


- Accueil
- Volume 72 (2019)
- Termites (Blattodea: Termitidae) diversity and assemblages in different yam fields habitats in central Benin


Visualisation(s): 4629 (60 ULiège)
Téléchargement(s): 136 (0 ULiège)

Termites (Blattodea: Termitidae) diversity and assemblages in different yam fields habitats in central Benin

Document(s) associé(s)
Version PDF originaleNotes de la rédaction
Received on November 21st 2018, accepted on February 26th 2019.
Résumé
Les termites sont connues pour causer des pertes économiques importantes dans les champs d’igname. Par conséquent, l’évaluation de la diversité et de l’abondance des termites dans les champs d’igname est essentielle pour leur gestion. Dans cette étude, nous avons comparé la richesse spécifique, l'abondance et la diversité de groupes fonctionnels des termites ravageurs dans les champs d’igname de plateaux (15) et bas-fonds (9), à travers 15 villages en utilisant la méthode normalisée de transect en ceinture. La diversité spécifique de termites rencontrée dans les champs d’igname a été comparée à celle de la forêt classée du Bénin d’Agoua. Huit genres classés en quatre sous-familles (Nasutitermitinae, Macrotermitinae, Amitermitinae et Coptotermitinae) regroupés en deux familles (Termitidae et Rhinotermitidae) ont été identifiés. Le plus grand nombre d'espèces (13) a été identifié dans les champs de plateaux, tandis que le plus petit nombre (10) a été identifié dans les bas-fonds. La faune de termites collectée dans les champs d'ignames et dans la forêt d'Agoua a été distincte sur base des indices de similarité. Amitermes evuncifer (Silvestri) a été l'espèce de termite la plus largement répandue et la plus dominante dans les champs d'ignames de plateaux, tandis que dans les bas-fonds, il s'est agi de Macrotermes subhyalinus (Rambur). Trois groupes fonctionnels ont été observés avec des consommateurs d’humus que l’on ne trouve que dans les champs d’igname de plateaux. Suite à l'analyse en composantes principales, les 24 transects échantillonnés ont été classés en quatre groupes ayant des termites avec différents comportements alimentaires. La prédominance des non-cultivateurs de champignons a été observée dans les groupes principalement composés de transects des champs d'igname de plateau. Trois habitats de nidification étaient représentés (monticule épigé, bois et sous-sol). Les résultats de la présente recherche pourraient aider à développer des stratégies efficaces contre les principaux termites ravageurs de l'igname identifiés.
Abstract
Termites are known to cause significant economic losses in yam fields. Therefore, assessing diversity and abundance of termites in yam fields is critical for their control. In this study, we compared the specific richness, abundance, and diversity of functional groups of termite pests in upland and lowland yam fields, across 15 villages using the standardized belt transect method. The specific termite diversity encountered in yam fields was compared with that of the Agoua classified forest. In general, eight genera classified into four subfamilies (Nasutitermitinae, Macrotermitinae, Amitermitinae and Coptotermitinae) grouped into two families (Termitidae and Rhinotermitidae) were identified. The highest number of the species (13) were identified in upland fields while the lowest number (10) were identified in the lowlands. Termite fauna collected from yam fields and from the Agoua forest were distinct, as showed by the similarity index. Amitermes evuncifer (Silvestri) was the most widely distributed and the most dominant termite species in upland yam fields while in lowland yam fields it was Macrotermes subhyalinus (Rambur). Three functional groups were observed with humus feeders only found in upland yam fields. Principal component analysis classified the 24 sampled transects into four groups with different termites feeding groups. The dominance of non-fungus-growers was observed in the groups mostly composed of transects in uplands yam fields. Three nesting habitats were represented (epigeal mound, wood, and subterranean). The results of this present research could help to develop efficient strategies against the identified key yam termite pests.
Table des matières
1lokoestellel@yahoo.fr
INTRODUCTION
2Termites are an important insect group in ecosystems across the tropics, including African savannas (Davies et al., 2015). They play a vital role in recycling wood and plant materials, and providing food for other animals (Jouquet et al., 2011). Their tunnelling efforts help to ensure that the soil is porous, aerated and enriched in minerals and nutrients to support better plant growth (Wood, 1996). In tropical areas, however, termites cause tremendous losses to agricultural crops. Approximately 2,800 described species of termites are known globally, but only 185 species have been reported as pests (Verma et al., 2009).
3Yams (Dioscorea sp.) correspond to a vital component of the agricultural sector in Benin in terms of food security, social and cultural values. Yam production requires soil high fertility and local farmers have kept with the traditional system of planting yams as the first crop after forest or a long fallow period (Diby et al., 2011; Akom et al., 2015). However, this cropping management is very often the cause of termite problems as termites become agricultural pests when humans destroy or modify their natural habitats as it is the case with yam cultivation (Wood, 1996). Consequently, the attacks of termites on yam remain an important problem in Benin and particularly in the Savanna woodlands of Central Benin (Loko et al., 2013a). Infested tubers present only one or two holes at harvest while its internal flesh is entirely consumed (Loko et al., 2013a). Termites being constantly present in the environment throughout the cropping season, economic losses in yam production are very often important. Such losses can reach more than 5 tons/ha or 50% of the yield in highly infested soils (Atu, 1993). Loko et al. (2015) recorded 10 species of termites recognized by farmers as pests of yam, among which A. evuncifer (Termitidae: Amitermitinae) and T. oeconomus (Termitidae: Nasutitermitinae) were designated as the most damaging ones. However, the diversity and abundance of termites in yam fields remains largely unknown.
4In central Benin, yam is cultivated both in upland and lowland ecosystems with an influence on the presence of a termite species depending on the vegetation type (Aslam et al., 2000), habitat disturbance (Jones et al., 2003) and fragmentation (Davies et al., 2003). Consequently, it is important to investigate the termite biodiversity in these two habitats. Additionally, in order to better assess their impact on yam cultivation, it is useful to classify termites according to functional groups based on the types of food substrate, as well as the locations of their nesting and feeding sites (Jouquet et al., 2011). The present work aimed to evaluate the composition, richness, abundance, feeding and nesting habits of termite species found in a yam fields in Central Benin.
MATERIAL AND METHODS
2.1. Study area and sampling sites
5The study was carried out in an area of Central Benin located between latitude 7°45' and 8° 40' North and longitude 2° 20' and 2° 35' East. The selected area represents 17% of the total surface of Benin with a Sudano-Guinean climate and a bimodal rainfall pattern unevenly distributed between two rainy seasons, ranging between 800 to 1200 mm per year (Adam & Boko, 1993). The soils are mainly ferruginous and relatively rich in minerals. The major crops grown in this area include maize, groundnuts, cassava, and yams. A total of fifteen villages were randomly selected throughout the different district of Central Benin (Figure 1). In each village, yam fields located respectively in the upland (15 villages) and lowland (9 villages) were prospected. To compare the species richness of termites in yam fields versus natural environment, a sampling of termite was made in the classified forest of Agoua, located in the western part of central Benin in the Bantè district (Figure 1). The Agoua forest was declared as protected area by Decree No. 8 104 SE of 4 November 1953 and covers 75,300 ha (Idrissou et al., 2011). In 2002, the Benin government initiated the restoration of the forest massif of Agoua, due to its importance in Benin’s ecosystem. High level of degradation, due to severe anthropogenic influences such as poaching, logging, grazing, and agricultural activities (Idrissou et al., 2011).
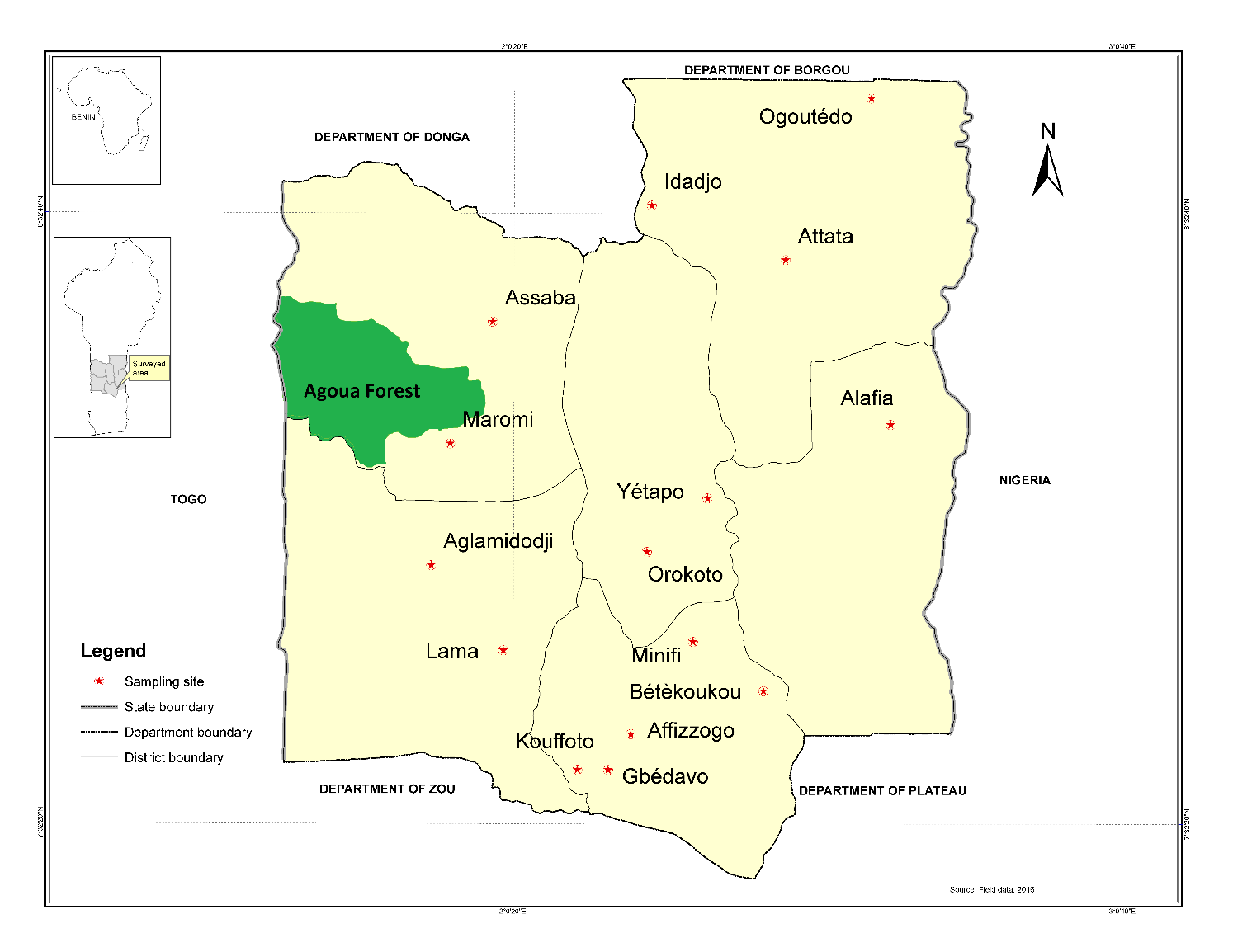
6Figure 1: Map of Central Benin showing the geographical positions of the sampling sites
2.2. Sampling of termites in yam fields
7Termites were sampled using a standardized method designed for rapid assessment of termite diversity described by Jones and Eggleton (2000). One transect (100 m long and 2 m wide) was delimited in each yam field and inside the Agoua forest zone. Each transect was divided into 20 contiguous sections (each 5m × 2m). Each section was sampled for 30 min by two peoples. Within each transect, the following microhabitats were searched for termites: i) soil surface (12 samples, each 12 cm x 12 cm to 10 cm depth); ii) leaf litter on soil surface and tree buttresses; iii) dead tree stumps, logs, branches and twigs; and iv) runways on trees and arboreal nests up to 2 m height. Soldier and worker termites were collected into vials containing 80 % ethanol.
2.3. Classification and termite identification
8Termite identification was carried out at the Evolutionary Ecology and Biology laboratory at the Université Libre de Bruxelles (ULB), based on caste soldiers or workers for groups of termites lacking the soldier castes. Specimens of the same genus were determined to species using various standard determination keys developed by Sands (1959, 1965, 1972), Bouillon and Mathot (1965), and Ruelle (1970).
2.4. Feeding and nesting groups classification
9Feeding behaviours of termites were derived from generic identity and feeding group classification of Donovan et al. (2001). Accordingly, species were assigned to the following four feeding groups and five feeding diets: group I (W), live-wood-feeding lower termites; group II (WF), fungus-growing wood feeders; group III (NF), non-fungus-growing wood feeders; group IV (I), soil–wood interface feeders (“humus feeders”); group V (S), true soil feeders.
10Termites were subsequently assigned to nesting groups according to Eggleton et al. (2002): (i) hypogeal nesting (found below ground); (ii) wood nesting (found in woody items); (iii) epigeal (in mounds protruding above ground); and (iv) arboreal (in tree nests).
2.5. Statistical analysis
11Sampling completeness was tested by constructing sample-based species accumulation curves and by recording the auto-similarity which is the mean between transects of the same habitat type (Dosso et al., 2010) using EstimateS 9.1.0 software (Colwell et al., 2013). The observed and estimated species accumulation curves were constructed after randomizing 500 times the sample in order to ensure the statistical representation of the target community (Cao et al., 2002). The species richness was determined by enumerating the number of species observed over the whole transects (Dosso et al., 2012). As we used presence-absence data, the relative abundance was defined as the number of encounters per transect, where the presence of one species in a quadrat represented one encounter (Magurran 2004). Using EstimateS 9.1.0 software, the Shannon index (H') and Simpson index (SI) were calculated. The β diversity or similarity index of Jaccard (Sj) was used to measure the similarity of species between sampling sites (yam field and Agoua forest) using the following formula:
 Where J = species common to both area, a = number of species found only on yam fields, b = number of observed species only on Agoua forest. This index varies from 0 (no similarity) to 1 (identical settings). The comparative zones are similar if Sj is ≥ 0.5 (Gbenyedji et al., 2011).
Where J = species common to both area, a = number of species found only on yam fields, b = number of observed species only on Agoua forest. This index varies from 0 (no similarity) to 1 (identical settings). The comparative zones are similar if Sj is ≥ 0.5 (Gbenyedji et al., 2011).
12To discover how termite species were distributed in the upland and lowland yam fields and better visualize the similarity of transects of both habitats, the UPGMA (Unweighted Pair-Group Method with Arithmetic Average) method was performed using the NTSYS-PC 2.21 software (Rohlf, 2009). In order to identify the patterns of termite feeding group abundance across the transects through the different habitats, a principal component analysis (PCA) was performed using Minitab 14 Software (Minitab the version 14, Minitab Inc., State College, Pa in the USA) by considering the transects as individuals and the type of feeding groups as variables.
RESULTS
3.1. Sampling efficiency
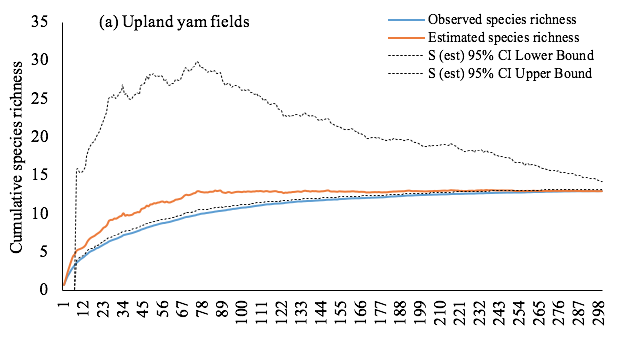
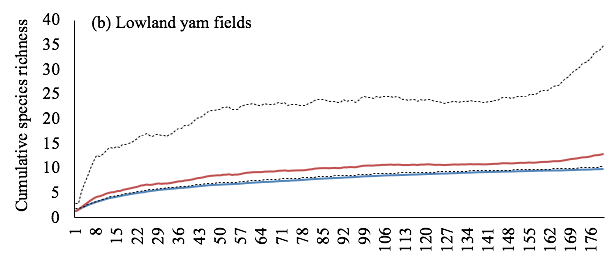
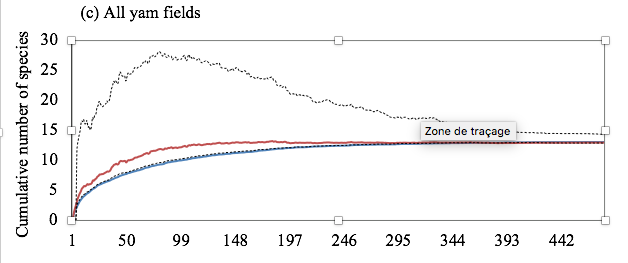 Figure 2: Sample-based accumulation curves of observed and estimated species richness in yam fields. Graphs show most likely estimates with 95% unconditional confidence intervals, calculated by the analytical formulas of Colwell et al. (2004).
Figure 2: Sample-based accumulation curves of observed and estimated species richness in yam fields. Graphs show most likely estimates with 95% unconditional confidence intervals, calculated by the analytical formulas of Colwell et al. (2004).
13The accumulation curves for observed and estimated species richness were similar among the upland yam fields (Figure 2a). Then, the species richness observed was considered a very good estimate of that expected in this habitat. In contrast, the higher accumulation curve for estimated species richness in the lowland yam fields suggests that a further sampling effort must be done to discover more species (Figure 2b). However, when data of the two habitats were pooled, the observed and expected species richness were similar, suggesting a good estimation of the overall termite diversity in yam fields (Figure 2c). With an auto-similarity values of 0.58 and 0.53, transects belonging respectively to upland and lowland yam fields did not differ in their taxonomic composition. The same trend was observed with pooled data from both yam field habitats, with an auto-similarity value of 0.55.
3.2. Species composition, richness and diversity
14Termites collected across yam fields belonged to two distinct families, Termitidae and Rhinotermididae, and were represented by three (Nasutitermitinae, Macrotermitinae and Termitinae) and one (Coptotermitinae) subfamilies respectively. The subfamily of Macrotermitinae was the most diverse with 5 species (Table 1), followed by Nasutitermitinae and Termitinae with 4 and 3 species (Table 1), respectively. The subfamily of Coptotermitinae is the least represented with one species. A total of 13 species were identified in yam fields (Figure 3). While, a total of 9 species of termites was found in Agoua forest belonging to Termitidae family. Nasutitermes arborum (Smeathman) and Promirotermes sp. were confined to Agoua forest (Table 1). Jaccard similarity index of 0.47 indicated a low similarity of the termite species compositions of the two habitats.

15Table 1: Details of termite taxa recorded from yam fields and Agoua forest in Central Benin
|
Termite taxa |
Environment |
|||
|
Family |
Subfamily |
Species |
Yam field |
Agoua forest |
|
Termitidae |
Nasutitermitinae |
Trinevitermes geminatus |
* |
* |
|
Trinevitermes togoensis |
* |
|||
|
Trinevitermes oeconomus |
* |
* |
||
|
Fulleritermes tenebricus |
* |
|||
|
Nasutitermes arborum |
* |
|||
|
Macrotermitinae |
Macrotermes bellicosus |
* |
* |
|
|
Macrotermes subhyalanus |
* |
* |
||
|
Macrotermes ivorensis |
* |
|||
|
Ancistrotermes cavithorax |
* |
* |
||
|
Microcerotermes sp. |
* |
|||
|
Termitinae |
Amitermes evuncifer |
* |
* |
|
|
Amitermes guineensis |
* |
|||
|
Pericapritermes sp. |
* |
* |
||
|
Promirotermes sp. |
* |
|||
|
Rhinotermididae |
Coptotermitinae |
Coptotermes sjostedti |
* |
|
16The four subfamilies (Nasutitermitinae, Macrotermitinae, Termitinae, Coptotermitinae) were recorded in both upland and lowland fields (Figure 4). However, in lowland yam fields, ten species were recorded against 13 in upland yam fields. The hierarchical cluster built on the basis of the termite species composition of transects of both habitats did not show a clear separation of transects as a function of different habitats (Figure 5). However, some cluster of the dendrogram revealed identical termite species composition in transects of different sampling sites. For example, transects in upland yam fields 3, 4 and 6 which correspond respectively to transects in Orokoto, Gbedavo and Minifi sampling sites were clustered similarly (Figure 5). Likewise, the termite species composition of transects in upland yam field 11 (Aglamidodji) was clustered similarly with transects in lowlands yam fields 4 (Gbedavo) and 5 (Betekoukou). Transects in upland yam field 14 (Affizzogo) and transects in lowland yam field 9 (Assaba) were completely detached from all the other transects due respectively to the presence of Microcerotermes sp. and Pericapritermes sp. in their termite species composition.
17In yam fields in general, the Simpson’s Index of 0.24 and Shannon’s Index of 1.78 was obtained (Table 2). A. evuncifer with a value of 0.15 and 0.37 for Simpson’s and for Shannon’s Indexes respectively was the dominant species followed by T. geminatus with a value of 0.05 and 0.34 for Simpson’s and Shannon’s indexes respectively. A similar trend was observed when upland yam fields were considered (Table 2). Also, in the lowland yam fields, M. subhyalinus and T. oeconomus with a value of 0.05 Simpson’s and respective value of 0.34 and 0.33 Shannon’s indexes were the dominant species (Table 3).
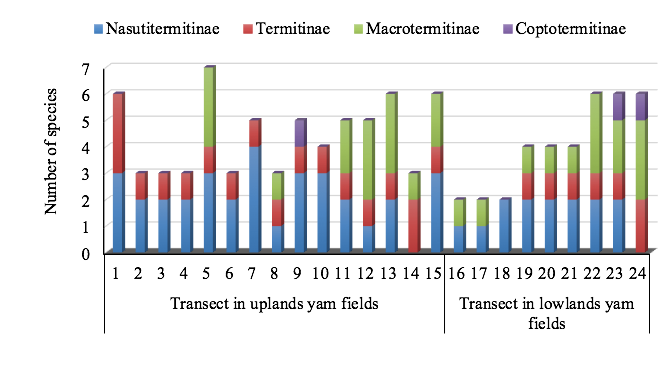 Figure 4: Taxonomic composition of termites from transect of the 24 prospected yam fields in central Benin.
Figure 4: Taxonomic composition of termites from transect of the 24 prospected yam fields in central Benin.
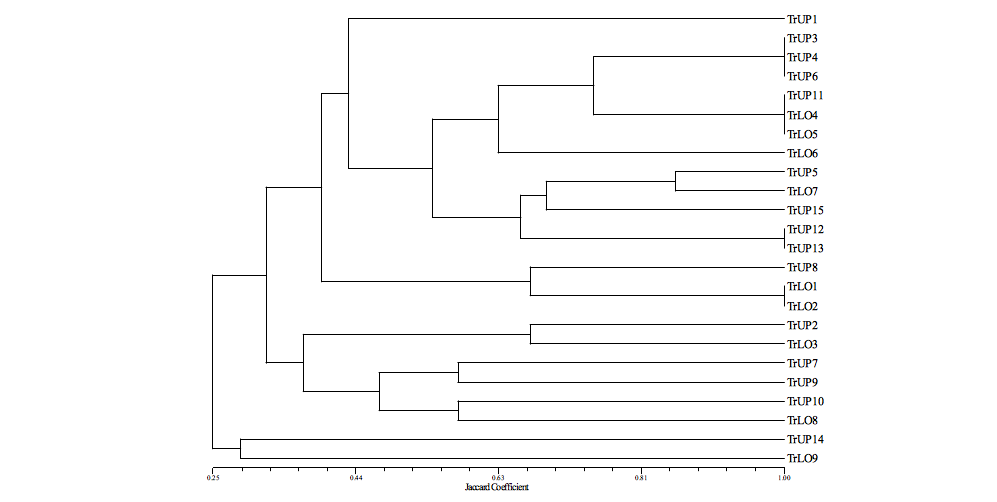

18Figure 5: Classification of uplands and lowlands transects based on their termite species composition with UPGMA using Jaccard coefficient. Abbreviations: TrUP 1-15, Transect in upland yam fields 1-15; TrLO 1-9, Transect in lowland yam fields 1-9.
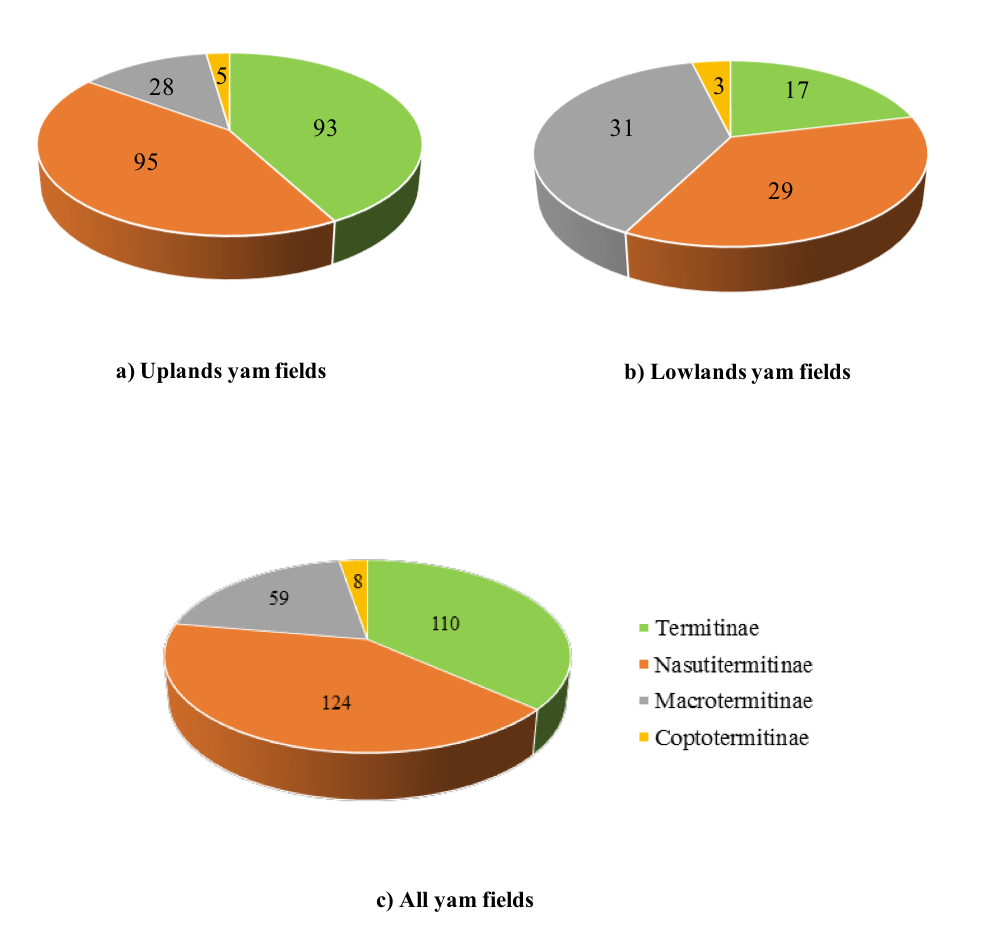
19Figure 6: Distribution of termite occurrences by Subfamily in yam fields in Central Benin.
3.3. Relative abundance
20Species from the Subfamily of Nasutitermitinae were most abundant in yam fields while the Coptotermitinae subfamily was the less represented (Figure 6a). In upland yam field, Nasutitermitinae and Termitinae were the most encountered species (Figure 6b) while species from the Subfamily Macrotermitinae were the most important in lowlands yam fields (Figure 6c).
21The relative abundance based on the number of each morphospecies encountered within yam field habitats types is given in Table 3. A total of 301 termites were recorded in the 24 transects surveyed. Overall, A. evuncifer was the most widely distributed and the most dominant termite species in yam fields, representing 34.5% of all samples. It was also the most dominant species in uplands yam fields, while M. subhyalinus was most dominant in lowland yam fields (Table 3). Amitermes guineensis, F. tenebricus and Pericapritermes sp. were only sampled from upland yam field.
22Table 2: Diversity index of number of individuals of different termite species collected from 24 yam field in central Benin.
|
Termite species |
Upland yam fields |
Lowland yam fields |
All yam fields |
|||
|
Simpson’s index |
Shannon’s index |
Simpson’s index |
Shannon’s index |
Simpson’s index |
Shannon’s index |
|
|
Amitermes evuncifer |
0.21 |
0.34 |
0.03 |
0.31 |
0.15 |
0.37 |
|
Amitermes guineensis |
0.00 |
0.11 |
0.00 |
0.00 |
0.00 |
0.11 |
|
Ancistrotermes cavithorax |
0.00 |
0.27 |
0.00 |
0.01 |
0.00 |
0.06 |
|
Coptotermes sjostedti |
0.00 |
0.06 |
0.00 |
0.06 |
0.00 |
0.07 |
|
Fulleritermes tenebricus |
0.00 |
0.04 |
0.00 |
0.00 |
0.00 |
0.05 |
|
Macrotermes bellicosus |
0.00 |
0.10 |
0.00 |
0.06 |
0.00 |
0.05 |
|
Macrotermes ivorensis |
0.00 |
0.04 |
0.00 |
0.09 |
0.00 |
0.05 |
|
Macrotermes subhyalanus |
0.00 |
0.07 |
0.05 |
0.33 |
0.01 |
0.20 |
|
Microcerotermes sp. |
0.00 |
0.00 |
0.00 |
0.02 |
0.00 |
0.01 |
|
Pericapritermes sp. |
0.00 |
0.36 |
0.00 |
0.00 |
0.00 |
0.06 |
|
Trinervitermes geminatus |
0.06 |
0.09 |
0.03 |
0.31 |
0.05 |
0.34 |
|
Trinervitermes oeconomus |
0.02 |
0.01 |
0.05 |
0.34 |
0.03 |
0.30 |
|
Trinervitermes togoensis |
0.00 |
0.07 |
0.00 |
0.12 |
0.00 |
0.11 |
|
Total |
0.29 |
1.56 |
0.16 |
1.65 |
0.24 |
1.78 |
3.4. Feeding and nesting groups
23Out of the total of 13 species recorded, 11 species were wood /litter/micro-epiphytes feeders and only one species was respectively wood feeder (C. sjostedti) and organic rich soil/humus feeder (Pericapritermes sp.) (Table 3). None of the observed species belonged to Group IV - true soil feeders. The feeding group III was only found in upland yam while in lowland yam fields two feeding groups (group I and II) were predominant (Figure 7).
24The principal component analysis identified four components among which the two first explain 69.3 % of the total variability (Table 4). These two first components were used to analyse the patterns of termites' feeding groups abundance across the transects of different habitats. PC1 is positively associated with group II (WF) and group I (W) and negatively to group II (NF) which is explained by 44.7 % of the total variation (Table 4). Only termites of group III are associated with PC2 with 24.6 % of the total variability (Table 4).
25For all sampling sites and based on the PC1 and PC2, the 24 transects were classified into four groups with different termite feeding group composition named G1 to G4 (Figure 8). The presence of soil-wood-feeders (group III) characterised the two transects grouped in G1. G2 included mainly transects in upland yam fields and was characterised by the dominance of non-fungus-growers (group II (NF)). G3 was characterized by the abundance of fungus and non-fungus growers (group II (WF and NF)) included in transects of lowland yam fields. Termites feeding on living wood (group I) characterized G4 which included only of one transect in lowland yam fields.
26Table 3: Feeding habitsb, nesting group and Occurrencesa of observed termite species in yam.
|
Species |
Feeding group |
Nesting group |
Occurrences |
Total occurrencs (n=480) |
|
|
Uplands (n=300) |
Lowlands (n=180) |
||||
|
Amitermes evuncifer |
II (NF) |
Wood-subterranean |
87 |
17 |
104 |
|
Amitermes guineensis |
II (NF) |
Wood-subterranean |
3 |
0 |
3 |
|
Ancistrotermes cavithorax |
II (WF) |
Wood |
6 |
1 |
7 |
|
Coptotermes sjostedti |
I (W) |
Wood |
5 |
3 |
8 |
|
Fulleritermes tenebricus |
II (NF) |
Epigeal mound |
4 |
0 |
4 |
|
Macrotermes bellicosus |
II (WF) |
Epigeal mound |
6 |
3 |
9 |
|
Macrotermes ivorensis |
II (WF) |
Epigeal mound |
2 |
1 |
3 |
|
Macrotermes subhyalanus |
II (WF) |
Epigeal mound |
13 |
25 |
38 |
|
Microcerotermes sp. |
II (NF) |
Wood |
1 |
1 |
2 |
|
Pericapritermes sp. |
III (I) |
Subterranean |
3 |
0 |
3 |
|
Trinervitermes geminatus |
II (NF) |
Epigeal mound |
49 |
15 |
64 |
|
Trinervitermes oeconomus |
II (NF) |
Epigeal mound |
33 |
11 |
44 |
|
Trinervitermes togoensis |
II (NF) |
Epigeal mound |
9 |
3 |
12 |
|
Total |
- |
- |
221 |
80 |
301 |
27a Numbers shown are occurrences; i.e. the number of quadrats (5 m × 2 m) in which each species was present.
28b Feeding habits: wood feeders I (W), fungus-growing wood feeders II (WF), non-fungus-growing wood feeders II (NF), soil–wood interface feeders III (I) and soil feeders IV (S).
29Three nesting habitats were represented in Table 3, with 53.84 % of the species nesting in epigeal mound, 38.46 % in wood, and 23.07 % in subterranean nests. Epigeal mound builders included F. tenebricus, M. bellicosus, M. ivorensis, M. subhyalanus, T. geminatus, T. oeconomus, and T. togoensis. Wood nesting species were A. evuncifer, A. guineensis, A. cavithorax, C. sjostedti and Microcerotermes sp. while subterranean nesters were A. evuncifer, A. guineensis, and Pericapritermes sp.
30Table 4: Eigen vectors and values of the principal component axes component analysis of abundance of termites feeding groups in transect of the different yam fields habitats.
|
Eigen vectors |
Eigen value |
Proportion |
Cumulative |
|
PC1 |
1.7867 |
0.447 |
0.447 |
|
PC2 |
0.9835 |
0.246 |
0.693 |
|
PC3 |
0.7954 |
0.199 |
0.891 |
|
PC4 |
0.4344 |
0.109 |
1.000 |
|
Variables |
PC1 |
PC2 |
PC3 |
|
III (I) |
-0.235 |
0.905 |
0.339 |
|
II (WF) |
0.643 |
0.096 |
-0.048 |
|
I (W) |
0.501 |
-0.118 |
0.777 |
|
II (NF) |
-0.529 |
-0.398 |
0.528 |
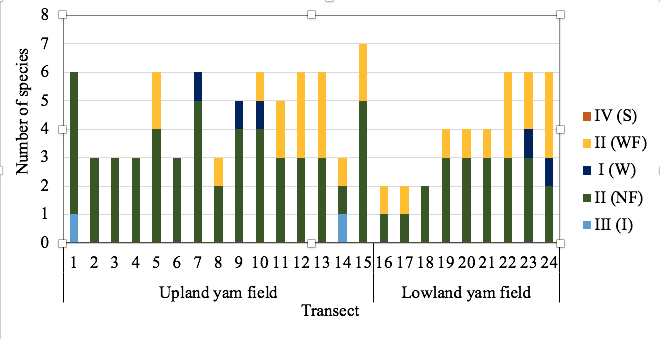
31Figure 7: Feeding group composition of termites from transect of the 24 prospected yam fields in Central Benin as defined by Donovan et al. (2001).
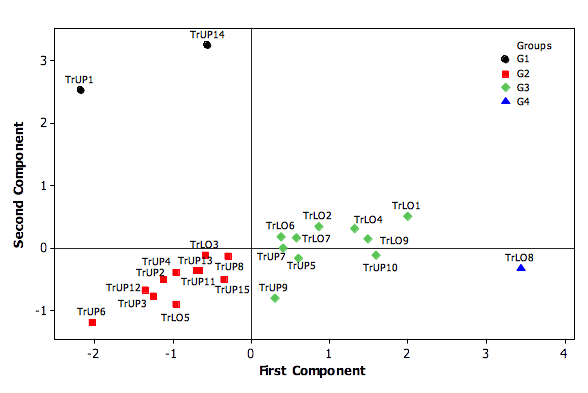

32Figure 8: Principal component analysis based on the abundance of termites feeding groups in transect of the different yam fields habitats. Abbreviations: TrUP 1-15, Transect in upland yam fields 1-15; TrLO 1-9, Transect in lowland yam fields 1-9.
DISCUSSION
33Our results allowed to infer that the termite species richness was efficiently assessed in the transects of upland yam fields, as illustrated by the converging observed and estimated rarefaction curves and the associated confidence intervals. In lowlands yam fields, however, the estimated trend curves indicated that additional samplings would be required to provide an accurate picture of the termite species richness. Nevertheless, pooling all samples from both habitats provided a good picture of the overall termite species composition in yam fields. Termitidae family displayed the highest number of species and abundance. This finding is comparable to those on others crops such as rice (Togola et al., 2012), maize (Ouédrago et al., 2015) and sugarcane (Junqueira et al., 2015), but also in tree plantations such as oil palms (Kon et al., 2012), coffee (Bhavana et al., 2015), teak (Gbenyedji et al., 2011), eucalypt (Calderon & Constantino, 2007) and neem (Adedeji et al., 2015). Termite species richness found in our study with 13 species was much higher compared with studies of termite diversity in agricultural sites in West Africa. For example, in rice fields in Benin (Togola et al., 2012) and Ivory Coast (Akpesse et al., 2008) where only 6 and 7 species were found respectively. Similarly, in maize fields, low termite diversity was found in Ivory Coast by Akpesse et al. (2008) with 4 species, respectively, composed only of Armitermes, Ancistrotermes, Microtermes, and Pseudacanthotermes genera.
34The evaluation of the termite diversity through the species richness both in yam fields and in the Agoua forest showed a higher diversity index in the former. The low species richness of termites in the Agoua forest was surprising because it is known that forests have greater structural complexity (larger trees, greater plant diversity and more dead wood), which is supposed to provide a larger array of nesting and feeding sites for termites (Jones & Prasetyo 2002). However, the distinctiveness of the termite assemblages in the Agoua forest can be attributed to several factors among which land use (Attignon et al., 2005), habitat disturbance (Eggleton et al., 2002) and habitat fragmentation (Davies, 2003) were considered as the key factors. In fact, when the Agoua forest was declared a protected area in 1953, the allocated area was 75.300 ha. Nevertheless, in 2002 it was reduced to 68.848 ha due to various anthropogenic influences such as, logging, grazing, and agricultural activities (Idrissou et al., 2011).This study showed that termite species richness was lower in the lowland yam fields (10 species), possibly due to unfavourable environmental conditions. In fact, regular inundation or high water table in lowland ecosystem inhibits soil-dwelling termites (Abensperg-Traun & Boer 1990). Morevoer, the absence of Kalotermitidae in lowland yam fields could be explained by the fact that lowlands are disturbed environments with considerably less trees and dead wood, decreasing the likely nesting sites of the group (Vasconcellos et al., 2010).
35The abundance of Nasutermitinae in yam fields is not surprising in view of agricultural practices promoting the conservation of straw mulch in the yam fields to curb soil erosion (Akinola & Owombo, 2012). Among this subfamily, Trinevitermes genus is corresponding to grass feeders and their main presence in the upland yam fields is certainly due to availability of abundant grass vegetation (Mugerwa et al., 2011; Dosso et al., 2012). As already reported by yam farmers of Central Benin (Loko et al., 2015), A. evuncifer was the most widely distributed and the most dominant termite species in yam fields also in the present study. In Africa, A. evuncifer is well known to attack various crops (Okwakol, 2000), being a main pest of rice in Benin (Togola et al., 2012), and neem in Nigeria (Adedeji et al., 2015).
36The dominance of Macrotermitinae subfamily in lowlands yam fields is not surprizing, because this subfamily is characterized by having a particularly soft cuticle with poor water-retaining properties (Moore, 1969), then preferring to construct their habitat in moist lowlands in order to avoid desiccation (Hesse, 1957). Also, termites mound of Macrotermitinae are known to have a special “drip tip”-like structures which allow the efficient flow of water over and around them (Eggleton, 2011). Moreover, their ability to live in disturbed habitats is due to their remarkable adaptation favored by the symbiotic relationship they have with Termitomyces fungus (Tenon et al., 2013). Among the Macrotermitinae, M. subhyalinus was the most dominant and abundant species in lowland yam fields. This species is known to be responsible for major damages in sugarcane plantation in Nigeria (Collins, 1984). In the present study, A. evuncifer and M. subhyalinus were identified as the dominant pest species both in upland and lowland yam fields. However, further studies need to be conducted for quantifying the destructive behavior of these two species as yam pest.
37The true soil feeders group was the one most sensitive to habitat disturbance (Jones et al., 2003). In fact, no termite species of this group was found in yam fields, confirming previous observations that the abundance of soil feeder termites was negatively correlated to habitat degradation (Eggleton et al., 2002; Jones & Prasad 2002; Jones et al., 2003). In contrast, the presence of organic rich soil/humus feeders in some upland yam fields could be an indicator of soil fertility (Poovoli & Rajmohana, 2016). In yam fields, farmers used grass straw to cover the yam mount as a strategy to overcome drought stress (Loko et al., 2013b) which can be an additional source of food for termites, especially for fungus-growing wood feeders groups. Fungus-growing termites, known to feed on crop residues, mulch and soil organic matter (Sileshi et al., 2005), were dominant in lowland yam fields. However, when there is scarcity of their primary food sources, they also attack crops including yams (Atu, 1993), and their damage is known to increase in soils with low organic matter content (Umeh & Ivbijaro, 1997; Uys, 2002; Van den Berg & Riekert, 2003).
CONCLUSIONS
38This study identified the main termite species attacking yams in uplands and lowlands of Central Benin. A total of thirteen species was found in yam fields against nine in natural environment (Agoua forest). The most dominant and abundant species were A. evuncifer and M. subhyalinus in upland and lowland yam fields respectively. Trophic composition of termites varied as a function of the habitat. Non-fungus-growers were the most abundant trophic group in upland yam fields, whereas fungus-growing termites were most abundant in lowland yam fields. In the light of these results, further studies will be needed to assess the impact of A. evuncifer and M. subhyalinus on yam yield.
ACKNOWLEDGEMENTS
39We thank anonymous reviewers for their comments on a previous version of the manuscript. This work was carried out with the technical support of Belgian National Focal Point to the Global Taxonomy Initiative (cbdgti@naturalsciences.be). We also thank Prof. Guy Josens by documentary the different species of termites recorded.
Bibliographie
Abensperg-Traun, M. & Boer, E.S., 1990. Species abundance and habitat differences in biomass of subterranean termites (Isoptera) in the wheatbelt of Western Australia. Austral Ecology, 15(2), 219–226.
Adam S. & Boko M., 1993. Le Bénin. Les éditions du Flamboyant/EDICEF.
Adedeji G.A., Emerhi, E.A. & Nyenke E. 2015. Incidence and Severity of Termites Infestations on Azadirachta indica A. Juss. Used as Avenue Trees in University of Port Harcourt, Nigeria. IOSR Journal of Agriculture and Veterinary Science, 8(2), 123-126.
Akinola A. & Owombo, P., 2012. Economic Analysis of Adoption of Mulching Technology in Yam Production in Osun State, Nigeria. International Journal of Agriculture and Forestry, 2(1), 1-6.
Akom M., Oti-Boateng, C., Otoo E. & Dawoe E., 2015. Effect of Biochar and Inorganic Fertilizer in Yam (Dioscorea rotundata Poir) Production in a Forest Agroecological Zone. Journal of Agricultural Science, 7, 211-222.
Akpesse A.A., Kouassi P.K., Tano Y. & Lepage M., 2008. Impact des termites dans les champs paysans de riz et de maïs en savane sub-soudanienne (Booro Borotou, Côte-d’Ivoire). Science et Nature, 5, 121–131.
Aslam M., Shaheen F.A. & Rehman A., 2000. Screening of sunflower (Helianthus annuus Linnaeus) Genotypes against the attack of Odontotermes obesus (Rambur) (Isoptera: Termitidae). Pakistan Journal of Biological Sciences, 3, 2238-2240.
Attignon S.E., Lachat T., Sinsin B., Nagel P. & Peveling R., 2005. Termite assemblages in West African semi-deciduous forest and teak plantation. Agriculture, Ecosystems and Environment, 110, 318-326.
Atu U.G. 1993. Cultural practices for the control of termite (Isoptera) damage to yams and cassava in south-eastern Nigeria. International Journal of Pest Management, 39, 462-466.
Bhavana K.V., Poovoli A., Rajmohana K. & Shweta M., 2015. A Comparison on Termite Assemblages in Coffee and Teak Plantations and Semi-Evergreen Forest - A case study in North Wayanad, Kerala, India. Tropical Agricultural Research, 26, 456-467.
Bouillon A. & Mathot G., 1965. Quel est ce termite Africain? Zooleo n°1, Leopoldville Univ, Leopoldville.
Calderon R.A. & Constantino R., 2007. A survey of the termite fauna (Isoptera) of an eucalypt plantation in Central Brazil. Neotropical Entomology, 36, 391-395.
Cao Y., Williams, D.D. & Larsen P.D., 2002. Comparison of ecological communities: the problem of sample representativeness. Ecological Monogaphs, 72, 313–318.
Collins N.M., 1984.Termite damage and crop loss studies in Nigeria-assessment of damage to upl & sugarcane. Tropical Pest Management, 30(1), 26-28.
Colwell R. K., 2013. Estimates, Version 9.1: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User's Guide). Freeware for Windows and Mac OS.
Colwell R.K., Mao C.X. & Chang J., 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology, 85(10), 2717-2727.
Davies A.B., Eggleton P., van Rensburg B.J. & Parr C.L., 2015. Seasonal activity patterns of African savanna termites vary across a rainfall gradient. Insectes Sociaux, 62(2), 157–165.
Davies R.G., Eggleton P., Jones, D.T., Gathorne-Hardy F.J. & Hernandez L.M., 2003. Evolution of termite functional diversity: analysis and synthesis of local ecological and regional influence richness on local species. Journal of Biogeography, 30(6), 847-877.
Diby L. N'G., Tie B.T., Girardin O., Sangakkara R. & Frossard E., 2011. Growth and Nutrient Use Efficiencies of Yams (Dioscorea spp.) Grown in Two Contrasting Soils of West Africa. International Journal of Agronomy, vol. 2011, Article ID 175958, 8 p.
Donovan S.E., Eggleton P. & Bignell D.E., 2001. Gut content analysis and a new feeding group classification of termites. Ecological Entomology, 26, 356-366.
Dosso K., Yéo K., Konaté S. & Linsenmair K.E., 2012. Importance of protected areas for biodiversity conservation in central Côte d’Ivoire: Comparison of termite assemblages between two neighboring areas under differing levels of disturbance. Journal of Insect Science, 12, 1-18.
Dosso K., Konaté S., Aidara D. & Linsenmair K.E., 2010. Termite diversity and abundance across fire-induced habitat variability in a tropical moist savanna (Lamto, Central Côte d’Ivoire). Journal of Tropical Ecology, 26, 323–334.
Eggleton P., Bignell D.E., Hauser S., Dibog L., Norgrove L. & Madong B., 2002. Termite diversity is a cross an anthropogenic disturbance gradient in humid forest zone of West Africa. Agriculture, Ecosystems and Environment, 90(2), 189-202.
Eggleton P., 2011. An Introduction to Termites: Biology, Taxonomy and Functional Morphology. In D.E. Bignell, Y. Roisin and N. Lo (Eds.), Biology of termites: A modern analysis. Springer, 1-26.
Gbenyedji J.N.B.K., Anani K.E., Amevoin K. & Glitho, I.A., 2011. Diversité spécifique des termites (Isoptera) dans deux plantations de tecks (Tectona grandis L.) au sud du Togo. International Journal of Biological and chemical Sciences, 5, 755-765.
Hesse P.R., 1957. Fungus combs in termite mounds. East African Agricultural and Forestry Journal, 23, 104- 105.
Idrissou L., Aarts N., Paassen A. & Leeuwis C., 2011. The discursive construction of conflict in participatory forest management: The case of the Agoua forest restoration in Benin. Conservation and Society, 9, 119-131.
Jones D.T. & Eggleton P., 2000. Sampling termite assemblages in tropical forests: testing a rapid biodiversity assessment protocol. Journal of Applied Ecology, 37, 191-203.
Jones D.T. & Prasetyo A.H., 2002. A survey of the termites (Insecta: Isoptera) of Tablong district, South Kaimantan. Indonesia. The Raffles bulletin of zoology, 50, 117−128.
Jones D.T., Susilo F.X., Bignell D.E., Hardiwinoto S., Gillison A.N. & Eggleton P., 2003. Termite assemblage collapse along a l &-use intensification gradient in lowland central Sumatra, Indonesia. Journal of Applied Ecology, 40, 380-391.
Jouquet P., Traoré S., Choosai C., Hartmann C. & Bignell D., 2011. Influence of termites on ecosystem functioning. Ecosystem services provided by termites. European Journal of Soil Biology, 47, 215-222.
Junqueira L.K., Gonçalves E.R. & Teixeira L.M.C., 2015. Termite Communities in Sugarcane Plantations in Southeastern Brazil: an Ecological Approach. Entomo Brasilis, 8, 105-116.
Kon T-W., Bong C-F.J., King J-H.P. & Leong C-T.S., 2012. Biodiversity of Termite (Insecta: Isoptera) in Tropical Peat Land Cultivated with Oil Palms. Pakistan Journal of Biological Sciences, 15, 108-120.
Loko Y.L., Dansi A., Agre A.P., Akpa N., Dossou-Aminon I., Assogba P., Dansi M., Akpagana K., & Sanni A., 2013b. Perceptions paysannes et impacts des changements climatiques sur la production et la diversité variétale de l’igname dans la zone aride du Nord- Ouest du Bénin. International Journal of Biological and Chemical Sciences, 7, 672-695.
Loko Y.L., Agre P., Orobiyi A., Dossou-Aminon I., Roisin Y., Tamò M. & Dansi A., 2015. Farmers’ knowledge and perceptions of termites as pests of yam (Dioscorea spp.) in Central Benin. International Journal of Pest Management, 62, 75-84.
Loko Y.L., Dansi A., Dansi M., Vodouhè R. & Sanni A., 2013a. Indigenous knowledge and management of yam (Dioscorea cayenensis – Dioscorea rotundata complex) pests and diseases in northern Benin. International Journal of Applied Biology and Pharmaceutical Technology, 4, 277-287.
Magurran A.E., 2004. Measuring biological diversity. Blackwell Science Ltd.
Moore B.P., 1969. Biochemical studies in termites. In: Krishna, K. and Weesner, F.M. (Eds.) Biology of termites, vol 1. Academic Press, New York, London, 407-432.
Mugerwa S., Nyangito, M., Nderitu, J., Bakuneta, C., Mpairwe, D. &, Zziwa, E., 2011. Farmers’ ethno-ecological knowledge of the termite problem in semi-arid Nakasongola. African Journal of Agricultural Research, 6, 3183-3191.
Okwakol, M.J.N., 2000. Changes in termite (Isoptera) communities due to the clearance and cultivation of tropical forest in Uganda. African Journal of Ecology, 38, 1-7.
Poovoli A. & Rajmohana K., 2016. Feeding group diversity of Termites (Isoptera: Insecta) in Kerala. Journal of Entomology and Zoology Studies, 4, 114-116.
Rohlf F.J., 2009. NTSYSpc: numerical taxonomy system. ver. 2.21c. Exeter Software, Setauket, New York.
Ruelle J.E., 1970. A revision of the termites of the genus Macrotermes from the Ethiopian region (Isoptera: Termitidae). Bulletin of the British Museum (Natural History). Zoology, 24, 363-444.
Sands W.A., 1965. A revision of the termite subfamily Nasutitermitinae (Isoptera, Termitidae) from the Ethiopian region. Bulletin of the British Museum (Natural History), (Entomology) Supp, 4, 1-172.
Sands W.A., 1959. A revision of the termite of genus Amitermes from the Ethiopian region (Isoptera, Termitidae, Amitermitinae). Bulletin of the British Museum, 8, 129-156.
Sands W.A., 1972. The soldierless termites of Africa (Isoptera: Termitidae). Bulletin of the British Museum (Natural History), (Entomology) supp, 18, 244 p.
Sileshi G., Mafongoya P.L., Kwesiga F. & Nkunika P., 2005. Termite damage to maize grown in agroforestry systems, traditional fallows and monoculture on nitrogen-limited soils in eastern Zambia. Agricultural and Forest Entomology, 7, 61–69.
Tenon C., Boga J-P., Yapi A. & Kouassi K.P., 2013. Effects of Continuous Cultivation of Soil on Termites (Isoptera) Diversity & Abundance in Savannas of Northern of Côte d’Ivoire. Asian Journal of Agriculture and Rural Development, 3, 632-649.
Togola A., Kotoklo E.A., Nwilene F.E., Amevoin K., Glitho, I.A., Oyetundji O.E. & Kiepe P., 2012. Specific diversity and damage of termites on upland rice in Benin. Journal of entomology, 9, 352-360.
Umeh V.C., & Ivbijaro M.F., 1997. Termite abundance and damage in traditional maize-cassava intercrops in Southwestern Nigeria. Insect Science and Its Application, 22, 41-46.
Uys V., 2002. A guide to the termite genera of southern Africa. Plant Protection. Research Institute Handbook no. 15. Agricultural Research Council, South Africa.
Van den Berg J., & Riekert H.F., 2003. Effect of planting and harvesting dates on fungus-growing termite infestations in maize. South African Journal of Plant and Soil, 20, 76-80.
Vasconcellos A., Bandeira A.G., Moura F.M.S., Araújo V.F.P., Gusmão M.A.B., & Constantino R., 2010. Termite Assemblages in three habitats under different disturbance regimes in the semi-arid Caatinga of NE Brazil. Journal of Arid Environments, 74, 298–302.
Verma M., Sharma S. & Prasad R., 2009. Biological alternatives for termite control: A review. International Biodeterioration and Biodegradation, 63, 959–972.
Wood T.G., 1996. The agricultural importance of termites in the tropics. Agricultural Zoology Reviews, 7, 117-155.
