (Waremme), 12.v.2008, 1 femelle, Noël leg. (GxABT) ; Huy, 5.vii.2011, 1 ad., Baugnée J.-Y. leg. (OFFH), 26.viii.2015, 10 ad., 28.viii.2015, 20 ad. et 30 juv., Vanmeerbeeck P. leg. (OBSBE) ; Montagne Saint-Pierre (Liège), 30.viii.2007, 1 mâle et 2 femelles, Aukema B. leg. (IRSNB), 12.viii.2007, 1 mâle, Aukema B. leg. (GxABT) ; Thier de Lanaye (Visé), 29.viii.2016, 1 ad., Boux K. leg. (OBSBE).
43Luxembourg : Givry (Bertogne), 8.v.2001, 1 femelle, De Bie leg. (GxABT).
44Namur : Gembloux, 2.vi.1996, 1 mâle et 1 femelle, Ghelici leg. (GxABT), 4.v.2000, 1 femelle, Vast leg. (GxABT), 17.x.2005, 1 femelle ex-larva, dans le jardin botanique, Baugnée J.-Y. leg. (GxABT) ; Grand-Manil (Gembloux), 29.v.1987, 1 mâle, Gernez leg. (GxABT).
45La carte 2 montre la répartition de Tritomegas sexmaculatus en Belgique sur base de l’ensemble des données reprises dans le texte ci-dessus. L'espèce est surtout présente dans le centre et l’ouest du pays.
46Tritomegas bicolor (Linnaeus, 1758)
47Pour cette espèce, nous avons recensé 1667 observations valides, qu’il serait fastidieux de détailler ici. Cette espèce a été signalée pour la première fois en Belgique à partir d’une femelle capturée le 10 août 1870 à Hollogne-sur-Geer (prov. de Liège).
48La carte 3 montre que Tritomegas bicolor est répandu dans toute la Belgique. Plus loin, nous esquissons l’évolution des populations de cette espèce dans notre pays.
Analyse des données et discussion
49Pour la Belgique, nous disposons donc d’un total de 1766 occurrences valides pour les trois espèces de Tritomegas confondues, majoritairement au stade adulte (1713 données – 96,99 %) et, dans une bien moindre mesure, au stade juvénile (53 données – 3,01 %). Les sources des données sont largement dominées par des spécimens de collection (1377 spécimens – 77,97 %), suivies par des prises de vues (366 photographies – 20,72 %) et enfin, de manière anecdotique, des citations dans la littérature spécialisée (23 données non redondantes avec les autres sources – 1,31 %).
50Le nombre total d’occurrences sur le territoire belge est inégal d’une région à une autre et d’une province à une autre : 77 % des observations ont été faites en Région wallonne, contre 20 % et 3 %, respectivement, pour les Régions flamande et Bruxelles-Capitale. Les provinces de Hainaut (444 données – 25 %), de Namur (395 données – 22 %) et de Liège (319 données – 18 %) réunissent à elles trois plus de la moitié des occurrences ; le solde étant partagé plus ou moins équitablement entre les autres provinces, entre 2 et 7 %.
51Comme le montre la carte 4, la couverture spatiale de l’ensemble des trois espèces de Tritomegas à l’échelle du pays est excellente.
52Plus de 60 % du matériel entomologique qui a permis d’établir ces cartes de répartition proviennent des récoltes d’étudiants de l’Agro-Bio Tech Gembloux (ULg) et de la Haute École Provinciale de Hainaut - Condorcet (954 et 171 occurrences sur 1766), qui réalisent leurs collections essentiellement en Région wallonne et au printemps. On pourrait donc craindre que cela n’introduise un biais dans nos données. Mais les données provenant d’autres sources (autres collections, portails d’encodage, littérature…) confirment ce que montrent déjà celles découlant des collections estudiantines.
53La méthodologie proposée par Stroot & Depiereux (1989) qui vise à établir des listes rouges d’invertébrés menacés, a été appliquée à nos chiffres. Cette démarche consiste à comparer les occurrences observées et théoriques (ou attendues) d’une espèce avant et après une date donnée et à calculer ensuite un indice en tenant compte de l’évolution globale de l’échantillonnage (toutes les espèces du groupe confondues) au cours de la période considérée. Cet indice sera d’autant plus élevé (c’est-à-dire significatif) que la proportion d’occurrence « avant » et « à partir de » pour une espèce donnée s’écartera de la proportion « avant » et « à partir de » globale.
54Tritomegas rotundipennis (Dohrn, 1862)
55Nous disposons d’un total de 44 observations validées, provenant toutes de la Région wallonne. Les nombres d’observations sont très variables d’une province à l’autre ; les plus élevés proviennent des provinces de Liège (24) et de Namur (12), les moins élevés celles de Luxembourg (6) et de Hainaut (2). Le nombe de stations différentes est de 20.
56La présence de T. rotundipennis a été décelée pour la première fois en Belgique en 2016 sur la base de photographies d’adultes et de juvéniles au stade V prises dans un jardin privé de la région de Spa (prov. de Liège). C’est suite à la parution d’un guide d’identification des punaises Pentatomoidea de France (Lupoli & Dusoulier, 2015) que le deuxième auteur de cette note a eu l’idée de réexaminer ses anciennes observations depuis avril 2007 jusqu’à ce jour et a ainsi confirmé la présence de T. rotundipennis. Des recherches ultérieures ont permis de signaler d’autres stations (19) et occurrences (44). La majorité des mentions proviennent des environs de Liège. La plus ancienne donnée pour le pays date de 1904. La présence de T. rotundipennis en Belgique est donc passée inaperçue pendant plus d’un siècle, probablement en raison de sa ressemblance avec T. bicolor. Son installation chez nous, en particulier dans l’est et le centre du pays (carte 1), est à présent bien établie. Des recherches plus poussées montreraient sans doute qu’elle est encore plus répandue que ne le montre cet article.
57En dehors de la Belgique, T. rotundipennis n’est renseignée que de six pays européens : Allemagne (Heckmann & Rieger, 2001), Autriche (Rabitsch, 2001), Espagne (Catalogne) (Goula et al., 2010 ; Ribes, 1978 et 1989 ; Ribes et al., 2004), France continentale (Lupoli et al. 2014), Italie continentale où elle semble s’étendre à l’intérieur du pays, et Sicile (Dohrn, 1862 ; Horváth, 1917 ; Servadei, 1967 ; Stoch, 2003) ainsi que de Slovénie (Gogala & Gogala, 1989). Ces deux derniers auteurs ont interprété les observations slovènes réalisées entre 1982et 1989 comme la conséquence de l’augmentation de l’aire de distribution de T. rotundipennis depuis une zone contiguë du nord de l’Italie, située à seulement 50 km de distance. Il est intéressant de noter une observation située à quelques kilomètres à peine de notre frontière avec la France, à proximité du cimetière de Marville (département de la Meuse), concernant un individu mâle trouvé le 19 avril 1996 (leg. et coll. Baugnée J.-Y.).
58À notre connaissance, en Belgique, la seule plante hôte renseignée est le lamier jaune (Lamium galeobdolon subsp. galeobdolon) ainsi que l’une de ses sous-espèces, le lamier argenté (Lamium galeobdolon subsp. argentatum). Cependant, à partir d’observations réalisées dans la nature et en élevage, le lamier blanc (Lamium album L., 1753) permet également la réalisation complète du cycle de développement (Heckmann & Rieger, 2001 ; Lupoli et al., 2014 ; Ribes, 1989 ; Wachmann et al., 2008). Nous avons trouvé T. rotundipennis dans les parties ombragées et humides de jardins privés, de lisières forestières ou de pelouses calcicoles, comme cela a déjà été mentionné par Mazuy (2008). Souvent, T. rotundipennis est accompagnée de T. bicolor. Un tel comportement avait déjà été signalé par Gogala & Hočevar (1990), ainsi que par Lupoli et al. (2014) qui ont pu observer les trois espèces de Tritomegas, au même moment, sur un même massif de lamier d’à peine 100 m2. Il nous semble que le repérage des insectes à l’œil nu est idéal lorsqu’ils se maintiennent sur les parties supérieures de leur plante hôte, particulièrement lorsque les rayons du soleil les atteignent directement. Ils s’y trouvent régulièrement lorsque la température avoisine les 10 à 15 °C. Cependant, les individus sont très sensibles à la brusque apparition d’ombre et réagissent en se laissant tomber immédiatement.
59Les adultes se rencontrent pratiquement tout au long de l’année, avec un pic au début du printemps (fin mars à mi-mai, graphique 1), pour laisser place petit à petit aux juvéniles aux mois de juin et juillet, qui donneront des imagos en fin de saison, lesquels hiberneront.
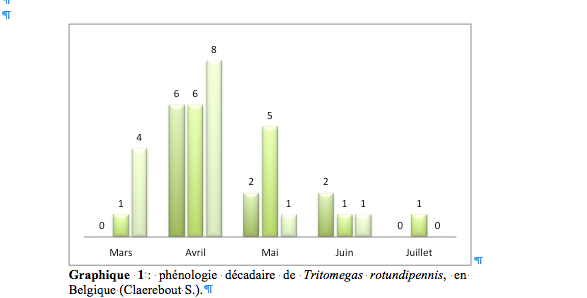
60Tritomegas sexmaculatus (Rambur, 1839)
61Nous disposons d’un total de 55 observations validées, provenant des Régions Bruxelles-Capitale, flamande et wallonne (respectivement 14, 9 et 32). Les nombres d’observations sont très variables d’une province à l’autre. En Région flamande, T. sexmaculatus est rencontré uniquement dans les provinces de Flandre occidentale et de Limbourg avec, respectivement, 8 et 1 observation(s). En Région wallonne, les observations les plus nombreuses s’effectuent dans la province de Hainaut (17), suivie des provinces de Liège (9) et de Namur (5). Le nombre de localités différentes est de 35.
62T. sexmaculatus a été découverte et identifiée officiellement pour la première fois en Belgique, le 12 août 2007, à la Montagne Saint-Pierre (Visé, prov. de Liège) (Aukema et al., 2007). Cependant, avant 2000, une population de Tritomegas, identifiée alors comme appartenant à l’espèce T. bicolor, était signalée au Moeraske à Bruxelles par Carlet G. et par les membres de la CEBE. Cette population fut observée et photographiée exactement au même endroit d’année en année (ix.2005, 17.viii.2006, 24.vii.2007 et 19.vii.2008) jusqu’à ce qu’elle soit correctement identifiée en tant que T. sexmaculatus fin de l’année 2008 par Baugnée J.-Y. (Coppée & Hanssens, 2006 ; Doornaert, 2007). Plus tard, Dethier & Steckx (2010) mentionneront sept données chorologiques inédites pour la Belgique. Tout récemment, la révision des collections a révélé l’existence de plusieurs spécimens plus anciens encore, dont le tout premier connu est une femelle provenant des environs de Mons, en 1984. À partir de 2011, les observations se multiplient, suggérant que cette espèce est bien installée chez nous, probablement depuis assez longtemps.
63T. sexmaculatus une espèce à tendance méridionale, connue également d'Europe centrale et orientale, mais où elle s’y fait plus rare. Elle est renseignée de nombreux pays, à l’exception des plus nordiques : Albanie, Allemagne, Autriche, Bosnie-et-Herzégovine, Bulgarie, Croatie, Danemark, Espagne, France et Corse, Géorgie, Grèce, Hongrie, Italie continentale (y compris Sicile et Sardaigne), Liechtenstein, Macédoine, Moldavie, Monténégro, Pays-Bas, Pologne, Portugal, Roumanie, Royaume-Uni, Russie (centre et sud des territoires européens), Serbie, Slovaquie, Slovénie, Suisse, Tchéquie, Turquie et Ukraine. Son aire de répartition s’étend aussi vers l’Asie : Azerbaïdjan, Arménie, Géorgie et Iran (Aukema B. & Rieger, 2006 ; Aukema et al., 2013).
64À proximité de nos frontières, en France, les données les plus septentrionales mentionnées jusqu’au début des années 2000, l’ont toujours été de la région parisienne, où elle y est bien implantée (Denosmaison, 2001). Mais depuis, d’autres localités encore plus au nord ont été découvertes, à Wimereux au sud-ouest de Calais (département du Pas-de-Calais) et aux environs de Reims (département de la Marne) (Lupoli & Dusoulier, 2015). Aux Pays-Bas, depuis son premier signalement en août 2002 à Het Zwindorp (Sluis, prov. de Zélande) (Aukema, 2003), elle a repoussé ses limites plus au nord jusqu’à Gassel (Grave, prov. du Brabant-Septentrional) et plus à l’est jusqu’à Venray (prov. de Limbourg). En ce qui concerne les populations allemandes, les plus proches connues de nos frontières sont celles de la région de Cologne (Rhénanie du Nord-Westphalie) (Aukema, 2003). Cette espèce n’est pas encore mentionnée du Grand-Duché de Luxembourg (Reichling, 2001).
65La ballote fétide (Ballota nigra subsp. meridionalis) est mentionnée comme la plante hôte préférentielle de T. sexmaculatus (Aukema, 2003 ; Bantock, 2011 ; Bunalski et al., 2010 ; Heiss, 1977 ; Wachmann et al., 2008 ; Werner, 2010). T. sexmaculatus ne se rencontre qu’exceptionnellement sur d’autres Lamiacées. Néanmoins, d’autres observateurs et nous-mêmes avons eu l’occasion d’observer, tant des imagos que des juvéniles, en grand nombre, sur le marrube (Marrubium vulgare L., 1753) (Aukema et al., 2007 ; Drost & Cuppen, 2008). Comme autres plantes hôtes évoquées dans la littérature, on retrouve la carotte (Daucus carota L., 1753) (Stichel, 1961), la menthe à longues feuilles (Mentha longifolia L., 1756), le marrube voyageur (Marrubium peregrinum L., 1753), diverses espèces de lamiers (Lamium spp.), l’ortie dioïque (Urtica dioica L., 1753) et la pariétaire officinale (Parietaria officinalis L., 1753) (Bunalski et al., 2010 ; Lis & Ziaja, 2008 et 2009).
66T. sexmaculatus est considérée comme une espèce sténotherme, incapable de supporter de grandes amplitudes thermiques. C’est une espèce plus thermophile que T. bicolor et qui évite apparemment les milieux les plus humides (Wachmann et al., 2008 ; Werner, 2010). T. sexmaculatus est donc susceptible de coloniser toutes sortes d’habitats ouverts, plutôt secs et sous un climat chaud, comme les bords de route, les voies de chemin de fer, les friches industrielles, les digues, les murs de pierre, les haies, les jardins, les villes, etc. (Aukema, 2003 ; Bantock, 2011 ; Lis & Ziaja, 2008). En outre, T. sexmaculatus serait affectée, plus que d’autres, par les changements climatiques récents, ce qui se traduirait par une remontée vers le nord des limites de son aire de distribution. C’est ce phénomène qui expliquerait, en Pologne, sa réapparition après cinquante ans d’absence (Lis & Ziaja, 2009) et le déplacement de cette limite à plus de 200 km vers le nord-ouest par rapport aux plus anciennes données polonaises, atteignant 52 degrés de latitude nord aux environs de « Pomeranian Lakeland » (Bunalski et al., 2010 ; Ziaja, 2013). L’explication se trouverait dans un changement de sa biologie, montrant un allongement du cycle de vie qui permettrait aussi au stade juvénile d’hiverner (pas seulement les adultes), fait tout à fait inhabituel pour les représentants européens de cette famille d’insectes (Lis & Ziaja, 2009). En Allemagne, à partir des années nonante, Werner (2010) constate une augmentation d’occurrences pour l’espèce, parfois même plus au nord que ce qui avait été observé jusque-là. La cause de ce phénomène serait également imputée aux changements climatiques récents. En Belgique, on constate aussi une augmentation significative des occurrences depuis 1990 et surtout depuis 2000.
67T. sexmaculatus est univoltine. Sa phénologie (graphique 2) est similaire à celle des autres Tritomegas, à ceci près que la période de ponte et le développement larvaire s’étalent sur une plus longue période, permettant de rencontrer des stades juvéniles plus tardivement dans la saison, jusqu’au mois de septembre ou même, à titre exceptionnel, fin octobre (Lis & Ziaja, 2008 ; Wachmann et al., 2008). Après avoir passé l’hiver au stade imaginal, les individus s’accouplent et les pontes sont déposées de mai à juin (Aukema, 2003). Des recherches récentes suggèrent qu’au sein de nombreuses populations, lors de conditions climatiques favorables, une partie importante des individus juvéniles hivernent (Kolak, 2015).
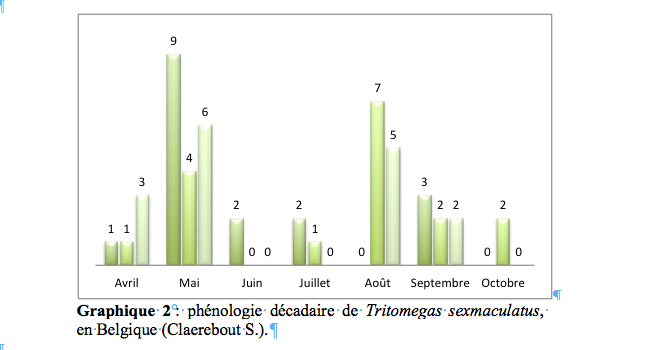
68Tritomegas bicolor (Linnaeus, 1758)
69Nous disposons d’un total de 1667 observations validées, provenant des Régions Bruxelles-Capitale, flamande et wallonne (respectivement 35, 352, et 1280). Les nombres d’observations sont très variables d’une province à l’autre. Les observations les plus nombreuses ont été faites dans les provinces de Hainaut (425), de Namur (378) et de Liège (286), totalisant 65 % des données. La Région Bruxelles-Capitale (35) et les provinces de Limbourg (26) et d’Anvers (61) sont les moins bien représentées.
70Dès 1879, Lethierry & Pierret et, peu après, Coubeaux (1891) et Lethierry (1892) signalent déjà la présence de T. bicolor sur l’ensemble du territoire national. Leurs observations seront suivies et confirmées par de nombreux hémiptérologues belges plus ou moins contemporains (Bosmans, 1975 ; Chérot, 1991 ; Dethier & Chérot, 1997 ; Kiriakoff, 1962 ; Mayné, 1965 ; Petit, 1994). Aujourd’hui encore, il s’agit de l’espèce du genre la plus répandue, la plus commune et la plus abondante chez nous.
71Au-delà de nos frontières, au sein du genre Tritomegas, T. bicolor est aussi le représentant le plus largement distribué mondialement (Aukema & Rieger, 2006). En Europe, cette espèce est renseignée de 35 pays. En Afrique du Nord, elle est présente en Algérie et au Maroc. En Asie, elle a également une très vaste répartition : Arménie, Azerbaïdjan, Chine (Liaoning), Corée, Géorgie, Iran, Israël, Kazakhstan, Kirghizstan, Mongolie, Ouzbékistan, Russie, Sibérie, Syrie, Turkestan et Turquie (Aukema & Rieger, 2006 ; Aukema et al., 2013).
72T. bicolor est régulièrement récoltée sur les Lamium spp. et, plus occasionnellement, sur les Stachys spp. (Bogacz & Tolsgaard, 2010 ; Ha1ászfy, 1954). Elle s’alimente également sur les boutons floraux ou les inflorescences de la ballote fétide (Ballota nigra subsp. meridionalis) (Bunalski et al., 2010 ; Petit, 1994). Une observation a été réalisée sur une plante ornementale appartenant à une autre famille, la lobélie érine (Lobelia erinus L., 1753), une Campanulacée (Petit, 1994). Bien que nous n’ayons jamais observé cette espèce ni sur des plantes potagères ni sur des arbres fruitiers, d’après Stichel (1961), les imagos se tiennent sur les inflorescences des prunelliers (Prunus spinosa L.,1753) et des arbres fruitiers à noyau en général, ainsi que sur les aubépines (Crataegus spp.). D'après cet auteur elle serait nuisible aux baies et à certains légumes. Perrier (1935) reprend les indications de Marchal (1898), Lameere (1900) et Lambertie (1908), observant cet insecte, souvent en groupes sur les racines des plantes potagères, et même sur les jeunes pousses des arbres fruitiers, mais alors isolé.
73Les habitats pouvant accueillir T. bicolor sont donc très nombreux : haies, lisières forestières, forêts clairsemées, accotements de voiries, jardins, friches, partout là où peuvent se retrouver ses plantes hôtes (Wachmann, 2008).
74Les imagos de T. bicolor hivernent au stade adulte, comme cela a déjà été observé de longue date (Horváth, 1892). Ils se retrouvent dans le sol, parfois à une profondeur de quinze centimètres, mais habituellement sous les mousses. Ils réapparaissent en mars-avril (graphique 3), migrant parfois vers de nombreuses plantes y compris vers les arbres fruitiers qu’ils ponctionnent notamment pour se réhydrater. Le pic d’abondance survient un mois plus tard, donnant lieu à de nombreux accouplements jusqu’en juillet. Au moment de la ponte, la femelle creuse une petite dépression, dans laquelle elle dépose une quarantaine d’œufs. Une vingtaine de jours plus tard, l’éclosion se produit (Southwood & Leston, 2005). Les œufs et les premiers stades juvéniles bénéficient de soins maternels, comportement fréquent chez les Cydnidae, qui impliquent essentiellement le transfert de bactéries symbiotiques vers la progéniture et la défense contre des parasitoïdes (Werner, 2010). En général, le stade juvénile dure six semaines. À la fin du mois d’août la nouvelle génération d’adultes apparaît et regagne assez rapidement les sites d’hivernage, formant parfois des agrégats.
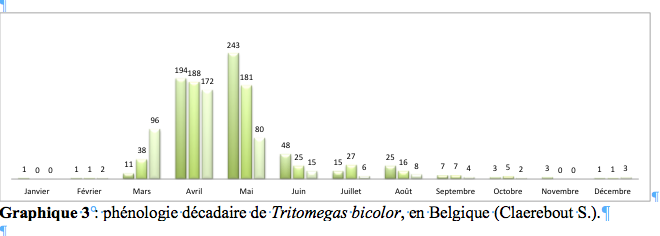
75Il n’est pas rare de rencontrer des Hétéroptères, appartenant notamment à la superfamille des Pentatomoidea, qui portent un ou plusieurs minuscules œufs blancs collés le plus souvent sur la tête, le pronotum ou les cories. Ces œufs ont été pondus par des tachinaires (Diptera: Tachinidae: Phasiinae), parasitant spécialement les Hétéroptères (Tschorsnig & Richter, 1998). Phasia pusilla Meigen, 1824 semble parasiter exclusivement les Cydnidae, tandis que, selon Herting (1971), Clytiomya continua (Panzer, 1798) serait un parasitoïde spécifique de T. sexmaculatus. Tschorsnig & Herting (1994) mentionnent Eliozeta pellucens (Fallén, 1820) et Phania incrassata Pandellé, 1894 comme parasitoïdes de T. bicolor. Toutes ces espèces de Tachinidae sont connues de Belgique (Gosseries & Wyatt, 1991) et la dernière a été signalée sur T. bicolor par Van de Weyer & Zeegers (2002). Jusqu’à présent, aucun parasitoïde spécifique de T. rotundipennis n’a été signalé.
Conclusions
76Tritomegas rotundipennis n’a été signalée que tout récemment dans notre pays. Cependant, la révision de la littérature et des collections a montré que cette espèce était présente chez nous depuis 1904 au moins. En raison de sa répartition dans toute la partie orientale du pays et de sa présence dans des milieux assez variés, il n’est pas interdit de penser que cette espèce est plus fréquente qu’il n’y paraît et que sa présence en Belgique est déjà ancienne et n’est pas nécessairement liée aux récentes modifications climatiques. Sa découverte tardive s’explique sans doute mieux par sa ressemblance avec T. bicolor, espèce omniprésente chez nous. Il faut néanmoins noter que la plupart des occurrences sont assez récentes, comme d’ailleurs en France (Lupoli et al., 2014).
77Le plus ancien spécimen connu de T. sexmaculatus en Belgique a été capturé en 1984. Cette espèce thermophile semble donc montrer une expansion récente, peut-être liée au réchauffement climatique. Cette impression est confirmée par l’application à nos chiffres de la méthodologie proposée par Stroot & Depiereux (1989).
78Les résultats des calculs montrent que seule l’espèce T. sexmaculatus présente un indice significatif et que l’on peut conclure à son expansion récente. Pour les deux autres espèces, tant avec la date-pivot 1950 qu’avec 1980, les indices sont beaucoup plus faibles et la tendance à une éventuelle expansion est moins nette. Les différences d’occurrences observées, parfois considérables comme chez T. bicolor, peuvent être attribuées à des efforts de capture plus intenses au cours de ces deux ou trois dernières décennies.
Remerciements
79MM. P. Lays (ULg) et M. Matthys ont rassemblé et envoyé aux auteurs les données belges de Tritomegas spp. provenant des collections entomologiques, respectivement, du Musée de l’Institut de Zoologie (Université de Liège) et du Cercle des Entomologistes Liégeois (C. E. L.), ainsi que de la Station scientifique des Hautes Fagnes (ULg, SSHF).
80Mme J. Bortels (GxABT) et MM. J. Constant (IRSNB), J.-B. Dutrifoy (HEPH - Condorcet), P. Rasmont et D. Évrard (UMONS), ainsi que J. Fagot (HEPL-La Reid) ont mis à disposition les spécimens belges de Tritomegas spp. présents dans les collections dont ils ont ou avaient la charge.
81M. Y. Barbier (SPW, DGO3, DEMNA) fournissant aux auteurs à titre gracieux les données de l’OFFH.
82MM. C. Farinelle et J.-Y. Baugnée signalant leurs observations personnelles aux auteurs, alors que de nombreux autres observateurs mentionnés dans le texte choisissaient de les rendre publiques sur le portail Observations.be/Waarnemingen.be.
83MM. K. Lock et R. Libeer pour leur travail de validation sur les deux portails d’encodage en ligne.
84MM. B. Aukema (Renkum, Pays-Bas) et D. J. Werner (Köln, Allemagne), ainsi que M. J. Bruers et Mme G. Viskens (Anvers, Belgique) ont transmis au premier auteur plusieurs de leurs travaux cités dans le présent article.
85Mme M. Van Assche (IRSNB, Belgique) a transmis aux auteurs de nombreux articles permettant d’améliorer le contenu de cette note.
86M. Q. Hubert (Centre Marie-Victorin, Cercles des Naturalistes de Belgique) a réalisé les cartes de répartition.
87M. S. Wullaert a bien voulu traduire le texte introductif en néerlandais.
88Enfin, le personnel du secrétariat de la Haute École Provinciale de Hainaut – Condorcet qui a permis de retrouver les auteurs de certaines collectes estudiantines intéressantes.
89Que toutes et tous en soient chaleureusement remerciés, ainsi que les 1074 personnes ayant transmis leurs données d’une manière ou d’une autre, sans lesquelles ce travail n’aurait pas pu aboutir.
Bibliographie
90Alderweireldt M., 2001. Enkele waarnemingen van landwantsen (Hemiptera) in de Blankaart (Woumen, West-Vlaanderen). Phegea,29(3): 89-94.
91Aukema B., 2003. Wantsennieuws uit Zeeland (Heteroptera). Nederlandse Faunistische Mededelingen,18: 1-16
92Aukema B., Bruers J. M. & Viskens G. M., 2007. Nieuwe en zeldzame Belgische wantsen II (Hemiptera: Heteroptera). Bulletin S.R.B.E./K.B.V.E., 143: 83-91.
93Aukema B., Bruers J. M. & Viskens G. M., 2014. Geotomus petiti Wagner, 1954, a new burrower bug for Belgium (Heteroptera: Cydnidae). Andrias,20: 15-19.
94Aukema B., Rieger C. & Rabitsch W., 2013. Catalogue of the Heteroptera of the Palaearctic Region. Vol. 6. Supplement. The Netherlands Entomological Society. 629 p.
95Aukema B. & Rieger C., 2006. Catalogue of the Heteroptera of the Palaearctic Region. Vol. 5. Pentatomomorpha II. The Netherlands Entomological Society. 550 p.
96Baugnée J.-Y., Dethier M., Bruers J., Chérot F. & Viskens G., 2003. Liste des punaises de Belgique (Hemiptera Heteroptera). Bulletin S.R.B.E./K.B.V.E., 139: 41-60.
97Bantock T., 2011. Tritomegas sexmaculatus (Cydnidae) arrives in Britain. Het News, 17/18: 4.
98Bogacz D. & Tolsgaard S., 2010. A study on the bug fauna from different Danish landscapes.Natural History Museum Aarhus. 22 p.
99Bosmans R., 1975. Voorkomen van de belgische wantsen. 1. Biologisch Jaarboek Dodonaea, 43: 78-89.
100Bunalski M., Korcz A. & Sienkiewicz P., 2010. Pluskwiaki różnoskrzydłe (Hemiptera: Heteroptera) północno-zachodniej Polski. 1. Rozmieszczenie gatunków z rodzaju Tritomegas Amyot et Serville, 1843 (Cydnidae). [True bugs (Hemiptera: Heteroptera) of north-western Poland. 1. The distribution of Tritomegas AMYOT et SERVILLE, 1843 (Cydnidae) species]. Wiad. entomol., 29(1): 5-13.
101Bruers J. & Viskens G., 2002. De verzameling Heteroptera van Atalanta. Entomo-Info,13(1): 17-27.
102Bruge H., 1995. Observations entomologiques réalisées en Belgique, au cours de l’été 1995. Accompagnées de quelques informations et commentaires « pédagogiques », spécialement destinés aux entomologistes amateurs. Bulletin et Annales de la Société royale belge d’Entomologie,131: 463-475.
103CEBE-MOB, 2016. www.cebe.be, consulté le 5.XII.2016
104Charlot M.-È. & Claerebout S., 2016. Clé de détermination photographique des Pentatomoidea de Belgique et des régions voisines. Éd. Cercles des Naturalistes de Belgique, 112 p.
105Chérot F., 1991. Les Hémiptères Hétéroptères de la Montagne Saint-Pierre et ses environs immédiats : liste préliminaire et considérations diverses (Hemiptera Heteroptera). Bulletin et Annales de la Société royale belge d’Entomologie,127(1-4): 33-45.
106Claerebout S., 2011. Conquête de l’Europe par une punaise nord-américaine invasive : la Punaise américaine des pins (Leptoglossus occidentalis Heidemann, 1910). L’Érable, 35(1): 14-21.
107Coppée J.-P. & Hanssens B., 2006. Observations Automne 2006 - Waarnemingen Najaar 2006 [01/09/2006 - 28/11/2006]. Écho du Marais,80: 19-26.
108Coubeaux E., 1891. Énumérations des Hémiptères de Belgique. I. Hémiptères Hétéroptères. Annales de la Société entomologique de Belgique,35: 388-395.
109Denosmaison J.-C., 2001. Hétéroptères nouveaux pour la région parisienne. L’Entomologiste,57(2): 84.
110Dethier M. & Chérot F., 1997. Nouvelles recherches sur les Hétéroptères de la Montagne Saint Pierre et note sur les Globiceps (Miridae). Bulletin et Annales de la Sociétéroyale belge d’Entomologie,133: 241-266.
111Dethier M. & Steckx É., 2010. Note sur quelques Hétéroptères intéressants pour la faune de Belgique. Natura Mosana,63(1): 9-15.
112Dohrn A., 1862. Drei neue europäische Heteroptera. Entomologischen Vereine zu Stettin, 23: 210-211.
113Doornaert A., 2007. La détermination des arthropodes, quelques réflexions.Écho du Marais,81: 18-19.
114Drost B. & Cuppen J. G. M., 2008. Entomofauna van Zuid-Limburg. Verslag van de 162e zomerbijeenkomst te Mechelen. Entomologische Berichten,68(4): 130-153
115Froeschner R. C., 1960. Cydnidae of the Western Hemisphere. Proceedings of the United States National Museum, 111: 337-680.
116Gogala A. & Gogala M., 1989. True bugs of Slovenia II (Insecta: Heteroptera). Biološki vestnik,37(1): 11-44.
117Gogala M. & Hočevar I., 1990. Vibrational songs in three sympatric species of Tritomegas. ScopoliaSuppl. 1: 117-123.
118Gosseries J. & Wyatt N.,1991. Tachinidae, p. 199-208. In: Grootaert P., De Bruyn L. & De Meyer M. (eds.). Catalogue of the Diptera of Belgium. Studiedocument van het KBIN, 70: 1-338.
119Goula M., Ribes J. & Serra A., 2010. Llista dels Heteròpters de Catalunya (Insecta, Hemiptera, Heteroptera). Versió 1, juny 2010. [Checklist of Heteroptera of Catalonia (Insecta, Hemiptera, Heteroptera)]. Centre de Recursos de Biodiversitat Animal, Facultat de Biologia, Universitat de Barcelona. 38 p.
120Halászfy Cs. E., 1954. A Synopsis of the Heteroptera of Hungary and the Neighbouring Areas. I. 1. Brachyplatidae ; 2. Cydnidae. Annales Historico-Naturales Musei Nationalis Hungarici,4:187-195.
121Heckmann R. & Rieger C., 2001. Wanzen aus Baden-Württemberg – Ein Beitrag zur Faunistik und Ökologie der Wanzen in Baden-Württemberg (Insecta, Heteroptera). Carolinea,59: 81-98.
122Heiss E., 1977. Zur Heteropterenfauna Nordtirols (Insecta: Heteroptera). VI. Pentatomoidea. Veröff. Mus. Ferd. Innsbruck, 57: 53-77.
123Herting B., 1971. A catalog of parasites and predators of terrestrial arthropods. Section A : Host or prey/enemy. Vol. I. Arachnida to Heteroptera. Commonwealth Agricultural Bureaux; Slough, England. 129 p.
124Horváth G., 1892. Chasses hivernales dans le midi de la France. Revue d’Entomologie,11:128-140.
125Horváth G., 1917. Heteroptera Palaearctica Nova vel Minus cognita. I. Annales Musei Nationalis Hungarici, 15: 365-381.
126Jordan K. H. C., 1958. Lautäußerungen bei den Hemipteren-Familien der Cydnidae, Pentatomidae und Acanthosomidae. Zoologischer Anzeiger, 161(5-6): 130-144.
127Kammerschen D., 1986a. Tritomegas rotundipennis (Dohrn, 1862) im Elsaß - ein zoogeographisch bemerkenswertes Vorkommen. Mit einer ergänzenden Beschreibung dieser und der beiden anderen mitteleuropäischen Tritomegas Taxa (Het.: Cydnidae). Marburger Entomologische Publikationen,2(3): 1-38.
128Kammerschen D., 1986b. Species-discrimination and geographic distribution in the cydnid genus Tritomegas (Heteroptera, Cydnidae). Proceedings of the 2nd International Congress concerning the Rhynchota Fauna of Balkan and Adjacent Regions, Mikrolimni - Greece : 33-34
129Kiriakoff S. G., 1962. De wantsen (Hemiptera Heteroptera) van de verzameling M. Goetghebuer. Biologisch Jaarboek Dodonaea, 30: 451-460.
130Kolak G., 2015. Nowe stanowisko Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) na Górnym Śląsku. Acta entomologica silesiana,23: 1-5.
131Kolak G. ,2015. Nowe stanowisko Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) na Górnym Śląsku. Acta entomologica silesiana,23: 1-5.
132Lambertie M., 1908. Notes sur les insectes nuisibles et utiles de l’Ordre des Hémiptères du sud-ouest de la France. Actes de la Société linnéenne de Bordeaux. Vol. LXII, Série 7, Tome 2 : 421-447.
133Lameere A., 1900. Manuel de la Faune de Belgique. Insectes inférieurs. Tome II. Lamertin, Bruxelles, 857 p.
134Lethierry L., 1892. Revue des Hémiptères de Belgique. Imprimerie Laroche-Delattre, Lille, 27 p.
135Lethierry L. & Pierret M., 1879. Premier essai d'un Catalogue des Hétéroptères de la Belgique. Annales de la Société entomologique de Belgique,22: 5-23.
136Leston D., 1954. Strigils and stridulation in Pentatomoidea (Hem.) : some new data and a review. The Entomologist’s monthly Magazine,90: 49-56.
137Lis J. A., 2006. Family Cydnidae Billberg, 1820 - burrowing bugs (burrower bugs), 119-147. In : Aukema B. & Rieger C. 2006. Catalogue of Heteroptera of the Palaearctic Region. Volume 5. Pentatomomorpha II. The Netherlands Entomological Society. 550 p.
138Lis J. A., Becker M. & Schaefer C. W., 2000. Chapter 12. Burrower Bugs (Cydnidae), p. 405-419. In : Schaefer C. W. & Panizzi A. R. Heteroptera of Economic Importance, CRC Press, 856 p.
139Lis J. A. & Ziaja D. J., 2008. Nowe dane o występowaniu i biologii Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) w Polsce. [New data on the occurrence and biologyof Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) in Poland].Nature Journal (Opole Scientific Society),41: 111-116.
140Lis J. A. & Ziaja D. J., 2009. Zmiany zasięgu Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) w Polsce efektem zmian klimatycznych ? [Changes in distribution range of Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) in Poland as a possible effect of climate changes ?].Nature Journal (Opole Scientific Society),42: 123-128.
141Lupoli R. & Dusoulier F., 2015. Les Punaises Pentatomoidea de France. Éd. Ancyrosoma, 429 p.
142Lupoli R., Derreumaux V. & Bertrand M., 2014. Nouvelles observations sur Tritomegas rotundipennis (Dohrn, 1862) (Hemiptera Cydnidae). L’Entomologiste,70(5): 257-262.
143Marchal C., 1898. Premières notes sur les Hémiptères de Saône-et-Loire. Bulletin de la Société d’histoire naturelle d’Autun,11: 557-593.
144Mayné R., 1965. Les Hémiptères de la Réserve domaniale du Westhoek. Pentatomoidea. Service des Réserves Naturelles domaniales et de la Conservation de la Nature, Travaux 1. 43 p.
145Mazuy C., 2008. Une rareté à Danjoutin ! CREN Infos,3: 1.
146Perrier R., 1935. La faune de la France illustrée. IV. Hémiptères, Anoploures, Mallophages et Lépidoptères. Librairie Delagrave, Paris, 243 p.
147Petit J., 1994. Quelques aspects peu connus du comportement d'un Hémiptère commun Tritomegas bicolor. Revue Verviétoise d’Histoire Naturelle,42: 43-45.
148Pluot-Sigwalt D. & Lis A. J., 2008. Morphology of the spermatheca in the Cydnidae (Hemiptera: Heteroptera) : Bearing of its diversity on classification and phylogeny. Eur. J. Entomol. , 105: 279-312.
149Rabitsch W., 2001. Notizen zur Wanzenfauna Österreichs (Insecta, Heteroptera). Linzer biologische Beiträge, 33(1): 83-86.
150Reichling L., 2001. Atlas des hétéroptères non-aquatiques du Luxembourg. Musée d'Histoire naturelle. Luxembourg. 134 p.
151Ribes J., 1978. Heteròpters nous o interessants per als Països Catalans. Bulletí de la Institució Catalana d’Historia Natural (Sec. Zool., 2), 42: 83-88.
152Ribes J., 1989. Miscellània Hemipterològica ibèrica (Heteroptera). Sessió Entomològica Institució Catala d’Història Natural. Societat Catalana de Lepidopterologia,6: 19-35.
153Ribes J., Serra A. & Goula M., 2004. Catàleg dels heteròpters de Catalunya (Insecta, Hemiptera, Heteroptera). [Catalog of Heteroptera of Catalonia (Insecta, Hemiptera, Heteroptera)]. Instotució Catalana d’Història Natural. Secció de Ciències Biològiques. Institut d'Estudis Catalans. Barcelona. 128 p.
154Rose L., 2000. Notions sur les Insectes Hémiptères avec relevés d’occurrences observées. Revue verviétoise d’Histoire naturelle,57: 141-160.
155Servadei A., 1967. Rhynchota (Heteroptera, Homoptera Auchenorrhyncha). Catalogo topografico e sinonimico. Fauna d'Italia, 9: 851. Bolonia
156Southwood T. R. & Leston D., 2005. Land and Water Bugs of the British Isles. Pisces Conservation. 440 p.
157Stichel W., 1961. Illustrierte Bestimmungstabellen der Wanzen. II. Europa (Hemiptera-Heteroptera Europae), 4: 667-698.
158Stoch F., 2003. FaunaItalia. http://www.faunaitalia.it/checklist/invertebrates/families/Cydnidae.html. Consulté le 30/11/2016
159Stroot Ph. & Depiereux E., 1989. Proposition d’une méthodologie pour établir des « Listes Rouges » d’Invertébrés menacés. Biological Conservation,48 : 163-179.
160Tschorsnig H.-P. & Herting B., 1994. Die Raupenfliegen (Diptera: Tachinidae) Mitteleuropas : Bestimmungstabellen und Angaben zur Verbreitung und Ökologie der einzelnen Arten. Stuttgarter Beiträge zur Naturkunde Serie A (Biologie),506: 1-170.
161Tschorsnig H.-P. & Richter V. A., 1998. Family Tachinidae. p. 691-827. In: Papp L. & Darvas B. (Eds.). Contributions to a Manual of Palaearctic Diptera (with special references to flies of economic importance). Volume 3. Higher Brachycera. Science Herald, Budapest. 880 p.
162Van de Weyer G. & Zeegers T., 2002. Aanvulling op de naamlijst van de Belgische sluipvliegen (Diptera: Tachinidae). Phegea,30(4): 193-199.
163Vidal J.-P. A., 1949. Hémiptères de l'Afrique du Nord et des Pays circum-Méditerranéens - Empire chérifien. Archives scientifiques du Protectorat français. Mémoires de la Société des sciences naturelles du Maroc,48: 1-235.
164Wachmann E., Melber A. & Deckert J., 2008. Wanzen. Band 4. Die Tierwelt Deutschlands 81. Verlag Goecke & Evers. 230 p.
165Wagner E., 1963. Untersuchungen über den taxonomischen Wert des Baues der Genitalien bei den Cydnidae (Hem. Het.). Acta Entomol. Mus. Natl. Pragae,35: 73-115.
166Werner D. J., 2010. Die Schwarznesselwanze Tritomegas sexmaculatus als rezenter Arealerweiterer und ihre Abtrennung von T. bicolor (Heteroptera: Cydnidae) : Verbreitung und Angaben zur Biologie. [The Bug Tritomegas sexmaculatus on Black Horehound as Host Plant, a Recent Area Expanding Species and its Separation from T. bicolor (Heteroptera: Cydnidae) : Distribution and Remarks on Biology.] Entomologie heute,22: 55-84.
167Ziaja D. J., 2013. Nowe stanowisko Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) w Polsce. [New record of Tritomegas sexmaculatus (Rambur, 1839) (Hemiptera: Heteroptera: Cydnidae) from Poland]. Heteroptera Poloniae - Acta Faunistica,7: 3-6.
PLANCHES
168Planche I - Imagos

169Planche II – Juvéniles au stade V

