


- Accueil
- Volume 44 (2026)
- Efficacité de champignons entomopathogènes contre la bruche du niébé Callosobruchus maculatus (Coleoptera : Chrysomelidae)


Visualisation(s): 0 (0 ULiège)
Téléchargement(s): 0 (0 ULiège)


Efficacité de champignons entomopathogènes contre la bruche du niébé Callosobruchus maculatus (Coleoptera : Chrysomelidae)

Résumé
Les infestations causées aux graines du niébé en stockage par Callosobruchus maculatus compromettent leur utilisation alimentaire et semencière. Au regard des effets délétères et de l’obsolescence de la lutte chimique couramment utilisée contre le ravageur, des alternatives durables de lutte s'imposent. A cet égard, nous avons évalué, en conditions de laboratoire, les effets létaux et sublétaux de trois souches fongiques, Beauveria bassiana (GHA), Metarhizium brunneum (USDA 4556 et V275) sur Callosobruchus maculatus, la bruche tropicale du niébé, par pulvérisation directe d’adultes aux suspensions fongiques de chacune de souches à 10 8 conidies/ml et par leur exposition aux semences du niébé contaminées aux solutions sporales. Beauveria bassiana (GHA) s’est montrée plus virulente avec des TL50 plus courts de cinq jours et 100 % de mortalités d’adultes en sept et neuf jours. En revanche, des TL50 de six jours et des taux de mortalités de 86,6% à 98,3% ont été obtenus après dix jours avec les souches M. brunneum USDA 4556 et V275, respectivement. Les taux d’inhibition de pontes sur les graines enregistrés variaient de 64,6% pour M. brunneum (V275) à 68,3% pour B. bassiana (GHA). Les traitements fongiques appliqués aux semences ont fourni une protection partielle en réduisant les taux d’infestations de graines par C. maculatus de 20% à 30%. Ces résultats suggèrent que les trois souches fongiques pourraient constituer des agents entomopathogènes efficaces contre C. maculatus et des alternatives potentielles aux pesticides chimiques couramment utilisés dans les greniers semenciers du niébé en milieu tropical.
Abstract
Infestations caused by Callosobruchus maculatus in stored cowpea seeds compromise their use as food and seed. Given the harmful effects and obsolescence of the chemical control method commonly used against this pest, sustainable alternatives are needed. In this regard, we evaluated, under laboratory conditions, the lethal and sublethal effects of three fungal strains, Beauveria bassiana (GHA) and Metarhizium brunneum (USDA 4556 and V275) on Callosobruchus maculatus, the tropical cowpea weevil, by directly spraying adults with fungal suspensions of each strain at 10⁸ conidia/ml and by exposing them to cowpea seeds contaminated with spore solutions. Beauveria bassiana (GHA) proved to be more virulent with shorter LT50s of five days and 100% adult mortality in seven and nine days. In contrast, TL50s of six days and mortality rates of 86.6% to 98.3% were obtained after ten days with the strains M. brunneum USDA 4556 and V275, respectively. The egg-laying inhibition rates on seeds ranged from 64.6% for M. brunneum (V275) to 68.3% for B. bassiana (GHA). Fungal treatments applied to cowpea seeds provided partial protection by reducing seed infestation rates by C. maculatus by 20% to 30%. These results suggest that the three fungal strains could be effective entomopathogenic agents against C. maculatus and potential alternatives to chemical pesticides commonly used in cowpea seed storage facilities in tropical environments.
Introduction
1Callosobruchus maculatus Fabricius 1775 (Coleoptera : Chrysomelidae) constitue l’un des ravageurs post-récolte les plus dommageables des graines du niébé (Vigna unguiculata L. Walp) en stockage (Huignard et al., 2011; Ileke et al., 2020; Ozdemir et al., 2020 ), une légumineuse tropicale essentielle à la sécurité alimentaire de plus de 200 millions de personnes en Afrique (Bello et al., 2019; Huignard et al.,2011) en Asie et en Amérique du sud comme source principale de protéines végétales fournies par ses graines (Boukar et al. 2019 ; Carneiro da Silva et al., 2019). Ce ravageur constitue le facteur limitant majeur de la production du niébé dans toutes les zones tropicales de cultures entrainant jusqu’à 100 % de perte de production en graines en l’espace de quelques mois d’entreposage (Kang et al.,2013 ; Seri-Kouassi et al., 2004). Les dommages causés sur graines stockées par C. maculatus impactent ainsi significativement les conditions de vie socio-économiques des ménages producteurs africains tout en affectant la situation nutritionnelle des ménages démunies ayant un accès très réduit aux protéines animales (Rabé et al., 2023).
2Les dégâts du ravageur débutent au champ à la phase de maturité des gousses par des pontes sur les gousses sèches et se poursuivent en stockage après pénétration et développement des larves au sein de graines pour émerger sous forme d’adultes dans les stocks (Huignard et al., 2011). Il en résulte une réduction de capacités germinatives des semences jusqu’à 80% (Mofunanya et Namgbe, 2016), la perte de poids de graines, l’altération des qualités visuelles, organoleptiques et nutritionnelles des graines (la richesse en protéines) et en définitive la perte de la valeur commerciale de la production en plus des pertes quantitatives (Haouel Hamdi et al., 2017 ; Ileke et al., 2020). Il a été démontré que des valeurs d’infestations de 1 à 2 % au champ sont susceptibles d’entrainer jusqu’à 100% de pertes de graines du niébé entreposées 3 à 5 mois seulement après récolte (Kang et al., 2013; Ozdemir et al., 2020). Le cycle de vie polyvoltin du ravageur, son aptitude à infester les graines du niébé au champ et au cours du stockage de graines et l’absence de diapause reproductive chez les adultes sont les principaux facteurs qui accentuent l’importance de ses dégâts et des moyens de lutte à mettre en place (Bouchikhi-Tani Zoheir et al., 2011; Huignard et al., 2011). La durée totale du cycle de vie de C. maculatus est d’environ 20 jours à 30°C, le développement larvaire nécessitant 189,8 degrés-jours au-dessus de 15,1°C et le développement nymphal 77,2 degrés-jours au-dessus de 12,4 °C (Kutcherov, 2020).
3La gestion de C. maculatus dans les greniers semenciers repose essentiellement sur le recours aux produits fumigants et insecticides synthétiques tels que le gaz de phosphine, le phosphure d’aluminium, la perméthrine et le pirimiphos méthyle(Bouchikhi-Tani Zoheir et al., 2011; Dugje et al., 2009; Zongo et al., 2015;). L’usage de ces derniers suscite de nos jours plusieurs controverses en raison des problèmes de résistance développée par C. maculatus face aux nombreux pyréthrinoïdes et organophosphorés couramment utilisés contre ce ravageur en milieu tropical (Zongo et al, 2021), du coût d’application des pesticides, des préoccupations sur la santé des consommateurs et l’environnement (Cherry et al., 2007; Khoobdel et al., 2019; Ozdemir et al., 2020). Les contraintes d’ordre économiques auxquelles font face la plupart des ménages producteurs du niébé africains limitent également leur accès aux pesticides chimiques, aux équipements de protection et à la formation sur leur manipulation tout en augmentant les risques d’exposition aux matières actives (Zongo et al., 2015).
4En Afrique tropicale, la production du niébé est également confrontée au problème d’un accès limité des producteurs aux semences de qualité dû au coût élevé de semences améliorées au niveau local (Rabé et al.,2023). Pour faire face à cet obstacle, la majorité de producteurs stockent une partie de leur récolte de graines durant des périodes allant à sept mois dans des greniers traditionnels, des emballages hermétiques et magasin de stockage afin de servir de semences au cours de la prochaine campagne agricole (Dugje et al., 2009 ; Huignard et al., 2011). Le coût élevé des traitements chimiques pousse les producteurs démunis à recourir aux pratiques traditionnelles de stockage telles que les méthodes de stockage hermétiques (Rabé et al.,2023) ou l’incorporation de la cendre du bois, du sable, de l’argile ou des poudres végétales au stocks des graines (Viaud, 1983), le stockage en gousses, le séchage solaire ou le fumage de graines (Kpatinvoh et al.,2009) malheureusement adaptées aux productions à petite échelle et très limitées en termes d’efficacité contre les ravageurs post-récoltes tel que C. maculatus (Viaud, 1983).
5Eu égard aux nombreuses contraintes de la lutte chimique, de nombreuses études se sont dernièrement concentrées sur le développement des méthodes de lutte alternatives sures, durables et facile à mettre en place contre différents ravageurs de cultures. Parmi celles-ci, les champignons entomopathogènes se sont révélés des candidats potentiels en raison de leurs multiples succès observés sur une large diversité d’insectes ravageurs de pré- et de post-récoltes (Cherry et al., 2007; Lacey et al., 2015; Zimmermann, 2007). Cette étude évalue l’efficacité de trois souches fongiques précisément les effets létaux (TL50 et taux de mortalité) et sublétaux (inhibition de pontes sur graines) de Beauveria bassiana (GHA) formulé dans le produit commercial Botanigard 22WP et de deux souches de Metarhizium brunneum (USDA 4556 et V275) sur les adultes de C. maculatus en testant deux méthodes d’inoculation fongique à savoir la pulvérisation directe d’adultes avec des suspensions fongiques et leur exposition aux semences du niébé traitées avec des solutions sporales. L’effet protecteur des traitements fongiques appliqués aux semences est également évalué contre les infestations causées aux graines par C. maculatus.
Matériels et méthodes
6Les adultes de C. maculatus utilisés pour les bioessais proviennent des élevages réalisés selon Haouel Hamdi et al. (2017) en cages de 30cmx30cm et en boîtes plastiques de 10cmx10cm dans une chambre conditionnée du laboratoire d’entomologie fonctionnelle et évolutive de Gembloux Agro Bio-Tech maintenue à 27 ± 2°C, 60 ± 10% d’humidité relative et dans l’obscurité totale (Segers et al., 2023). La méthode d’élevage consiste à exposer 250 g de graines saines de niébé aux insectes adultes dans des cages ou des boîtes d’élevage pendant 2 jours pour recueillir les pontes. Les graines sont ensuite retirées et tamisées pour séparer les insectes adultes puis transférées dans de nouvelles boîtes d’élevage afin d'obtenir des adultes d'âge homogène après émergence. Une génération complète est obtenue en environ 21 jours.
7Trois souches fongiques ont été utilisées dans les essais biologiques notamment la souche commerciale Beauveria bassiana (GHA), agent actif du produit Botanigard 22WP (Certis, Europe, Belgique) et deux souches de Metarhizium brunneum (USDA 4556 et V275). La souche B. bassiana (GHA) a été produite à partir d’un stock de culture conservé dans une solution de glycérol à 10 % à −80°C, au laboratoire d’entomologie fonctionnelle et évolutive de Gembloux Agro-Bio Tech. Les souches de M. brunneum ont été obtenues auprès du département des biosciences de l’Université de Swansea (Royaume-Uni). Les trois souches ont été isolées selon Beris et al. (2024) sur milieu SDA (Sabouraud Dextrose Agar) préparé en dissolvant un mélange de 12g du Sabouraud Dextrose Broth et 6 g d’agar bactériologique dans 400 ml d’eau distillée, puis incubés pendant 15 jours à 25°C. Les conidies ont été prélevées des cultures dans du Tween 80 (0,05 % v / v) (Merck KGaA, Darmstadt, Allemagne) par grattage à l'aide d'une spatule stérile. Les suspensions obtenues ont été filtrées à l’étamine et la concentration en conidies a été ajustée à une dose unique de 1 × 10 8 conidies/ml à partir des dilutions de 100 fois dans du Tween 80 (0,05 % v / v) à l'aide d'un hémocytomètre Neubauer ® (Assistant, Sondheim vor der Rhön, Allemagne). En suite, 100 µl de suspension conidienne à 10 5 conidies de chaque souche ont été déposés sur milieu SDA et incubé à 23°C pendant 24 h pour évaluer la viabilité des conidies (Dessauvages et al., 2024). Seules les suspensions de conidies avec un taux de germination supérieur à 90 % ont été utilisées dans les bioessais (Jaber, 2018).
8Deux tests biologiques ont été effectués sur 600 adultes sexés de C. maculatus âgés de 3 jours en conditions de laboratoire. Le premier bioessai consistait à évaluer les effets létaux (TL50 et taux de mortalité) et sublétaux (inhibition de pontes sur graines) de trois souches fongiques en étude sur les adultes de C. maculatus pulvérisés directement avec les solutions sporales. Le second test visait à évaluer les mêmes effets létaux sur les adultes exposés aux semences du niébé traitées aux suspensions de trois souches fongiques et la ponte sur semences traitées au cinquième jour d’exposition. Chaque bioessai était constitué de cinq traitements en trois répétitions appliqués sur des lots de 20 adultes sexés de C. maculatus (10 mâles et 10 femelles) dont la suspension fongique concentré à 10 8 UFC/ml de B. bassiana (GHA); M. brunneum (V275 et USDA 4556), un contrôle négatif constitué d’une solution à 0,05 % (v / v) de Tween 80 (Asan et al., 2017) et un contrôle positif constitué d’une solution de 400µl/l d'insecticide karate ® zeon (Syngenta,Bâle,Suisse) contenant 100 g/L de λ-cyhalothrine (soit une solution à 40 mg/L de lamba-cyhalothrine).
9Des lots de 20 adultes sexés de C. maculatus anesthésiés au CO2 (soit un total de 300 adultes sexés) ont été disposés sur papiers filtres dans des boites de pétri de 90 mm. Les adultes ont été pulvérisés manuellement sous un cône tronqué (Figure 1 : 2) selon Segers et al. (2021) par un millilitre de chaque traitement. Les adultes traités ont été ensuite transférés en boîtes plastiques aérées de 10cm×10cm contenant 10 grammes de graines sèches saines du niébé disposées sur boîtes de pétri de 90mm pour piéger les pontes sur les graines. Les graines étaient renouvelées tous les trois jours durant 12 jours puis conservés à -20°C pour comptabiliser les pontes avec trois répétitions de 10 graines sélectionnées aléatoirement dans chaque lot (pontes totales de femelles mortes et vivantes). Les graines utilisées étaient nettoyées et désinfectées au départ à l’éthanol 70 %. Les essais étaient maintenus dans des conditions contrôlées à 25±2°C, 60±10% d’humidité relative et dans l’obscurité totale (conditions similaires à celles de stockage) pour le suivi de mortalités d’adultes traités toutes les 24 heures pendant 10 jours (Bouchikhi-Tani Zoheir et al., 2011).
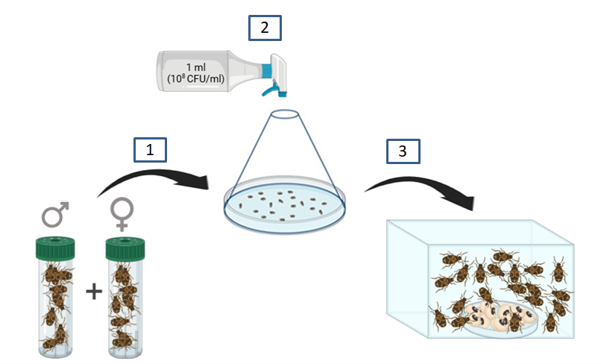
Figure 1 : Pulvérisation manuelle d’adultes sexés (1) de Callosobruchus maculatus anesthésiés au CO2 par différents traitements sous un cône tronqué (2) et disposition d’adultes traités en boîtes contenant les graines sèches du niébé pour le suivi des mortalités journalières et des pontes sur graines (3). © R. Lugendo
10Des lots de 10 grammes de graines sèches du niébé de la variété dite « aux yeux noirs » ont été nettoyés à l’eau distillée stérile et désinfectés en surface à l’éthanol 70%. Chaque lot de graines a été traité selon Jaber & Enkerli (2016) et Jaber (2018) par immersion dans 10 ml de solution sporale de chacune des souches et de traitements en étude dans des tubes falcons stériles de 50 ml pendant 10 minutes. Le mélange a été agité au vortex pendant 1 minute et les graines ont été séchées sous hotte pendant 30 minutes sur boîtes de Pétri de 90 mm, avant d’être transférées dans des boites en plastiques de 10cm×10cm (Cherry et al., 2005). Des lots de 20 adultes sexés de C. maculatus (soit un total de 300 adultes) ont été récupérés et transférés dans chacune de boîtes contenant des graines du niébé contaminées. Les adultes ont ensuite été maintenus en conditions contrôlées à 25±2°C ,60±10% d’humidité relative et dans l’obscurité totale pour le suivi de mortalités journalières pendant 10 jours. La ponte sur les semences traitées a été évaluée au cinquième jour d’exposition après la séparation de graines avec les insectes. Les graines traitées ont été retirées des boîtes d’essai après cinq jours puis conservées en boîtes de pétri de 90mm à 30°C, 60±10% d’humidité relative dans la chambre d’élevage de C. maculatus au laboratoire durant 25 jours après lesquels le taux d’infestation de chaque lot a été déterminé suivant le protocole de Ileke et al., (2020). Les adultes morts ont été retirés des boîtes d’essai, désinfectés en surfaces par immersion dans l’éthanol 70% pendant 2 minutes puis rincés trois fois à l’eau distillée stérile. Ces adultes ont été placés sur papiers filtres humides et maintenues à l’incubateur à 25°C pour confirmer l’infection fongique à partir de l’émergence du mycélium des souches testées sur la cuticule des insectes.
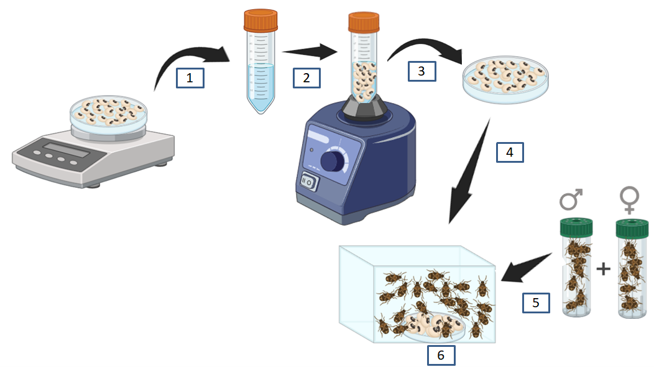
Figure 2 : Traitement des semences du niébé par immersion dans la solution sporale des souches fongiques et selon différents traitements en tubes falcons de 50 ml (1,2), agitation du mélange au vortex (3), suivi du transfert des semences traitées (4) et celui d’adultes sexés de Callosobruchus maculatus (5) en boîtes d’essai (6) pour le suivi des mortalités journalières pendant 10 jours. © R. Lugendo
11Le traitement statistique des données de mortalités, les statistiques comparatives des différents traitements et la conception des graphiques ont été réalisées grâce au logiciel R studio software version 1.3.9.5.9 et R version 4.2.0. L’estimation des temps létaux médians (TL50) des souches fongiques a été réalisée à l’aide de la méthode de Kaplan-Meier à partir d'observations du temps jusqu'à l'événement (décès), une méthode non paramétrique, qui attribue une probabilité de survie sur la base du temps de survie observé (Kaplan et Meier, 1958). La comparaison des courbes de survie de différents traitements a été réalisée à l’aide du test log-rank, un test non paramétrique distribué comme un test du chi carré en utilisant les packages ¨survival¨, ¨survminer¨ et dplyr du logiciel R. Pour évaluer l’effet des différents traitements sur le nombre d’œufs de C. maculatus déposés sur les graines, un modèle linéaire généralisé (GLM) ajusté avec une distribution de poisson adaptée aux données de comptage a été utilisé à l’aide de la fonction glm. Aussi, l’effet des traitements sur le taux d’infestation de graines par C. maculatus après 25 jours de conservation en laboratoire a été évalué à l’aide d’un modèle mixte généralisé (GLMM) ajusté avec une distribution binomiale adaptée aux données de proportions en intégrant un effet aléatoire de la répétition à l’aide de la fonction glmmTMB() du package glmmTMB. Des comparaisons multiples des différents traitements avec le traitement contrôle ont été réalisées à l’aide du test post hoc de Dunnett en considérant un seuil de signification de 5% et intervalles de confiance de 95%.
Résultats
12Une différence très significative entre les différents traitements a été observée (χ 2 (0,95 ; 4) = 199, p < 0,001). Parmi les trois souches fongiques testées, B. bassiana (GHA) a été la plus virulente sur les adultes traités de C. maculatus avec un TL50 plus court correspondant à cinq jours (χ 2 (0,95 ; 1) = 103, p < 0,001) et un taux de mortalité de 100% après sept jours de traitement. Des TL50 correspondant à six jours (χ 2 (0,95 ;1) = 92,3 ; p < 0,001) et (χ 2 (0,95;1) = 65, p < 0,001) ont été respectivement obtenus avec les souches M. brunneum USDA 4556 et V275 ainsi que des taux de mortalités d’adultes équivalent à 98,3% et 88,3% après dix jours de traitement. En revanche, un TL50 de huit jours et un taux de mortalité de 75% ont été enregistrés chez les adultes traités à l’insecticide chimique lambda cyhalothrine (Figure 3 et Tableau 1).
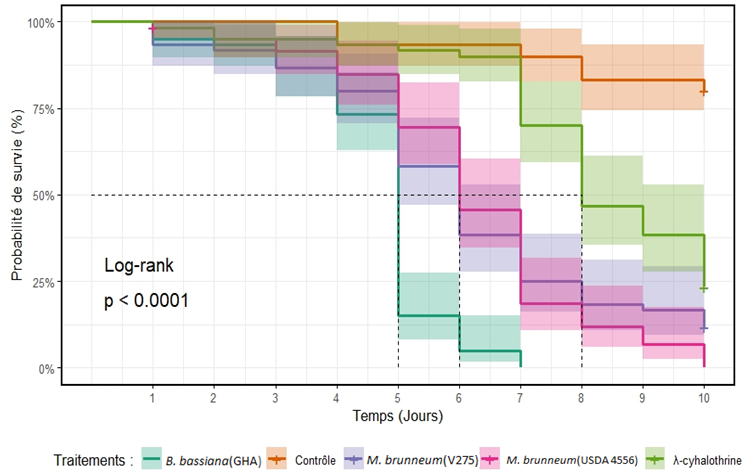
Figure 3 : Courbes de survie de Callosobruchus maculatus pulvérisés directement avec les suspensions fongiques. Les lignes pointillées indiquent les estimations de TL50. Chaque courbe de survie est présentée avec les intervalles de confiance à 95 %.
13Un effet hautement significatif des traitements a été observé (χ² (0,95 ; 4) = 208,05 ; p˂0,001) sur le nombre d’œufs observés sur les graines et l’oviposition au cours du temps d’observation post-traitement (χ² (0,95 ; 12) = 121,90 ; p < 0,001). Toutes les souches fongiques testées ont entrainé une réduction significative de la fécondité de C. maculatus sur les graines (Figure 4) au cours de différents moments d’observation post-traitement (effets estimés négatifs allant de -0,405 à -5,77; Z= –3,686 à –3,156 ; p<0,001) comparativement au traitement contrôle. Une inhibition complète de la ponte d’ insectes traités avec la souche B. bassiana (GHA) a été observée au neuvième jour de traitement (effets estimés négatifs = -5,782 ; Z= –3,162 ; p<0,001). Par ailleurs, les pontes d’adultes traités avec les souches M. brunneum (USDA 4556 et V275) ont été complètement inhibées douze jours après traitement (effets estimés négatifs =-5,772 et -5,771 ; Z= –3,156 et –3,156 ; p<0,001). Les pontes totales observées sur les graines durant la période d’observation ont montré, par rapport aux graines traitées avec de l’eau distillée stérile additionnée de Tween 80 (0,05 %), une inhibition de ponte de 68,3 % sur les graines provenant des boîtes contenant des adultes traités avec B. bassiana (GHA), contre 65,0 % et 64,6 % respectivement pour les souches de M. brunneum V275 et USDA 4556 (Tableau 1).
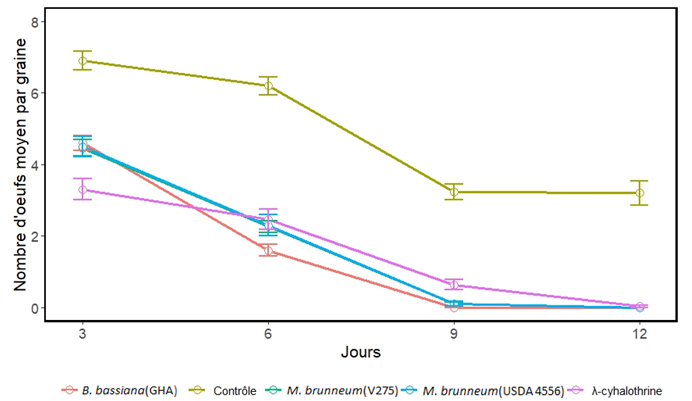
Figure 4 : Evolution des pontes sur graines de Callosobruchus maculatus soumis aux différents traitements pendant la période d’observation
14Une différence hautement significative entre les différents traitements (χ²(0,95 ; 4) = 111, p < 0,001) a été observée. Beauveria bassiana (GHA) a été la souche la plus efficace en termes de létalité d’adultes exposés aux semences traitées avec un TL50 plus court correspondant à cinq jours et un taux de mortalité d’adultes de 100% au neuvième jour d’exposition. Des TL50 équivalent à six jours et des taux de mortalités de 91,6% et 86,6% ont été obtenus avec les souches USDA 4556 et V275 de M. brunneum après dix jours d’exposition d’adultes aux semences traitées. En outre, un TL50 de sept jours et un taux de mortalité de 81,7% ont été enregistrés auprès d’adultes exposés sur graines traitées à l’insecticide lambda cyhalothrine (Figure 5 et Tableau 1.)
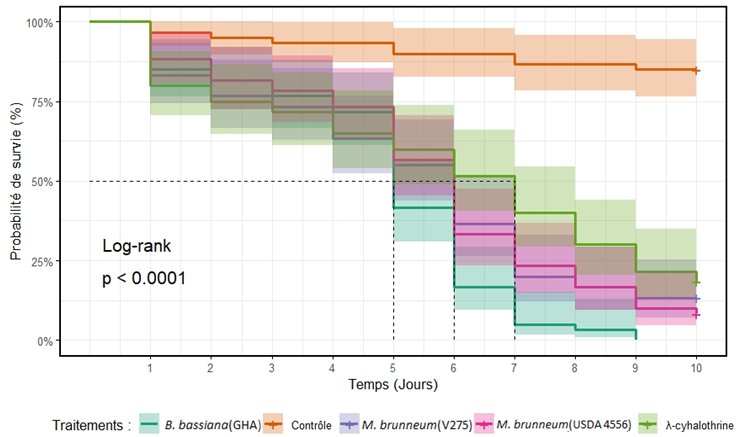
Figure 5 : Courbes de survie de Callosobruchus maculatus exposés aux semences traitées avec les suspensions fongiques. Les lignes pointillées indiquent les estimations de TL50. Chaque courbe de survie est présentée avec les intervalles de confiance à 95 %.
15L’effet des différents traitements a été également évalué sur l’oviposition de C. maculatus sur semences traitées au cinquième jour d’exposition. Un effet significatif du facteur traitement (χ²(0,95 ; 4) =10,80 ; p =0,028) sur le nombre d’œufs comptabilisés sur les semences traitées a été observé. Le traitement des semences avec B. bassiana (GHA) (effet estimé négatif = –0,209; Z= –2,654 ; p=0,029) et M. brunneum (USDA 4556) (effet estimé négatif = –0,191 ; Z= –2,450 ; p=0.050) a entraîné une réduction statistiquement significative des pontes sur graines par rapport au traitement contrôle. Par ailleurs, les traitements M. brunneum (V275) (effet estimé négatif = –0,068 ; Z= –0,908 ; p=0,739) et λ-cyhalothrine (effet estimé négatif = –0,17 ; Z= –2,288 ; p=0.076) n’ont pas eu d’effet significatif sur l’oviposition sur graines comparés au traitement contrôle. Les valeurs moyennes de pontes comptabilisées par graine ont été de 9,77 ± 2,56; 9,93 ± 2,41; 11,2 ± 2,18 œufs sur semences traitées respectivement avec B. bassiana (GHA); M. brunneum (USDA 4556 et V275) et de 10,1 ± 2,8 et 12,0 ± 3,6 œufs sur celles traitées avec l’insecticide lambda cyhalothrine et à l’eau distillée stérile + Tween 80 (0,05%) (Figure 5 et Tableau 1).
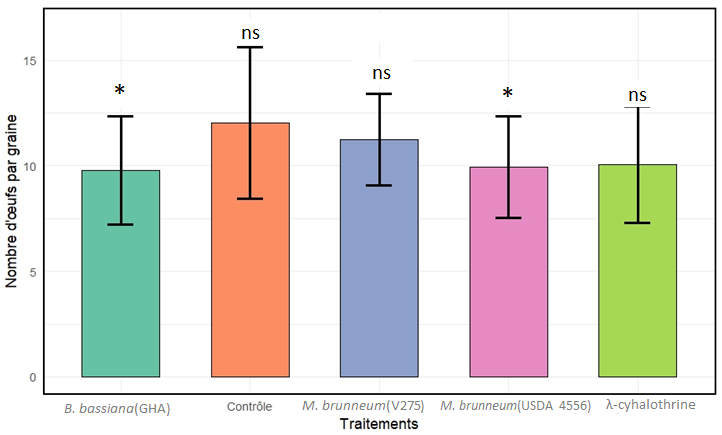
Figure 6 : Fécondité de Callosobruchus maculatus sur semences traitées au cinquième jour d’exposition (moyenne ± écart-type du nombre d’œufs ; *=significatif ; ns = non significatif)
16Une différence statistiquement significative a été observée entre les taux d’infestations de semences traitées avec les suspensions de B. bassiana (GHA) (effet estimé négatif = -2,09 ; Z = -2.54 ; p = 0,029) et de M. brunneum (USDA 4556) (effet estimé négatif = -1,95 ; Z = -2.35; p = 0,049) par rapport à celui observé sur graines traitées à l’eau distillée stérile + Tween 80 (0,05%) (traitement contrôle). En revanche, aucune différence significative n’a été observée entre le traitement contrôle et les traitements avec M. brunneum (V275) (effet estimé négatif = -1,62; Z = -1,93; p = 0,12) et la λ-cyhalothrine (effet estimé négatif = -1,79; Z = -2,15; p = 0,07) en termes de taux d’infestation observés sur semences traitées. Le trempage des semences avec les suspensions de B. bassiana (GHA) et M. brunneum (USDA 4556) a réduit les taux d’infestation par C. maculatus de 30% et 27% comparativement au traitement contrôle. Des taux de réduction moyens d’infestation équivalent à 20 % et 23% ont été respectivement obtenus avec les traitements M. brunneum (V275) et l’insecticide lambda cyhalothrine. Les proportions moyennes de graines bruchées équivalent à 63,3 ± 15,2% ; 66,6 ± 20,8% et 73,3 ± 15,2% ont été respectivement obtenues sur semences traitées avec B. bassiana (GHA) ; M. brunneum USDA 4556 et V275 contre 70,0 ± 17,3% et 93,3 ± 5,7% de graines infestées obtenues sur semences traitées au lambda cyhalothrine et à l’eau distillée stérile + Tween 80 (0.05%). (Figure 7 et Tableau 2).
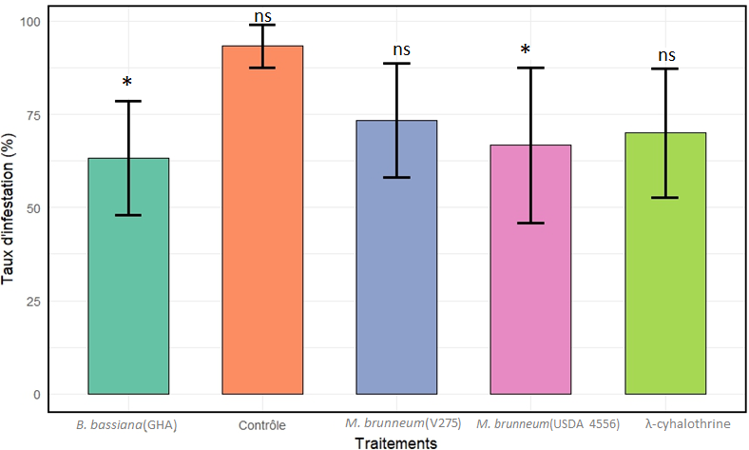
Figure 7’ : Taux d’infestation des graines par Callosobruchus mmaculatus selon les différents traitements appliqués après 25 jours de conservation (moyennes d’individus ± écart-types ; *=significatif ; ns = non significatif)
Tableau 1 : Synthèse des résultats d’effets létaux et sublétaux des souches fongiques sur Callosobruchus maculatus (TL50= temps létal 50, TM= taux de mortalité, IP= inhibition de pontes, PD= pulvérisation directe sur les adultes, IS= immersion des semences, n = nombre total d'adultes de C. maculatus, Chisq = statistique du chi-carré du test log-rank ; DL = degré de liberté lié au test log-rank ; p = valeur p associée au test log-rank).
|
Estimations de survie de Kaplan-Meier |
Test Log-Rank Vs contrôle |
Pontes totales |
Taux d’inhibition de pontes |
|||||||||||||||||||||||||||||||
|
Bioessai |
Observations |
Méthode de pulvérisation |
Traitements |
n |
Mortalités observées |
Médiane (TL50) |
Chisq |
DL |
p |
|||||||||||||||||||||||||
|
TL50 +TM +IP |
PD |
B. bassiana (GHA) |
60 |
60 |
5 |
103 |
1 |
<0,001 |
186 œufs |
0,68 |
||||||||||||||||||||||||
|
1 |
TL50 +TM+IP |
PD |
λ-cyhalothrine (40 mg/L) |
60 |
46 |
8 |
36,1 |
1 |
<0,001 |
193 œufs |
0,67 |
|||||||||||||||||||||||
|
TL50 +TM+IP |
PD |
M. brunneum (USDA 4556) |
60 |
59 |
6 |
92,3 |
1 |
<0,001 |
207 œufs |
0,65 |
||||||||||||||||||||||||
|
TL50 +TM+IP |
PD |
M. brunneum (V275) |
60 |
53 |
6 |
65 |
1 |
<0,001 |
205 œufs |
0,65 |
||||||||||||||||||||||||
|
TM |
PD |
Tween 80 (0,05%) |
60 |
12 |
>10 |
/ |
/ |
/ |
586 œufs |
- |
||||||||||||||||||||||||
|
TL50+ TM |
IS |
B. bassiana (GHA) |
60 |
60 |
5 |
95,4 |
1 |
<0,001 |
- |
- |
||||||||||||||||||||||||
|
TL50+ TM |
IS |
λ-cyhalothrine (40 mg/L) |
60 |
49 |
7 |
53,7 |
1 |
<0,001 |
- |
- |
||||||||||||||||||||||||
|
2 |
TL50+ TM |
IS |
M. brunneum (USDA 4556) |
60 |
55 |
6 |
72,7 |
1 |
<0,001 |
- |
- |
|||||||||||||||||||||||
|
TL50+ TM |
IS |
M. brunneum (V275) |
60 |
52 |
6 |
63,3 |
1 |
<0,001 |
- |
- |
||||||||||||||||||||||||
|
TM |
IS |
Tween 80 (0,05%) |
60 |
9 |
>10 |
/ |
/ |
/ |
- |
- |
||||||||||||||||||||||||
Discussion
17La présente étude a démontré la virulence de la souche commerciale B. bassiana (GHA) et de deux souches de M. brunneum (USDA 4556 et V275) sur C. maculatus traités directement avec des suspensions fongiques et exposés aux semences de niébé traitées. Beauveria bassiana (GHA) a été la souche la plus virulente dans les deux bioessais avec des TL50 plus courts correspondant à cinq jours et des taux de mortalités d’adultes les plus élevés équivalent à 100%. Ces résultats suggèrent que B. bassiana (GHA) et les deux souches de M. brunneum (USDA 4556 et V275) peuvent constituer des agents entomopathogènes efficaces contre C. maculatus. L’efficacité de trois souches démontrée dans la présente étude confirme leur virulence déjà prouvée sur d’autres espèces d’insectes ravageurs telles que Bruchus rufimanus Boheman 1833, la bruche tempérée de féveroles (Vicia faba L.) (Lugendo et al., 2025) et Curculio elephas (Gyllenhal,1836) le charançon du châtaignier (Castanea sativa Mill.) (Asan et al., 2017). La virulence de différentes souches fongiques appartenant aux genres Beauveria et Metarhizium a également été prouvée avec succès sur d’autres insectes ravageurs de stocks telles que Rhyzopertha dominica (Fabricius, 1792), le ravageur de grains de blé entreposés (Mahdneshin Z. et al. 2009), Prostephanus truncatus (Horn, 1878) (Bourassa et al., 2001 ; Meikle et al., 2001) et Sitophilus zeamais Motschulsky, 1885 (Hidalgo et al. 1998) deux ravageurs les plus dommageables de grains de maïs en stock et Oryzaephilus surinamensis (Linnaeus, 1758) le cucujide dentelé des grains de riz et de blé stockés (Khoobdel, Pourian et Alizadeh, 2019). Par ailleurs, la performance spécifique de B. bassiana (GHA) sur C. maculatus s’explique également par son large spectre d’hôtes incluant une diversité de Coléoptères ravageurs de graines des légumineuses et des céréales en stock (Batta, 2007 ; Cherry et al., 2007; Ozdemir et al., 2020; Pande & Mishra, 2018). Des résultats similaires aux nôtres soit 100 % de mortalités d’adultes de C. maculatus traités aux suspensions de la souche B. bassiana (0362) après six jours et des TL50 variant entre 3,11 à 6,13 jours ont été rapportés par l’étude de Cherry et al. (2005). Ozdemir et al. (2020) ont également obtenus 100% de mortalité d’adultes de C. maculatus traités aux solutions sporales de B. bassiana (TR 217) et M. anisopliae (TR 106) huit jours après traitement et des TL50 respectifs de 4,61 jours et 5,34 jours.
18L’infectiosité de B. bassiana et M. brunneum par simple contact de spores fongiques avec les insectes ravageurs a aussi été démontrée par plusieurs études (Asan et al., 2017; Jarrold et al, 2007). Il a été également prouvé que l’infection fongique peut se transmettre horizontalement des surfaces ou objets contaminés tels que les graines vers les insectes ravageurs non contaminés (Quesada-Moraga et al., 2008). Ces observations mettent en lumière la transmission de l’infection fongique de trois souches appliquées sur les semences du niébé vers les adultes sains de C. maculatus et leur virulence observée chez les adultes exposés sur semences contaminées. Nos résultats sont cohérents avec l’étude de Pande & Mishra (2018) qui a également rapporté la virulence d’une autre souche de B. bassiana (2028) chez les adultes de Callosobruchus spp exposés aux graines sèches du haricot vert (Vigna radiata L.) traitées aux suspensions fongiques avec un taux de mortalité de 99% d’adultes au dixième jour d’exposition. Cherry et al. (2005) ont également démontré la virulence des souches B. bassiana (0362) et de M. anisopliae (0351) chez les adultes de C. maculatus exposés aux graines du niébé mélangées avec les conidies sèches de deux souches avec des taux de mortalités d’adultes de l’ordre de 90%.
19L’efficacité des trois souches étudiées a été également démontrée avec succès sur la fécondité d’adultes de C. maculatus. Toutes les souches testées ont réduit significativement la fécondité d’adultes infectés sur les graines sèches du niébé au cours des différents moments d’observation des pontes. Un résultat similaire a été également observé avec les trois souches dans notre récente étude chez les adultes de Bruchus rufimanus, le ravageur le plus dommageable de graines de féveroles (Vicia faba) au cours d’un bioessai effectué en laboratoire (Lugendo et al., 2025). Ces différentes observations convergent avec celle Vinayaga Moorthi et al., (2015) ayant également démontré que l’infection fongique peut impacter considérablement et/ou inhiber la fécondité d’un insecte mycosé.
20En revanche, nos résultats ont démontré que le traitement chimique à base de l’insecticide lambda cyhalothrine a causé moins de mortalité d’adultes de C. maculatus dans nos deux bioessais par rapport aux traitements fongiques. La preuve d’une résistance de C. maculatus aux nombreux pyréthrinoïdes et organophosphorés couramment appliqués dans les greniers de stockage du niébé en zones tropicales a été fournie par l’étude de Zongo et al. (2021). De ceux-ci figure la lambda cyhalothrine commercialisée localement sous le nom lambda super (Zongo et al., 2015). L’implication chez C. maculatus des enzymes telles que les mono-oxygénases du cytochrome P450 dans la dégradation d’insecticides et la détoxication d’adultes traités, réduisant ainsi l’efficacité des traitements chimiques a été également démontrée par l’étude de Zongo et al. (2021). Ces raisons pourraient expliquer l’inefficacité du traitement chimique observé sur C. maculatus au cours de nos expériences. Elles soulèvent par ailleurs, une nécessité de réflexion sur des approches non chimiques de gestion durable du ravageur qui permettraient de réduire à un seuil acceptable les pertes quantitatives, qualitatives et économiques occasionnées par ses dégâts.
21Les résultats obtenus ont démontré des effets protecteurs intéressants des souches B. bassiana (GHA) et M. brunneum (USDA 4556) sur semences du niébé traitées avec des suspensions des deux souches avec des taux de réduction moyennes d’infestations de C. maculatus de 30% et 27% comparativement aux graines traitées à l’eau distillée stérile +Tween 80. Ces résultats suggèrent que B. bassiana et M. brunneum peuvent constituer des biopesticides intéressants susceptibles de réduire les dégâts causés par C. maculatus dans les stocks de graines et d’offrir une protection aux semences. En revanche, des réductions des taux d’infestation de graines de 20% et 23% par rapport au traitement contrôle ont été obtenus avec M. brunneum (V275) et la λ-cyhalothrine. Deux hypothèses pourraient justifier la protection partielle offerte aux semences par certains traitements utilisés notamment la faible persistance de la formulation fongique liquide appliquée sur les semences durant la conservation et les conditions de températures élevées maintenues de manière constante dans la chambre d’élevage de C. maculatus où étaient incubées les graines après traitement.
22Il a été démontré que le niveau de protection de semences par les champignons entomopathogènes varie en fonction du type de formulation fongique utilisée ainsi que d’une espèce ou d’une souche fongique (Meikle et al., 2001). Il a été également prouvé que les formulations fongiques sèches telles que les conidies sèches des champignons entomopathogènes offrent plus d’efficacité vis-à-vis des ravageurs des stocks par rapport aux formulations liquides en raison de leur persistance élevée dans les stocks et la facilité d’auto-dissémination ou de propagation des spores fongiques sèches entre insectes congénères (Cherry et al., 2005; Meadow et al., 2000). Des études antérieures ont également démontré que les facteurs environnementaux hostiles tels que les températures élevées et basses, les rayonnements UV etc. réduisent significativement l’efficacité des champignons entomopathogènes (Muniz et al., 2020; Quesada-Moraga et al., 2023) et peuvent affecter négativement l’efficacité pesticides chimiques tels que la lambda cyhalothrine (Mudassir et al.,2015). Le choix d’une formulation fongique adéquate et la prise en compte des conditions environnementales de stockage de semences tels que la température de stockage seraient des facteurs clés à prendre en considération dans une stratégie d’intégration des biopesticides fongiques dans la lutte contre les ravageurs de stocks tels que C. maculatus.
23La conservation de semences du niébé après récolte constitue un problème majeur auquel se heurte la plupart de producteurs en milieu tropical. Différentes méthodes de stockage de semences du niébé sont utilisées par ces derniers pour faire face aux infestations de C. maculatus. Une majorité d’agriculteurs africains stockent leurs graines après récoltes pendant des périodes allant de 3 à 7 mois dans des greniers traditionnelles conçus en matières végétales ou en argile (banco), des réservoirs aménagés localement, des récipients hermétiques (fûts en plastics), des sacs en polyéthylènes à triples couches ou des magasins de stockage (chez quelques acteurs impliqués dans le commerce de graines) en y incorporant divers produits de protection de graines (Dugje et al., 2009). Les semences sont souvent mélangées avec une matière inerte (couche de sable, d’argile ou de cendre de 20 cm d'épaisseur alternées avec les graines), aux poudres chimiques telles que l’actellic super dust (pirimiphos-méthyle)(Guèye et al.,2011; Swella & Mushobozy,2007),le spintor (Spinosad),le Dursban (chlorpyriphos-éthyl)(Guèye et al., 2011), ou traitées aux fumigants et insecticides de synthèse tels que la phosphine, le phosphure d’aluminium, la perméthrine ou le pirimiphos- méthyle (concentré émulsifiable) avant et pendant le stockage (Kpatinvoh et al., 2009 ; Zongo et al., 2015 ). . Cependant, la majorité de ces pesticides ne sont pas agréés au niveau local en raison de leur forte toxicité et leur manipulation pose des sérieux problèmes aux agriculteurs (Zongo et al., 2015). De plus, bon nombre de ces substances sont devenues inefficaces en raison de la résistance développée par C. maculatus (Idrissi et al., 2010 ; Kanda et al., 2009 ; Williamson et al., 2008 ; Zongo et al., 2015). Les pratiques traditionnelles de protection de semences utilisées dans les greniers offrent également une efficacité très limitée contre C. maculatus tout en restant adaptées aux exploitations à petite échelle (Viaud, 1983).
24Dans ce contexte, les stratégies de gestion de C. maculatus basée sur les biopesticides fongiques constituent des pistes potentielles et intéressantes pour la mise en place des méthodes de lutte efficaces, durables, respectueuses de l’environnement et facile en termes de mise en œuvre par les agriculteurs. Une première approche consisterait à sélectionner des souches fongiques efficaces adaptées aux conditions climatiques tropicales. Cette stratégie permettrait de contourner les effets des facteurs environnementaux hostiles tels que les températures élevées dans les installations de stockage et d’améliorer la persistance et l’infectiosité des agents fongiques dans les greniers semenciers (Quesada-Moraga et al., 2023). Différentes souches fongiques aux larges spectres d’hôtes ont été identifiées et certaines d’entre elles sont de nos jours commercialiséesà l’occurrence de Beauveria bassiana (GHA) Botanigard 22WP (Certis, Europe, Belgique)(Lugendo et al., 2025) ; Beauveria bassiana ATCC 74040 Naturalis® (CBC ,Europe,Srl)(Atzeni et al., 2020), Metarhizium brunneum GranMet® (Agrifutur srl, Alfianello (BS), Italie), Metarhizium brunneum Met52® (Novozymes BioAg Ltd, Saskatoon, Canada) (Benvenuti et al., 2019), etc. Une seconde stratégie consisterait à la mise en place des formulations fongiques sèches facilement applicables par les producteurs dans les dispositifs de stockage telles que des poudres pulvérisables, des poudres sèches associées aux matières inertes tels que le talc, le kaolin, l’amidon applicables sur les graines par enrobage à sec, les granulés secs, la terre de diatomée etc. La coformulation de la poudre sèche de kaolin avec une souche de B. bassiana (isolat IMI389521) a permis d’améliorer significativement le contrôle d’adultes d' Oryzaephilus surinamensis et de Tribolium confusum Jacquelin du Val 1861 sur du blé stocké, avec une mortalité atteignant jusqu'à 99% après 14 jours (Storm et al., 2016). L’utilisation des poudres pulvérisables de B. bassiana a déjà été testé avec succès sur Sitophilus zeamais par Hidalgo et al. (1998). Khoobdel et al. (2019) ont obtenu une mortalité élevée d’adultes de C. maculatus et d’Oryzaephilus surinamensis en appliquant aux graines du niébé et du blé une combinaison d’une souche de B. bassiana(IRAN441C) avec de la terre de diatomées commerciale (DE). Des études approfondies devraient donc être menées afin d’évaluer l’efficacité de ces différentes stratégies dans des conditions de stockage à grande échelle et étudier les risques potentiels associés. Les analyses effectuées à l'aide d'une CLHP (Thermo Scientific Ultimate 3000) par Pande & Mishra (2018) n’ont détecté aucune production de mycotoxine dans les graines de Vigna radiata traitées de manière optimale par quatre concentrations différentes de B. bassiana suggérant que les graines traitées avec B. bassiana devraient être destinées à des fins comestibles. Ces différentes études mettent en évidence le statut des biopesticides fongiques en tant qu’alternative prometteuse aux fumigants et insecticides chimiques largement utilisés contre les ravageurs des stocks.
Conclusions
25La présente étude a mis en évidence les effets létaux et sublétaux de trois souches fongiques à savoir : B. bassiana (GHA), M. brunneum (V275 et USDA4556) sur les adultes de la bruche tropicale du niébé C. maculatus pulvérisés directement aux suspensions fongiques et exposés aux semences du niébé contaminées. Beauveria bassiana (GHA) s’est montré plus virulente tant en termes de létalité et d’inhibition de pontes d’adultes traités et exposés aux semences traitées comparativement aux souches de M. brunneum V275 et USDA4556. Ces résultats suggèrent que ces souches peuvent constituer des agents de lutte biologique performants contre la bruche tropicale du niébé C. maculatus et une alternative de taille aux insecticides chimiques. Néanmoins, des études supplémentaires sont envisageables pour évaluer leur efficacité au champ et en condition de stockage à grande échelle.
Bibliographie
Asan, C., et al., 2017. An innovative strategy for control of the chestnut weevil Curculio elephas (Coleoptera : Curculionidae) using Metarhizium brunneum. Crop Protection,102,147‑153. https://doi.org/10.1016/j.cropro.2017.08.021
Atzeni, R. et al. 2020. « Genome Sequence of Beauveria bassiana Strain ATCC 74040, a Widely Employed Insect Pathogen», Microbiology Resource Announcements, 9(23). https://doi.org/10.1128/mra.00446-20.
Batta, Y. A., 2007. Biocontrol of almond bark beetle (Scolytus amygdali Geurin-Meneville, Coleoptera : Scolytidae) using Beauveria bassiana (Bals.) Vuill. (Deuteromycotina : Hyphomycetes). Journal of Applied Microbiology, 103(5), 1406‑1414. https://doi.org/10.1111/j.1365-2672.2007.03369.x
Bello, S., Babalakoun, A. O., Zoudjihékpon, J., & Coulibaly, K. A., 2019. Diversité de l’entomofaune du niébé (Vigna unguiculata (L.) Walpers) au Nord-Ouest du Bénin. Journal of Applied Biosciences, 132(1), 13424. https://doi.org/10.4314/jab.v132i1.4
Benvenuti, C. et al., 2019. « A new device for auto-disseminating entomopathogenic fungi against Popillia japonica: a study case. », Bulletin of Insectology, 72 (2): 219-225.
Bouchikhi-Tani Zoheir, Anouar, K. M., Mourad, B., & Pujade-villar, J., 2011. Lutte contre les trois bruches Acanthoscelides obtectus (Say, 1831), Bruchus rufimanus Boheman 1833 et Callosobruchus maculatus (Chrysomelidae : Bruchinae ) par les huiles essentielles extraites d’ Origanum glandulosum (Lamiacées). Regiment de la cosa natural, 177‑186. http://dx.doi.org/10.3406/ecmed.2013.1282
Boukar, O., et al., 2019. Cowpea [Vigna unguiculata (L.) Walp.] Breeding’, in J.M. Al-Khayri, S.M. Jain, and D.V. Johnson (eds) Advances in Plant Breeding Strategies: Legumes: Volume 7. Cham: Springer International Publishing, pp. 201–243. https://doi.org/10.1007/978-3-030-23400-3_6
Bourassa, C., et al., 2001. Effects of Entomopathogenic Hyphomycetes against the Larger Grain Borer, Prostephanus truncatus (Horn) (Coleoptera : Bostrichidae), and Its Predator, Teretriosoma nigrescens Lewis (Coleoptera: Histeridae). Journal of Invertebrate Pathology, 77(1), 75‑77. https://doi.org/10.1006/jipa.2000.4986
Carneiro da Silva, A., et al., 2019. Cowpea : A Strategic Legume Species for Food Security and Health. In : Jimenez-Lopez J.C. & Clemente A., eds. Legume Seed Nutraceutical Research. IntechOpen. ISBN 978-1-78985-398-8, 96 p. https://doi.org/10.5772/intechopen.79006
Cherry, A. J., Abalo, P., & Hell, K., 2005. A laboratory assessment of the potential of different strains of the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) to control Callosobruchus maculatus (F.) (Coleoptera : Bruchidae) in stored cowpea. Journal of Stored Products Research, 41(3), 295‑309. https://doi.org/10.1016/j.jspr.2004.04.002
Cherry, A. J., Abalo, P., Hell, K., & Korie, S., 2007. Farm-scale trials to compare the entomopathogenic fungus Beauveria bassiana with pirimiphos methyl + deltamethrin and essential oil of lemon grass for protection of stored cowpea against Callosobruchus maculatus (Coleoptera : Bruchidae). Annals of Applied Biology, 151(1),1‑10. https://doi.org/10.1111/j.1744-7348.2007.00146.x
Dessauvages, K., Scheifler, M., Francis, F., & Ben Fekih, I., 2024. A New Isolate Beauveria bassiana GxABT-1 : Efficacy against Myzus persicae and Promising Impact on the Beet Mild Yellow Virus-Aphid Association. Insects, 15(9), Article 9. https://doi.org/10.3390/insects15090697
Dugje I.Y., et al., 2009. Production du niébé en Afrique de l’Ouest : Guide du paysan. IITA, Ibadan, Nigéria. 20 p. https://www.researchgate.net/publication/237489635_
El-Maziani, I., et al., 2016. Etude des dégâts qualitatifs et quantitatifs dus aux Bruches sur les légumineuses au Maroc. Revue Marocaine de Protection des Plantes, 9, 83‑99.
Guèye, M.T. et al., 2011. « Lutte contre les ravageurs des stocks de céréales et de légumineuses au Sénégal et en Afrique occidentale : synthèse bibliographique », BASE, 15(1), p. 183‑194.
Haouel Hamdi, S., et al., 2017. Nutritional alterations and damages to stored chickpea in relation with the pest status of Callosobruchus maculatus (Chrysomelidae). Journal of Asia-Pacific Entomology, 20(4), 1067‑1076. https://doi.org/10.1016/j.aspen.2017.08.008
Hidalgo, E., Moore, D., & Le Patourel, G., 1998. The effect of different formulations of Beauveria bassiana on Sitophilus zeamais in stored maize. Journal of Stored Products Research, 34(2‑3), 171‑179. https://doi.org/10.1016/S0022-474X(97)00041-6
Huignard, J.; Glitho, I.A.; Monge, J.-P.; Regnault-Roger, C., 2011. Insectes ravageurs des graines de légumineuses: Biologie des Bruchinae et lutte raisonnée en Afrique, 1st ed.; Quae: Versailles Cedex, France,Volume 146, pp. 1773–7923. https://www.quae.com/produit/1074/9782759217281/
Ileke, K. D., Adesina, J. M., Nwosu, L. C., & Olagunju, A., 2020. Perforation index assessment of cowpea seeds against cowpea bruchid, Callosobruchus maculatus (Fabricius) (Coleoptera : Chrysomelidae), infestation using Piper guineense. The Journal of Basic and Applied Zoology, 81(1). https://doi.org/10.1186/s41936-020-00195-7
Jaber, L. R., 2018. Seed inoculation with endophytic fungal entomopathogens promotes plant growth and reduces crown and root rot (CRR) caused by Fusarium culmorum in wheat. Planta, 248(6), 1525‑1535. https://doi.org/10.1007/s00425-018-2991-x
Jaber, L. R., & Enkerli, J., 2016. Effect of seed treatment duration on growth and colonization of Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biological Control, 103, 187‑195. https://doi.org/10.1016/j.biocontrol.2016.09.008
Jarrold, S. L., Moore, D., Potter, U., & Charnley, A. K., 2007. The contribution of surface waxes to pre-penetration growth of an entomopathogenic fungus on host cuticle. Mycological Research, 111(2), 240‑249. https://doi.org/10.1016/j.mycres.2006.10.007
Kanda M., et al., 2009. Le maraîchage périurbain à Lomé : pratiques culturales, risques sanitaires et dynamiques spatiales. Cahiers Agricultures, 18 (4): 356-363.https://doi.org/10.1684/agr.2009.0319
Kang, J. K., Pittendrigh, B. R., & Onstad, D. W., 2013. Insect Resistance Management for Stored Product Pests : A Case Study of Cowpea Weevil (Coleoptera: Bruchidae). Journal of Economic Entomology, 106(6), 2473‑2490. https://doi.org/10.1603/EC13340
Kaplan, E. L., & Meier, P., 1958. Nonparametric Estimation from Incomplete Observations. Journal of the American Statistical Association, 53(282), 457–481. https://doi.org/10.2307/2281868
Khoobdel, M., Pourian, H.R., & Alizadeh, M., 2019. Bio-efficacy of the indigenous entomopathogenic fungus, Beauveria bassiana in conjunction with desiccant dust to control of coleopteran stored product pests. Journal of Invertebrate Pathology, 168, 107254. https://doi.org/10.1016/j.jip.2019.107254
Kpatinvoh B., et al., 2009. Problématique de la conservation du niébé (Vigna unguiculata (L), Walp) en Afrique de l’Ouest : étude d’impact et approche de solution. Journal of Animal &Plant Sciences, 2016. Vol.31, Issue 1: 4831-4842
Kutcherov D., 2020. Stagewise resolution of temperature-dependent embryonic and postembryonic development in the cowpea seed beetle Callosobruchus maculatus (F.). BMC Ecol. 2020 Sep 11;20(1):50. doi: 10.1186/s12898-020-00318-2. PMID: 32917176; PMCID: PMC7488527.
Lacey, L. A. et al., 2015. Insect pathogens as biological control agents : Back to the future. Journal of Invertebrate Pathology, 132, 1‑41. https://doi.org/10.1016/j.jip.2015.07.009
Lugendo, A. R. et al., 2025. Efficacy of Entomopathogenic Fungi Against Bruchus rufimanus (Coleoptera : Chrysomelidae) in Laboratory and Field Trials Using Dropleg Spraying Technique. Agriculture, 15(4), Article 4. https://doi.org/10.3390/agriculture15040376
Mahdneshin Z. ; Hassan Safaralizadah M. and Ghosta Y., 2009. Study on the efficacy of iranian Isolates of Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metsch.) Sorokin against Rhyzopertha dominica F. (Coleoptera : Bostrichidae) (p. Journal of Biological Sciences 9 (2):170-174).
Meadow, R., Vandenberg, J. D., & Shelton, A. M., 2000. Exchange of Inoculum of Beauveria bassiana (Bals.) Vuill. (Hyphomycetes) Between Adult Flies of the Cabbage Maggot Delia radicum L.(Diptera :Anthomyiidae). Biocontrol Science and Technology, 10(4), 479‑485. https://doi.org/10.1080/09583150050115061
Mofunanya, A. A. J., & Namgbe, E. E., 2016. Assessment of damage due to Callosobruchus maculatus (Coleoptera: Bruchidae) infestation on germination and nutrient quality of Vigna unguiculata L.(Walp). International Organization of Scientific Research (IOSR) Journal of Agriculture and Veterinary Science, 9(12), 96-101.
Mudassir, M. et al., 2015. Post-exposure temperature influence on the toxicity of conventional and new chemistry insecticides to green lacewing Chrysoperla carnea ( Stephens ) ( Neuroptera : Chrysopidae ). Saudi Journal of Biological Sciences, 22(3), 317‑321. https://doi.org/10.1016/j.sjbs.2014.10.008
Muniz, E. R. et al., 2020. Efficacy of Metarhizium anisopliae conidia in oil-in-water emulsion against the tick Rhipicephalus microplus under heat and dry conditions. BioControl, 65, 339‑351. https://doi.org/10.1007/s10526-020-10002-5
Ozdemir, I. O., Tuncer, C., Erper, I., & Kushiyev, R., 2020. Efficacy of the entomopathogenic fungi; Beauveria bassiana and Metarhizium anisopliae against the cowpea weevil, Callosobruchus maculatus F. (Coleoptera : Chrysomelidae : Bruchinae). Egyptian Journal of Biological Pest Control, 30(1), 20‑24. https://doi.org/10.1186/s41938-020-00219-y
Pande, R., & Mishra, H. N., 2018. Efficacy of non formulated entomopathogenic fungi (Beauveria bassiana) to control the storage insect pests of green gram (Vigna radiata) seeds. Indian Journal of Agricultural Research, 52(3), 295‑299. https://doi.org/10.18805/IJARe.A-4913
Quesada-Moraga, E. et al., 2008. « Horizontal transmission of Metarhizium anisopliae among laboratory populations of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) », Biological Control, 47(1), p. 115‑124.https://doi.org/10.1016/j.biocontrol.2008.07.002.
Quesada-Moraga, E. et al., 2023. Key role of environmental competence in successful use of entomopathogenic fungi in microbial pest control. Journal of Pest Science, 97, 1‑15. https://doi.org/10.1007/s10340-023-01622-8
Rabé, M. M., Baoua, I. B., & Baributsa, D., 2023. Enhancing Cowpea Productivity in the Sahel : Exploring Seed Access among Smallholder Farmers in South-Central Niger. Horticulturae, 9(12), Article 12. https://doi.org/10.3390/horticulturae9121287
Segers, A., Carpentier, J., Francis, F., & Caparros Megido, R., 2023. Gene Silencing of laccase 1 Induced by Double-Stranded RNA in Callosobruchus maculatus (Fabricius 1775) (Coleoptera : Chrysomelidae) Suggests RNAi as a Potential New Biotechnological Tool for Bruchid’s Control. Agriculture, 13(2), Article 2. https://doi.org/10.3390/agriculture13020412
Segers A. et al., 2021. Susceptibility of Bruchus rufimanus Boheman 1833 (Coleoptera: Chrysomelidae) to three entomopathogenic fungi : Limits of conidial suspensions sprayings and potential alternatives in integrated pest management strategy. 2021 International Congress on Invertebrate Pathology and Microbial Control & 53rd Annual Meeting of the Society for Invertebrate Pathology. http://dx.doi.org/10.13140/RG.2.2.23290.06086
Seri-Kouassi, B. P. et al., 2004. Action des huiles essentielles de deux plantes aromatiques de Côte-d’Ivoire sur Callosobruchus maculatus F. du niébé. Comptes Rendus Chimie, 7(10‑11), 1043‑1046. https://doi.org/10.1016/j.crci.2003.12.031
Storm, C. et al., 2016. Improving Efficacy of Beauveria bassiana against Stored Grain Beetles with a Synergistic Co-Formulant. Insects, 7(3), https://doi.org/10.3390/insects7030042
Swella, G.B. & Mushobozy, D.M.K. 2007. « Evaluation of the efficacy of protectants against cowpea bruchids (Callosobruchus maculatus (F.)) on cowpea seeds (Vigna unguiculata (L.) Walp.) », Plant Protection Science, 43(2), p. 68‑72. https://doi.org/10.17221/2256-PPS.
Viaud, P. 1983. La protection des Légumineuses contre les Bruches : Vigna unguiculata et Callosobruchus maculatus [Col.]. Bulletin de la Société entomologique de France, 88(3), 241‑249. https://doi.org/10.3406/bsef.1983.18306
Vinayaga Moorthi, P., Balasubramanian, C., Selvarani, S., & Radha, A., 2015. Efficacy of sub lethal concentration of entomopathogenic fungi on the feeding and reproduction of Spodoptera litura. SpringerPlus, 4(1), 681. https://doi.org/10.1186/s40064-015-1437-1
Williamson S., Ball A. & Pretty J., 2008. Trends in pesticide use and drivers for safer pest management in four African countries. Crop Protection, 27 (10): 1327-1334. https://doi.org/10.1016/j.cropro.2008.04.006
Zimmermann, G. 2007. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Science and Technology, 17(6), 553‑596. https://doi.org/10.1080/09583150701309006
Zongo, S., Z, I., Waongo A., G. O., Doumma A., S. M., & Sanon., A., 2015. Risques Liés à l’utilisation d’insecticides au cours du stockage du niébé ( Vigna Unguiculata L. Walp .) dans la région centrale du Burkina-Faso. Rev. Cames, Vol.03, Num.01. 2015. ISSN 2424-7235. https://www.researchgate.net/publication/296831078
Zongo, S., et al., 2021. Metabolic resistance to pyrethroids (Py) and organophosphates (Op) in Callosobruchus maculatus (fab.) (Coleoptera: Chrysomelidae: Bruchinae) a major pest of stored cowpeas in West Africa. International Journal of Pest Management, 67(4), 338–345. https://doi.org/10.1080/09670874.2020.1778812
Pour citer cet article
A propos de : Adrien Rodrigue Lugendo*
Entomologie fonctionnelle et évolutive, Université de Liège-Gembloux Agro-Bio Tech, Gembloux, Belgique
Département d’agrovétérinaire, Institut Supérieur Pédagogique de Bukavu (ISP-Bukavu), Bukavu, Sud-Kivu, République Démocratique du Congo
A propos de : Arnaud Segers
Entomologie fonctionnelle et évolutive, Université de Liège-Gembloux Agro-Bio Tech, Gembloux, Belgique
A propos de : Ibtissem Ben Fekih
Entomologie fonctionnelle et évolutive, Université de Liège-Gembloux Agro-Bio Tech, Gembloux, Belgique
A propos de : Rudy Caparros Megido
Entomologie fonctionnelle et évolutive, Université de Liège-Gembloux Agro-Bio Tech, Gembloux, Belgique
A propos de : Gaston Amzati Sefu
Faculté des sciences Agronomiques, Université Evangélique en Afrique (UEA/Bukavu), Bukavu, Sud-Kivu, République Démocratique du Congo
A propos de : Frédéric Francis
Entomologie fonctionnelle et évolutive, Université de Liège-Gembloux Agro-Bio Tech, Gembloux, Belgique
