


- Startpagina tijdschrift
- Volume 78 (2025)
- Effects of cashew tree density on infestations by the wood borer, Apate terebrans (Pallas 1772) (Coleoptera, Bostrichidae), in Côte d'Ivoire, West Africa


Weergave(s): 854 (1 ULiège)
Download(s): 200 (1 ULiège)

Effects of cashew tree density on infestations by the wood borer, Apate terebrans (Pallas 1772) (Coleoptera, Bostrichidae), in Côte d'Ivoire, West Africa

Documenten bij dit artikel
Version PDF originaleRésumé
Apate terebrans (Pallas 1772) affecte considérablement le développement et la productivité des anacardiers (Anacardium occidentale Linnaeus 1753) en Afrique de l'Ouest, en forant des galeries à l’intérieur des arbres. La vie cachée de ce ravageur limite l'efficacité des méthodes de lutte par contact, et nécessiterait des méthodes de lutte culturales. Cette étude a évalué les effets de la densité des anacardiers sur les infestations par A. terebrans, dans dix vergers de deux zones agro-climatiques (représentées par Bondoukou et Bouna), au nord-est de la Côte d'Ivoire. Dans chaque verger, les anacardiers ont été dénombrés dans 13 quadrats de 400 m2, et le nombre d'infestations a été suivi mensuellement sur ces anacardiers, de juin 2018 à juillet 2020. Les indices de Blackman et de Morisita ont été utilisés pour évaluer la distribution des infestations. La densité à Bondoukou, 515 ± 235,5 anacardiers.ha-1, était significativement supérieure à celle de Bouna (235 ± 39,6 anacardiers.ha-1). Cependant, avec un coefficient de variation (CV) de 29,44%, la densité d’anacardiers à Bouna était plus homogène qu'à Bondoukou (CV = 58,64%). Dans les deux zones agro-climatiques, le nombre d'infestations a varié significativement entre les 13 quadrats du verger. Ainsi, la régression linéaire simple a montré que les fortes densités d’anacardiers réduisaient modérément et significativement les infestations. La distribution parcellaire d'anacardiers infestés était agrégée à Bondoukou et aléatoire à Bouna. Des méthodes d'analyses de répartition utilisant la distance entre les plants contribueraient à trouver des distances optimums entre les anacardiers qui limiteraient naturellement les infestations par A. terebrans.
Abstract
Apate terebrans (Pallas 1772) affects considerably the development and productivity of cashew trees (Anacardium occidentale Linnaeus 1753) in West Africa, by digging galleries inside the trees. The hidden life of this pest limits the effectiveness of contact control methods, and would require cultural control methods. This study assessed the effects of cashew tree density on A. terebrans infestations, in ten orchards in two agro-climatic zones (represented by Bondoukou and Bouna), in north-eastern Côte d'Ivoire. In each orchard, cashew trees were counted in 13 quadrats of 400 m2, and the number of infestations was monitored monthly on these cashew trees, from June 2018 to July 2020. The Blackman and Morisita indices were used to assess the distribution of infestations. The density in Bondoukou, 515 ± 235.5 cashew trees.ha-1, was significantly higher than that in Bouna (235 ± 39.6 cashew trees.ha-1). However, with a coefficient of variation (CV) of 29.44%, the cashew density in Bouna was more homogeneous than in Bondoukou (CV = 58.64%). In both agro-climatic zones, the number of infestations varied significantly among the 13 orchard quadrats. Thus, simple linear regression showed that high cashew tree densities moderately and significantly reduced infestations. The plot distribution of infested cashew trees was aggregated in Bondoukou and random in Bouna. Distribution analysis methods using the distance among plants would contribute to find optimum distances among cashew trees, which would naturally limit infestations by A. terebrans.
Reçu le 28 octobre 2024, accepté le 25 février 2025.
Cet article est distribué suivant les termes et conditions de la licence CC-BY (http://creativecomons.org/licenses/by/4.0/deed.fr)
INTRODUCTION
1The cashew tree, Anacardium occidentale L. 1753 (Anacardiaceae), is widely cultivated in the tropical countries of Asia and Africa, due to its socio-economic and environmental importance (Balogoun et al., 2014; Ruf et al., 2019). The fruit of cashew nut is mainly used in the food industries (Das & Arora, 2017). The cashew nut shell is increasingly used to produce biofuel (Gwoda et al., 2024). In 2020, the United Nations Food and Agriculture Organisation (FAO) estimated that world production of raw cashew nuts amounted to 4,180,000 tonnes harvested from 7,100,000 ha, with West Africa being the main producer (Chen et al., 2023).
2In Côte d'Ivoire, the area under cashew trees has increase from around 8,000 ha when this crop was introduced in 1960 as a reforestation plant in the north part of the country to more than 1,400,000 ha in 2024 (FIRCA, 2024). This dynamic of cashew tree adopting by farmers was largely due to the commercial value of the cashew nut fruit, but also to the plant's hardiness and low soil and climate requirements (Lautié et al., 2001; Bassett et al., 2018). As a result, cashew is currently the main perennial cash crop in the northern part of the country, thereby contributing for 58% to its agricultural income (Koffi & Oura, 2019). Consequently, since 2015, Côte d'Ivoire has been the world's leading producer of raw cashew nuts, with production of more than 1,000,000 tonnes in 2022 (Ouattara, 2017; FIRCA, 2024).
3However, this high production is essentially based on an increased area under cultivation. Indeed, cashew orchards in Côte d’Ivoire are facing a number of major agronomic and phytosanitary constraints (Kouakou et al., 2018; Ouali N'Goran et al., 2020; Soro et al., 2020a), that limit its yields to around 450 kg/ha compared with the 1,200 kg.ha-1 at experimental stations (Djaha et al., 2010). Most of the current orchards in Côte d'Ivoire were created on the basis of 'individual' cultivation practices by farmers, without any substantial technical or financial support from the State (Ruf et al., 2019). In this regard, Kambou et al. (2019) reported high cashew tree densities, reaching eight times the Ivorian standard of 100 plants.ha-1. According to a study by Soro et al. (2020a) carried out in 250 plantations covering 43 cashew-growing localities in Côte d'Ivoire, 98% of orchards have been established by direct sowing, and only 6% of growers thinned their orchards to reach the standard of 100 plants.ha-1. Meanwhile, they observed a significant influence of the diversity of cultural practices on insect damage, but their analyses did not focus specifically on one pest. In addition, several studies (Thomas et al., 2017; Beaumelle et al., 2021) highlighted the influence of the agricultural landscape on the dispersal and abundance of arthropod crop pests.
MATERIALS AND METHODS
Sampling sites selection and description
4The study was conducted in the localities of Bondoukou and Bouna, approximately 180 km apart, in the northeast of Côte d'Ivoire (Figure 1). In Bondoukou, the climate is transitional equatorial, with four seasons: a long and short rainy season from April to June and September to October respectively; a long and short dry season from November to March and July to August respectively. The monthly temperature ranged between 20 - 33 °C, with an annual rainfall of 942 - 1625 mm. The climate in Bouna is wet and dry Sudanese, with one rainy season from March to October and one dry season from November to February. The monthly temperature ranged between 20 - 35 °C, with an annual rainfall of 992 - 1383 mm (Kouakou et al., 2017). The natural vegetation consists of forest and wooded savannah in Bondoukou, and shrub and tree savannahs in Bouna (Guillaumet & Adjanohoun, 1971).
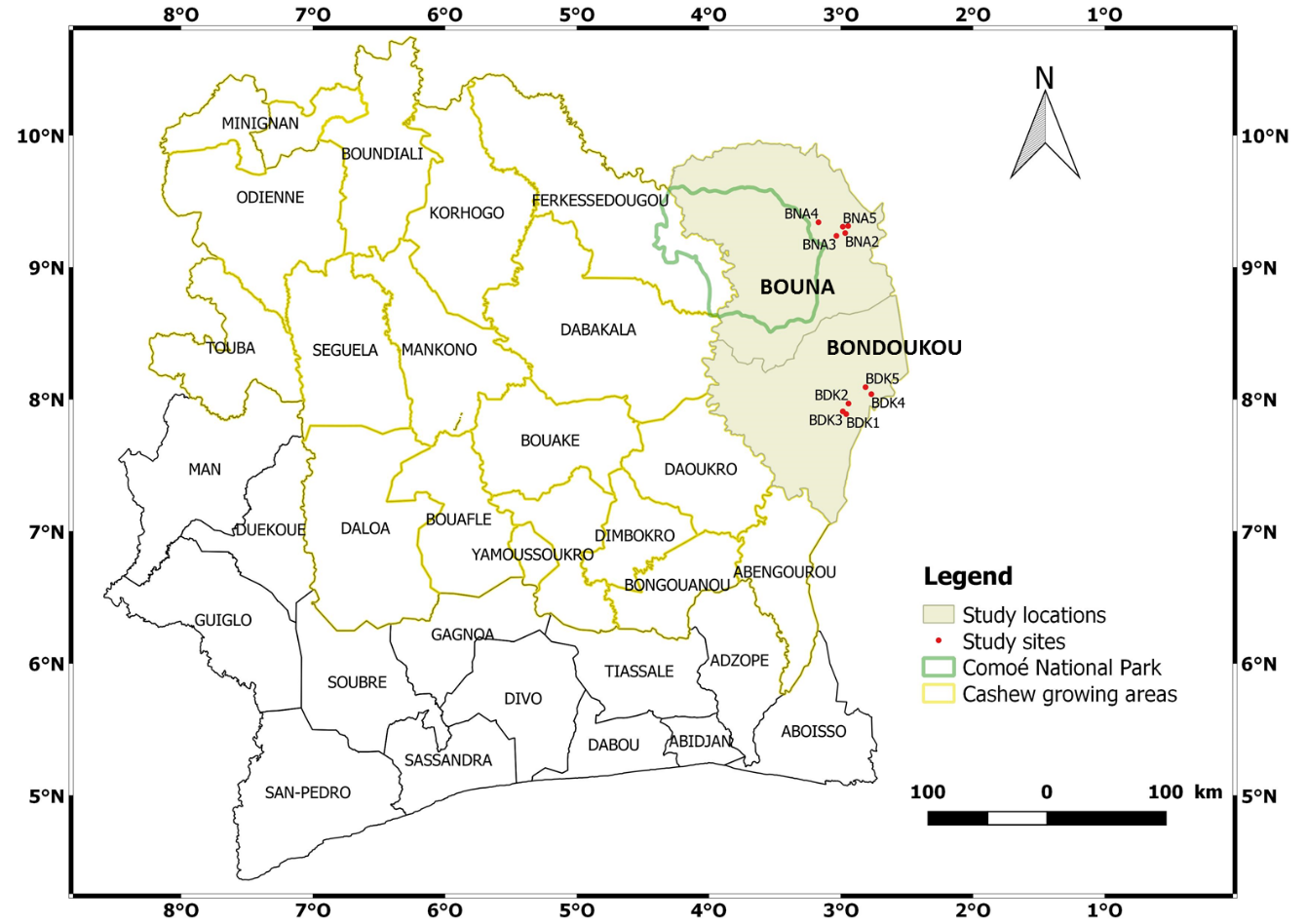
Figure 1: Map of Côte d’Ivoire showing the study location
5In each locality, five study sites of at least 2 ha in size, 15 to 20 years old, infested by A. terebrans but not subject to chemical insecticide applications (at least two years prior to the study) were selected. All these sites were planted in bulk by direct sowing, using unimproved seeds. The systematic sampling using quadrats was used because it allows the density and heterogeneity of cashew trees to be assessed. This sampling method is also used to assess the spatial distribution of A. terebrans infestations and the effect of cashew tree density on these infestations. In each site, a plot of 100 m x 100 m (i.e., 1 ha) was delimited, starting from an edge. This plot was then subdivided into 13 quadrats of 20 m x 20 m (i.e., 400 m2) at 20 m intervals. All cashew trees in the 13 quadrats were then numbered, marking a total sample of 1924 cashew trees in both localities. Distance between two study sites was approximately 2.5 to 30 km, and each of them was geo-referenced by Global Positionning System (Table 1).
Table 1: Geographical coordinates and age of the study sites
 Assessment of cashew density
Assessment of cashew density
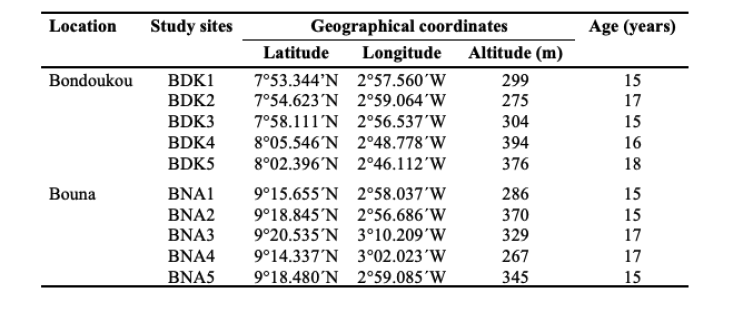
6Cashew density is the ratio of the total number of cashew trees in a quadrat to the area of that quadrat. The density of cashew trees in each quadrat was determined by counting the cashew trees in the 13 quadrats of each study site, i.e., a total area of 0.52 ha. The density per hectare was determined by extrapolation. The variation in the number of cashew trees among the 13 quadrats of each study site was assessed using the coefficient of variation (CV):
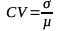
7With σ and µ representing the standard deviation and the mean of the number of cashew trees per quadrat of a study plot, respectively.
8Given the cryptic life of A. terebrans, the monitoring of its infestations was done through the galleries bored inside the cashew tree (Agboton et al., 2017). The presence of the insect inside the cashew tree is characterised by the presence of fresh sawdust from the cashew wood at the gallery opening, known as the 'fresh entry hole (FEH)', and under the infested cashew tree (Figure 2).
9Thus, in this study, a FEH is equivalent to one infestation of A. terebrans. For two consecutive years, from July 2018 to June 2020 (i.e., two cashew production seasons), the 1924 marked cashew trees were inspected once at the end of each month, in order to count all the FEH present on each infested cashew tree.

Figure 2: Illustration of Apate terebrans infestations on a live cashew tree. (A) A. terebrans at the entrance to the fresh hole; (B) Sawdust under an infested cashew tree; (C) Gallery of A. terebrans
Assessment of the spatial distribution of cashew trees infested by A. terebrans
10The contiguous quadrat method was used to analyse the spatial distribution of infested cashew trees at plot level (Agbangla et al., 2015). This method is based on the examination of the mean and variance of the number of infested cashew trees per quadrat (Jayaraman, 1999). The type of distribution of infestations (aggregate, regular or random) was determined using the Blackman (DIB) and Morisita (DIM) dispersion indices (Amaral et al., 2014; Agbangla et al., 2015; Hayes & Castillo, 2017). The simultaneous use of these two indices aimed at confirming the result obtained by each of the two indices.
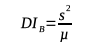
11With s2 and µ representing the variance and mean of the number of infested cashew trees per quadrat, respectively, in each study site.
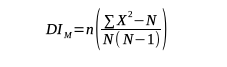
12With n: total number of quadrats; X: number of infested cashew trees in a single quadrat; ∑X2: sum of all X2 values; N: total number of infested cashew trees in the 13 quadrats of each study site.
13For both dispersion index models, DI values greater than 1 indicate that the distribution of infestation is aggregated, while DI values less than 1 indicate a regular distribution of infestation. The distribution is random when DI = 1 (Canard & Poinsot, 2004).
Collection of climatic data
14Two thermo-hygrometers (EasyLob EL-USB-2) installed in each area, close to the study sites, were used to simultaneously record temperature and relative air humidity during the study.
Data analysis
15The collected data were processed and analysed using Excel 2013 and RStudio 3.6.2 (Rstudio Team, 2019).
16The coefficient of variation was used to determine the variation in the number of cashew trees among the 13 quadrats in each study site. Kruskal Wallis test was used, after verification of normality (Shapiro’s test) and homogeneity of variances (Bartlett’s test), to compare cashew tree densities of the different study sites in each locality. Wilcoxon post hoc test was used for pairwise comparisons. For the comparison of this variable between the two localities, the student’s t-test was used, after testing for normality (Shapiro's test) and homogeneity of variance (Levene's test).
17Kruskal Wallis test was also used to compare the average number of Apate terebrans infestations of the different study sites in each locality, considering the quadrat as a replicate. For each study site, the proportions of infestations in the 13 quadrats were compared using Fisher's test, after verification of Cochran's rule.
18Simple linear regression was used to assess the effect of cashew tree density on infestation by A. terebrans. To do this, cashew tree densities of the 13 quadrats of each study site were related to the infestations counted in these quadrats. However, in order to limit the influence of the shade of large trees (other than cashew trees), the quadrats in which they were located were not used in this analysis. Thus, in one study site, the maximum number of quadrats not taken into account was three.
19The Chi-square test (χ2) for α = 5% was used to test whether the dispersion indices values are significantly different from 1 (random dispersion), considering:
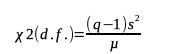
20With d.f. (degree of free) = q − 1, q being the number of quadrats in each study site, s2 and µ representing the variance and the mean number of infested cashew trees per quadrat in each study site, respectively (David & Moore, 1954).
RESULTS
Cashew tree densities
21The average cashew tree density varied significantly among the five study sites in each area (d.f. = 4; p < 0.01) (Figure 3). A mean (± standard deviation) of 515 ± 235.5 cashew trees.ha-1 was obtained in Bondoukou, which was significantly higher than the density of 235 ± 39.6 cashew trees.ha-1 in Bouna.
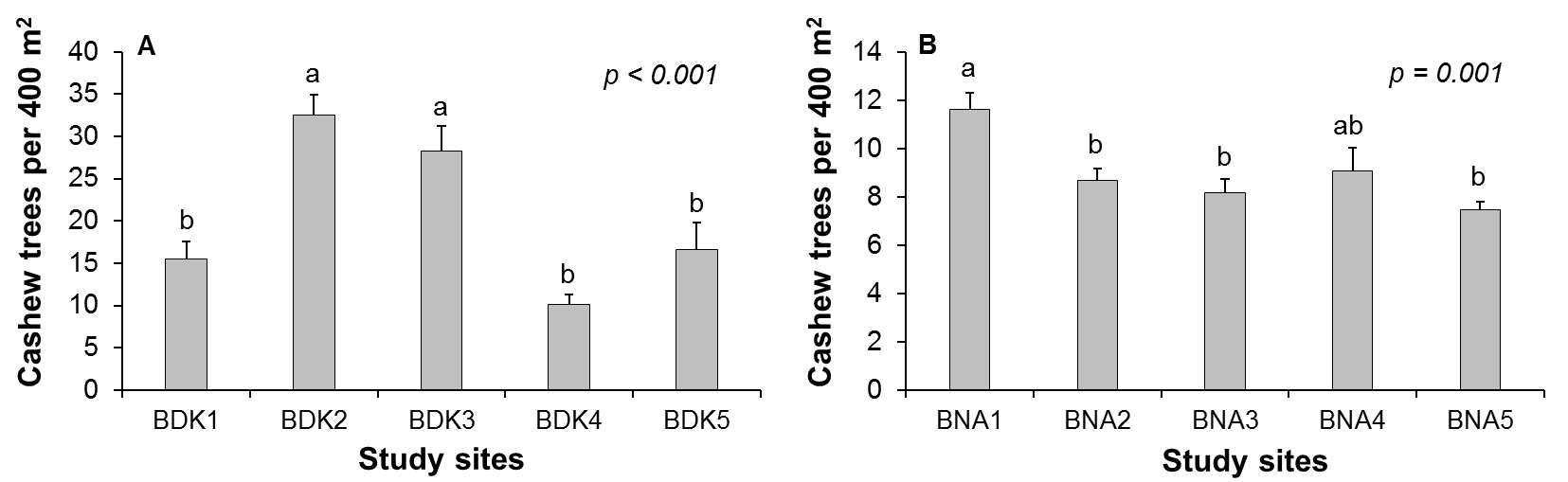
Figure 3: Average (± SD) density of cashew trees within the 400 m2 quadrats of study sites in Bondoukou (A) and Bouna (B) during the 2018 to 2020 production seasons in Côte d’Ivoire. Significant differences are indicated by different letters (p < 0.05)
22The coefficient of variation (CV) of cashew tree density in the 13 quadrats of the study sites ranged from 26.67 to 70.34% in Bondoukou, and from 19.58 to 38.0% in Bouna. At the scale of the locality, the CV of cashew tree density was 58.64% in Bondoukou and 29.44% in Bouna. Thus, the density of cashew trees within the Bondoukou study sites appears more heterogeneous than that of Bouna (Figure 4).
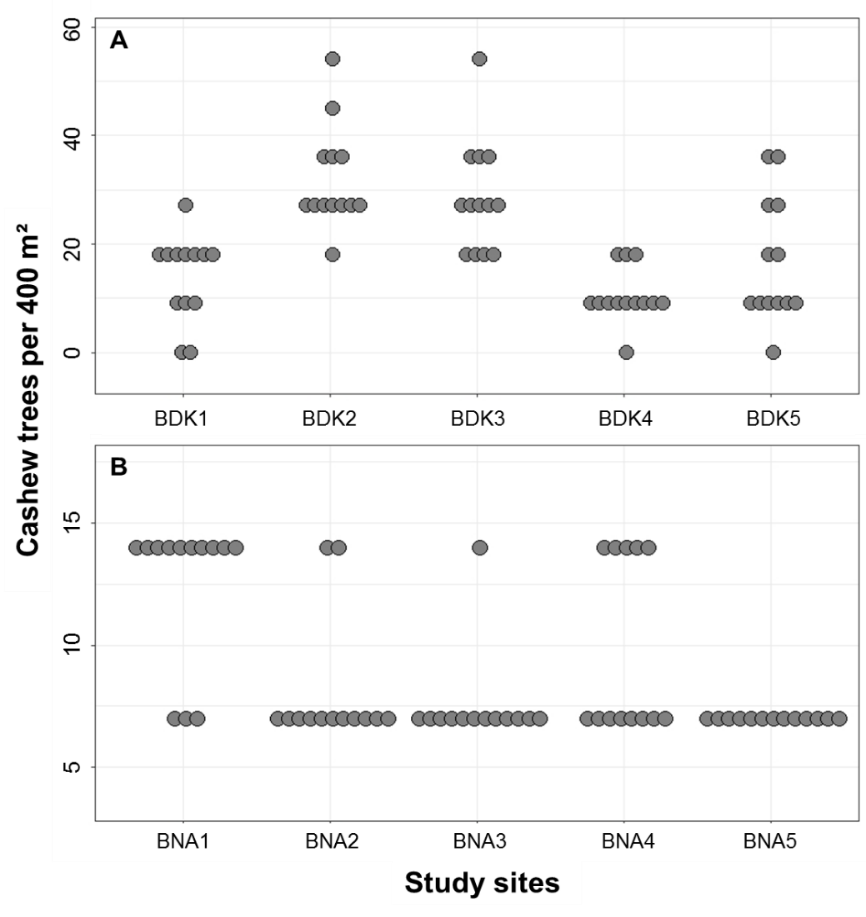
Figure 4: Variation in the number of cashew trees within the 13 quadrats in the study sites in Bondoukou (A) and Bouna (B) during the 2018 to 2020 production seasons in Côte d’Ivoire
Abundance of A. terebrans infestations
23In each locality, the number of infestations varied significantly among the five study sites (Figure 5). In Bondoukou, mean (± standard deviation) of 99.2 ± 37.4 infestations and 14 ± 3.87 infested cashew trees (ICTs) per study site were observed in the first sampling year, compared with 73.8 ± 45.85 infestations and 7.2 ± 2.95 ICTs per study site in Bouna, during the same period. In the second year of sampling, the means were 101 ± 47.88 infestations and 9.8 ± 7.39 ICTs in Bondoukou, and 61 ± 57.76 infestations and 6.4 ± 6.54 ICTs in Bouna.
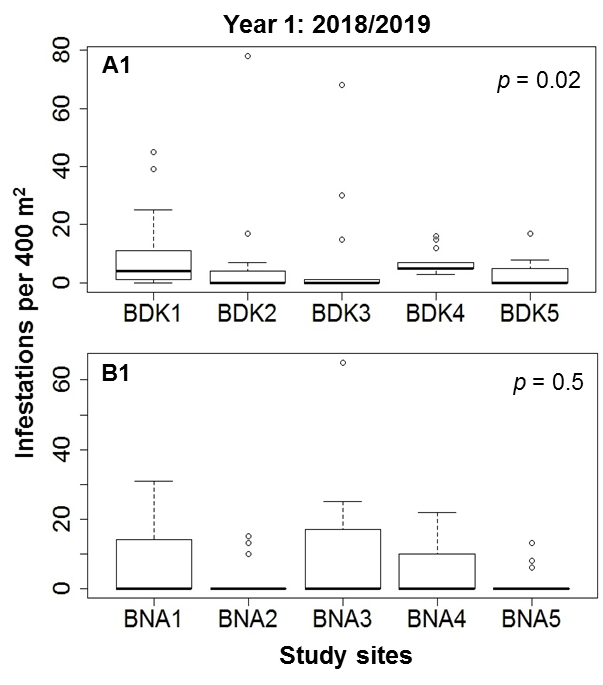
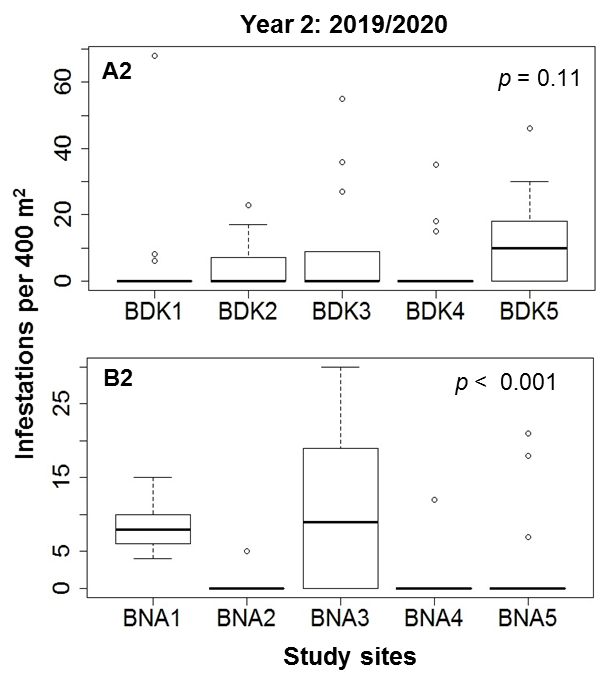
Figure 5: Comparison of infestation abundance among study sites in Bondoukou (A1, A2) and Bouna (B1, B2) during two cashew crop seasons (2018/2019 and 2019/2020) in Côte d’Ivoire
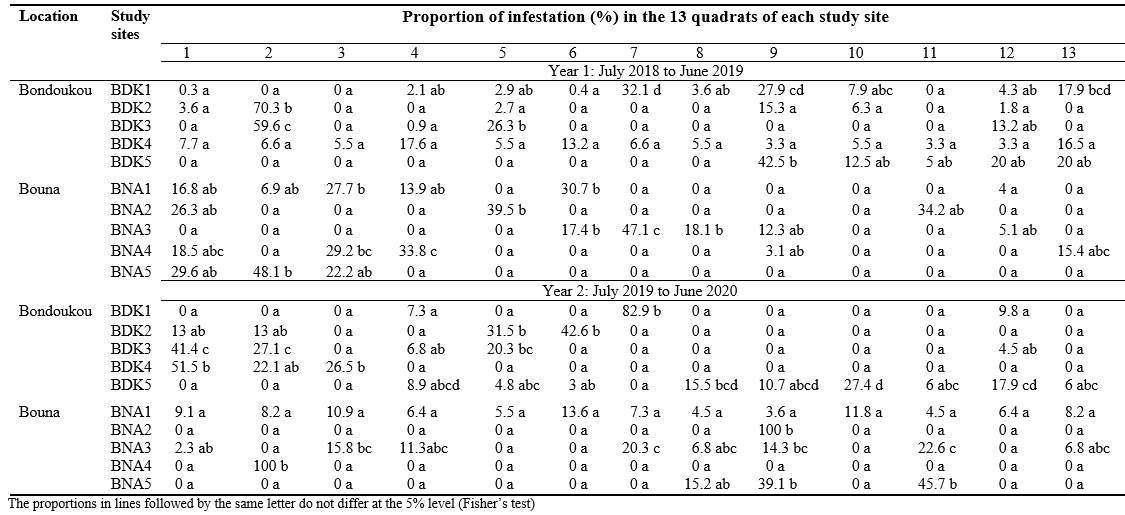
24The proportion of infestations varied very significantly between the 13 quadrats at each study site in Bondoukou and Bouna (d.f. = 12; p < 0.0001). This proportion varied from 0 to 100% for one quadrat (Table 2).
Table 2: Comparison of proportions of Apate terebrans infestation in the 13 quadrats of each study site in Bondoukou and Bouna during two cashew cropping seasons (2018/2019 and 2019/2020) in Côte d’Ivoire
 Effect of cashew tree density on the abundance of A. terebrans infestations
Effect of cashew tree density on the abundance of A. terebrans infestations
25Linear regression analysis showed that high cashew density negatively influenced infestation abundance (p < 0.05) in both localities. The relationship between these two variables was moderate and significant for four study sites in Bondoukou (BDK1, BDK2, BDK3 and BDK4) and two study sites in Bouna (BNA1 and BNA3). However, for a given site, the relationship between cashew tree density and infestation abundance was only significant for the first year of sampling (Table 3).
Table 3: Relationship between cashew tree density and the abundance of Apate terebrans infestations in Bondoukou and Bouna during two cashew cropping seasons (2018/2019 and 2019/2020) in Côte d’Ivoire
|
Location |
Study sites |
Regression parameters |
|||
|
Linear regression |
R2 |
p-value |
d.f. |
||
|
Year 1: July 2018 to June 2019 |
|||||
|
Bondoukou |
BDK1 |
y = - 1.54x + 35.37* |
0.52 |
0.02 |
8 |
|
BDK2 |
y = - 1.32x + 50.90* |
0.46 |
0.02 |
9 |
|
|
BDK3 |
y = - 0.88x + 36.11* |
0.55 |
0.05 |
9 |
|
|
BDK4 |
y = - 0.76x + 14.69* |
0.53 |
0.005 |
11 |
|
|
BDK5 |
y = - 0.17x + 06.71 |
0.30 |
0.08 |
9 |
|
|
Bouna |
BNA1 |
y = - 1.98x + 31.99 |
0.15 |
0.21 |
10 |
|
BNA2 |
y = 0.65x - 02.40 |
0.03 |
0.57 |
10 |
|
|
BNA3 |
y = - 3.75x + 42.54 |
0.13 |
0.28 |
9 |
|
|
BNA4 |
y = 1.34x - 7.16 |
0.29 |
0.09 |
9 |
|
|
BNA5 |
y = 1.33x - 7.88 |
0.11 |
0.29 |
10 |
|
|
Year 2: July 2019 to June 2020 |
|||||
|
Bondoukou |
BDK1 |
y = - 1.75x + 32.93 |
0.33 |
0.08 |
8 |
|
BDK2 |
y = 0.21x - 1.53 |
0.03 |
0.58 |
9 |
|
|
BDK3 |
y = - 0.48x + 26.04 |
0.08 |
0.40 |
9 |
|
|
BDK4 |
y = - 0.2x + 7.21 |
0.01 |
0.80 |
11 |
|
|
BDK5 |
y = 0.61x + 4.34 |
0.28 |
0.09 |
9 |
|
|
Bouna |
BNA1 |
y = - 1.26x + 23.40* |
0.68 |
0.001 |
10 |
|
BNA2 |
y = 0.26x - 1.78 |
0.18 |
0.17 |
10 |
|
|
BNA3 |
y = - 4x + 40* |
0.58 |
0.007 |
9 |
|
|
BNA4 |
y = 0.17x - 0.58 |
0.04 |
0.57 |
9 |
|
|
BNA5 |
y = 2.26x - 13.29 |
0.10 |
0.31 |
10 |
|
26The equation is given by y = ax + b, where y is the abundance of infestations, x is the cashew density factor, a is the directing coefficient and b is the intercept. *Asterisks indicate a significant regression at the 5% level
Effect of cashew tree density on the spatial distribution of A. terebrans infestations
27The plot distribution of infestations by A. terebrans was aggregated in both areas. The distribution of infested cashew trees (ICTs) varied from one study site to another. In Bondoukou, three aggregative, one random and one regular distributions of ICTs were observed in each year (p < 0.05). In contrast, no site had an aggregative distribution in Bouna (Table 4). At the locality scale, the distribution of ICTs was aggregative in Bondoukou (Year 1: DIB = 1.49; DIM = 1.40; Chi2 = 81.88; Year 2: DIB = 2.41; DIM = 2.62; Chi2 = 132.71; with d.f. = 55; p = 0.05). In Bouna, the distribution was random (Year 1: DIB = 1.30; DIM = 1.56; Chi2 = 74.13; Year 2: DIB = 1.15; DIM = 1.21; Chi2 = 65.44; with d.f. = 57; p = 0.05).
Table 4: Distribution indices of cashew trees infested by Apate terebrans in Bondoukou and Bouna during two cashew cropping seasons (2018/2019 and 2019/2020) in Côte d’Ivoire
|
Location |
Study sites |
Distribution indices |
||||
|
DIB |
DIM |
d.f. |
Chi2 |
Repartition |
||
|
Year 1: July 2018 to June 2019 |
||||||
|
Bondoukou |
BDK1 |
1.18 |
1.10 |
9 |
10.64 |
Random |
|
BDK2 |
2.52 |
2.02 |
10 |
25.25 |
Aggregative |
|
|
BDK3 |
1.91 |
2.14 |
10 |
19.11 |
Aggregative |
|
|
BDK4 |
0.12 |
0.25 |
12 |
1.46 |
Regular |
|
|
BDK5 |
2.2 |
2.2 |
10 |
22 |
Aggregative |
|
|
Bouna |
BNA1 |
1.45 |
1.45 |
11 |
16 |
Random |
|
BNA2 |
1.51 |
2.4 |
11 |
16.6 |
Random |
|
|
BNA3 |
0.87 |
0.73 |
10 |
8.67 |
Regular |
|
|
BNA4 |
1.40 |
1.57 |
10 |
14 |
Random |
|
|
BNA5 |
1.51 |
2.4 |
11 |
16.6 |
Random |
|
|
Year 2: July 2019 to June 2020 |
||||||
|
Bondoukou |
BDK1 |
1.22 |
1.66 |
9 |
11 |
Random |
|
BDK2 |
1.95 |
2.36 |
10 |
19.5 |
Aggregative |
|
|
BDK3 |
3.01 |
2.68 |
10 |
30.15 |
Aggregative |
|
|
BDK4 |
0.83 |
0 |
12 |
10 |
Regular |
|
|
BDK5 |
2.04 |
1.58 |
10 |
20.38 |
Aggregative |
|
|
Bouna |
BNA1 |
0 |
0 |
11 |
0 |
Regular |
|
BNA2 |
0.91 |
0 |
11 |
10 |
Regular |
|
|
BNA3 |
1.6 |
1.6 |
10 |
16 |
Random |
|
|
BNA4 |
1.53 |
3.67 |
10 |
15.33 |
Random |
|
|
BNA5 |
1.51 |
2.4 |
11 |
16.6 |
Random |
|
28DIB and DIM: respectly Blackman and Morisita dispersion indices
Climatic conditions
29The average (± standard deviation) monthly temperatures during years 1 and 2 were 29.28 ± 1.79 °C and 27.70 ± 1.84 °C in Bondoukou, compared to 30.30 ± 1.71 °C and 29.30 ± 1.98 °C in Bouna. Those of hygrometry were 65.93 ± 12.69% and 71.16 ± 14.09% in Bondoukou, against 52.18 ± 24.68% and 58.26 ± 22.82% in Bouna.
DISCUSSION
30The significant differences between cashew densities are mainly due to the implementation of a 'national cashew thinning programme' by farmers in Bouna as opposed to Bondoukou. This programme, initiated by the government of Côte d'Ivoire, aims at reducing cashew trees density in the orchards to a standard of 100 cashew trees.ha-1, which would optimise the yield of the orchards (FIRCA, 2024). The application of this programme therefore creates a certain homogeneity of cashew tree density at plot level, which is reflected in a low coefficient of variation in Bouna, unlike in Bondoukou. Furthermore, the wide variation in cashew density within the original orchards, as is the case in Bondoukou, is mainly the result of the historical context of the establishment of the crop. Ivorian orchards were generally created in bulk, without staking, and with a view to protecting the environment (Ruf et al., 2019). This patch heterogeneity is known to influence insect dynamics (Alford et al., 2018; Snyder et al., 2021).
31We believe that relative humidity is relatively higher in high-density cashew quadrats than in low-density ones, as reported by Yang et al. (2021) in China. They have observed that the high rice density created a relatively humid microclimate that was more favourable to leafhoppers and planthoppers. The low abundance of A. terebrans in high-density cashew quadrats would be justified by the negative effect of humidity on bostrichids (Nansen et al., 2002; Hodges et al., 2003), in particular on A. terebrans (Aliko & Ouali N’Goran 2022). Similar results were obtained by Krob et al. (2022) in Philippines, who showed that high cotton density reduced trips damage. In addition, bostrichids prefer wood with relatively low moisture content (Ivie, 2002; Cookson, 2004). Several authors (Thackeray et al., 2016; Falk et al., 2018) have shown a direct and/or indirect (e.g., physiological changes in the host plant) influence of climatic and biotic factors on the temporal dynamics of insects. Furthermore, in the present study, the relationship between cashew tree density and infestations by A. terebrans was found to be influenced by other factors, including the border effects in the high cashew density quadrats. Indeed, a large number of infestations were sometimes recorded at the edge of these quadrats, especially when they were contiguous to low cashew density quadrats, or when the edge of these quadrats was less shaded. The border effects on insects have been mentioned by Amala & Shivalingaswamy (2018). Otherwise, the weak relationship observed between A. terebrans infestations and cashew tree density during the second year of sampling could be justified by the induced defences of infested cashew trees in the first year of sampling (Boone et al., 2011; Goiana et al., 2019). Indeed, it was observed that 74% of cashew trees attacked by A. terebrans in the first year of sampling were no longer attacked in the second year. This observation is in accordance with Agboton et al. (2017). In addition, given that there are different cashew varieties within each study site, the preference of one variety could influence the relationship between cashew density and infestation (Schröder et al., 2015; Reisenman et al., 2016).
32An aggregated distribution of A. terebrans infestations was observed in the study sites in both localities, as reported by Agboton et al. (2017) in cashew orchards in Benin. This type of distribution is known for several bostrichids such as Prostephanus truncatus (Horn 1878) and Rhyzopertha dominica (Fabricius 1792) (Nansen et al., 2002; Steel et al., 2012). According to Jayaraman (1999), Canard & Poinsot (2004), several causal mechanisms such as environmental heterogeneity, gregarious behaviour and breeding mode, could explain such a distribution. For example, it has been reported that R. dominica males release two aggregation pheromones, Dominicalure-1 and Dominicalure-2, which attract conspecifics to located food resources (Bashir et al., 2003; Edde et al., 2007). As for the distribution of cashew trees infested by A. terebrans, it was globally aggregated in Bondoukou and random in Bouna. These two different types of distribution would be the result of the plot structure of the cashew trees. Indeed, the heterogeneity of plots within the plot influences the dynamics of crop pest (Wang et al., 2023). Therefore, the large heterogeneity of cashew trees observed in Bondoukou would strongly justifies the aggregative distribution of infested cashew trees. The insect mainly attacks cashew trees in quadrats characterised by a low density of wood. In Bouna, the microclimate being relatively homogeneous, the spatial barrier appears reduced for the pest. As a result, the pest is dispersed throughout the plot in a more or less homogeneous manner. These results confirm those of Yéo et al. (2019) on the spatial distribution of cashew trees infested by A. terebrans in Bondoukou and Bouna, in Côte d’Ivoire.
CONCLUSIONS
33This study showed a large heterogeneity in cashew tree density in Bondoukou and Bouna (north-east of Côte d'Ivoire), which significantly influenced the abundance and patchy distribution of A. terebrans infestations. We recommend that entomological studies in cashew orchards planted in bulk take into account the existence of heterogeneity in cashew tree density and its probable influence on insects. Further studies should be carried out to determine the optimum spacing among cashew trees, as well as on the varietal resistance of cashew trees, with a view to sustainably controlling A. terebrans infestations and improving cashew tree productivity.
Acknowledgements
The authors are grateful to the African Centre of Excellence on Climate Change, Biodiversity and Sustainable Agriculture, the University Agency of the Francophony (AUF), the Research Institute for Development (IRD), the International Foundation for Science (IFS) and the Scientific and Technological Cooperation Committee (COMSTECH) of the Organization of Islamic Cooperation (OIC) for their financial support. They are also grateful to the technician supervising the fieldwork, Mr. DUA Fofié Daniel, as well as to the various farmers who made their plots available.
Bibliographie
Agbangla M.M., Aoudji A.K.N., Akouehou G.S., Gbetoho J.A., Samon K., Ayina O., Cannière C. & Ganglo J.C., 2015. Structural and Ecological Characteristics of Commercial Timber species' population: a Basis for Silviculture in Niaouli Forest Stands (Southern Benin). Tropicultura, 33(3), 238-252.
Agboton C., Onzo A., Ouessou F.I., Goergen G., Vidal S. & Tamò M., 2014. Insect fauna associated with Anacardium occidentale (Sapindales, Anacardiaceae) in Benin, West Africa. Journal of Insect Science, 14(229). https://doi.org/10.1093/jisesa/ieu091
Agboton C., Onzo A., Korie S., Tamò M. & Vidal S., 2017. Spatial and Temporal Infestation Rates of Apate terebrans (Coleoptera: Bostrichidae) in Cashew Orchards in Benin, West Africa. African Entomology, 25(1), 24-36.
Alford L., Tougeron K., Pierre J.-S., Burel F., & Baaren J.v., 2018. The effect of landscape complexity and microclimate on the thermal tolerance of a pest insect. Insect Science, 25, 905–915. https://doi.org/10.1111/1744-7917.12460
Aliko Y.J. & Ouali N’Goran S.-W.M., 2022. Seasonal Fluctuation of The Populations of The Stem Borer, Apate terebrans (Coleoptera: Bostrichidae) in Cashew Orchards in North-Eastern Côte d'Ivoire (West Africa). Egyptian Academic Journal of Biological Sciences, 15(4), 1-12. https://doi.org/10.21608/eajbsa.2022.265487
Amala U. & Shivalingaswamy T.M., 2018. Effect of intercrops and border crops on the diversity of parasitoids and predators in agroecosystem. Egyptian Journal of Biological Pest Control, 28, 11. https://doi.org/10.1186/s41938-017-0015-y
Amaral M.K., Péllico Netto S., Lingnau C. & Figueiredo Filho A., 2014. Evaluation of the Morisita index for determination of the spatial distribution of species in a fragment of araucaria forest. Applied Ecology and Environmental Research, 13(2), 361-372. DOI: 10.15666/aeer/1302_361372
Balogoun I, Saïdou A., Ahoton E.L., Amadji L.G., Ahohuendo C. B., Adebo I.B., Babatounde S., Chougourou D., Adoukonou-Sagbadja H. & Ahanchede A., 2014. Characterization of cashew’s production systems in the main cropping zones of Benin. Agronomie Africaine, 26(1), 9-22.
Bashir T., Hodges R.J., Birkinshaw L.A., Hall D.R. & Farman D.I., 2003. Phenotypic plasticity of Rhyzopertha dominica pheromone signaling: the effects of different hosts and presence of conspecific females on male produced aggregation pheromone. Journal of Chemical Ecology, 29, 945-959.
Bassett T.J., Koné M. & Pavlovic N.R., 2018. Power Relations and Upgrading in the Cashew Value Chain of Côte d’Ivoire. Development and Change, 49(5), 1223–1247. DOI: 10.1111/dech.12400
Beaumelle L., Auriol A., Grasset M., Pavy A., Thiéry D. & Rusch A., 2021. Benefits of increased cover crop diversity for predators and biological pest control depend on the landscape context. Ecological Solutions and Evidence, 2, e12086. https://doi.org/10.1002/2688-8319.12086
Boone C.K., Brian H. Aukema B.H., Bohlmann J., Carroll A.L. & Raffa K.F., 2011. Efficacy of tree defense physiology varies with bark beetle population density: a basis for positive feedback in eruptive species. Canadian Journal of Forest Research, 41(6), 1174-1188. https://doi.org/10.1139/x11-041
Canard A. & Poinsot D., 2004. Some typical statistical methods for studying populations and stands using the quadrat method. Technical data sheet, University of Rennes 1, France, 34 p.
Chen Y.-Y., Li N.-y., Guo X., Huang H.-j., Garcia-Oliveira P., Sun J.-j., Zhang J.-D., Prieto M.A., Guo Z.-q. & Liu C., 2023. The nutritional and bio-active constituents, functional activities, and industrial applications of cashew (Anacardium occidentale): A review. Food Frontiers, 4, 1606-1621. https://doi.org/10.1002/fft2.250
Cookson L.J., 2004. Treatment Methods for the Protection of Hardwood Sapwood from Lyctine Borers. Forest and Wood Products Research and Development Corporation, Australia, 23 p.
Das I. & Arora A., 2017. Post-harvest processing technology for cashew apple – A review. Journal of Food Engineering, 194, 87–98. https://doi.org/10.1016/j.jfoodeng.2016.09.011
David F.N. & Moore P.G., 1954. Notes on contagious distribution in plant populations. Annals of Botany, 18, 47-53.
Deguine J.-P., Aubertot J.-N., Flor R.J., Lescourret F., Wyckhuys K.A.G. & Ratnadass A., 2021. Integrated pest management: good intentions, hard realities. A review. Agronomy for Sustainable Development, 41, 38. https://doi.org/10.1007/s13593-021-00689-w
Djaha A.J.-B., N’guessan A.K., Ballo C.K. & Aké S., 2010. Seed germination of two elite cashew trees (Anacardium occidentale L.) intended for use as rootstock in grafting operations in Côte d’Ivoire. Journal of Applied Biosciences, 32, 1995-2001.
Edde P.A., Phillips T.W., Robertson J.B. & Dillwith J.W., 2007. Pheromone Output by Rhyzopertha dominica (Coleoptera: Bostrichidae), as Affected by Host Plant and Beetle Size. Annals of the Entomological Society of America, 100(1), 83-90.
Fahrig L., Baudry J., Brotons L., Burel F.G., Crist T.O., Fuller R.J., Sirami C., Siriwardena G.M. & Martin J.L., 2011. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecology Letters, 14, 101-112.
Falk M.A., Lindroth R.L., Keefover-Ring K. & Raffa K.F., 2018. Genetic variation in aspen phytochemical patterns structures windows of opportunity for gypsy moth larvae. Oecologia, 187, 471–482. https://doi.org/10.1007/s00442-018-4160-0
FIRCA, 2024. Fonds Interprofessionnel pour la Recherche et le Conseil Agricoles, https://firca.ci/blog/le-firca-et-la-filiere-anacarde/, (30/04/2024).
Goiana E.S.S., Dias-Pini N.S., Muniz C.R., Soares A.A., Alves J.C., Vidal-Netoa F.C. & Da Silva C.S.B., 2019. Dwarf-cashew resistance to white fly (Aleurodicus cocois) linked to morphological and histochemical characteristics of leaves. Pest Management Science, 76, 464–471. DOI: 10.1002/ps.5531
Guillaumet J.L. & Adjanohoun E., 1971. La végétation. In: Le milieu naturel de la Côte d'Ivoire. O.R.S.T.O.M. n° 50, 157–263.
Gwoda S., Valette J., Sidibé S.S., Piriou B., Blin J. & Ouédraogo I.W.K., 2024. Use of cashew nutshell liquid as biofuel blended in diesel: Optimisation of blends using additive acetone–butanol–ethanol (ABE (361)). Chemical Engineering Science, 9, 100117. https://doi.org/10.1016/j.clce.2024.100117
Hayes J.J. & Castillo O., 2017. A New Approach for Interpreting the Morisita Index of Aggregation through Quadrat Size. International Journal of Geo-Information, 6, 296. https://doi.org/10.3390/ijgi6100296
Hodges R.J., Addo S. & Birkinshaw L., 2003. Can observation of climatic variables be used to predict the flight dispersal rates of Prostephanus truncatus? Agricultural and Forest Entomology, 5, 123–135.
Ivie M.A., 2002. Bostrichidae Latreille 1802. In: Arnett R.H. Jr., Thomas M.C., Skelley P.E. & Frank J.H. Editor, 2002. American Beetles, Volume 2, CRC Press, Boca Raton, pp. 233-244.
Jayaraman K., 1999. Manuel de statistique pour la recherche forestière. Kerala Forest Research Institute Peechi, Thrissur, Kerala (Inde), 242 p.
Kambou D., Koné T., Soumahoro B.A., Toure Y., Kassi K.F.J.-M., Tuo S., Koné D. & Koné M., 2019. Structuring of the agricultural population of the cashew nut sector (Anacardium occidentale (L.) Anacardiaceae) and characterization of plantations in the Bounkani and Gontougo regions of Côte d’Ivoire. International Journal of Innovative and Applied Research, 26(4), 1159-1169.
Koffi S.Y. & Oura K.R., 2019. The factors of cashew nuts adoption in the cotton basin of Côte d’Ivoire. Cahiers Agricultures, 28(24), 1-8. https://doi.org/10.1051/cagri/2019025
Kouadio K.F., Akesse E.N., Ouali N’Goran S.-W.M. & Koné D., 2023. Distribution and damage of Diastocera trifasciata (Coleoptera: Cerambycidae) and Apate terebrans (Coleoptera: Bostrichidae), major insect pests of the cashew tree in Côte d'Ivoire. Journal of Applied Biosciences, 186, 19588-195600. https://doi.org/10.35759/JABs.186.5
Kouakou K.E. et al., 2017. Redefinition of homogeneous climatic zones in Cote d’Ivoire in a context of climate change. International Journal of Engineering Research & Technology, 8(11), 453-462.
Kouakou C.K., Konan A.D.S.-H., N’Da Adopo A., N’Da H.A., Djaha A.J.-B., Minhibo M.Y., Djidji A.H., Dosso M. & N’Guessan A.E., 2018. Agro-morphological characterization of cashew (Anacardium occidentale L.) ecotypes from North-Central of Côte d’Ivoire. Fruits, 73(5), 255–264. https://doi.org/10.17660/th2018/73.5.1
Krob J.L., Stewart S.D. & Brown S.A., 2022. Effects of plant density, seed spacing, and seed treatment on thrips injury to cotton. Crop Protection, 161, 106059. https://doi.org/10.1016/j.cropro.2022.106059
Lautié E., Dornier M., Filho M.Z. & Reynes M., 2001. The cashew tree and its products: main characteristics and possible uses. Fruits, 56, 235-248.
Lindell C.A., Irish-Brown A., Rothwell N.L. & Wallis A.E., 2023. Pest and disease risk and management in high-density perennial crops: Current knowledge and areas of future research. Crop Protection, 165, 106150. https://doi.org/10.1016/j.cropro.2022.106150
Nansen C., Meikle W.G. & Korie S., 2002. Spatial Analysis of Prostephanus truncatus (Bostrichidae: Coleoptera) Flight Activity Near Maize Stores and in Different Forest Types in southern Benin, West Africa. Annals of the Entomological Society of America, 95(1), 66-74.
Onzo A., Biaou J.T. & Agboton C., 2018. Dead-wood collection and burning: an effective control measure against the wood borer Apate terebrans in cashew orchards in Northern-Benin. Journal of Applied Biosciences, 121, 12168-12180. https://dx.doi.org/10.4314/jab.v121i1.7
Ouali N’Goran S.-W.M., Akessé E.N., Ouattara G.M. & Koné D., 2020. Process of attack on cashew tree branches by Diastocera trifasciata (Coleoptera: Cerambycidae) and the relationship between these attacks and the phenological stages in the Gbêkê Region (Central Côte d’Ivoire). Insects, 11, 456. https://doi.org/10.3390/insects11080456
Ouattara G.M., 2017. Analysis of the dynamics of raw cashew nut supply in Côte d'Ivoire: an application of the ARDL (Self-regressive with Staggered Delay) approach. European Scientific Journal, 14(34), 292-306. https://doi.org/10.19044/esj.2018.v14n34p292
Reisenman C.E., Lei H. & Guerenstein P.G., 2016. Neuroethology of Olfactory-Guided Behaviour and Its Potential Application in the Control of Harmful Insects. Frontiers in Physiology, 7, 271, 21 p. https://doi.org/10.3389/fphys.201600271
RStudio Team, 2019. Integrated Development for R. RStudio, Inc., Boston, MA URL http://www.rstudio.com
Ruf F., Koné S. & Bebo B., 2019. Côte d’Ivoire’s cashew nut boom: A social and ecological transition of the cotton and cocoa systems. Cahiers Agricultures, 8(21), 1-12. https://doi.org/10.1051/cagri/2019019
Rumschlag S.L., Mahon M.B., Hoverman J.T., Raffel T.R., Carrick H.J., Hudson P.J. & Rohr J.R., 2020. Consistent effects of pesticides on community structure and ecosystem function in freshwater systems. Nature Communications, 11, 6333. https://doi.org/10.1038/s41467-020-20192-2
Schröder M.L., Glinwood R., Webster B., Ignell R. & Krüger K., 2015. Olfactory responses of Rhopalosiphum padi to three maize, potato, and wheat cultivars and the selection of prospective crop border plants. Entomologia Experimentalis et Applicata, 157(2), 241-253. https://doi.org/10.1111/eea.12359
Soro S., Sanogo S., Ouattara G.M., Silue N., Koné D. & Kouadio Y.J., 2020a. Descriptive analysis and vanguards agronomic factors of the sanitary status of the cashew orchard in Côte d'Ivoire (Anacardium occidentale L.) en Côte d’Ivoire. European Scientific Journal, 16(30), 72-86. https://doi.org/10.19044/esj.2020.v16n30p72
Soro S., Tuo S., Ouattara G.M., Traore M.M., Koné D. & Kouadio Y.J., 2020b. Inventory of pesticides use in cashew nuts orchards in the north of Côte d'Ivoire. International Journal of Development Research 10(11), 41925-41929.
Steel R., Elmouttie D. & Hamilton G., 2012. Geostatistical analysis of adult Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae) in wheat stored at constant temperatures. Journal of Stored Products Research, 50, 8-14.
Snyder L.D., Gomez M.I., Mudrak E.L. & Power A.G., 2021. Landscape-dependent effects of varietal mixtures on insect pest control and implications for farmer profits. Ecology, 31(2), e02246. https://doi.org/10.1002/eap.2246
Thackeray S.J., Henrys P.A., Hemming D., Bell J.R., Botham M.S., Burthe S., Wanless S. 2016. Phenological sensitivity to climate across taxa and trophic levels. Nature, 535, 241–245. https://doi.org/10.1038/nature18608
Thomas S.M., Simmons G.S. & Daugherty M.P., 2017. Spatio temporal distribution of an invasive insect in an urban landscape: introduction, establishment and impact. Landscape Ecology, 32, 2041–2057. https://doi.org/10.1007/s10980-017-0565-0
Vasconcelos S., Mendes L.F., Beja P., Hodgson C.J. & Catarino L., 2014. New Records of Insect Pest Species Associated with Cashew, Anacardium occidentale L. (Anacardiaceae), in Guinea-Bissau. African Entomology, 22(3), 673-677.
Wang C.-J., Wang S.-J., Yu C.-M., Wang X.-T., Wang R. & Wan J.-Z., 2023. Habitat heterogeneity and topographic variation as the drivers of insect pest distributions in alpine landscapes. Acta Ecologica Sinica, 43(4), 596-603. https://doi.org/10.1016/j.chnaes.2022.08.005
Yang G.T., Wang X.C., Nabi F., Wang H.N., Zhao C.K., Peng Y.L., Ma J. & Hu Y.G., 2021. Optimizing planting density and impact of panicle types on grain yield and microclimatic response index of hybrid rice (Oryza sativa L.). International Journal of Plant Production, 15, 447–457. https://doi.org/10.1007/s42106-021-00150-8
Yéo K., Aliko Y.J. & Ouali-N’Goran S.-W.M., 2019. Dégâts et facteurs d’influence des infestations de Apate terebrans (Coleoptera, Bostrichidae, Pallas, 1772), ravageur de l’anacardier en Côte d’Ivoire : cas des localités de Bondoukou et de Bouna. Reb-Pasres, 3(3), 11-21.
(57 Réf.)
