


- Startpagina tijdschrift
- Volume 37 (2019)
- Numéro 2
- Variabilité des caractères morphologiques mesurables de la chèvre Djallonké dans les zones agro-écologiques du Togo


Weergave(s): 10418 (15 ULiège)
Download(s): 0 (0 ULiège)


Variabilité des caractères morphologiques mesurables de la chèvre Djallonké dans les zones agro-écologiques du Togo

Nota's van de redactie
Reçu le 06. 05.18 et accepté pour publication le 24.04.19
Résumé
Au Togo, les petits ruminants jouent un rôle très important dans la vie socio – économico – culturelle et les caprins sont plus nombreux que les ovins. La race caprine la plus rencontrée est la chèvre Djallonké ou chèvre naine. Le mode d’élevage généralement traditionnel de cette race favorise des croisements anarchiques et pourrait constituer une vraie menace pour la race. Cependant, peu d’études ont été faites pour caractériser cette race qui est présente dans toutes les régions du pays et dans beaucoup de pays de la sous - région. Dans la présente étude, des enquêtes ont été faites à travers six régions agro-écologiques du pays (la zone du littoral, la zone forestière, la zone de Savane guinéenne, la zone de l’Atakora, la forêt sèche et la zone soudanienne). Au total, 330 caprins dont 35 mâles et 295 femelles ont été choisis au hasard dans les élevages et ont fait l’objet de mensurations. Les paramètres morpho – biométriques mesurés ont été les suivants : la hauteur au garrot (HG), la longueur scapulo- ischiale (LSI), le tour de poitrine (TP), la longueur des cornes (LC), la longueur des oreilles (LO), la longueur de la queue (LQ), la longueur des poils (LP), et le poids vif (PV). Les moyennes obtenues pour ces paramètres sont les suivantes: HG = 41,56 ± 3,66 cm ; LSI = 50,62 ± 4,55 cm ; TP = 61,28 ± 5,40 cm ; LC = 6,52 ± 2,22 cm, LO = 9,42 ± 0,90 cm ; LQ = 9,17 ± 1,12 cm ; LP = 2,55 ± 1,05 cm et PV = 17,51 ± 4,43 kg. Des analyses de variances (ANOVA) et des analyses multivariées (ACP et AFD) montrent que ces résultats sont comparables à ceux trouvés par plusieurs auteurs dans la sous - région pour cette race.
Abstract
Variability of Measurable Morphological Characters of the West African Dwarf Goats (WAD goats) in the Agro-ecological Areas of Togo
In Togo, small ruminants play a very important socio - economic - cultural role. Goats are the most numerically important compared to sheep. The most encountered goat breeds in this side of Africa are the West African Dwarf goats (WAD goats). Traditionally, extensive raising methods used for this race favors anarchic cross breeding and may constitute a real threat for the race. However, only few studies have been carried on to characterize this race, which exists in all Agro-ecological regions of the country, and also in the neighboring countries. In this study, surveys have carried on in the six Agro-ecological regions (Coastal, Forest, Guinea-Savannah, Atakora, Dry forest, and Sudan). 330 goats in total, including 35 males, were randomly selected in farms and were subject to measurements. Data have been analyzed using variance analysis (ANOVA) and multivariate analysis (PCA and AFD). These methods resulted in the means of morphological - biometric parameters, like: height at the withers (41.56 ± 3.66 cm), scapular ischial length (50.62 ± 4.55 cm), chest (61.28 ± 5.40 cm), horn length (6.52 ± 2.22 cm), ear length (9.42 ± 0.90 cm) tail length (9 17 ± 1.12 cm), hair length (2.55 ± 1.05 cm) and body weight (17.51 ± 4.43 kg). These figures are the same for this race in comparison to those found by other authors in the sub – region.
Inhoudstafel
Introduction
1L’élevage reste la deuxième activité pratiquée par 93,4% des agriculteurs (8). Il assure de multiples fonctions dans la vie des éleveurs et des pays. Plus de 600 millions de personnes vulnérables dépendent des activités liées à l’élevage pour leur survie (11). Au Togo, l’élevage est pratiqué par la plupart des ménages, et l’élevage des petits ruminants vient en seconde position après celui des volailles en matière d’effectifs. Les conditions climatiques sont favorables à la pratique de l’élevage des petits ruminants à cause de la disponibilité du pâturage et de la faible pression parasitaire. La chèvre naine de l’Afrique de l’Ouest encore appelée chèvre Djallonké est celle qui est la plus rencontrée à travers tout le pays (4).
2La chèvre Djallonké se caractérise par une précocité de reproduction, une bonne prolificité, une aptitude à se reproduire tout au long de l’année et une adaptation aux conditions climatiques tropicales. Malgré ces qualités, la chèvre Djallonké présente un poids adulte faible variant entre 20 – 25 kg pour les boucs et entre 18 – 20 kg pour les chèvres, contrairement aux poids moyens des sujets de la race sahélienne de plus en plus rencontrée dans les zones péri- urbaines du pays dont les poids moyens sont de 40 kg pour les boucs et 27 – 30 kg pour les femelles (22). Dans le souci d’améliorer les performances des caprins Djallonké, les éleveurs et certains projets de développement de l’élevage, font des croisements avec des races sahéliennes ou européennes plus performantes pour des fins de production de viande ou de lait. Compte tenu du mode d’élevage généralement traditionnel de cette race, qui favorise des croisements anarchiques, la race Djallonké est menacée de disparition. Peu d’études ont été faites sur la caractérisation de cette race au Togo. Les travaux d’Amegée a fait des recherches en 1986 sur les performances d’engraissement de la chèvre Djallonké au Togo et en 1987 sur ses performances de reproduction. Peu d’études ont été faites dans le pays non sur sa caractérisation tant morphologique que génétique. Une description phénotypique reposant sur une analyse statistique des traits morphométriques contribuerait donc à définir une morphologie standard et pourrait guider les programmes d'amélioration génétique à l'avenir (15) de cette race dans le pays. La présente étude vise la caractérisation bio – morphologique de la race Djallonké dans les élevages villageois du Togo.
Matériel et méthode
Site de l’étude
3Cette étude a couvert tout le territoire national divisé en six zones agro écologiques caractérisées comme suit:
-
la zone du littoral (région Maritime) qui est caractérisée par un côtier frais et une végétation moins dense ; rencontre des cultures et des jachères, des fourrés, des buissons, des savanes herbeuses littorales, des savanes à termitières buissonnantes qui coexistent avec les îlots de forêt (16). Les mangroves, les prairies et savanes inondables sont également présentes à l’extrême sud-est du pays (2).
-
la zone de forêt mésophile qui correspond à la zone humide de l’Atakora ou région des plateaux ouest, elle est caractérisée par des précipitations abondantes et une strate de sous - bois des herbes à larges feuilles. La zone des monts du Togo correspond à la partie méridionale appelée unité de l’Atakora (23). Elle est constituée des plaines du Litimé et d’Ahlon, les plateaux Akébou, Akposso, Dayes et Kloto.
-
la zone de savane humide abondamment arrosée ou région des plateaux - est, c’est une savane arborée, constituée de l’ensemble du dahoméen dans le bassin du Mono ;
-
la zone de l’Atakora correspondant à la région Centrale, elle couvre partiellement la zone des massifs de collines avec une strate herbacée composée de graminées en touffes : Andropogon sp. , Hypparrhenia sp… Cette zone correspond aux grandes étendues de plaines du centre du pays avec des altitudes situées entre 200 et 400 mètres. Elle appartient aux vastes pénéplaines précambriennes méridionales et centrales du pays. Les formations végétales dominantes sont “des savanes boisées guinéennes” plus ou moins arborées (6). On y trouve également des forêts claires et des forêts galeries discontinues le long des principaux cours d’eau.
-
la zone des montagnes du nord - est ou région Kara, elle est une caractérisée par le massif Kabyè et les monts Défalé qui encadrent le plateau de Niamtougou et la plaine de la Binah. C’est une zone de forêts denses sèches, de forêts claires et de savanes herbeuses à arborées avec Parkia biglobosa, Vitellaria paradoxa et Isoberlinia spp. La strate herbacée est composée de graminées en touffes ;
-
la zone Soudanienne ou la région des savanes et des plaines du nord correspondent à la partie septentrionale du pays. Elle fait partie des formations de couvertures du bassin de la Volta. Le principal type de végétation est la savane arborée soudanienne où subsistent quelques lambeaux de forêt-galerie le long des berges de l’Oti. Cette végétation est aussi caractérisée par la présence d’Andansonia digitata, des arbres épineux et une strate herbacée constituée des herbes Aristida sp, Cloris sp, Hypparrrhenia sp….
4L’étude a concerné 18 préfectures à raison de trois préfectures par zone agro écologique. Les sites ou lieux de mensurations sont mentionnés sur la figure 1.
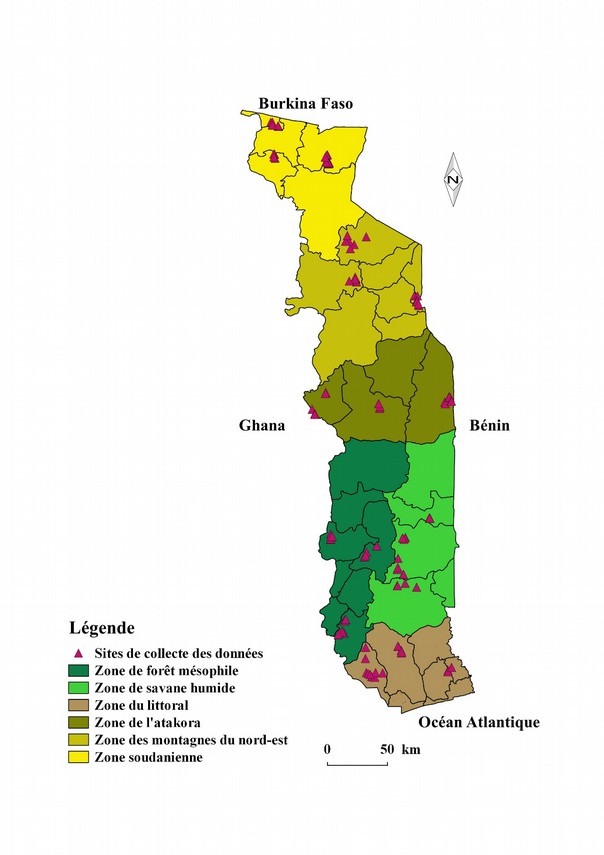
Figure 1 : Carte du Togo indiquant les lieux des mensurations.
Matériel animal
5L’étude a porté sur trois cent trente (330) caprins de race Djallonké dont 35 mâles et 295 femelles appartenant à 135 élevages différents choisis au hasard dans les dix - huit préfectures. Par rapport à l’âge, les caprins ayant fait l’objet de cette étude avaient un âge compris entre un an et plus de quatre ans, répartis comme suit : un à deux ans (111 têtes), entre deux et trois ans (112 têtes), entre trois et quatre ans (60 têtes) et quatre ans et plus (47 têtes). La photo 1 montre les chèvres Djallonké dans différentes régions.

Photo 1 : Chèvres Djallonké dans différentes régions.
Méthode
Echantillonnage des éleveurs et des animaux
6Les élevages ont été choisis dans les différentes zones agro – écologiques. Le choix des éleveurs s’est effectué en collaboration avec un conseiller agricole et les élevages étaient éloignés les uns des autres d’une distance d’au moins un kilomètre afin de réduire le risque de prendre des animaux apparentés compte tenu de la divagation surtout en saison sèche.
7Au sein d’un élevage, les mensurations sont faites sur deux à cinq animaux ayant au moins l’âge d’un an déterminé à partir du contrôle de la dentition. Au moins un animal selon le nombre et l’état des dents adultes est choisi en commençant par ceux ayant deux dents adultes, puis en terminant par ceux ayant les huit dents en état d’usure. Par rapport au sexe, s’il y a des mâles adultes castrés ou entiers, ils sont tous choisis pour la mensuration. Seuls les animaux présentant les caractéristiques des chèvres Djallonké (absence de pendeloques, oreilles non pendantes et taille moyenne) sont choisis pour les mensurations.
Mensurations morpho - biométriques
8Sur les animaux, les caractères quantitatifs suivants ont été mesurés :
-
la hauteur au garrot (HG), elle est mesurée de la pointe des pieds au bout de l’épaule de la chèvre ;
-
la longueur scapulo - ischiale (LSI), c’est la longueur mesurée entre la pointe de l’épaule et la pointe de la croupe ;
-
le tour thoracique (TT), c’est la circonférence prise au niveau du thorax de l’animal ;
-
la taille des oreilles (LO), c’est la longueur comprise entre la base et la pointe des oreilles ;
-
la longueur des cornes (LC) est mesurée de la base au sommet de la corne en suivant la courbure de celle – ci ;
-
la longueur de la queue (LQ); elle est prise à l’aide d’une règle décimètre, la queue reste tendue sur cette règle plate afin de noter sa longueur ;
-
la longueur des poils (LP); ce sont les poils au niveau de la nuque qui sont prélevés et mesurés à l’aide d’un décimètre ;
-
le poids vif de l’animal (PV) ; déterminé après la pesée de l’animal avec une balance de portée 50 kg ayant une précision de 200 grammes.
9Toutes les longueurs ont été relevées avec un mètre ruban, une canne-toise pour la hauteur au garrot, une balance de portée 50 kg avec une précision de 200 grammes pour le poids vif (PV) des animaux.
Analyses statistiques des données
10Les données ont été traitées avec le logiciel SPSS 20 pour l’analyse des variances et des moyennes. XLSTAT a permis de faire l’Analyse en Composantes Principales (ACP) et l’Analyse Factorielle Discriminante (AFD) au seuil de signification de 5%.
Résultats
Analyse descriptives des données quantitatives mesurées
11Les caractères quantitatifs mesurés sur les caprins au Togo concernent 330 caprins dont 35 boucs (soit 11%) et 295 chèvres (89%). La structure des troupeaux est largement en faveur des femelles au sein des troupeaux. Les tableaux 1 à 3 traduisent les résultats des analyses des caractères quantitatifs selon les régions, le sexe et l’âge des caprins.
Hauteur au garrot (HG)
12La moyenne de la hauteur au garrot pour les 330 caprins de l’étude est de 41,56 ± 3,66 cm.
13Le facteur « zone agroécologique » a eu un effet sur la hauteur au garrot des caprins de cet échantillon (Tableau 1). Les chèvres issues des zones littorales, de la zone forestière et de la savane humide situées dans la partie méridionale du pays sont plus courtes sur pied que leurs soeurs de la zone septentrionale (zone soudanienne).
14Le sexe n’a pas eu d’influence sur la hauteur au garrot des animaux dans cette étude (Tableau 2).
15L’effet de l’âge sur la hauteur au garrot de la chèvre Djallonké dans cette étude est ressorti dans l’analyse statistique. Les chèvres âgées de trois ans au moins sont plus hautes que les jeunes chèvres (Tableau 3).
Tableau 1 : Résultats des analyses des différents paramètres bio- morphologiques des caprins selon les zones agro écologiques.
|
Région |
HG |
TT |
LSI |
LQ |
TO |
LP |
LC |
Poids |
|
MARITIME |
39,79 ± 3,86 c |
59,88 ± 4,68 |
47,6 ± 4 c |
8,75 ± 1,25 c |
9,22 ± 0,94 b |
2,94 ± 1,63 a |
6,52 ± 2,09 ab |
15,30 ± 4,62 b |
|
PLTO EST |
41,16 ± 2,9 bc |
62,26 ± 6,14 |
50,02 ± 4,17 b |
9,38 ± 0,99 ab |
9,76 ± 1,02 a |
2,57 ± 0,73 a |
7,36 ± 2,64 a |
17,63 ± 4,41 a |
|
PLTO OUE |
41,19 ± 4,12 bc |
60,74 ± 3,93 |
49,5 ± 3,93 b |
9,09 ± 1,08 bc |
9,22 ± 0,93 b |
2,74 ± 1,25 a |
6,51 ± 2,05 ab |
17,64 ± 3,3 a |
|
CENTRALE |
41,78 ± 3,6 b |
62,1 ± 6,28 |
52,75 ± 4,49 a |
9,62 ± 1,26 a |
9,6 ± 0,84 ab |
2,43 ± 0,87 ab |
6,7 ± 2,02 ab |
18,04 ± 4,5 a |
|
KARA |
41,52 ± 3,43 bc |
61,33 ± 5,32 |
50,65 ± 3,97 b |
9,02 ± 1,04 bc |
9,34 ± 0,88 ab |
2,58 ± 0,96 a |
5,88 ± 2,09 b |
18,13 ± 4,2 a |
|
SAVANES |
43,77 ± 2,99 a |
61,2 ± 5,62 |
52,93 ± 4,68 a |
9,15 ± 0,98 abc |
9,39 ± 0,66 ab |
2,08 ± 0,33 b |
6,17 ± 1,82 b |
18,63 ± 4,40 a |
|
Moyenne ± SD |
41,54 ± 3,48 |
61,25 ± 5,33 |
50,58 ± 4,21 |
9,17 ± 1,1 |
9,42 ± 0,88 |
2,56 ± 0,96 |
6,53 ± 2,12 |
17,45 ± 4,33 |
|
F (5%) |
7,11 |
1,39 |
11,98 |
3,9 |
3,4 |
4,31 |
3,15 |
3,88 |
|
Probabilité (%) |
< 0,01 |
22,5 |
< 0,01 |
0,2 |
0,5 |
0,1 |
0,9 |
0,2 |
HT : hauteur au garrot ; TT : tour thoracique ; LSI : longueur scapulo – ischiale ; LQ : longueur de la queue ; TO : taille des oreilles ; LP : longueur des poils ; LC : longueur des cornes
Sur une colonne les moyennes portant les lettres différentes sont significativement différentes
Tableau 2 : Résultats des analyses des différents paramètres bio- morphologiques des caprins selon le sexe des caprins.
|
Sexe |
HG |
TT |
LSI |
LQ |
TO |
LP |
LC |
Poids |
|
F |
41,67 ± 3,48 |
61,56 ± 5,28 a |
50,97 ± 4,27 a |
9,15 ± 1,1 |
9,45 ± 0,91 |
2,52 ± 1,08 |
6,42 ± 2,11 a |
17,71 ± 4,4 a |
|
M |
40,69 ± 4,92 |
58,92 ± 5,96 b |
47,69 ± 5,74 b |
9,38 ± 1,27 |
9,26 ± 0,79 |
2,78 ± 0,69 |
7,43 ± 2,53 b |
15,83 ± 4,43 b |
|
Moyenne ± SD |
41,18 ± 4,2 |
60,24 ± 5,62 |
49,33 ± 5 |
9,27 ± 1,19 |
9,35 ± 0,85 |
2,65 ± 0,89 |
6,93 ± 2,32 |
16,77 ± 4,41 |
|
F5% |
2,26 |
7,65 |
17,04 |
1,24 |
1,31 |
1,83 |
6,35 |
5,72 |
|
Probabilité (%) |
13,3 |
0,6 |
< 0,01 |
26,7 |
25,3 |
17,7 |
1,2 |
1,7 |
Sur une colonne les moyennes portant les lettres différentes sont significativement différentes
Tableau 3 : Résultats des analyses des différents paramètres bio- morphologiques des caprins selon l’âge.
|
Age (an) |
HG |
TT |
LSI |
LQ |
TO |
LP |
LC |
Poids |
|
1-2 ans |
39,89 ± 4,22 b |
55,54 ± 3,39 d |
47,31 ± 4,04 c |
9,00 ± 1,17 |
8,97 ± 0,67 b |
2,35 ± 0,63 |
5,56 ± 1,90 c |
13,47 ± 2,16 c |
|
2- 3 ans |
40,02 ± 3,10 b |
58,06 ± 4,69 c |
47,89 ± 3,94 c |
9,20 ± 1,19 |
9,33 ± 0,87 ab |
2,78 ± 0,97 |
5,72 ± 1,98 c |
14,72 ± 3,24 c |
|
3- 4 ans |
42,49 ± 4,00 a |
62,28 ± 4,14 b |
51,43 ± 3,95 b |
9,25 ± 1,20 |
9,59 ± 0,94 a |
2,52 ± 1,01 |
6,40 ± 1,83 b |
18,20 ± 3,81 b |
|
≥ 4 ans |
42,25 ± 3,02 a |
64,19 ± 5,16 a |
52,74 ± 4,23 a |
9,12 ± 0,97 |
9,43 ± 0,90 a |
2,44 ± 1,20 |
7,56 ± 2,31 a |
20,00 ± 4,39 a |
|
Moyenne ± SD |
41,16 ± 3,58 |
60,02 ± 4,35 |
49,84 ± 4,04 |
9,15 ± 1,13 |
9,33 ± 0,84 |
2,52 ± 0,96 |
6,31 ± 2,01 |
16,60 ± 3,4 |
|
F5% |
11,52 |
43,69 |
30,1 |
0,51 |
4 |
2,11 |
15,23 |
42,1 |
|
Probabilité (%) |
< 0,01 |
< 0,01 |
< 0,01 |
67,9 |
0,8 |
9,8 |
< 0,01 |
< 0,01 |
Sur une colonne les moyennes portant les lettres différentes sont significativement différentes
Tableau 4 : Matrice de corrélation (Pearson (n))
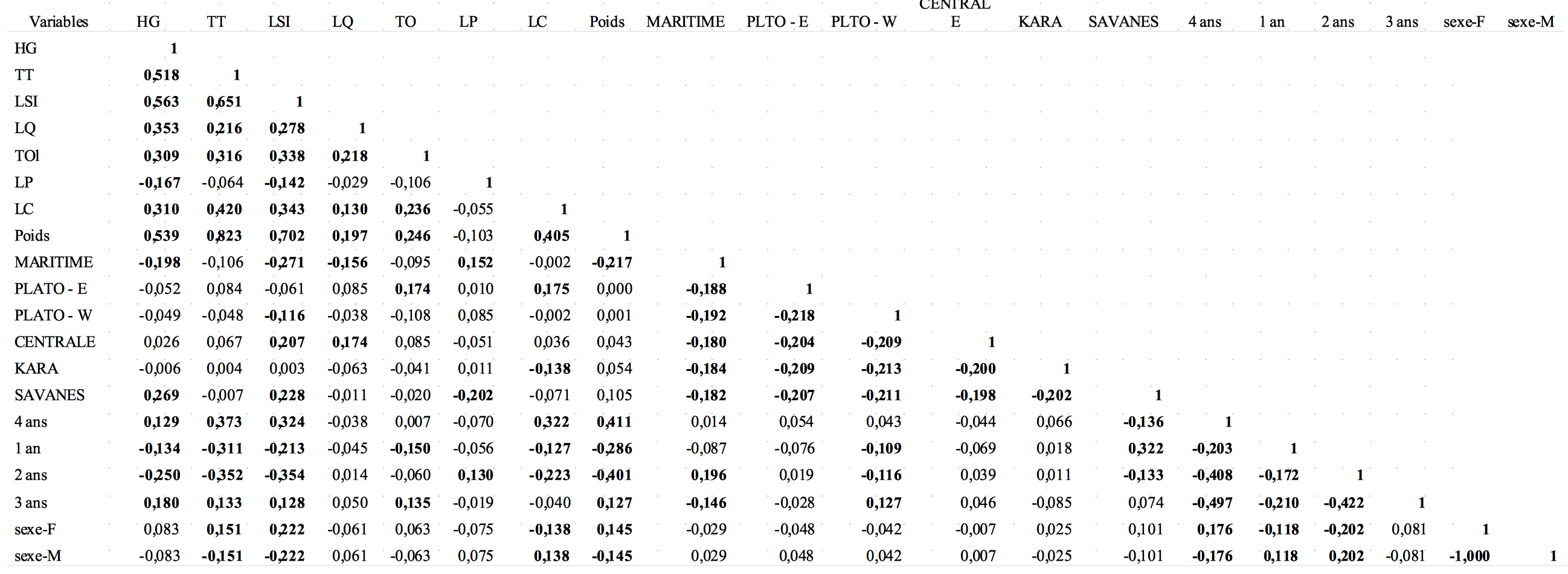 Les valeurs en gras sont significativement différentes de 0 à un niveau de signification alpha=0,05
Les valeurs en gras sont significativement différentes de 0 à un niveau de signification alpha=0,05
Tour Thoracique (TT)
16La moyenne pour les 330 échantillons est de 61,28 ± 5,40 cm. L’analyse statistique montre qu’il n’y a aucune influence de la zone agro-écologique sur ce facteur. Les chèvres de cette étude ont un tour thoracique comparable dans toutes les zones.
17Par rapport au sexe, les femelles ont un tour thoracique plus large (61,56 ± 5,28) que les mâles (58,92 ± 5,96). Ceci s’explique par le fait que les mâles trouvés lors de l’échantillonnage étaient plus jeunes que les femelles.
18L’âge des animaux a eu un effet significatif (F = 43,69 à P=1%) sur le TT. En effet, le TT augmente avec l’âge des animaux. Les caprins de quatre ans au moins ont eu le TT le plus large (64,19 ± 5,16).
Longueur scapulo – ischiale (LSI)
19La moyenne générale de la LSI pour les 330 caprins est de 50,62 ± 4,55 cm.
20L’analyse statistique a montré que la moyenne de la LSI des caprins de la zone du littoral est significativement plus faible (F = 11,98 à P = 1%) par rapport à celles des caprins des autres régions. Les caprins des régions savanes et centrale étaient les plus longs. La LSI suit la tendance de la hauteur au garrot.
Longueur de la queue (LQ)
21La longueur moyenne de queue de l’échantillon de l’étude est de 9,17 ± 1,12 cm. C’est une queue courte le plus souvent relevée. La différence n’est très significative entre les 6 zones agroécologiques. Les facteurs sexe et âge n’ont pas eu d’effet sur la longueur de la queue.
Longueur des oreilles (TO)
22La longueur moyenne des oreilles des caprins étudiés est de 9,42 ± 0,90 cm.
Longueur des cornes (LC)
23La longueur moyenne des cornes est de 6,52 ± 2,22 cm. La longueur moyenne des cornes a significativement varié d’une région à une autre. Les caprins de la région des Plateaux ont des cornes plus longues que ceux des régions de la Kara et des Savanes qui ont des cornes plus courtes.
Longueur des poils (LP)
24Les poils de tous les animaux sont courts, la moyenne pour l’ensemble est de 2,55 ± 1,05 cm. Les caprins de la région des Savanes, proche du Sahel ont les poils les moins longs par rapport aux caprins des autres régions. L’âge et le sexe n’ont eu aucun effet significatif sur la longueur des poils. Avec de telles longueurs des poils, la chèvre naine du Togo peut être classée comme animaux à poils ras.
Poids des animaux
25Le poids de l’échantillon de cette étude est en moyenne de 17,56 ± 4,32 kg. Mis à part les chèvres de la région Maritime qui ont une moyenne de poids 15,30 ± 4,62 kg significativement différente, les moyennes de poids des chèvres des autres régions sont presque identiques, 17,63 ± 4,41 kg dans les Plateaux Est et 18,63 ± 4,40 kg dans les Savanes. L’analyse statistique a montré qu’il existe une corrélation entre le poids moyen des caprins et la valeur moyenne du TT. Cette corrélation est traduite par la figure 2 qui représente la variation du poids des caprins en fonction du tour thoracique, cette corrélation est traduite par la droite d’équation :
26Y= 0,659x – 22,25 : x étant la valeur du TT en cm et Y le poids en kg avec comme coefficient de détermination ajusté de 0,678.
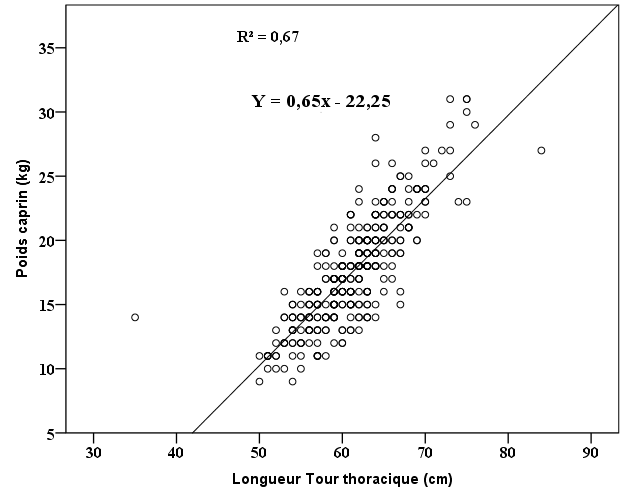
Figure 2 : Variation du poids des caprins en fonction de leur tour thoracique.
Analyse multivariée des paramètres mesurés
Analyse en Composantes Principales (ACP)
27Le tableau 4 montre les relations de corrélations qui existent entre les différents caractères phénotypiques quantitatifs étudiés en variables principales d’une part et entre ces caractères et les facteurs « région » et « sexe » en variables supplémentaires. Les deux premiers axes F1 et F2 de l’ACP sont les seuls dont les valeurs propres sont supérieures à 1. Ils résument environ 56,48% des observations dans le cadre de notre travail. La figure 3 montre que toutes les variables (actives et supplémentaires) sont représentées sur le plan factoriel F1 et F2. Par ailleurs, une forte corrélation positive est observée entre la HG, la LSI, le TT, la TO, la LC, la LQ et le poids des animaux avec les régions Savanes, Centrale et Plateaux Est. En outre, ces paramètres sont corrélés positivement avec le sexe femelle et négativement au sexe mâle qui se retrouve dans le plan opposé à celui des paramètres étudiés. Les âges de 3 et 4 ans aussi sont corrélés avec ces paramètres.
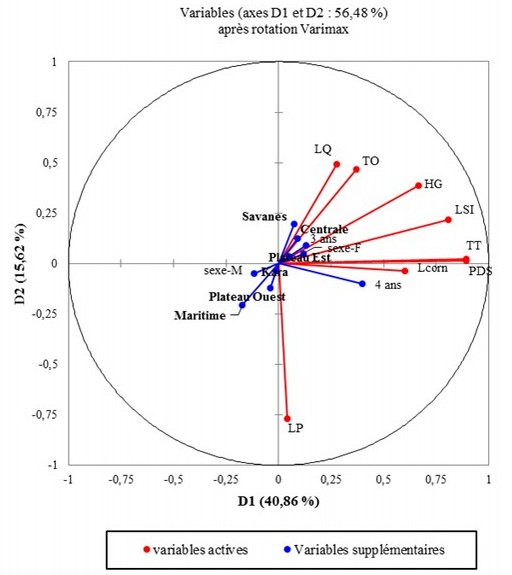
Figure 3: Cercle de corrélation ACP.
Analyse Factorielle Discriminante
28Avec les facteurs « région » et « sexe » comme variables explicatives et les mesures morpho - biométriques (HG, TT, LSI, LQ, TO, LC, LP, Poids) et l’âge des caprins comme variables dépendantes, les axes F1 et F2 de la figure 4 expliquent 82,66% de la variabilité entre les différentes zones agroécologiques. La figure 5 montre que les facteurs les plus discriminants sur les deux axes et qui caractérisent mieux les caprins sont le TT, la LSI, la LO, la LC et la LQ. Il ressort de la matrice de confusion du tableau 5 que les individus sont bien classés dans 39,51% seulement et que 200 individus soit 60,49% ont été reclassés. Ce taux élevé de reclassement prouve que « la région » n’est pas un facteur très influent dans le regroupement des caprins au Togo. D’autres facteurs intrinsèques liés aux animaux ou au système d’élevage pourront expliquer les différences.
29Tout comme dans le cas de l’ACP, les figures (4 et 5) de l’AFD ont fait ressortir l’absence totale de groupes distincts de caprins. Les caprins de la Savane sont caractérisés par une HG et une LSI élevées associées à des poils et des cornes courts. Les caprins de la région Centrale sont caractérisés par une LSI, une LQ, une LC et une HG élevées. Les caprins de la région Maritime se distinguent des autres par leur petite taille (HG, LSI), un poids moyen avec une LP plus élevée.
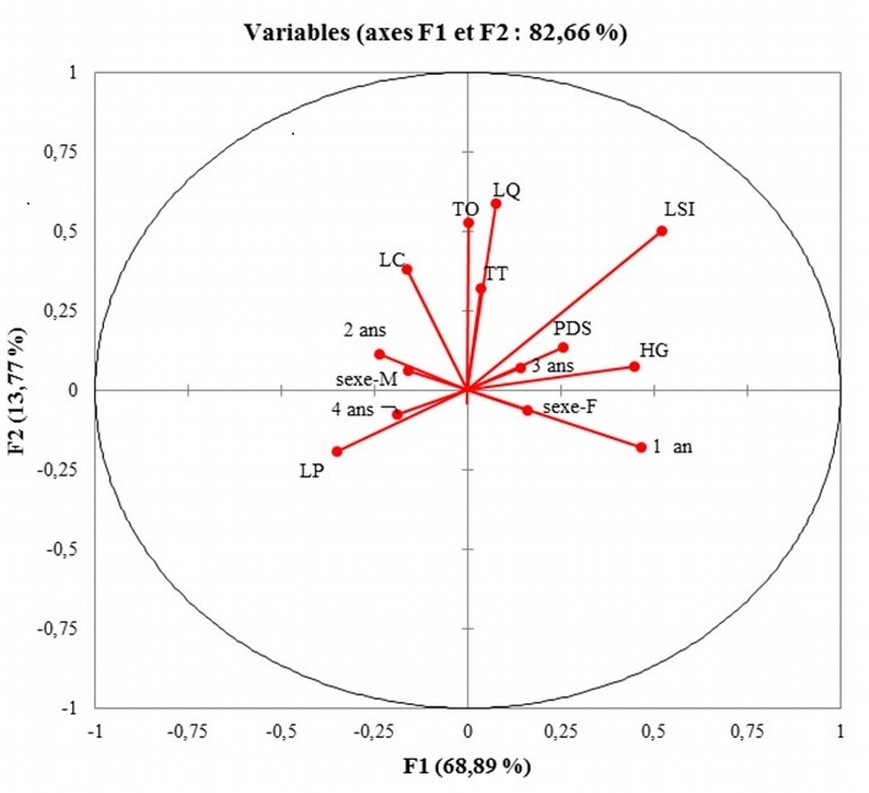
Figure 4: Cercle de corrélation AFD.
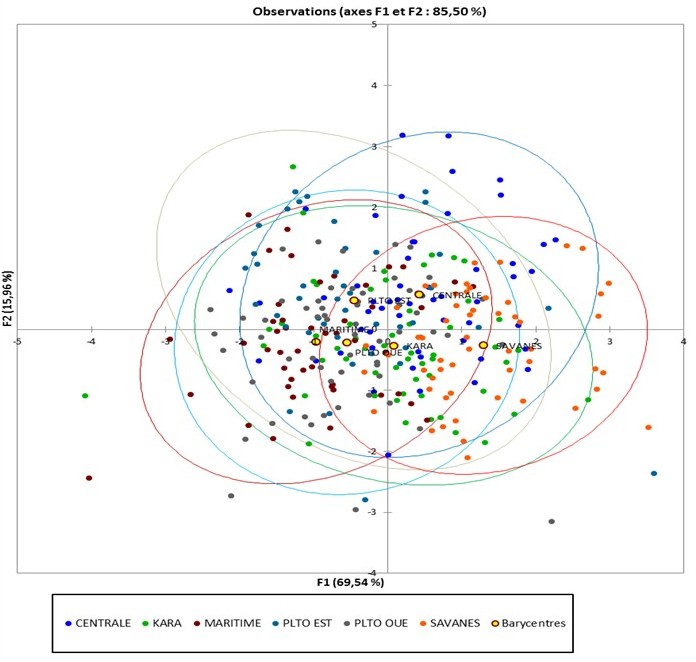
Figure 5 : Répartition des individus dans le plan factoriel (F1 – F2)
Tableau 5 : Matrice de confusion pour l'échantillon d'estimation (région)
|
de \ Vers |
Centrale |
Kara |
Maritime |
Plateau Est |
Plateau Ouest |
Savanes |
Total |
% correct |
|
Centrale |
19 |
8 |
10 |
5 |
4 |
8 |
54 |
35,19% |
|
Kara |
9 |
13 |
7 |
6 |
13 |
8 |
56 |
23,21% |
|
Maritime |
3 |
3 |
21 |
12 |
8 |
0 |
47 |
44,68% |
|
Plateau Est |
8 |
7 |
9 |
23 |
11 |
0 |
58 |
39,66% |
|
Plateau Ouest |
10 |
6 |
8 |
10 |
23 |
3 |
60 |
38,33% |
|
Savanes |
12 |
5 |
0 |
0 |
6 |
31 |
54 |
57,41% |
|
Total |
61 |
42 |
55 |
56 |
65 |
50 |
329 |
39,51% |
Discussion
30Le taux de présence des boucs dans les élevages observé lors de cette enquête est inférieur à celle des troupeaux du Botswana 31% pour les mâles et 69% pour les femelles (16). Le faible taux des mâles est le résultat d’une exploitation précoce de ceux – ci par les éleveurs qui gardent plus longtemps les femelles. Différentes raisons sont invoquées par ceux - ci pour justifier cette exploitation précoce des mâles : i) le vol car selon les éleveurs, les boucs entiers divaguent plus et donc sont susceptibles d’être volés ; ii) la constitution du noyau reproducteur justifiant le nombre élevé des femelles. Cette dernière raison a été également rapportée au Botswana (16).
31La moyenne de la HG pour l’ensemble des caprins trouvée est conforme à ce qui a été signalée précédemment pour cette race au Togo (3). Elle est proche de celles rapportées pour les chèvres de même race au Nigéria (9), au Ghana (14) et au Sénégal (12). Les individus caractérisés dans cette étude appartiennent donc à la race de chèvre naine plus petite que la chèvre Mossi du Burkina Faso dont la hauteur peut atteindre 48,4 ± 4,10 cm (25) et les chèvres de Lubumbashi en République Démocratique du Congo avec 53 cm (21). La HG est le paramètre qui le plus fait référence pour décrire le format des mammifères (18). Toute race animale ayant une hauteur au garrot inférieure à 60 cm est dite race naine (20), dans ce sens, on peut dire que les chèvres caractérisées dans cette étude sont des chèvres naines.
32L’effet de la zone écologique sur la hauteur au garrot confirme l’hypothèse selon laquelle la taille augmente des zones humides et froides vers les zones sèches (19). Aussi, la variabilité entre les populations peut refléter certaines différences génétiques et adaptation spécifique de chaque population locale aux conditions environnementales (15).
33L’influence du sexe sur la HG n’a pas été significative comme l’ont aussi observé Ebegbulem et ses collaborateurs au Nigéria sur la chèvre de la même race et au Congo par Ngona et ses collaborateurs sur les chèvres de Lubumbashi. Toutefois, chez la chèvre Djallonké au Ghana (14), la chèvre Tswana au Botswana (16) et chez la chèvre locale des hautes terres Hararghe en Ethiopie (7) il a été observé que le sexe a un effet sur la hauteur au garrot.
34L’âge a eu une influence sur la hauteur au garrot. Ce résultat est conforme avec celles faites sur la chèvre de même race (9 ; 13 ; 14), sur la chèvre Mossi (25), sur les chèvres d’Ouganda (23) et d’Ethiopie (7).
35Le tour thoracique des chèvres n’est pas influencé par la zone écologique. La moyenne de 61,28 ± 5,40 cm est proche de celles rapportées au Sénégal (12) chez la chèvre Djallonké (54 à 62 cm), mais légèrement supérieure à celles enregistrées au Ghana pour la même race (14).
36L’influence du sexe ressortie dans cette étude sur le TT a été rapportée sur les caprins au Burkina Faso (25). Par contre, en Ouganda, il a été constaté que les mâles avaient toujours les mesures les plus grandes par rapport aux femelles de même âge chez la chèvre naine de l’Afrique de l’Est (23). Hagan et collaborateurs au Ghana en 2012 ont trouvé une moyenne pour les femelles légèrement supérieure à celle des mâles. Cependant Gueye au Sénégal a trouvé une moyenne de tour thoracique des mâles plus grande que celle des femelles lorsque les animaux sont de même âge.
37L’effet de l’âge sur le TT est signalé dans plusieurs études sur les chèvres naines (12 ; 9 ; 14) et sur d’autres races, chèvre Mossi (25); chèvres du Niger (19) et les chèvres en Ouganda (23).
38La moyenne de la longueur scapulo – ischiale (LSI) trouvée dans cette étude est la même que celles rapportées par d’autres auteurs (9 ; 11 ; 14 ; 25). Cette valeur est légèrement supérieure à celles trouvées chez la chèvre de la même race au Nigéria (28). L’analyse a montré que la LSI est influencée par la zone agro écologique, le sexe et l’âge. Hagan et collaborateurs ont signalé au Ghana en 2012, l’influence de la zone d’élevage, du sexe et de l’âge sur la longueur du corps des chèvres naines. Les femelles, dans l’ensemble plus âgées que les mâles, ont été plus longues que ces derniers. Les animaux adultes ont une LSI supérieure à celle des jeunes. Chez la chèvre Mossi au Burkina (25) et la chèvre Djallonké au Sénégal (12) les mêmes observations sont faites. Par contre au Nigéria, aucune différence significative entre les moyennes des LSI aussi bien selon le sexe que l’âge des animaux n’a été établie (9). L’influence de la zone d’élevage peut être expliquée par les différences dans les pratiques de gestion des chèvres, les conditions agro-climatiques et les ressources biophysiques (27).
39La longueur de la queue des caprins étudiés n’a été influencée par aucun facteur. Par contre pour cette même race au Ghana, une influence de l’âge des animaux sur la longueur de la queue a été signalée (15), les caprins les plus âgés ont une longue queue.
40La longueur moyenne des oreilles est voisine de celles trouvées par d’autres auteurs (9 ; 14 ; 26) mais inférieure à celle signalée au Sénégal (12). La zone agro-écologique et l’âge ont significativement influencé la taille des oreilles des caprins. Cette influence peut être expliquée par des métissages réalisés dans le temps par les éleveurs. Les zones dans lesquelles les animaux ont eu des oreilles moins longues (Maritime et Plateaux Ouest) sont des zones très pluvieuses. Les races sahéliennes et les produits du métissage résistent ou survivent faiblement. Certains individus rencontrés dans les autres zones pourraient être des individus issus de croisements lointains. Certains auteurs ont signalé une influence significative de l’âge sur la taille des oreilles (14 ; 19). L’influence du sexe n’est pas ressortie de l’analyse statistique ce qui est conforme aux résultats d’autres auteurs au Nigéria et au Sénégal (9 ; 12).
41La longueur moyenne des cornes des caprins dans les différents élevages enquêtés est inférieure à la longueur trouvée au Burkina Faso chez la race Mossi (25) et au Ghana (14) chez les chèvres Djallonké. L’effet sexe et âge des caprins sont aussi statistiquement significatifs sur la longueur des cornes des animaux. L’effet de l’âge sur la longueur des cornes a été signalé au Ghana (14). Par contre, au Sénégal, l’effet du sexe a été ressorti, les mâles ayant des cornes plus longues que les femelles (12).
42La zone agro écologique, l’âge et le sexe ont influencé significativement le poids des animaux. Les femelles ont été plus lourdes que les mâles et les animaux âgés de plus de deux ans plus lourds que les jeunes de moins de deux ans, ceci est conforme à l’étude faite au Nigeria (9).
43Le nombre limité des mâles adultes dans les élevages peut expliquer la faible corrélation entre le sexe mâle et les différents paramètres étudiés. Cette observation confirme celle faite au Niger (19). L’homogénéité relative de la population caprine du Togo (dans les 6 zones agroécologiques) pour certains caractères mesurés pourrait s’expliquer par les migrations des populations, les échanges ou les vente de géniteurs entre les éleveurs (10).
Conclusion
44Cette étude a confirmé que la chèvre Djallonké est présente dans toutes les zones agroécologiques et domine toutes les autres races de caprins du Togo. Sa taille varie du Sud vers le Nord du pays. Le format des chèvres Djallonké augmente notamment du point de vue hauteur au garrot, tour du thorax et poids du sud vers le nord. Une hauteur au garrot de 60 cm et un tour de thorax de 50 cm contre un poids de 16 Kg est considérée comme un maximum pour un adulte Djallonké des régions sud. Dans les régions du nord la HG, le TT et le poids atteignent respectivement 65cm, 58 cm et 22 kg. Ces valeurs doivent être considérées comme des limites de taille que la chèvre Djallonké adulte ne peut dépasser. L’influence de l’âge sur ces paramètres montre que la chèvre Djallonké continue à croitre après deux ans et cette croissance se stabilise après l’âge de 4 ans.
45Les analyses multivariées utilisées (analyses en Composantes principales et en Facteurs Discriminants) ont montré qu’il y a une très faible structuration des caprins d’une zone agroécologique à une autre. Des analyses moléculaires de ces caprins permettront de mieux expliquer les différences observées sur le plan phénotypique et de voir s’il existe ou non des sous – groupes génétiques.
Remerciement
46Nous remercions le Programme de Productivité Agricole de l’Afrique de l’Ouest du Togo (PPAAO/WAAPP -TOGO) qui a financé la collecte des données de cette étude. Nous présentons toute notre reconnaissance aux éleveurs qui ont accepté de mettre les animaux à notre disposition pour les mensurations et aux Conseillers agricoles de l’Institut de Conseil et d’Appui Technique (ICAT).
Bibliographie
-
Adomefa K. & Bonfoh B., 2003. Etat des Ressources Génétiques Animales au Togo, Rapport National, Lomé, Ministère de l’Agriculture et de l’Elevage, 80 p.
-
Afidegnon D., 1999, Les mangroves et les formations associées du Sud – Est du Togo : Analyse éco-floristique et cartographie par télédétection spatiale. Thèse de doctorat FDS/UB. Lomé Togo 237 p.
-
Amégée Y., 1986, Performance d’engraissement et qualités bouchères de la chèvre Djallonké In. Rev. Elevage Med. Vet. Pays Trop., 39, 78-85.
-
Amégée Y., 1987, Performance de reproduction de la chèvre Djallonké au Togo. In: Goat production in the humid tropics. Proceeding of a workshop at University of Ifê, Ilé-Ifé Nigeria pp 137 -139.
-
Anèlle P., 2007, Genetic characterization of commercial goat populations in South Africa. Animal Breeding and Genetics in the Faculty of Natural and Agricultural Science. University of Pretoria. 62 p.
-
Brunel J. F., 1981, Végétation. In : Atlas du Togo. Ed. Jeune Afr., Paris, pp 16 -17.
-
Dereje T., Berhanu B. & Aynalem H., 2013, Morphological Characterization of Indigenous Hararghe Highland Goat Breed in Their Native Environment, West Hararghe, Ethiopia. In: Am. Eurasian J. Sci. Res., 8, 2, 72-79.
-
DSID, 2012, Productions Animales au Togo, www.countrystat.org
-
Ebegbulem V.N., Ibe S.N., Ozung P.O. & Ubua J.A., 2011, Morphometric trait characteristics of West African Dwarf goats in Abia State, South East Nigeria. Contin. J. Agri. Sci., 5, 2, 1- 6.
-
FAO, 2007, L’état des ressources zoogénétiques pour l’alimentation et l’agriculture dans le monde – en bref. Dafydd Pilling & Barbara Rischkowsy. Rome
-
FAO, 2015, The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture. B.D. Scherf & D. Pilling. FAO Commission on Genetic Resources for Food and Agriculture Assessments. Rome (available at http://www.fao.org/3/a-i4787e/index.htlml).
-
Gueye A., 1997, Moutons et chèvres du Sénégal : caractérisation morpho - biométrique et typage sanguin. Thèse de doctorat, Ecole Inter Etat des Sciences et Médecine Vétérinaires (EISMV), Université Cheik Anta Diop de Dakar, 108 p.
-
Getachew B.F., 2016, Characterization of African Goat Production and Productivites: The Case of Ethiopia: Rev. Appl. Anim. Husb. Rural Develop. 2009, 1, 1-8: www.sasas.co.za/aahrd/ Appl. Anim. Husb. Rural Develop. 2009, 2.
-
Hagan J.K., Apori S.O., Bosompem M., Ankobea G. & Mawuli A., 2012, Morphological Characteristics of Indigenous Goats in the Coastal Savannah and Forest Eco-Zones of Ghana. J. Anim. Sci. Adv., 2, 10, 813-821 ISSN: 2251-7219. Educ. Psychol. Meas., 20, 141-151.
-
Ibnelbachyr M., Boujenane I. & Chikhi A., 2015, Morphometric differentiation of Moroccan indigenous Draa goat based on multivariate analysis Animal Genetic Resources, 57, 81-87. Food and Agriculture Organization of the United Nations, 2015 doi: 10.1017/S2078633615000296.
-
Katongole J.B.D., Sebolai B. & Madimabe M. J., 1996, Morphological characterisation of the Tswana goat. In: S.H.B. Lebbie & E. Kagwini, Small Ruminant Research and Development in Africa. Proceedings of the Third Biennial Conference of the African Small Ruminant Research Network, UICC, Kampala, Uganda 5-9 December 1994 pp 43- 46.
-
Kokou K., 1998, Les mosaïques forestières au sud du Togo : biodiversité, dynamique et activités humaines. Thèse de doctorat, Université de Mompellier II, 140 p.
-
Manallah I. & Dekhili M., 2011, Caractérisation morphologique des caprins dans la zone des hautes plaines de Sétif. Agric., 2, 7 -13
-
Mani M., Marichatou H., Issa M., Chaibou I., Sow A., Chaibou M. & Sawadogo J.G., 2014, Caractéristiques phénotypiques de la chèvre du sahel au Niger par analyse des indices de primarité et des paramètres qualitatifs. In: Anim. Genetic Res., 54, 11-19, Food and Agriculture Organization of the United Nations, 2014 doi:10.1017/S2078633613000507 pp. 11- 20.
-
Mason I.L., 1984, Goat Evolution of Domestical Animals. Ed. Longman, London, 86-93.
-
Ngona I.A., Beduin J.M., Khang’Maté A.B.F. & Hanzen C., 2012, Etude descriptive des caractéristiques morphométriques et génitales de la chèvre de Lubumbashi en République Démocratique du Congo. In : Rev. Elev. Med. Vet. Pays Trop., 65, 3-4, 75-79.
-
Sangaré M., 2005, Synthèse des résultats acquis sur l’élevage des petits ruminants dans les écosystèmes de production d’Afrique de l’Ouest. URPAN CIRDES Burkina Faso. 2005. Pp. 90-134.
-
Semakula J., Mutetikka D., Kugonza R. D. & Mpairwe D., 2010, Variability in Body Morphometric Measurements and their Application in Predicting Live Body Weight of Mubende and Small East African Goat Breeds in Uganda In Middle-East. J. Sci. Res., 5, 2, 98-105.
-
Sylvain J-P., Collart J., Aregba A., Godonou S., 1986, Notice explicative de la carte géologique au 500 000è du Togo. Mem n° 6, Dir. Gén. Min/Bur. Nat. Rech Min Lomé.
-
Traoré A., Tamboura H.H., Kaboré A., Yaméogo N., Bayala B. & Zare I., 2006, Caractérisation morphologique des petits ruminants (ovins et caprins) de race locale « Mossi » au Burkina. Anim. Genet. Ressour. Inf., 39, 39-50.
-
Traoré A., Tamboura H.H., Kaboré A., Royo L.J., Fernandez I., Alvarez I., Sangaré M., Bouchel D., Poivey J.P., François D., Sawadogo L. & Goyache F., 2008, Multivariate analyses on morphological traits of goats in Burkina Faso. Arch. Tierz., Dummerstorf, 51, 6, 588-600.
-
Yadav D.K., Jain A., Kulkarni V.S., Govindaiah M.G., Aswathnarayan T. & Sadana D.K. 2013. Classification of four ovine breeds of southern peninsular zone of India, morphometric study using classical discriminant function analysis. Springer Plus, 2, 29.
-
Yakubu A., Salako A.E., Imumorin I.G., Ige A.O. & Akinyemi M.O., 2010, Discriminant analysis of morphometric differentiation in the West African Dwarf and Red Sokoto goats. South Afr. J. Anim. Sci., 40, 4, 381- 387.
Om dit artikel te citeren:
Over : A. Y. Djagba
Togolais, PhD, Institut Togolais de Recherche Agronomique, Chef de Programme National Ovins Caprins de Kolokopé, Anié, Togo.
Email: atouga16@yahoo.fr
Over : B. Bonfoh
Togolais, PhD, Directeur Général de l’Institut Togolais de Recherche Agronomique, Chargé de Recherche au CAMES, Lomé Togo.
Over : G.-K. Dayo
Togolais, PhD, Maitre de Recherches CAMES (Génétique Animale). Centre International de Recherche-Développement sur l’Élevage en zone Subhumide. Chef de l'Unité sur les Maladies à Vecteurs et Biodiversité (UMaVeB, ex URBIO). Coordonnateur Régional du Projet de Valorisation des ressources génétiques animales et aquacoles locales dans l'espace UEMOA, Bobo-Dioulasso, Burkina Faso.
Over : K. Aklikokou
Togolais, PhD, Professeur titulaire, Université de Lomé, Faculté Des Sciences, Département de Physiologie/Toxicologie, Lomé, Togo.
Over : H. Bassowa
Togolais, Ingénieur Agronome Zootechnicien, Régisseur au Programme National Ovins Caprins de Kolokopé, Anié Togo.
