
Uppermost Famennian bryozoans from Ratingen (Velbert Anticline, Rhenish Massif/Germany) – Taxonomy, facies dependencies and palaeobiogeographic implications
Abstract
Three bryozoan species are described from the uppermost part of the Velbert Formation (uppermost Famennian, “Strunian”) from the classical section Ratingen-Cromford (northwestern flank of Velbert Anticline, western Rhenish Massif, Germany). The described fauna includes one new genus Ratingella gen. n., one new species Crustopora fistulosa sp. n., and one species in open nomenclature, Nikiforovella sp. The hitherto known bryozoans from the “Strunian” of the Velbert Anticline constitute the most important bryozoan fauna from the Famennian ‘Condroz Shelf’, bordering on the southwesterncoast of Laurussia between southwestern England and Pomerania (northern Poland). Coinciding, it is the most diverse European bryozoan fauna from that time slice. Facies differences along the northwestern flank of the Velbert Anticline between a clear-water ramp segment in the southwest and a marl ramp segment in the northeast are responsible for a completely different faunal association. Encrusting taxa diminish towards the north, while erect growing, branched taxa predominate and the general diversity increases. The studied fauna points to connections with the Asiatic part of Russia, Kazakhstan, and China and implies a general Palaeotethyan character for latest Famennian bryozoans in Eurasia.
1. Introduction
1Famennian bryozoans are widely distributed on Palaeotethyan platforms and adjacent epeiric seas. However, taxonomic knowledge is strongly biased. Besides a Middle Famennian faunula from Iran (Ernst & Mohammadi, 2009), a Late Devonian fauna from Afghanistan (Gorjunova, 2006) and a latest Famennian faunula from the Canning Basin (NW Australia, Ross, 1961), knowledge is mostly based on faunas from the GUS states, Mongolia and northern China (see compilation in Tolokonnikova & Ernst, 2010). Only few publications dealt with Famennian bryozoans from central and western Europe, which, due to unfavourable siliciclastic facies in the preceding time slices, are restricted to the latest Famennian (“Strunian”). No Famennian faunas at all are known from the siliciclastic strata of the Iberian Peninsula or the deeper-water limestones of southern France (Pyrenees, Montagne Noire).
2All the latest Famennian (“Strunian”) western and central European bryozoans have been described from the mixed carbonate-siliciclastic deposits of the “Condroz shelf”, bordering on the southwestern coast of Laurussia (Paproth et al., 1986). Besides Whidborne (1896, 1898), who mentioned four species from South England, all known bryozoans are from the Velbert anticline, westernmost Rhenish Mountains (Germany), which, at present, is the hotspot of our knowledge. Weber & Wise Jackson (2006) gave an overview of these bryozoans, recorded in the classical monograph of Nekhoroshev (1932) and in the faunal list of Michels (1986). Altogether, they listed 20 bryozoan species from the "Etroeungt", i.e. from the uppermost Devonian. However, most of these species are inadequately described and illustrated, and many are in open nomenclature. Their identification is mainly uncertain. Recently, Ernst & Herbig (2010) and Tolokonnikova et al. (2014) started to explore the fauna from the Velbert Anticline. Herein we add the description of three species, including a new genus and two new species, from the classical profile Ratingen-Cromford, also called “Blauer See” at the westernmost tip of the Velbert anticline (Fig. 1). The unexpected increase in diversity from the spatial relatively short outcrop belt along the northwestern flank of the Velbert Anticline provokes a discussion on facies dependency and palaeobiogeographic relations of the “Strunian” bryozoans from that region.
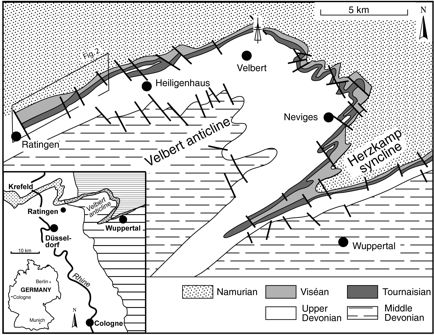
3Figure 1. Geological sketch of the Velbert Anticline. Known uppermost Famennian (“Strunian”) bryozoan localities are situated along the northwestern flank between Ratingen (described herein) and Heiligenhaus, and north of Velbert (research well Velbert-4). Inset: Geological overview of the anticline at the northwestern margin of the Rhenish Mountains showing the narrow “Strunian”–Viséan outcrop belt (stippled in grey) between widely outcropping Devonian in the south (wide horizontal ruling) and Namurian (narrow horizontal ruling) in the north. The strata continue in subcrop below Tertiary-Quaternary sediments of the Niederrhein Embayment (white, subcrop of “Strunian-Viséan stippled in white) towards the Ardennes (modified from Herbig & Mamet, 2006).
2. Geological and stratigraphical setting
4The Velbert Anticline is the westernmost fold structure of the Rhenish Mountains which exposes Middle Devonian to Mississippian strata east of the Rhine River. The northeast plunging, internally strongly tectonized anticline crops out in the region between Ratingen (NE of Düsseldorf) and Wuppertal over a length of about 15 km along strike and 10 km across strike (Fig. 1). Its wide Middle Devonian core is surrounded by Frasnian reefal and very fine-grained peri-reefal limestones (“Massenkalk”, “Flinzkalk”), which grade into shales, siltstones and sandstones of the Late Devonian and finally into a cyclic, mixed siliciclastic-carbonate succession of the latest Famennian (“Strunian”) age. A narrow outcrop belt of predominantly calcareous Tournaisian and Viséan strata follows, succeeded by widely outcropping Namurian siliciclastics.
5The bryozoans described herein have been derived from the uppermost (“Strunian”) part of the Velbert Formation (Paeckelmann, 1913). This formation, with an estimated thickness of 500–1000 m, and in most parts poorly exposed and intensively folded, faulted, and foliated (Bärtling & Paeckelmann, 1928), spans most of the Famennian. The oldest, impure limestone beds yielded Middle Famennian conodonts (Early marginifera Zone: Ribbert & Lange, 1993; Late marginifera-Early trachytera Zone: Amler et al. 1994). From that time onward, and into the Tournaisian, the northwestern flank of the Velbert anticline belonged to the shallow Northwest European Condroz shelf (Franke et al., 1975, p. 316). The Velbert Formation at the northeastern nose and further towards the southern flank of the anticline graded into the hemipelagic “Cypridina shale facies” of the Rhenohercynian Culm basin, which include a few nodular limestone intercalations. These limestone beds are fully developed further southeastward in the adjoining narrow Herzkamp syncline (Paeckelmann, 1922; see reviews in Amler & Herbig, 2006, Clausen & Korn, 2008). The southwestern prolongation of the Condroz Shelf is observed in subcrop of the Niederrhein Embayment (Ribbert, 1998) and across the Ardennes (Aachen region/westernmost Germany, Belgium) into the Avesnois (northern France) and finally into the Pilton beds of southwest England (Paul, 1937; Conil, 1964; Mamet et al., 1965, 1970; see Amler, 1995 for review). Towards the northeast and east relicts are known from deep wells from the Rügen Island (Hoffmann et al., 1975; see review in Zagora, 2008) and Pomerania (northern Poland, Matyja, 1988, 1993).
6The latest Famennian “Strunian transgression” (TR-Zyklus IIf; Johnson et al., 1985, 1986) caused the diachronous onset of cyclic, mixed siliciclastic carbonate succession all along the Condroz shelf. The term “Strunian” is used as a chronostratigraphical unit for that uppermost Famennian time slice in Belgium and in many other parts of the world in time-equivalent shallow-water deposits (Streel et al., 2006). However, in spite of a proposal, that its base should coincide with the base of the Late expansa Conodont Zone and its top with the Devonian-Carboniferous boundary, it is not a formally accepted chronostratigraphic unit (Streel et al., 2006). In the historical type area in the Avesnois (N France) and in southern Belgium, the calcareous Etroeungt Formation is the lithostratigraphic equivalent and, therefore, in older literature also addressed as “Etroeungt” time interval. However, due to the diachronous onset of carbonate sedimentation, the “Strunian” base is in the upper part of the underlying, predominantly sandy Evieux Formation further northeast (Bultynck & Dejonghe, 2001: fig. 7, see also Amler & Herbig, 2006). In the Velbert Anticline, first Kayser (1882) noted the equivalence of beds, now placed into to the Velbert Formation, with the “Etroeungt” of the Avesnois, a fact which was stressed by Paul (1937) and extended to include the Pilton beds from Southwest England. Later, Paul (1939) made the most thorough contribution to the upper Famennian of the Velbert anticline. In ascending order, he discerned Velbert beds, Angertal beds, and Etroeungt beds s.s. However, already Conil & Paproth (1968, p. 57) and Paproth et al. (1983, p. 213) discarded the usage of Angertal beds and Etroeungt beds as regionally usable lithostratigraphic units because of their very local recognition in parts of the northwestern flank of the Velbert Anticline and general rapid facies variations. Therefore, all units were placed in the Velbert Formation. This was also adapted after renewed mapping in the review of Amler et al. (1994), and finally in the most recent compilation of the Uppermost Famennian and Mississippian of the Velbert anticline by Amler & Herbig (2006).
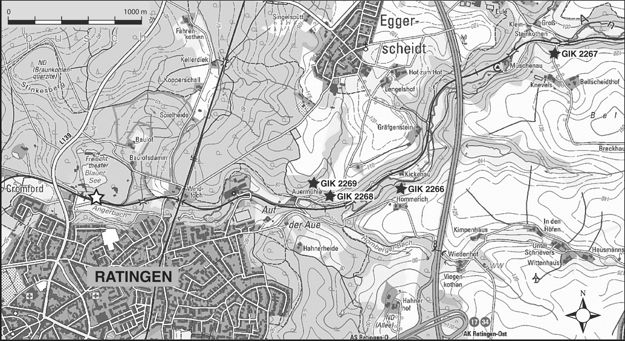
7Figure 2. Location of the section Ratingen-Cromford (white asterisk) and bryozoan localities of Ernst & Herbig (2010) (black asterisks; repository numbers of the Institute of Geology and Mineralogy, University of Cologne). See Table 4 for faunal contents.
8The studied bryozoans, at first observed by chance with a hand-lens, were collected during field work in July 2013 from bed 10 of the classical section Ratingen-Cromford (Fig. 2) at the northern margin of the town of Ratingen (topographic map 1 : 25 000, sheet 4607 Heiligenhaus/Kettwig, N 51° 18' 25.44", E 6° 51' 35.4"). The bryozoan-bearing bed is located at the paved driveway which runs parallel along the northern side of the railway track, some ten meters west of a small quarried slope side exposing beds 2–8 (Fig. 3). No further bryozoans were observed during field work in these beds, which remained unsampled. It has to be mentioned, however, that also some bryozoans described by Nekhoroshev (1932) had been derived from that stratigraphic interval (beds 2-14: “Fenestella cf. longa”, “Acanthoclema? cavernosa”, “Monotrypella? cromfordensis” – the latter herein assigned to the new genus Ratingella). The section was most thoroughly measured by Paeckelmann and Schindewolf in 1930. Their unpublished records including extended fossil lists and the still used bed numbers were the base of the description by Paul (1939, p. 652-658). Later descriptions of the Strunian part of the section are from Conil (1964), who mentioned stromatoporoids and rugose corals from beds 4-8, and Conil & Paproth (1968). The latter mentioned in the four lowermost mixed siliciclastic carbonate cycles (beds 2–14b, but note that we recognized only three cycles – see Fig. 3) the stratigraphically diagnostic foraminifers Quasiendothyra communis communis (Rauzer-Chernoussova), Qu. communis regularis Lipina and rare “Quasiendothyra with fibrous wall layer”, i.e. the first representatives of the Quasiendothyra kobeitusana group. The first Qu. kobeitusana kobeitusana (Rauzer-Chernoussova) was recorded in bed 16. Paproth & Streel (1970) recorded a palynoflora from bed 3, which was dated as questionable “Pli subzone” of the pusillites–lepidophytus (PL) Florizone. From bed 19 they listed an assemblage from the Pls1 Subzone of the PL Florizone. Today, both assemblages belong to the Retisporalepidophyta–Knoxisporites literatus (LL) Biozone according to the revised Famennian miospore zonation of Maziane et al. (1999), which also includes the underlying lepidophyta–verrucosa (LV) Biozone established in Paproth et al. (1983). The stratigraphic summary of the known foraminifers and spores from the interval embracing the bryozoan-bearing bed 10 indicates its position close to the base of the Strunian stage (Fig. 3) within the Middle or Late expansa Conodont Zone (Maziane et al., 1999, fig. 5; see also Higgs et al., 2013). In lithostratigraphic terms, the discussed part of the section at Ratingen-Cromford has to be correlated with the basal Comblain-au-Pont Formation from eastern Belgium (= Etroeungt Formation according to Amler & Herbig 2006).
9A later compilation of the section including a complete fossil list was given in Paproth & Streel (1982) and somewhat supplemented and modified in Amler et al. (1994), who figured some productellids from the brachiopod-rich bed 13 (see Paul, 1939, p. 654). Surprisingly, the bryozoan-bearing bed 10, a 4 m thick unit consisting of thick-bedded, dark bluish-grey, in part nodular to flasered limestones with shale interbeds, was characterized by those authors to be almost devoid of fossils except for crinoid ossicles. The bryozoan faunula has been derived from strongly sandy, micritic crinoid-bryozoan packstone; all bioclasts are fragmented. The microfacies represents the reworked debris of crinoid-bryozoan meadows flourishing between storm wave base and fair weather base. It is correlated with Facies I of Herbig & Mamet (2006).
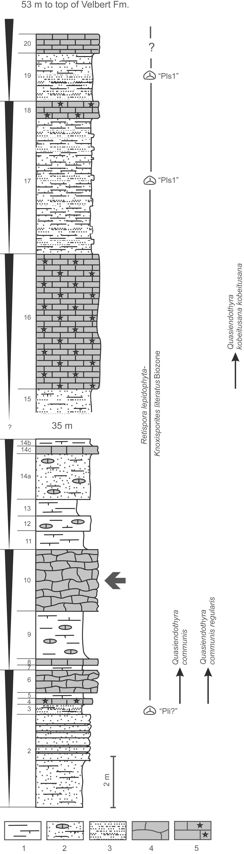
10Figure 3. The uppermost Famennian (“Strunian”) part of the Velbert Formation at Ratingen-Cromford based on Paeckelmann and Schindewolf in Paul (1939), amendments in Conil & Paproth (1968), and biostratigraphic data discussed in the text. Bryozoan-bearing bed 10 and smaller transgressive cycles with diminishing siliciclastic influx and increasing carbonate content indicated. 1 – Siltstone, in part calcareous to marly; 2 – sandy siltstone, in part calcareous and/or with carbonate nodules; 3 – fine-grained, mica-bearing sandstone; 4 – flasered and nodular limestone; 5 – limestone, indifferent / crinoidal limestone.
11From the collected material 10 thin sections were prepared. The bryozoan fauna was studied with a transmitted light binocular microscope. Studied material is housed at the Senckenberg Museum (Frankfurt am Main, Germany), numbers SMF23.099- SMF23.109.
3. Systematic palaeontology
12Phylum Bryozoa Ehrenberg, 1831
13Class Stenolaemata Borg, 1926
14Order Trepostomata Ulrich, 1882
15Suborder Amplexoporina Astrova, 1965
16Family Atactotoechidae Duncan, 1939
17Genus Ratingella gen. n.
18Etymology. The genus name refers to the type locality Ratingen, western Germany.
19Type species. Monotrypella (?) cromfordensis Nekhoroshev, 1932, designated in the present publication. Ratingen, Germany; upper Famennian, Upper Devonian.
20Diagnosis. Encrusting colonies; autozooecia prismatic, growing from thick epitheca; basal diaphragms common to rare, thick, originating in wall cortex; exilazooecia few to absent; acanthostyles absent; autozooecial walls laminated, integrated with distinct zooecial boundaries in endozone; merged, with distinct reversal U-shaped lamination without distinct zooecial boundaries in exozone, containing laminated cortex; maculae consisting of macrozooecia present.
21Comparison. Ratingella gen. n. differs from Leptotrypa Ulrich, 1883 in absence of acanthostyles. It differs from Anomalotoechus Duncan, 1939 in absence of acanthostyles and development of thin encrusting colonies instead of massive and branched ones in the latter genus.
22Occurrence. The new genus Ratingella contains four species: R. cromfordensis (Nekhoroshev, 1932) from the Upper Devonian (Famennian) of Germany, R. tubulosa (Nekhoroshev, 1977) from the Upper Devonian (Famennian) of Kazakhstan, R. texta (Troizkaya, 1963) from the Upper Devonian (Frasnian-Famennian) of Kazakhstan, and R. cavernosa (Fritz, 1944) from the Upper Devonian (Famennian) of New Mexico, USA.
23Ratingella cromfordensis (Nekhoroshev, 1932) (Pl. 1 A–F; Table 1)
241932 Monotrypella (?) cromfordensis Nekhoroshev: 65-66, pl. 4, figs 4, 4a-c.
25Material. SMF23.101-SMF23.102.
26Description. Encrusting colony, 0.67-0.97 mm in thickness. Autozooecia growing from 0.02-0.04 mm thick epitheca, bending sharply in exozone. Autozooecial apertures polygonal. Autozooecial diaphragms thick, usually 1-3 in each autozooecium present, originating in wall cortex. Exilazooecia and acanthostyles absent. Autozooecial walls laminated, with distinct zooecial boundaries, 0.02-0.04 mm thick in endozones; thickened, showing distinct reversal U-shaped lamination without distinct zooecial boundaries, containing laminated cortex, 0.04-0.10 mm thick in exozones. Maculae distinct, consisting of macrozooecia, 1.2-1.4 mm in diameter.
27Comparison. Ratingella cromfordensis (Nekhoroshev, 1932) differs from R. tubulosa (Nekhoroshev, 1977) from the Famennian of Kazakhstan, in larger autozooecial apertures (0.20-0.29 mm vs. 0.17-0.25 mm in R. tubulosa), and in larger macrozooecia (0.29-0.41 mm vs. 0.26-0.30 mm in R. tubulosa). Ratingella cromfordensis differs from R. texta (Troizkaya, 1963) from the Upper Devonian (Frasnian-Famennian) of Kazakhstan in presence of macrozooecia. Furthermore, Ratingella cromfordensis shows also wider range of aperture width in non-macular area than the latter species (0.20-0.29 mm vs. 0.26-0.29 mm in R. texta). No measurements are available in the original description of the species Ratingella cavernosa (Fritz, 1944) from the Upper Devonian (Famennian) of New Mexico, USA. This species seems to lack macrozooecia and to possess larger autozooecial apertures than R. cromfordensis. Measurements of the aperture width from the figure 6 of the plate 12 (Fritz, 1944) resulted in values of 0.27-0.33 mm for Ratingella cavernosa, whereas aperture width in R. cromfordensis ranges within 0.20-0.29 mm.
28Occurrence. Ratingen-Cromford, Germany; bed 10 at the profile Ratingen-Cromford, uppermost Velbert Formation (“Strunian”, uppermost Famennian, Upper Devonian).
29Table 1. Descriptive statistics for Ratingella cromfordensis (Nekhoroshev, 1932). Abbreviations: N = number of measurements, X = mean, SD = sample standard deviation, CV = coefficient of variation, MIN = minimal value, MAX = maximal value.
|
N |
X |
SD |
CV |
MIN |
MAX |
|
|
Autozooecial aperture width, mm |
20 |
0.22 |
0.028 |
12.71 |
0.20 |
0.29 |
|
Autozooecial aperture spacing, mm |
20 |
0.28 |
0.033 |
11.98 |
0.16 |
0.31 |
|
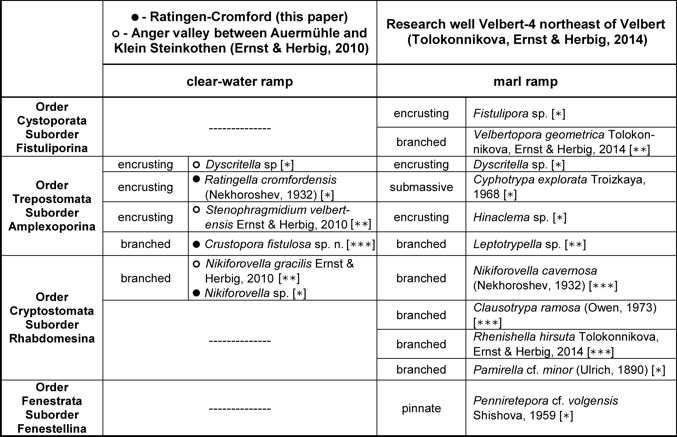
Macrozooecia width, mm |
7 |
0.34 |
0.040 |
11.64 |
0.29 |
0.41 |
|
Macrozooecia spacing, mm |
5 |
0.44 |
0.029 |
6.49 |
0.41 |
0.49 |
|
Exozonal wall thickness, mm |
10 |
0.07 |
0.021 |
30.61 |
0.04 |
0.10 |
30Family Crustoporidae Dunaeva & Morozova, 1967
31Genus Crustopora Dunaeva, 1964
32Type species. Crustopora tuberculata Dunaeva, 1964, by original designation. Lower Carboniferous (Namurian); Ukraine.
33Diagnosis. Encrusting and branched colonies; autozooecial apertures rounded-polygonal; autozooecial diaphragms thin; exilazooecia abundant, small, often gathered in clusters, possessing thin diaphragms; acanthostyles small, common to abundant; maculae consisting of macrozooecia present.
34Comparison. Crustopora Dunaeva, 1964 differs from Hinaclema Sakagami & Sugimura, 1987 in having small acanthostyles. Toulopora Nakrem et al., 2009 differs in having acanthostyles of two sizes: sparsely distributed large ones, and a row of regularly distributed small ones inflecting each zooecial aperture. Crustopora differs from Tabuliporella Nikiforova, 1933 in lacking hemiphragms.
35Occurrence. Several species are known from the Middle Devonian (Givetian) to the Lower Carboniferous (Serpukhovian) of Kazakhstan, Siberia, Altai and Ukraine. The species described as Crustopora dubia Nekhoroshev, 1977 from the Lower Devonian (Emsian) of Kazakhstan has no acanthostyles and is excluded from this genus.
36Crustopora fistulosa sp. n. (Pl. 2 A–I; Table 2)
37Etymology. The species name refers to the presence of abundant small exilazooecia (derived from Latin "fistulosa" = porous, cavernous).
38Holotype. SMF23.106.
39Paratypes. SMF23.099, SMF23.103-SMF23.105, SMF23.107-SMF23.109.
40Locus typicus. Ratingen-Cromford, Germany.
41Stratum typicum. Bed 10 of the Ratingen-Cromford section, uppermost Velbert Formation (“Strunian”, uppermost Famennian, Upper Devonian).
42Diagnosis. Branched colonies with sharply separated exozones; autozooecial apertures polygonal; autozooecial diaphragms thick, common in the transition between endozone and exozone, originating from wall cortex; exilazooecia abundant, small, usually separating autozooecial apertures, locally forming clusters, containing thin diaphragms; acanthostyles small, common; autozooecial walls laminated, merged without visible zooecial boundaries, containing indistinct laminated cortex; maculae consisting of macrozooecia and clusters of exilazooecia.
43Description. Branched colonies. Branches 1.43-2.10 mm in diameter, with 0.23-0.59 mm wide endozones and 0.78-1.20 mm wide exozones. Exozones distinctly separated from endozones. Autozooecia growing for a long distance in endozone, diverging at angles of 17-21° (20.2° at average) in the outer endozone and abruptly bending at angles of 73-87° (82° at average) in exozone, having polygonal transverse section in endozone. Autozooecial apertures rounded-polygonal. Autozooecial diaphragms common, thick, deflected proximally, concentrated in the transition between endozone and exozone, originating from the wall cortex. Exilazooecia originating at the base of exozone, small, abundant, 6-11 surrounding each autozooecial aperture, forming clusters between apertures, generally separating them in 1-2 rows, locally not completely separating autozooecia, containing up to 5 thin diaphragms. Acanthostyles small, common, having distinct hyaline cores and laminated sheaths, distributed evenly on the colony surface. Autozooecial walls granular, 0.005-0.010 mm thick in endozone; finely laminated, merged without visible zooecial boundaries, containing indistinct laminated cortex, 0.035-0.055 mm thick in exozone. Maculae consisting of macrozooecia and clusters of exilazooecia, 0.94-1.17 mm in diameter.
44Comparison. Crustopora fistulosa sp. n. differs from C. tuberculata Dunaeva, 1964 from the Serpukhovian of Ukraine in smaller apertures (0.09-0.20 mm vs. 0.16-0.25 mm in C. tuberculata). Crustopora fistulosa differs from C. lubrica (Trizna, 1958) from the Tournaisian of Kuznets Basin in smaller apertures (0.09-0.20 mm vs. 0.16-0.20 mm in C. lubrica).
45Table 2. Descriptive statistics for Crustopora fistulosa sp. n. Abbreviations as for Table 1.
|
N |
X |
SD |
CV |
MIN |
MAX |
|
|
Branch diameter, mm |
6 |
1.81 |
0.277 |
15.32 |
1.43 |
2.10 |
|
Exozone width, mm |
6 |
0.42 |
0.121 |
29.23 |
0.23 |
0.59 |
|
Endozone width, mm |
6 |
0.98 |
0.146 |
14.97 |
0.78 |
1.20 |
|
Autozooecial aperture width, mm |
25 |
0.14 |
0.032 |
23.01 |
0.09 |
0.20 |
|
Autozooecial aperture spacing, mm |
25 |
0.25 |
0.049 |
19.70 |
0.17 |
0.35 |
|
Macrozooecia width, mm |
8 |
0.25 |
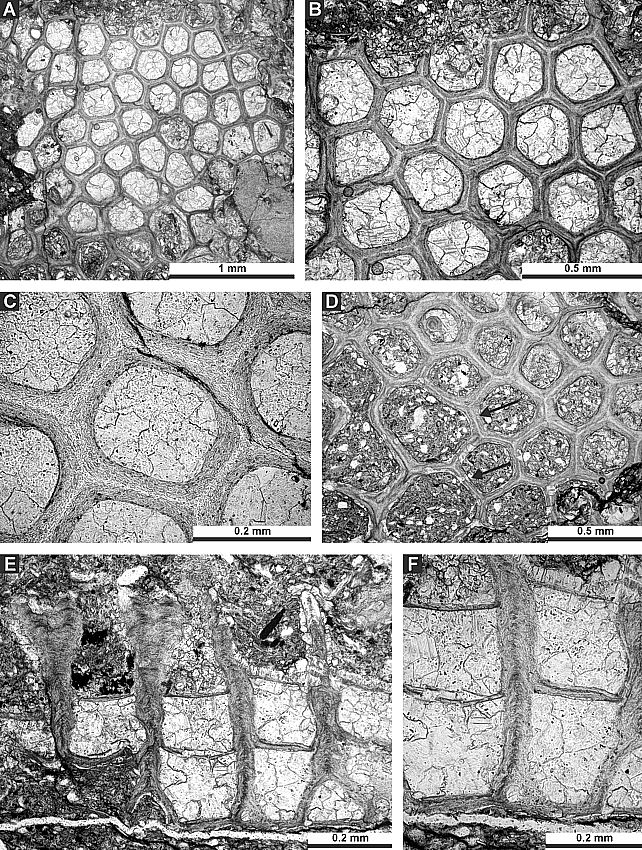
0.024 |
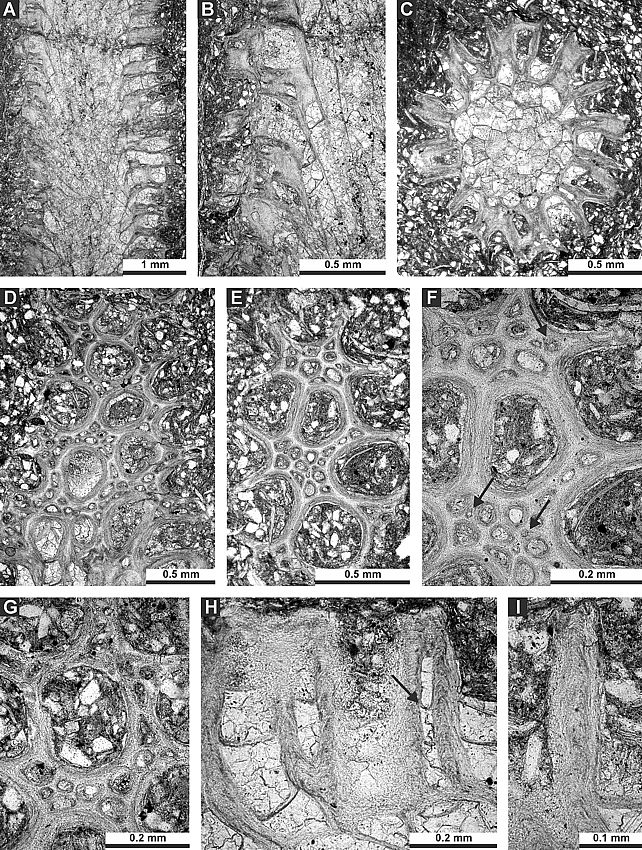
9.71 |
0.22 |
0.30 |
|
Macrozooecia spacing, mm |
7 |
0.37 |
0.056 |
14.91 |
0.32 |
0.48 |
|
Acanthostyle diameter, mm |
23 |
0.030 |
0.007 |
22.65 |
0.020 |
0.045 |
|
Exilazooecia width, mm |
25 |
0.04 |
0.013 |
35.99 |
0.02 |
0.08 |
|
Exilazooecia per aperture |
24 |
8.1 |
1.393 |
17.14 |
6.0 |
11.0 |
|
Exozonal wall thickness, mm |
20 |
0.053 |
0.013 |
24.69 |
0.035 |
0.075 |
46Order Cryptostomata Vine, 1884
47Suborder Rhabdomesina Astrova & Morozova, 1956
48Family Nikiforovellidae Gorjunova, 1975
49Genus Nikiforovella Nekhoroshev, 1948
50Type species. Nikiforovella alternata Nekhoroshev, 1948, by original designation. Lower Carboniferous; Altai, Russia.
51Diagnosis. Branched colonies. Autozooecia diverging at low angles from distinct median axis. Hemisepta absent, diaphragms rare. Autozooecial walls laminated, with dark zooecial boundaries. Metazooecia few between longitudinally successive autozooecial apertures; acanthostyles common to abundant. Longitudinal ridges absent.
52Comparison. Nikiforovella Nekhoroshev, 1948 is similar to Streblotrypella Nikiforova, 1948, but differs from it mainly in shape of autozooecia, which bend at higher angles in exozone, and absence of longitudinal ridges. Moreover, styles can be absent in Streblotrypella.
53Occurrence. Devonian to Permian; worldwide.
54Nikiforovella sp. (Pl. 3 A-E; Table 3)
55Material. Single specimen SMF23.100.
56Description. Branched colony exposing an incomplete longitudinal section with 0.58 mm wide exozone. Autozooecia growing in spiral pattern from the median axis, abruptly bending in exozone. Autozooecial apertures oval to circular, arranged in regular diagonal rows. Autozooecial diaphragms absent. Metazooecia originating at the base of exozone, 1-2 arranged in clusters between longitudinally successive autozooecial apertures. Acanthostyles large, 1-3 arranged between longitudinally successive autozooecial apertures, having distinct hyaline cores and laminated sheaths. Autozooecial walls granular, 0.010-0.015 mm thick in endozone; finely laminated, without visible zooecial boundaries, 0.023-0.055 mm thick in exozone.
57Comparison. Nikiforovella sp. differs from N. amazarica Nekhoroshev, 1960 from the Famennian of the Amur region of Russia in having larger apertures (aperture width 0.12-0.17 mm vs. 0.07-0.08 mm in N. amazarica). Nikiforovella sp. is similar to N. kasakhstanica Nekhoroshev, 1948 from the upper Tournaisian (Mississippian) of central Kazakhstan, but differs from it in having 1-2 metazooecia instead of 6 metazooecia between autozooecial apertures the latter species.
58Occurrence. Ratingen-Cromford, Germany; bed 10 of the Ratingen-Cromford section, uppermost Velbert Formation (“Strunian”, uppermost Famennian, Upper Devonian).
59Table 3. Descriptive statistics for Nikiforovella sp. Abbreviations as for Table 1.
|
N |
X |
SD |
CV |
MIN |
MAX |
|
|
Autozooecial aperture width, mm |
30 |
0.14 |
0.013 |
9.58 |
0.12 |
0.17 |
|
Autozooecial aperture spacing, mm |
25 |
0.27 |
0.023 |
8.58 |
0.23 |
0.32 |
|
Autozooecial aperture spacing diagonally, mm |
25 |
0.17 |
0.012 |
7.27 |
0.14 |
0.20 |
|
Metazooecia width, mm |
20 |
0.043 |
0.011 |
25.81 |
0.025 |
0.075 |
|
Acanthostyle diameter, mm |
20 |
0.034 |
0.006 |
18.44 |
0.025 |
0.050 |
|
Number of metazooecia between apertures |
20 |
1.6 |
0.503 |
31.41 |
1.0 |
2.0 |
|
Number of acanthostyles between apertures |
20 |
1.8 |
0.894 |
49.69 |
1.0 |
3.0 |
|
Exozonal wall thickness, mm |
7 |
0.038 |
0.013 |
34.15 |
0.023 |
0.055 |
4. Discussion and conclusions
4.1. Diversity and facies relations of the bryozoans from the upper Velbert Formation
60We rely on data assembled by Ernst & Herbig (2010), Tolokonnikova et al. (2014) and herein. Classical data from Nekhoroshev (1932) and faunal lists from Michels (1986) are omitted due to taxonomic uncertainties (see also compilation in Weber & Wyse Jackson, 2006). Altogether 16 taxonomically modern treated bryozoan taxa are known from the uppermost Velbert Formation along the northwestern flank of the Velbert anticline, two of them with certain restrictions in specific affiliation (‘cf.’), five others in open nomenclature (Table 4). All have been derived from different horizons of the Strunian. This means that at present the Velbert Anticline is the most important Strunian bryozoan locality in Europe. In general, bryozoans are common and contribute up to 10 % of the total invertebrate fauna (Michels, 1986, from fine-grained, in part calcareous siliciclastics at the northern tip of the anticline near Velbert), but in most cases the zooecia are too strongly fragmented for precise determinations.
61Table 4. Taxonomic composition and growth types of uppermost Famennian (“Strunian”) bryozoans from the northwestern flank of the Velbert Anticline demonstrating the differences between clear-water ramp segment (left column) and marl ramp segment (right column). [] – single specimen; [] – rare to moderately common; [] – common.
62Strong differences are observed between the fauna from the southwestern segment of the Velbert Anticline between Ratingen-Cromford and Klein Steinkothen, described by Ernst & Herbig (2010) and herein, and from the Velbert-4 well, located northeast of Velbert at the northern tip of the syncline (Fig. 1). Only two genera (Dyscritella, Nikiforovella) and not a single common species co-occur. The diversity, also on order/suborder level, is higher in the north, with a clear dominance of rhabdomesines, as already noted by Michels (1986: predominance of “Rhabdomeson”). In the north, encrusting and submassive growing taxa contribute about 35 % to the fauna, although they are rare in absolute numbers. Branched and pinnate growth forms predominate and are common. In the southwestern segment, encrusting taxa (on generic level, without separating Nikiforovella gracilis Ernst & Herbig, 2010 and Nikiforovella sp.) make up 60 % of the total fauna. They might be somewhat more common in absolute numbers than in the north, but still branched taxa, especially Crustopora fistulosa sp. n. are frequent.
63The observed differences are due to a southwest-northeast facies gradient along the northwestern flank of the Velbert Anticline. They corroborate the observations of all earlier authors (e.g. Paul, 1939; Böger, 1962; Conil & Paproth, 1968) and, especially, the facies model of Herbig & Mamet (2006). This model points to a southeast dipping carbonate ramp consisting of a high-energy clear-water ramp segment in the southwest extending from the westernmost outcrops in Ratingen-Cromford to the western edge of Heiligenhaus. It is characterized by packstone and grainstone dominated by Girvanella–Paracelligelloides–Quasiendothyra. This segment of the ramp, inhabited by the bryozoans, is described in Ernst & Herbig (2010) and herein. Obviously, higher turbulence and availability of bioclasts favoured the growth of encrusting taxa. Branching, erect growing taxa obviously were a component of bryozoan-crinoid meadows flourishing below fair-weather wave base (cf. Herbig & Mamet, 2006).
64Towards the northeast, an intermediate “Menselina ramp” followed, which was replaced by a muddy ramp consisting of packstone and wackestone, characterized by Issinella–Palaeoberesella–tournayellids. These segments are seen in the surroundings of Heiligenhaus. From both segments, no determinable bryozoans are recorded up to now.
65Finally, Herbig et al. (2013, 2014) recognized further transition into an extremely muddy ramp at the northern tip of the anticline (Velbert-4 core). It is composed predominantly of marly bioclastic mudstone and wackestone, and inhabitated by a filter feeding community (crinoid–bryozoan-brachiopod association) without calcareous algae and foraminifera. This segment was termed marl ramp in Tolokonnikova et al. (2014) and yielded the diverse bryozoans described therein. The fauna inhabited low-energetic environments, with relative high influx of fine-grained silicilastics. The organisms were rooted in the soft bottom, growing erect and branched, in part as gracile pinnate colonies (Penniretеpora cf. volgensis Shishova, 1959). Few encrusting forms probably used brachiopods and crinoids as suitable substrates and might be transported into the depositional realm by tempestites (Herbig et al., 2014).
66Outcropping Strunian deposits in that region, i.e. in the surroundings of Velbert, are predominantly silty to fine-grained sandy, in part with shale and carbonate content (see sections in Michels, 1986). The reduced carbonate content in contrast to the Velbert-4 core is most probably due to the weathering of the carbonate content.
4.2. Palaeobiogeographic relations
67The new genus Ratingella gen. n. is restricted to the Upper Devonian (Frasnian-Famennian), with four species recorded till now. One species Ratingella cromfordensis (Nekhoroshev, 1932) is known from the Famennian of Germany. R. cavernosa (Fritz, 1944) is known from the Famennian of New Mexico, USA. Two species are known from Kazakhstan: Ratingella texta (Troizkaya, 1963) (Frasnian-Famennian), and R. tubulosa (Nekhoroshev, 1977) (Famennian).
68The genus Crustopora Dunaeva, 1964 is known with several species from the Devonian and Carboniferous of Eurasia. The earliest recorded species C. dubia Nekhoroshev, 1977, which was described from the Lower Devonian of Kazakhstan, may not belong to this genus. This species develops encrusting colonies and has no acanthostyles. In fact, its morphology is similar to that of the genus Mongoloclema Shishova, 1970 (Lower-Middle Devonian of Eurasia). The latter genus produces dendroid colonies, however. The earliest confirmed Crustopora species is C. aliena Tolokonnikova, 2010 from the Middle Devonian (Givetian) of Gornyi Altai, Russia. Other Crustopora species, which were reported, are: Crustopora fallacis Nekhoroshev, 1977 from the Upper Devonian (Frasnian) of Kazakhstan, C. lubrica (Trizna, 1958) from the Lower Carboniferous (Tournaisian) of the Kuznetz Basin, Russia, C. undulata Balakin, 1975 from the Lower Carboniferous (lower Viséan) of Uzbekistan, C. ramosa Dunaeva, 1964, C. tuberculata Dunaeva, 1964, and C. tubulosa Dunaeva, 1964, all from the Lower Carboniferous (Namurian) of the Donetz Basin, Ukraine.
69The genus Nikiforovella Nekhoroshev, 1948 occurs predominantly in the Carboniferous, with few species surviving in the Lower Permian. The Devonian records of this genus are known exceptionally from the Famennian: N.? verellaformis Lu, 1999 (Xinjiang, China), N. nitida Troizkaya, 1979 (late Famennian, Kazakhstan), and N. amazarica Nekhoroshev, 1960 (late Famennian, Amur region, Russia). This genus shows an unusual diversity at the Devonian-Carboniferous boundary and in the lower Mississippian. The uppermost Famennian (“Strunian”) part of the Velbert Formation contains at least three species of this genus: Nikiforovella sp. described in the present paper, N.cavernosa (Nekhoroshev, 1932) known from the interval of beds 2-14 in the Ratingen-Cromford section and from the Velbert-4 research well (Tolokonnikova et al., 2014), and, finally, N. gracilis Ernst & Herbig, 2010, described from localities near Ratingen. Unstudied material from the Belgian part of the basin contains abundant and diverse representatives of the genus Nikiforovella. This genus has been recently reported from the Viséan of SW Spain (Ernst & Rodríguez, 2013).
70In spite of the strongly biased knowledge of Famennian bryozoans, the general relations of Late Devonian and Early Carboniferous (Mississippian) genera discussed herein suggest faunal migrations between Western Europe, Kazakhstan, Far East of Russia, and China along shelves and platforms of the Palaeotethys, and thus, a common Palaeotethyan bryozoan realm, though internally differentiated as shown by Tolokonnikova & Ernst (2010). As for the late Famennian (“Strunian”) this is especially confirmed by the widespread genus Nikiforovella.
4. Acknowledgements
71We are grateful to Markus Aretz (Toulouse) for his thoughtful review of an earlier version of the manuscript and for his final remarks, and to Patrick Wyse Jackson for his helpful comments. Zoya Tolokonnikova thanks the Deutscher Akademischer Austauschdienst (DAAD), grant PKZ A/13/00100, for the financial support of a field trip in the Rhenish Massif in 2013. Part of her work has been supported by the Russian Government Program of Competitive Growth of Kazan Federal University. Andrej Ernst is grateful to the “Deutsche Forschungsgemeinschaft (DFG)” for financial support (project ER 278/6.1). This study is a contribution to the IGCP 596 “Mid-Palaeozoic climate and biodiversity".
5. References
72Amler, M.R.W., 1995. Die Bivalvenfauna des Oberen Famenniums West-Europas. - 1. Einführung, Lithostratigraphie, Faunenübersicht, Systematik 1. Pteriomorphia. Geologica et Palaeontologica, 29, 19-143.
73Amler, M.R.W. & Herbig, H.-G., 2006. Ostrand der Kohlenkalk-Plattform und Übergang in das Kulm-Becken im westlichsten Deutschland zwischen Aachen und Wuppertal. In Deutsche Stratigraphische Kommission (ed.), Stratigraphie von Deutschland VI. Unterkarbon (Mississippium). Schriftenreihe der Deutschen Geologischen Gesellschaft, 41, 441-477.
74Amler, M.R.W., Rathmann, S. & Richter, E., 1994. Henry Pauls “Etroeungt-Schichten“ des Bergischen Landes. Biostratigraphie und Biofazies am Nordrand des Velberter Sattels. In Hackler, C., Heinrich, A & Krause, E.-B. (eds), Geologie, Paläontologie und Vor- und Frühgeschichte zwischen Lippe und Wupper. Archäologie im Ruhrgebiet, 2, 73-98.
75Astrova, G.G., 1965. Morphology, history of development and system of the Ordovician and Silurian Bryozoa. Trudy Paleontologicheskogo Instituta, 106, 1-432 [in Russian].
76Astrova, G.G. & Morozova, I.P., 1956. About systematics of the order Cryptostomata. Doklady Akademii Nauk SSSR, 110 (4), 661-664 [in Russian].
77Balakin, G.V., 1975.Bryozoa. InMikhno, N. M. & Balakin, G. V. (eds), Foraminifery i mshanki nizhnego karbona Chatkal’skich gor [Foraminifers and bryozoans from the Lower Carboniferous of Chatkalsk Mountains]. Izdatelstvo “Fan”, Tashkent, 62-125 [In Russian].
78Bärtling, R. & Paeckelmann, W., 1928. Erläuterungen zur Geologischen Karte von Preußen und benachbarten deutschen Ländern. Blatt Velbert. 109 pp.
79Böger, H., 1962. Zur Stratigraphie des Unterkarbons im Velberter Sattel. Decheniana, 114 (2), 133-170.
80Borg, F., 1926. Studies on Recent cyclostomatous Bryozoa. Zoologiska Bidrag från Uppsala,10, 181-507.
81Bultynck, P. & Dejonghe, L. 2001. Devonian lithostratigraphic units (Belgium). In Bultynck, P. & Dejonghe, L. (eds), Lithostratigraphic scale of Belgium. Geologica Belgica, 4 (1-2), 39-69.
82Clausen, C.-D. & Korn, D., 2008. Höheres Mitteldevon und Oberdevon des nördlichen Rheinischen Schiefergebirges (mit Velberter Sattel und Kellerwald). In Deutsche Stratigraphische Kommission (ed.), Stratigraphie von Deutschland VIII. Devon. Schriftenreihe der Deutschen Geologischen Gesellschaft, 52, 439-481.
83Conil, R. (with collaboration of Lys, M. & Paproth, E.), 1964. Localités et coupes types pour l’étude du Tournaisien inférieur (révision des limites sous l’aspect micropaléontologique). Académie royale de Belgique, Mémoires de la Classe des Sciences, 2ème série, 15 (4), 1-105.
84Conil, R. & Paproth, E., 1968. Mit Foraminiferen gegliederte Profile aus dem nordwest-deutschen Kohlenkalk und Kulm. Decheniana, 119 (1/2), 51-94.
85Dunaeva, N.N., 1964. Novye mshanki otryada Trepostomata iz nizhnego karbona Donetskogo bassejna [New bryozoans of the order Trepostomata from the Lower Carboniferous of Donetz Basin]. Paleontologicheskij Zhurnal, 1964 (2), 39-44 [in Russian].
86Dunaeva, N.N. & Morozova, I.P., 1967. Peculiarities of development and systematical position of some Upper Paleozoic Trepostomata. Paleontologicheskiy Zhurnal, (1967) 4, 86-94 [in Russian].
87Duncan, H., 1939. Trepostomatous Bryozoa from the Traverse Group of Michigan. University of Michigan Paleontology Contributions, 5 (10), 171–270.
88Ehrenberg, C.G., 1831. Symbolae Physicae, seu Icones et descptiones Corporum Naturalium novorum aut minus cognitorum, quae ex itineribus per Libyam, Aegiptum, Nubiam, Dongalaam, Syriam, Arabiam et Habessiniam, studia annis 1820-25, redirent. Pars Zoologica, 4, Animalia Evertebrata exclusis Insectis. Berolini, 10 pls.
89Ernst, A. & Herbig, H.-G., 2010. Stenolaemate bryozoans from the Latest Devonian (uppermost Famennian) of western Germany. Geologica Belgica,13 (3), 173-182.
90Ernst, A. & Mohammadi, M., 2009. Stenolaemate bryozoans from the Geirud Formation (Upper Devonian/Lower Carboniferous) of Central Alborz (Iran). Paläontologische Zeitschrift, 83 (4), 439-447.
91Ernst, A. & Rodríguez, S., 2013. Stenolaemate bryozoan fauna from the Mississippian of Guadiato Area, southwestern Spain. Spanish Journal of Palaeontology, 28 (2), 173-192.
92Franke, W., Eder, W. & Engel W., 1975. Sedimentology of a Lower Carboniferous shelf-margin (Velbert Anticline, Rheinisches Schiefergebirge, W-Germany). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 150 (3), 314-353.
93Fritz, M.A.,1944. Upper Devonian Bryozoa from New Mexico. Journal of Paleontology, 18 (1), 31-41.
94Hoffmann, N., Lindert, W., Weyer, D. & Illers, K.-H., 1975. Zum Unterkarbon-Vorkommen auf den Inseln Rügen und Hiddensee. Zeitschrift fuer geologische Wissenschaften, 3, 851-873.
95Gorjunova, R.W., 1975.Permian bryozoans of the Pamir. Trudy Paleontologicheskogo Instituta Akademiyi Nauk SSSR, 148, 1-125 [in Russian].
96Gorjunova, R.V., 2006. New bryozoans from the Devonian of Afghanistan and the Carboniferous of Iran. Paleontological Journal, 40 (6), 626–635.
97Herbig, H.-G. & Mamet, B., 2006. A muddy to clear ramp, latest Devonian, Velbert Anticline (Rheinisches Schiefergebirge, Germany). Geologica et Palaeontologica, 40, 1-25.
98Herbig, H.-G., Lobova, D. & Seekamp, V., 2013. Tales of a core: sea-level history, global crises and changing depositional architecture – The Famennian-Viséan of «Velbert 4», westernmost Rhenish Massif, Germany. In Žák, J., Zulauf, G. & Röhling, H.-G. (eds), Crustal evolution and geodynamic processes in Central Europe. Proceedings of the joint conference of the Czech and German geological societies held in Plzeň (Pilsen), September 16–18, 2013. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften, 82, 49.
99Herbig, H.-G., Lobova, D. & Seekamp, V., 2014. Sea-level history during the birth of a foreland Basin: the Famennian–Visean of ‘‘Velbert 4’’, westernmost Rhenish Massif, Germany. In Rocha, R., Pais, J., Kullberg, J.C. & Finney, S. (eds), Strati 2013, First International Congress on Stratigraphy. At the Cutting Edge of Stratigraphy, 397-402.
100Higgs, K.T., Prestianni, C., Streel, M. & Thorez, J., 2013. High resolution miospore stratigraphy of the Upper Fammenian of eastern Belgium, and correlation with the conodont zonation. Geologica Belgica, 16 (1-2), 84-94.
101Johnson, J.G., Klapper, G. & Sandberg, Ch.A., 1985. Devonian eustatic fluctuations in Euramerica. Geological Society of America, Bulletin, 96, 567-587.
102Johnson, J.G., Klapper, G. & Sandberg, Ch.A., 1986. Late Devonian eustatic cycles around margin of Old Red Continent. In Bless, M.J.M. & Streel, M. (eds), Late Devonian events around the Old Red Continent. Annales de la Société géologique de Belgique, 109, 141-147.
103Kayser, E., 1882. Beiträge zur Kenntnis von Oberdevon und Culm am Nordrande des Rheinischen Schiefergebirges. Jahrbuch der Preußischen Geologischen Landesanstalt [für 1881 (2)], 51-91.
104Lu, L., 1999.Famennian-Tournaisian bryozoans of the Aergati Mt., NW Xinjiang. In Palaeozoic fossils of northern Xinjiang, China. Nanjing Institute of Geology and Palaeontology, Academia Sinica, Nanjing, 37-47, 142-186 [In Chinese].
105Mamet, B., Mikhailoff, N. & Mortelmans, G., 1970. La stratigraphie du Tournaisien et du Viséen inférieur de Landelies. Comparison avec les coupes du Tournaisis et du bord nord du synclinal de Namur. Mémoires de la Société belge de Géologie, Paléontologie et d´Hydrologie, 9, 1-80.
106Mamet, B., Mortelmans, G. & Sartenaer, P., 1965. Réflexions à propos du Calcaire d'Etroeungt. Bulletin de la Société belge de Géologie, 74 (1), 41-51.
107Matyja, H., 1988. Famennian facies of Pomerania, northwestern Poland, and the paleogeography of northern Europe. In McMillan, N.J., Embry, A.F. & Glass, D.J. (eds), Devonian of the World: Proceedings of the 2nd International Symposium on the Devonian System. Canadian Society of Petroleum Geologists, Memoir, 14 (Vol. I), 27-94.
108Matyja, H., 1993. Upper Devonian of Western Pomerania. Acta Geologica Polonica, 43 (1-2), 28-94.
109Maziane, N., Higgs, K.T. & Streel, M., 1999. Revision of the late Famennian miospore zonation scheme in eastern Belgium. Journal of Micropalaeontology, 18 (1), 17-25.
110Nakrem, H.A., Błaźejowski, B. & Gaździcki, A., 2009.Lower Permian bryozoans from southern and central Spitsbergen, Svalbard. Acta Palaeontologica Polonica, 54 (4), 677-698.
111Nekhoroshev, V.P., 1932. Die Bryozoen des deutschen Unterkarbons. Abhandlungen der Preussischen Geologischen Landesanstalt, Neue Folge, 141, 1-74.
112Nekhoroshev, V.P., 1948. Devonian Bryozoa of the Altai (Paleontologiya SSSR: Devonskie mshanki Altaya). Paleontology of the USSR, 3, 1-172 [in Russian].
113Nekhoroshev, V.P.,1960. Nekotorye vidy paleozoiskikh kriptostomat SSSR [Some species of Cryptostomata of USSR].InMarkowskii, B.P. (ed.), Novye Vidy Drevnikh Rastenii i Bespozvonochnykh SSSR [New species of fossil plants and invertebrate animals], 1: 288-283; VSEGEI, Moscow [in Russian].
114Nekhoroshev, V.P., 1977. Devonian bryozoans of Kazakhstan. Trudy VSEGEI, N.S., 186, 1-192 [In Russian].
115Nikiforova, A.I., 1933. The Carboniferous deposits of Central Asia: contributions to the knowledge of the Lower Carboniferous Bryozoa of Turkestan. Trudy Vsesoyusnogo Geologo-razvedochnogo obyedineniya, 207, 1-76 [in Russian].
116Nikiforova, A.I., 1948. Nizhne-kamennougolnye mshanki Karatau [Lower Carboniferous Bryozoa of Karatau].AN KazSSR, Alma-ata, 53 p. [in Russian].
117Michels, D., 1986. Ökologie und Fazies des jüngsten Ober-Devon von Velbert (Rheinisches Schiefergebirge). Göttinger Arbeiten für Geologie und Paläontologie, 29, 1–86.
118Owen, D.E., 1973. Carboniferous Bryozoa from County Tyron. Geological Journal, 8 (1), 297-306.
119Paeckelmann, W., 1913. Das Oberdevon des Bergischen Landes. Abhandlungen der preußischen geologischen Landesanstalt, Neue Folge, 70, 1-356.
120Paeckelmann, W., 1922. Oberdevon und Untercarbon der Gegend von Barmen. Jahrbuch der Preußischen Geologischen Landesanstalt, Neue Folge, 41 (2), 52-147.
121Paproth, E. & Streel, M., 1970. Corrélations biostratigraphiques près de la limite Dévonien/Carbonifère entre les facies littoraux ardennais et les facies bathyaux rhénans. In Colloque sur la stratigraphie du Carbonifère. Les Congrès et Colloques de l’Université de Liège, 55, 365-398.
122Paproth, E. & Streel, M., 1982 (eds). Devonian-Carboniferous transitional beds of the northern "Rheinisches Schiefergebirge".– IUGS, Commission on Stratigraphy, Working Group on the Devonian/Carboniferous Boundary, Guidebook, 63 pp., Liège.
123Paproth, E., Conil, R., Bless, M.J.M., Boonen, P., Bouckaert, J., Carpentier, N., Coen, M., Delcambre, B., Deprijck, C., Deuzon, S., Dreesen, R., Groessens, E., Hance, L., Hennebert, M., Hibo, D., Hahn, G., Hahn, R., Hislaire, O., Kasig, W., Laloux, M., Lauwers, A., Lees, A., Lys, M., Op De Beek, K., Overlau, P., Pirlet, H., Poty, E., Ramsbottom, W., Streel, M., Swennen, R.,Thorez,J., Vanguestaine, M., Van Steenwinkel, M. & Vieslet, J.L., 1983. Bio- and lithostratigraphic subdivisions of the Dinantian in Belgium, a review. Annales de la Société géologique de Belgique, 106, 185-239.
124Paproth, E., Dreesen, R. & Thorez, J., 1986. Famennian paleogeography and event stratigraphy of northwestern Europe. In Bless, M.J.M. & Streel, M. (eds), Late Devonian events around the Old Red Continent. Annales de la Société géologique de Belgique, 109, 175-186.
125Paul, H., 1937. The relationship of the Pilton Beds of North Devon to their equivalents on the Continent. Geological Magazine, 74 (10), 433-442.
126Paul, H., 1939. Die Etroeungt-Schichten des Bergischen Landes. Jahrbuch der Preußischen geologischen Landesanstalt, 59 (für 1938), 647-726.
127Ribbert, K.H., 1998. Das Famenne im Untergrund der Niederrheinischen Bucht. Fortschritte in der Geologie von Rheinland und Westfalen, 37, 81-107.
128Ribbert, K.-H. & Lange, F.G. (1993). Klastika und Carbonate im Mittel- und Oberdevon des Velberter Sattels. 145 Hauptversammlung der Deutschen Geologischen Gesellschaft, Krefeld 1993, Exkursionsführer, 5-16 [Geologisches Landes-Amt Nordrhein-Westfalen].
129Ross, J.R.P., 1961. Ordovician, Silurian and Devonian Bryozoa of Australia. Bureau of Mineral Resources Bulletin, 50, 1–172.
130Sakagami, S. & Sugimura, A., 1987. Hinaclema, a new Carboniferous bryozoan genus from the Hina Limestone, southwest Japan. Proceedings Japan Academy, B (Physical and Biological Sciences), 63, 246-249.
131Shishova, N.A., 1959. New species of the genus Penniretepora from the Carboniferous of the Moscow region. Materialy k Osnovam paleontologii, 3, 16-27 [in Russian].
132Shishova, N.A., 1970. Some new Silurian and Devonian bryozoans from Mongolia. In Astrova, G.G. & Chudinova, I.I. (eds), New species of Paleozoic Bryozoa and corals, 28-31. Nauka, Moscow [in Russian].
133Streel, M., Brice, D. & Mistiaen, B., 2006. Strunian. In Dejonghe, L. (ed.), Chronostratigraphic units named from Belgium and adjacent areas. Geologica Belgica, 9 (1-2), 105-109.
134Tolokonnikova Z.A., 2010. New bryozoans from the Devonian of Gorny Altai (Russia). Paleontological Journal, 44 (2), 151–156.
135Tolokonnikova, Z. & Ernst, A., 2010. Palaeobiogeography of Famennian (Late Devonian) bryozoans. Palaeogeography, Palaeoclimatology, Palaeoecology, 298, 360-369.
136Tolokonnikova, Z., Ernst, A. & Herbig, H.-G., 2014. Famennian (Upper Devonian) bryozoans from borehole Velbert 4, Rhenish Massif (Germany). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 273 (1), 25-44.
137Trizna, V.B., 1958. Rannekamennougolnye mshanki Kuznetzkoi kotloviny [Early Carboniferous bryozoans of the Kuznetz depression]. Trudy Vsesoyuznogo Neftanogo Nauchno-Issledovatel’skogo Geologo-Razvedochnogo Institua, 122, 1-436 [in Russian].
138Troizkaya, T.D., 1963. Bryozoans of the Upper Silurian, Middle, Upper Devonian and Lower Carboniferous. In: Stratigrafiya i fauna paleozoiskikh otlozhenii khrebta Tarbagataya, 209-227. Moscow State University, Moscow [in Russian].
139Troizkaya, T.D., 1968. Devonian Bryozoa of Kazakhstan. Nedra, Moscow, 239 p. [in Russian].
140Troizkaya, T.D., 1979. Bryozoans of Masterovi Horizon in Central Kazakhstan. Paleontological Journal, 4, 31-39 [in Russian].
141Ulrich, E.O., 1882. American Palaeozoic Bryozoa. The Journal of the Cincinnati Society of Natural History, 5, 121-175, 233-257.
142Ulrich, E.O., 1883. American Palaeozoic Bryozoa. The Journal of the Cincinnati Society of Natural History, 6, 245-279.
143Ulrich, E.O., 1890. Palaeozoic Bryozoa: III. Report of the Geological Survey of Illinois, 8, 283-688.
144Vine, G.R., 1884. Fourth report of the committee, consisting of Dr. H. C. Sorby and Mr. G. R. Vine, appointed for the purpose of reporting on fossil Bryozoa. In Report of the British Association for the Advancement of Science (Southport, 1883), John Murray, London, 161-209.
145Weber, H.M. & Wyse Jackson, P.N., 2006. Bryozoen. In Deutsche Stratigraphische Kommission (ed.), Stratigraphie von Deutschland VI. Unterkarbon (Mississippium). Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften, 41, 101–105.
146Whidborne, G.F., 1896. A preliminary synopsis of the faunas of the Pickwell Down, Baggy, and Pilton Beds. Proceedings of the Geological Association, 14, 371-377.
147Whidborne, G.F., 1898. A monograph of the Devonian fauna of the south of England. The fauna of the Marwood and Pilton Beds. Palaeontographical Society, Monograph 52, v. 3, no. 3, p. 214-236, pl. 29-38.
148Zagora, K., 2008. Rügen/Hiddensee (und Usedom). In Deutsche Stratigraphische Kommission (ed.), Stratigraphie von Deutschland VIII, Devon, 574-577.
149Manuscript received 17.04.2014, accepted in revised form 25.03.2015, available on line 14.04.2015.
150Plate 1. A-F: Ratingella cromfordensis (Nekhoroshev, 1932). A-B: tangential section, SMF23.101. C-D: tangential section (arrows – macrozooecia), SMF23.102. E-F: longitudinal section, SMF23.101.
151Plate 2. A-I: Crustopora fistulosa sp. n. A-B: longitudinal section, paratype SMF23.104. C: branch transverse section, holotype SMF23.106. D-G: tangential section (F: arrows – acanthostyles), holotype SMF23.106. H: longitudinal section showing diaphragms in autozooecia and exilazooecia (arrow – diaphragm in exilazooecium), holotype SMF23.106. I: branch transverse section showing wall microstructure with cortex, holotype SMF23.106.
152Plate 3. A-E: Nikiforovella sp., SMF23.100. A-C: tangential section showing autozooecial apertures, metazooecia and acanthostyles. D-E: longitudinal section.
Pour citer cet article
