
A reappraisal of the stratigraphy of the upper Miocene unit X in the Maaseik core, eastern Campine area (northern Belgium)
Abstract
The stratigraphy of the Tortonian-Messinian sequence from the Maaseik core, located on the shoulder of the Roer Valley Graben (RVG) in the eastern Campine area in northern Belgium, was improved. The analysis of the marine palynomorphs (dinoflagellate cysts and acritarchs) from the uppermost part of the Breda Formation, the unnamed unit X and the basal part of the Lower Waubach Member led to the recognition of the mid to upper Tortonian Hystrichosphaeropsis obscura biozone. Therefore deposition of this entire analyzed sequence took place sometime between 8.8 to 7.6 Ma. Paleoenvironmental interpretation of the palynomorphs points to shallow marine conditions and most probably a stressed environment during the deposition of unit X. A comparison with the time equivalent stratigraphy in the nearby Belgian Campine, the Dutch RVG and the German Lower Rhine Basin allowed the identification of the Inden Formation and required a shift in the base of the Kieseloolite Formation compared to the earlier lithostratigraphic interpretation of the Maaseik core. The regional stratigraphic scheme shows the progressive northwestward extension of the river facies from the Lower Rhine during the late Tortonian.
1. Introduction
1The 302 m deep Maaseik borehole (BGD 49W0220, DOV kb18d49w-B220), located in the Roer Valley Graben (RVG) affecting part of northeast Limburg, was drilled in 1980 as a reconnaissance borehole at the initiative of the Geological Survey of Belgium (Fig. 1). A stratigraphic interpretation of this core was published by Vandenberghe et al. (2005). Lithologically the pre-Quaternary section can be subdivided in two main parts: above 198 m depth mainly pale-colored and quartz-rich sand with clays and brown coal or lignite towards its top and below 198 m depth occurs green glauconite-rich sand. The basal part of the pale-colored sand between 192.7 m and 198 m depth is singled out because of its finer grain size (modal size between 128 and 174 µm), high mica content, faint lamination, a few percentages of glauconite pellets and high natural gamma ray (GR) signature. This basal part of the pale-colored sand above 198 m was interpreted as a shallow marine deposit. Its position in the regional lithostratigraphy was unknown and therefore it was given the name unit X by Vandenberghe et al. (2005). The overlying fluviatile quartz-rich sand and the clays and brown coal above 192.7 m depth was recognized as the Kieseloolite Formation while the glauconite sand below 198 m was recognized as the marine Breda Formation.
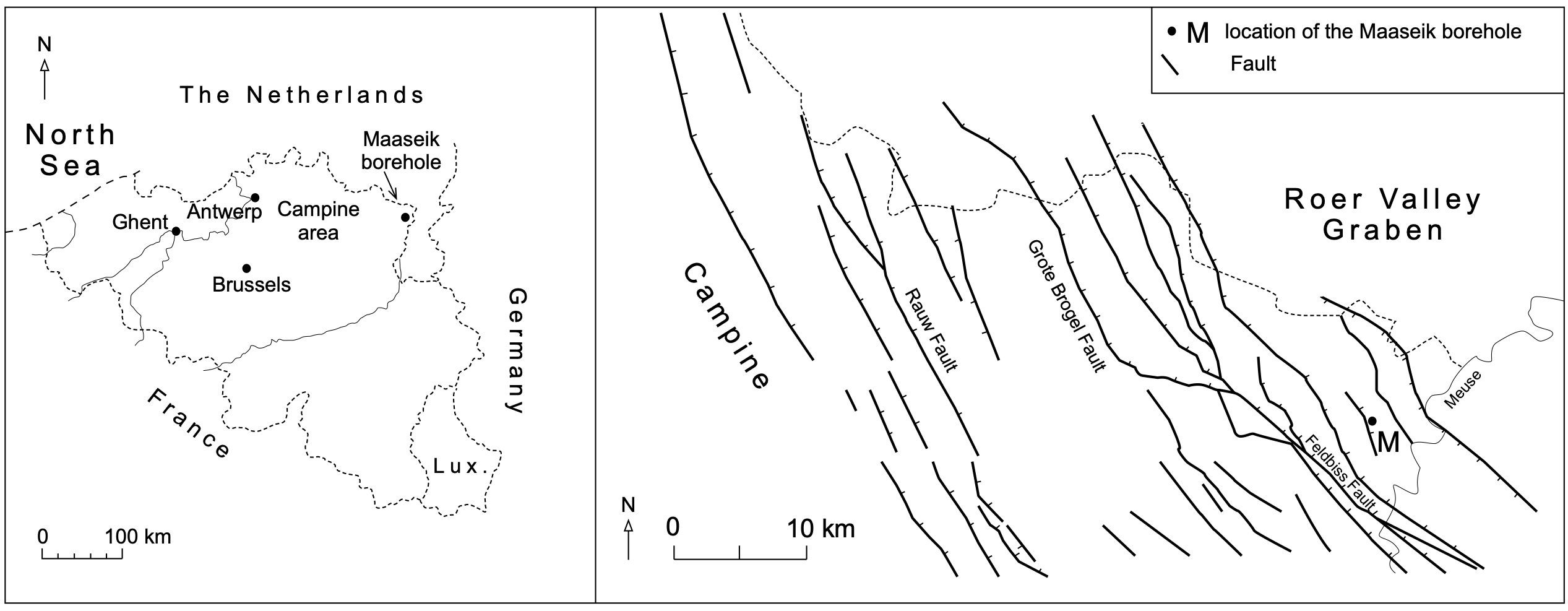
Figure 1. Location of the study area in northern Belgium. The fault pattern is part of the western border of the Roer Valley Graben (modified after Vandenberghe et al., 2005).
2In addition to mollusks and calcareous microfossils in the Breda Formation, also dinoflagellate cysts including acritarchs and green algae were sampled between 191.5 m and 295.8 m (16 samples). With respect to the dinoflagellate investigations the sample at 191.5 m just above unit X in the base of the Kieseloolite Formation was impoverished in species and compared to the uppermost ‘unnamed’ Subzone in the Nieder Ochtenhausen research core in northern Germany (Strauss et al., 2001); based on this comparison a Syltian (regional North Sea stage) uppermost Miocene age was proposed for unit X. The top part of the Breda Formation below showed a DN9 association with the transition to a DN8 association occurring between samples at 220.5 m and 227.5 m (DN biozones sensu de Verteuil & Norrris, 1996); the associations of dinoflagellates and of calcareous microfossils in the samples between 198 m and about 235 m point to associations comparable to those in the Dessel and Deurne Members of the upper Miocene Diest Formation in the Campine and the Antwerp area. Note that within unit X no samples were analyzed and only the base of the Kieseloolite Formation just above unit X was sampled at 191.5 m.
3Fifty samples were taken for an investigation with terrestrial palynomorphs in order to study the pale-colored quartz-rich sand. Thirty-one samples yielded a reliable pollen assemblage, the lowest one at 197.90 m which is the very base of unit X (Vandenberghe et al., 2005). With respect to the pollen assemblages it was concluded that the base of unit X did not basically differ from the Kieseloolite Formation content except for its low total pollen content, the very low content of Ericales and the presence of Mastixia occurring commonly in the Miocene, whilst the Kieseloolite Formation section was interpreted to be of Pliocene age.
4Recent attempts to incorporate the Maaseik 49W0220 core stratigraphy in regional stratigraphic schemes have proposed different correlation solutions (Vandenberghe et al., 2005; Dusar et al., 2012; Vernes et al., 2018; Vandenberghe et al., 2020, this volume). In these recent studies the Waubach Member above unit X was subdivided in an upper and a lower part and the latter often indicated as Inden Formation (e.g. Menkovic & Westerhoff, 2010; Dusar et al., 2012; Vernes et al., 2018, fig. 7.3; Verhaegen, 2020, this volume; Vandenberghe et al. 2020, this volume). Such a subdivision in the Waubach Member is figured in Wong et al. (2007, p. 163) with the boundary between the Lower and Upper Waubach Member situated at the turn of the Tortonian to Messinian.
5In order to improve the data set for the interpretation of the stratigraphic succession in the Maaseik core a reinvestigation of the dinoflagelate cyst stratigraphy in particular of unit X was required since unit X itself was not sampled for dinocysts in the original study by Vandenberghe et al. (2005). Therefore unit X, the base of the Lower Waubach Member just above unit X, and the top of the Breda Formation just below unit X were resampled and reinvestigated making use of recent progress in the dinoflagellate zonation of the North Sea Basin.
2. Material and methods
6In order to assess the stratigraphy of the sediments of unit X, a renewed sampling in the core repository of the Geological Survey of Belgium and a subsequent palynological analysis was carried out. Three samples were taken in unit X itself, at 196.5, 195.5 and 194.5 m, and an additional sample in the uppermost part of the Breda Formation at 202.5 m and in the lowermost part of the Kieseloolite Formation at 191 m in the Waubach Member. The relative abundances of marine palynomorphs in these newly investigated samples were combined with sample 201.5 m from the Breda Formation and sample 191.5 m from the Waubach Member as previously reported in Vandenberghe et al. (2005) (Table 1).
7All samples investigated for palynological analysis in this study and in Vandenberghe et al. (2005) were processed according to the procedure described in Louwye et al. (2004). The maceration involved a treatment with HCl for the removal of carbonates and a subsequent treatment with HF for the removal of silicates. A short ultrasonication for 15 s was carried out for an adequate dispersion. The residues were filtered on a nylon screen with a 16 µm mesh size, and strew mounted on one slide with glycerine jelly. The microscopic analysis was carried out with a Zeiss AxioImager A1 transmitted light microscope at 200x and 400x magnifications until a minimum of 250 palynomorphs were counted in non-overlapping traverses. The remainder of the slide was then counted for rare specimens. The nomenclature used is after Williams et al. (2017).
3. Results
8Only samples 202.5 m and 201.5 m (Breda Formation) and samples 191.5 m and 191.0 m (Waubach Member of the Kieseloolite Formation) yielded enough palynomorphs for a count of 250 palynomorphs. All samples from unit X (196.5 m, 195.5 m and 194.5 m) yielded a very low diversity and abundance of marine palynomorphs and the targeted count of 250 palynomorphs was not reached in one slide (Table 1).
9Reworked marine palynomorphs are virtually absent in the upper part of the Breda Formation, only sample 201.5 m holds two reworked Paleogene specimens (Vandenberghe et al., 2005). The Waubach Member is devoid of reworked marine palynomorphs (Vandenberghe et al., 2005; this study), as is the case in the unit X samples.
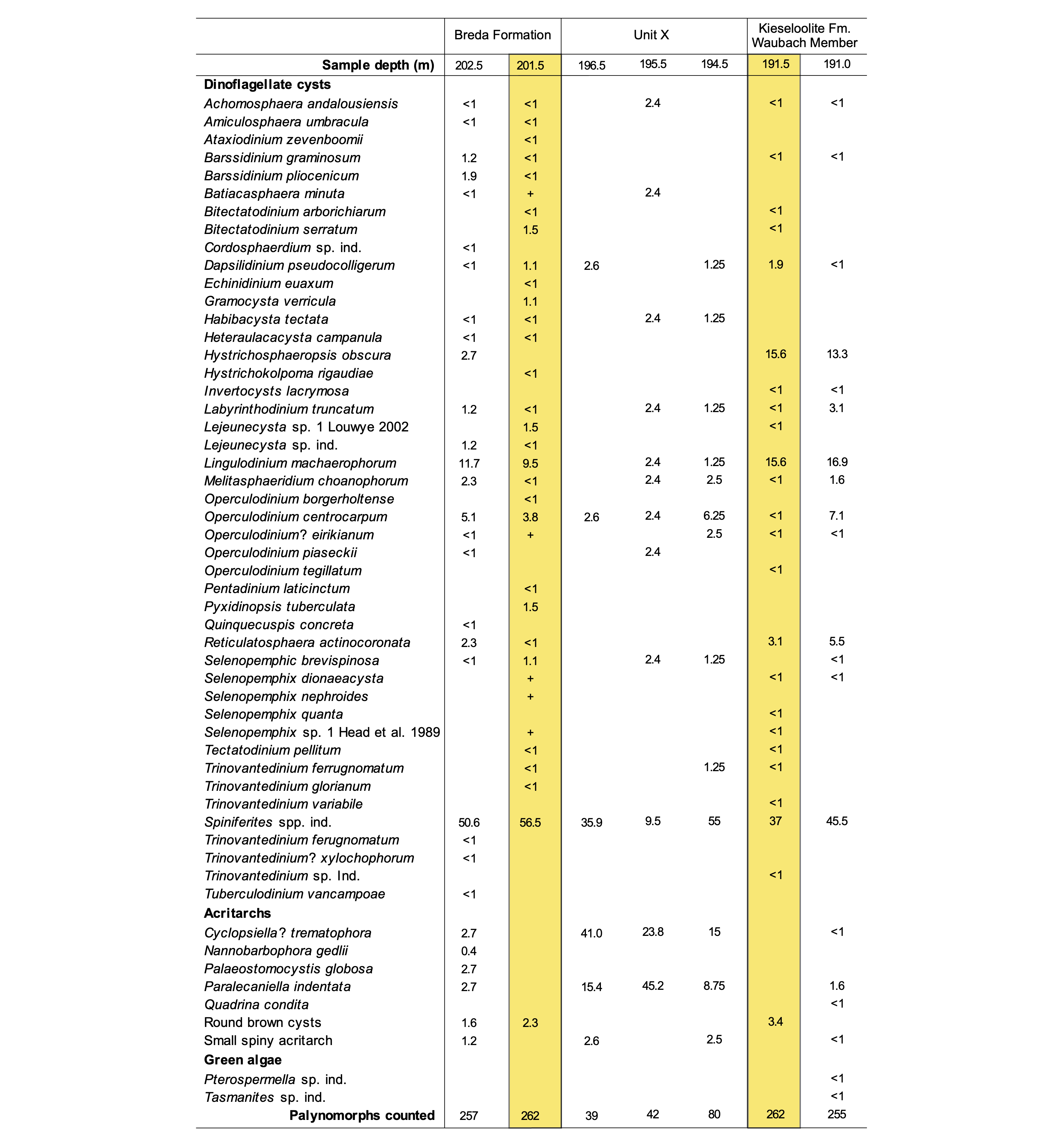
Table 1. Marine palynomorphs recorded in samples from the Breda Formation (samples 202.5, 201.5), unit X (sample nos. 196.5, 195.5, 194.5) and Lower Waubach Member (191.5, 191.0). Samples 201.5 and 191.5 were already studied by Vandenberghe et al. (2005). Percentages of the total palynomorph count are given. Core depth in m.
3.1. Biostratigraphy
10Based on the biozonation by de Verteuil & Norris (1996), Vandenberghe et al. (2005) concluded that the sediments in the 220.5 m to 191.5 m interval, representing the top of the Breda Formation and the basal sample of the Kieseloolite Formation (Waubach Member) and thereby automatically including the unit X in between their two topmost samples, hold the DN9 Hystrichosphaeropsis obscura Zone. This de Verteuil & Norris (1996) zone is defined from the highest occurrence (HO) of Sumatradinium soucouyantiae to the HO of the eponymous species. An additional diagnostic bioevent is the HO of Palaeocystodinium spp. at the upper boundary of the subjacent zone. The chronostratigraphic position of the DN9 zone, according to de Verteuil & Norris (1996) is lower to middle upper Miocene, i.e. upper Tortonian. Nevertheless, based on the comparison with the Nieder Ochtenhausen research core (northern Germany) (Strauss et al., 2001), Vandenberghe et al. (2005) could also propose a Syltian age, the regional North Sea stage of latest Tortonian and Messinian age, for the basal sample of the Kieseloolite Formation (Lower Waubach Member) at 191.5 m and proposed also this age for the unit X.
11The assemblages in the interval 202.5 m to 191 m, in the newly collected samples and in the samples studied in Vandenberghe et al. (2005), are now biostratigraphically interpreted with the more recent and age-calibrated dinoflagellate cyst biozonation by Dybkjær & Piasecki (2010) (Fig. 2). All samples in the interval 202.5 m to 191 m hold their Hystrichosphaeropsis obscura Zone, defined as the interval from the HO of Barssdinium evangelinae to the HO of the eponymous species. Other bioevents are the HO of Labyrinthodinium truncatum close to the upper boundary, and the HO occurrence of Palaeocystodinium spp. coinciding with the lower boundary of the zone (Table 1). Dybkjær & Piasecki (2010) provide an age of 8.8 to 7.6 Ma for the zone, i.e. mid to late Tortonian. Neither the upper nor the lower boundary of the H. obscura zone are recognized in the samples implying that unit X was deposited somewhere in the time interval between 8.8 and 7.6 Ma. Dybkjær & Piasecki (2010) parallel their H. obscura zone with the DN9 Zone of de Verteuil & Norris (1996), the DN9 Zone of Köthe (2003), the upper partim of the Aum Zone of Strauss et al. (2001), and the M14 Zone of Munsterman & Brinkhuis (2004).
12Therefore the present biostratigraphic study confirms the late Tortonian DN9 stratigraphic assessment and invalidates the Syltian age interpretation discussed in Vandenberghe et al. (2005).
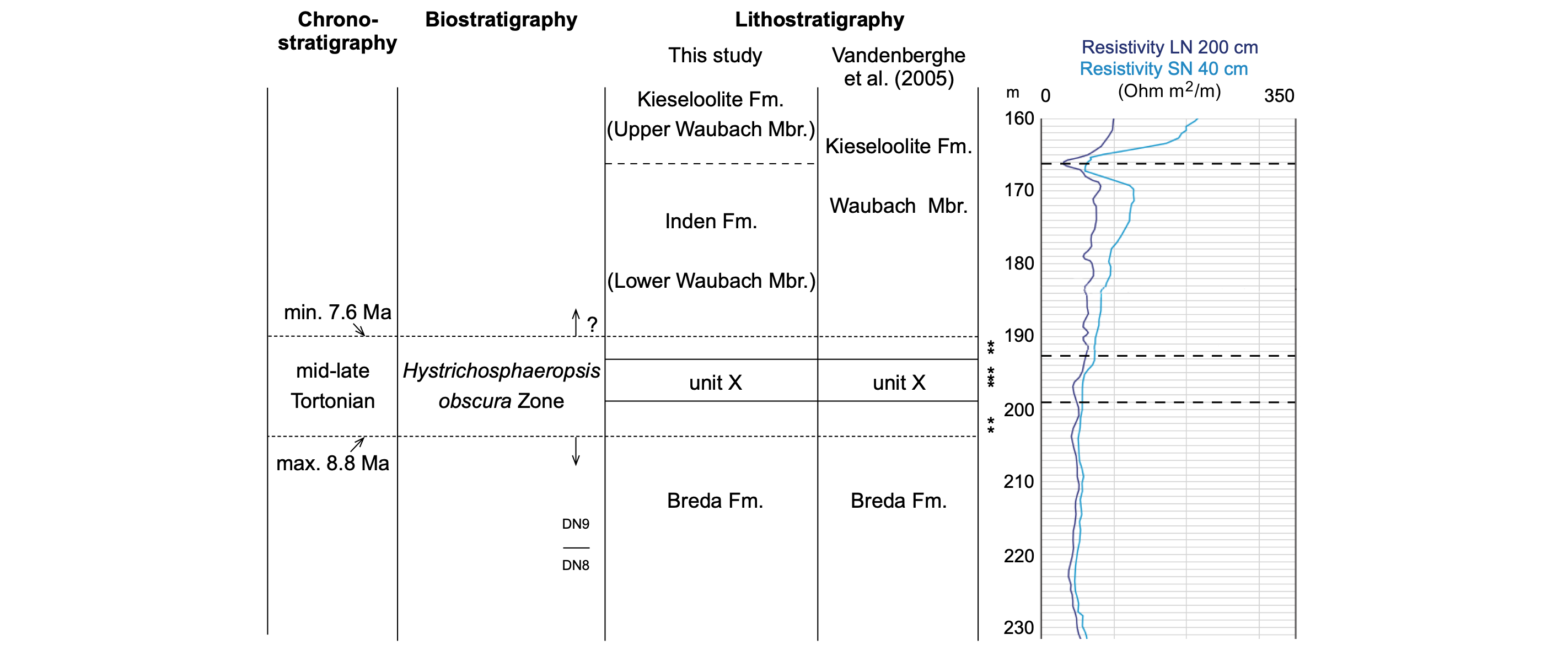
Figure 2. Chronostratigraphic position of unit X based on the interpretations presented in the current study. Position of the samples (*) newly investigated in the present study. They all hold the H. obscura Zone, age calibrated between 7.6 and 8.8 Ma indicating a mid-late Tortonian age (7.246 to 11.63 Ma). The position of the DN8/DN9 dinoflagellate cyst zone boundary is taken from Vandenberghe et al. (2005), and is based on the dinoflagellate cyst biozonation by de Verteuil & Norris (1996). The lithostratigraphic interpretations discussed in the present paper are compared to the lithostratigraphy interpreted in Vandenberghe et al. (2005). Note that all samples are located with respect to core depths and lithological core descriptions and that a slight misfit exists between drilling depth and the geophysical borehole depths, recorded after coring.
3.2. Paleoenvironmental and taphonomical considerations
13The dinoflagellate cyst assemblages recorded in the upper part of the Breda Formation (samples 202.5 m and 201.5 m) typically reflect a neritic depositional environment, as testified a.o. by the abundance of the neritic Spiniferites species. No indications of oceanic influence are observed given the absence of indicator species of typical genera such as Impagidinium and Nematosphaeropsis. Based on the lithology of the glauconite-rich Breda Formation section Vandenberghe et al. (2005) advance a depositional depth close to the minimal depth for glauconite formation being around 10 m – 20 m.
14The diversity and abundance of marine palynomorphs in the unit X samples is very low compared to the samples from the upper part of the Breda Formation and even from the base of the Waubach Member of the Kieseloolite Formation. Given the numbers of the thin-walled species of the genera Cyclospiella and Selenopemphix, no selective mild oxidation nor mechanical degradation can be postulated as a causal taphonomic mechanism behind the poor diversity and abundance. Dilution of the palynological signal by a high sedimentation rate is also an unlikely mechanism taking into account the selective presence of species. The dinoflagellate cyst assemblage from unit X is not readily interpreted given this very low diversity and abundance. The absence of the above-mentioned oceanic species is indicative of a neritic environment, corroborated by the relative high numbers of Spiniferites species. However, the presence of high numbers of the acritarch Paralecaniella indentata is noteworthy. Louwye & Laga (2008) discussed records of high abundance of Paralecaniella indentata in Cenozoic deposits. This acritarch abounds often in stressed marginal marine environments characterized by environmental parameters such as, a.o., increased stratification, reduced salinities, and shoaling phases. Species of the thin-walled acritarch genus Cyclopsiella are recorded in relatively high number in unit X. Matsuoka & Head (1992) suggest that Cyclopsiella most likely has a colony-like mode of life attached to the substrate. Louwye & Laga (2008) concluded in their review that acmes of Cyclopsiella can be indicative of shallow marine to nearshore, high-energetic environments. Deposition of unit X took most probably place in a stressed marine environment, unfavorable for marine palynomorphs.
15Marine palynomorphs are also present in the basal samples of the Waubach Member of the Kieseloolite Formation (191.5 m and 191 m). This presence of marine palynomorphs is rather surprising given the paleoenvironmental interpretation of the greater part of the Waubach Member as river deposits (Vandenberghe et al., 2005; Westerhoff, 2009). However, Westerhoff (2003) already reported that the depositional setting of the Waubach Member was partly marine at its base. The preservation of the palynomorphs is however good, and no reworked pre-Neogene palynomorphs are recorded. The palynomorph diversity is lower in the upper sample of the two analyzed (sample 191.5 m: 24 species, sample 191 m: 20 species), but no data is available on whether or where the marine signal fades out higher in the sequence. The presence of the marine palynomorphs may indicate that the Waubach Member river plain initially developed close to the sea.
16The marine palynomorphs sampled near the base of unit X reflect a similar depositional environment as for the upper part of the Breda Formation, although the diversity is circa one third lower. The palynomorphs reflect a neritic depositional environment with abundant Spiniferites. Similarly as in the top of the Breda Formation, genera such as Impagidinium and Nematosphaeropsis are absent.
4. Discussion
4.1. Hystrichosphaeropsis obscura Zone in the top Breda Formation, the unit X and the base of Lower Waubach Member. Time equivalence of the Lower Waubach Member with the Inden Formation of the Lower Rhine Basin
17The recognition of the Hystrichosphaeropsis obscura Zone of Dybkjær & Piasecki (2010) in the samples of the top Breda Formation, the unit X and the basal part of the Waubach Member of the Kieseloolite Formation situates this interval with three lithostratigraphic units in one single dinoflagellate cyst biochron age calibrated between 7.6 and 8.8 Ma in the later part of the Tortonian, and equivalent to the de Verteuil & Norris (1996) DN9 dinoflagellate cyst biozone (Fig. 2). Earlier analyses already established the boundary between the dinoflagellate cyst DN8 and DN9 zones in the Maaseik core between 220.5 m and 227.5 m. The calcareous microfossils (benthic foraminifers and ostracods) in the top of the Breda Formation in the Maaseik core holding the DN9 zone (de Verteuil & Norris, 1996) all point to a comparison with the microfossil content of the Deurne and Dessel Members of the Diest Formation in the Antwerp and Campine area (Vandenberghe et al., 2005).
18In the subsiding RVG, where the Maaseik core is located, the switch from fully marine deposits of the Breda Formation to the quartz-rich shallow/marginal marine unit X and finally to the continental river plain dominated sediments of the Lower Waubach Member occurred during the later part of the Tortonian between 8.8. and 7.6 Ma. In the Lower Rhine Basin, to the southeast, the transition from the marginal marine Ville Formation including several lignite seams to the fluvial dominated Inden Formation occurred already at the transition of the Serravallian to the Tortonian (Schäfer & Utescher, 2014, fig. 3; Utescher et al., 2020). These last authors subdivide the Inden Formation in three sequences each starting with a fluviatile to lacustrine deposit and evolving into a brown coal seam. Based on the time constraints given by Utescher et al. (2020, fig. 3) and the time constraints imposed by the Hystrichosphaeropsis obscura biochron, the marginal marine unit X deposits are time equivalent with the top middle or upper sequence of the meandering fluviatile to lacustrine Inden Formation (beds 7C to 7F in the Inden Formation). It appears that during that time the continental realm in the Lower Rhine Basin expanded westward into the Belgian northeast Limburg. The deposition of the glauconitic marine Breda Formation sediments was replaced by the sedimentation of marginal marine quartz sand of unit X. Wong et al. (2007, p. 160) also state that the Inden Formation in western direction interdigitates with the marine Breda Formation.
4.2. The stratigraphic position of the Lower and Upper Waubach Members, the Inden Formation and the Kieseloolite Formation in the RVG
19In the Lower Rhine Basin at the turn of the Tortonian to the Messinian, the coarse-grained braided river Hauptkies facies developed holding silicified oolites as in the Kieseloolite Formation (Schäfer & Utescher, 2014). In the southern Netherlands, Wong et al. (2007, p. 163) interpret the base of the Upper Waubach Member as the base of the Messinian but still incorporate the Lower Waubach Member in the Kieseloolite Formation. On the other hand, in recent studies of the Maaseik core stratigraphy (Dusar et al., 2012, p. 18; Vernes et al., 2018, fig. 7.3) Lower Waubach Member is split from the Kieseloolite Formation and named Inden Formation. This last practice is introduced in the Netherlands since the official introduction of the Inden Formation in the nomenclature (Menkovic & Westerhoff, 2010) and has been duly considered in the recent stratigraphic scheme of Munsterman et al. (2019, fig. 8). Unfortunately, in the Maaseik core no good stratigraphic markers are available higher up in the Waubach members above its very base which is still containing an upper Tortonian dinoflagellate cyst assemblage. Neither the palyno-stratigraphic analysis reported in Vandenberghe et al. (2005, p. 13) is decisive and hesitates between a Miocene and Pliocene age. Better stratigraphic control in the Maaseik core only resumes with the overlying Brunssum clay and lignite complex of which the pollen spectra resemble those of the Maat lignite in the top of the Mol Formation to the west (Fig. 3); the latter formation in the west forms the top of the Pliocene (Laga et al., 2001).
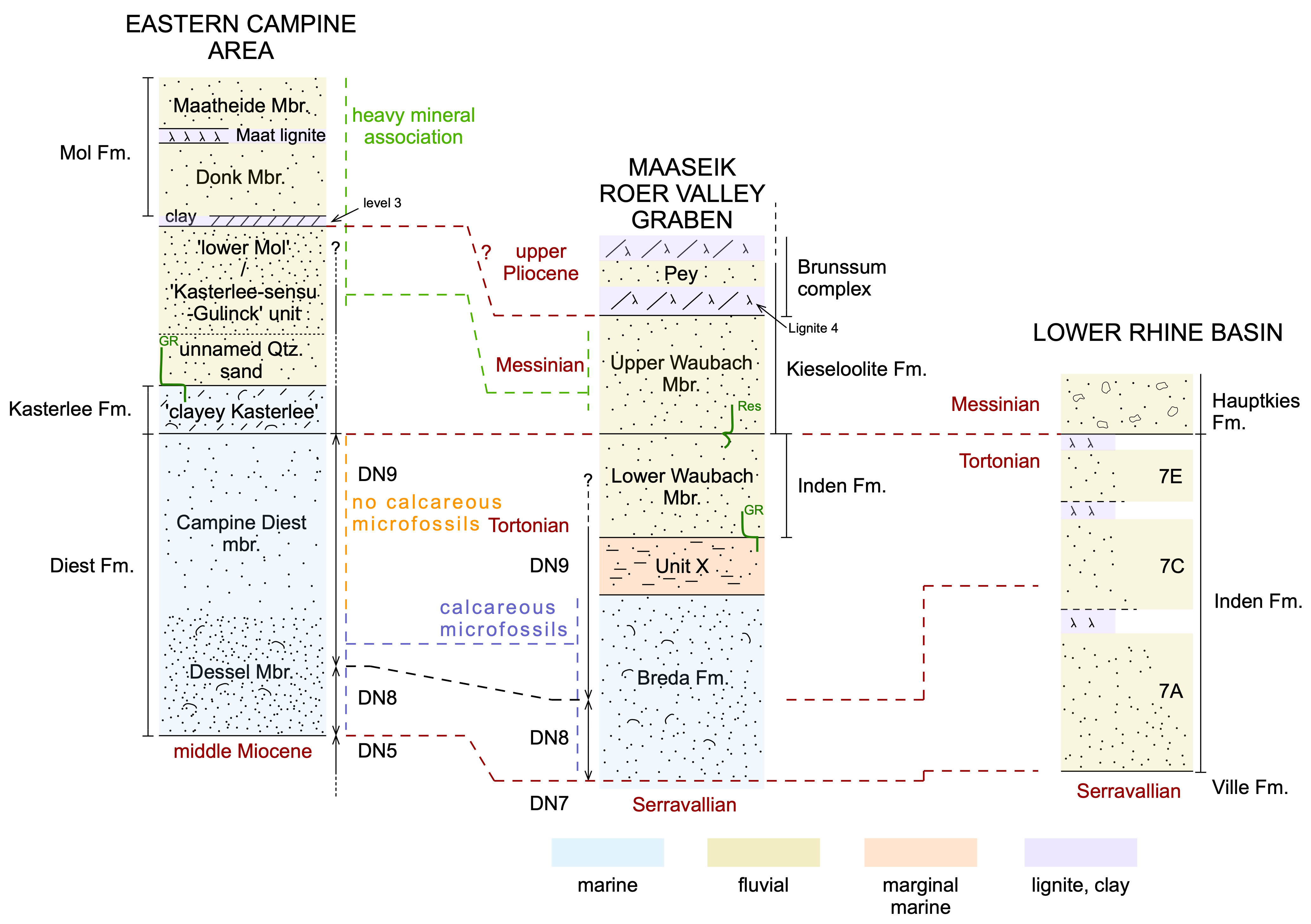
Figure 3. Correlation between schematic lithostratigraphic columns of the upper Miocene to Pliocene of the eastern Campine, the Maaseik core in the Roer Valley Graben and the Lower Rhine Basin. The eastern Campine succession is discussed in Vandenberghe et al. (2020, this volume), the Maaseik core lithostratigraphic succession in the column is after Vandenberghe et al. (2005) whilst to the right of the column the interpretations of the Inden and Kieseloolite Formations as discussed in the present text are indicated. The Lower Rhine Basin stratigraphy is from Schäfer & Utescher (2014) and Utescher et al. (2020). The GR (Gamma Ray), Res (Resistivity) and lignite 4 markers are discussed in Vandenberghe et al. (2020, this volume). Level 3 in the Eastern Campine section is after Vandenberghe et al. (2020, this volume, figs 5 & 6). The heavy minerals are discussed in Verhaegen (2020, this volume).
4.3. A comparison of the eastern Campine stratigraphic succession with the succession in the Maaseik core
20The standard lithostratigraphic sequence in the eastern Campine, located on the shoulder area of the RVG, differs, logically, from the lithostratigraphical sequence in the RVG. The eastern Campine sequence is described in detail by Vandenberghe et al. (2020, this volume): the Diest Formation at the base is overlain by the ‘clayey Kasterlee’ unit of the Kasterlee Formation, followed by the ‘lower Mol’ or ‘Kasterlee-sensu-Gulinck’ unit of uncertain affiliation with a package of unnamed quartz-rich sand at its base and with a clay layer developed at its top. This unit is finally overlain by the Mol Formation which contains in its upper part the Maat lignite horizon (Fig. 3). In this eastern Campine area, the boundary between the DN9 and DN10 associations, and hence the Tortonian-Messinian boundary occurs between the Diest Formation (DN9) and the Kasterlee Formation (DN10).
21In this area the DN9 biozone is recognized in parts of the marine glauconite-bearing Dessel Sand and the superjacent Diest Sand in the Mol (BGD 031W0221, DOV kb17d31w-B212) and Retie (BGD 031W0243, DOV kb17d31w-B228) cores while more to the northwest the DN9 zone is only recognized in the Dessel Sand in e.g. the Oostmalle core (BGD 029E0249, DOV kb16d29e-B276). The DN9 zone has the greatest thickness in the Mol – Retie area (c. 75 m) and thins out in a northwestern direction, and has a thickness of c. 14 m in the Oostmalle area. The DN9 sediments of the Diest Formation are part of a large-scale west to northwestward prograding wedge which is relayed in the northwest by a Diest Formation package containing the DN10 Zone (Louwye & Laga, 2008; Vandenberghe et al., 2014).
22These data imply that the DN9 holding unit X together with the very base of the Lower Waubach Member are time equivalent with the top of the Diest Formation. As reported above, downwards in the Maaseik section the boundary between the DN9 and DN8 dinoflagellates Zones occurs about 20 m below the top of the Breda Formation whilst in the Diest Formation it occurs within its lower Dessel Member (Figs 2, 3).
23The Messinian dinoflagellate Zone DN10 (de Verteuil & Norris, 1996) was observed in the ‘clayey Kasterlee’ unit of the Kasterlee Formation and also in the lower part of the ‘lower Mol’ or ‘Kasterlee-sensu-Gulinck’ unit (eight samples over about 10 m height in the ON-Dessel-2 core) above which the sediment becomes barren (Louwye et al., 2007; Vandenberghe et al., 2020, this volume).
24Assuming that the base of the Upper Waubach Member corresponds indeed with the base of the Messinian as figured in Wong et al. (2007, p. 163) and Munsterman et al. (2019, fig. 8), this implies that the Kasterlee Formation and at least the lower part of the ‘lower Mol’ or ‘Kasterlee-sensu-Gulinck’ unit which contain the Messinian DN10 biochron are time equivalent with the Upper Waubach Member. In that case, one might even hypothesize that the clay layer developed at the top of the ‘lower Mol’ or ‘Kasterlee-sensu-Gulinck’ unit could be related to the basal clay in the Brunssum Complex (Fig. 3). The heavy mineral analysis (Verhaegen, 2020, this volume) supports the correlation of the ‘lower Mol’ or ‘Kasterlee-sensu-Gulinck’ unit with the upper part of the Waubach Member and also indicates that the heavy mineral association of the Lower Waubach Member is significantly different from the association in the Upper Waubach Member.
25In the Netherlands, the river plain facies of the Lower Waubach Member is correlated and equated to the Tortonian Inden Formation in the Lower Rhine Basin (Menkovic & Westerhoff, 2010), a practice applied by Dusar et al. (2012) and Vernes et al. (2018) to the Lower Waubach Member in the Maaseik core. This seems to imply that the dinoflagellate DN9 biochron identified only in its basal layer could extend over the whole Lower Waubach Member.
26During late Tortonian times the fluviatile environment that existed in the RVG expanded westwards. As a consequence in the Maaseik area, the marine glauconite sand of the Breda Formation was replaced by the quartz-rich marginal marine facies of the unit X. Subsequently this marginal marine facies was also replaced by the Inden-type river system depositing the Lower Waubach Sand and Gravel in the Maaseik area. Following the Dutch practice and the facies and time equivalence of the fluvial Hauptkies in the Lower Rhine Basin with the Kieseloolite in the RVG (Schäfer & Utescher, 2014), the base of the Kieseloolite Formation should be put at the base of the Upper Waubach Member. The lithological change between quartz-rich sand and green glauconite-rich sand, as observed in many borehole logs, justifies a main lithostratigraphic distinction at that level. Placing the Lower Waubach Member in the upper Tortonian also implies its equivalence with the upper part of the Diest Formation, just like unit X. This equivalence is not contradicted by the available heavy mineral analysis (Verhaegen, 2020, this volume). The correlation of the top of unit X with the ‘clayey Kasterlee’ unit based on a marked increased GR signal (Vandenberghe et al., 2005, 2020, this volume) and the correlation of the boundary between the Lower and Upper Waubach Members with the top of the clay layer overlying the ‘lower Mol’ or ‘Kasterlee-sensu-Gulinck’ unit as proposed in Vandenberghe et al. (2020, this volume) cannot be upheld.
5. Conclusions
27The marine palynological analysis of this study allowed to situate the top of the Breda Formation, unit X and the basal layer of the Lower Waubach Member in the same upper Tortonian DN9 Zone. These data, together with the practice in the Netherlands of considering the whole Lower Waubach Member as Tortonian, implies a time equivalence of the unit X and the Lower Waubach Member with the top part of the Diest Formation in the eastern Campine area. During late Tortonian times the marine influence in the Maaseik area was replaced first by the marginal marine unit X and later by river plain deposits in the Lower Waubach Member.
28This fluviatile facies is comparable to the time equivalent Inden Formation in the Lower Rhine Basin, and therefore, the Lower Waubach Member has been correlated to the Inden Formation in several recent interpretations of the Maaseik core.
29The coarse sediments of the Upper Waubach Member, considered in the Netherlands as Messinian in age, are time equivalent with the DN10 dinoflagellate containing ‘clayey Kasterlee’ unit of the Kasterlee Formation and ‘lower Mol’ or ‘Kasterlee-sensu-Gulinck’ units. The base of the Upper Waubach Member corresponds in time to the base of the Hauptkies in the Lower Rhine Basin and marks the base of the Kieseloolite. More precise correlations of the Maaseik core section will only be possible if more data characterizing the sediments become available.
6. Acknowledgements
30Marleen De Ceukelaire (Geological Survey of Belgium) kindly provided the samples for this study. Armin Menkovic (Geological Survey of the Netherlands) is thanked for information on the Inden Formation in the Netherlands. The constructive remarks by reviewers Jasper Verhaegen, Linda Prinz and Martin Salamon are kindly acknowledged.
7. References
31de Verteuil, L. & Norris, G. 1996. Miocene dinoflagellate stratigraphy and systematics of Maryland and Virginia. Micropaleontology, Supplement 42, 1–172. https://doi.org/10.2307/1485926
32Dusar, M., De Ceukelaire, M. & Walstra, J., 2012. Stratigrafische interpretatie en correlatie van het Quartair en continentale Neogeen. Unpublished report. Belgische Geologische Dienst, Koninklijk Belgisch Instituut voor Natuurwetenschappen, 81 p.
33Dybkjær, K. & Piasecki, S., 2010. Neogene dinocyst zonation for the eastern North Sea Basin, Denmark. Review of Palaeobotany and Palynology, 161/1-2, 1–29. https://doi.org/10.1016/j.revpalbo.2010.02.005
34Köthe, A., 2003. Dinozysten-Zonierung im Tertiär Norddeutschland. Revue de Paléobiologie, Genève, 22, 895–923.
35Laga, P., Louwye, S. & Geets, S., 2001. Paleogene and Neogene lithostratigraphic units (Belgium). In Bultynck, P. & Dejonghe, L. (eds), Guide to a revised lithostratigraphic scale of Belgium. Geologica Belgica, 4/1-2, 135–152. https://doi.org/10.20341/gb.2014.050
36Louwye, S. & Laga, P., 2008. Dinoflagellate cyst stratigraphy and palaeoenvironment of the marginal marine Middle and Upper Miocene of the eastern Campine area, northern Belgium (southern North Sea Basin). Geological Journal, 43, 75–94. https://doi.org/10.1002/gj.1103
37Louwye, S., Head, M. & De Schepper, S., 2004. Dinoflagellate cyst stratigraphy and palaeoecology of the Pliocene in northern Belgium, southern North Sea Basin. Geological Magazine, 141/3, 353–378. https://doi.org/10.1017/S0016756804009136
38Louwye, S., De Schepper, S., Laga, P. & Vandenberghe, N., 2007. The Upper Miocene of the southern North Sea Basin (northern Belgium): a palaeoenvironmental and stratigraphical reconstruction using dinoflagellate cysts. Geological Magazine, 144/1, 33–52. https://doi.org/10.1017/S0016756806002627
39Matsuoka, K. & Head, M., 1992. Taxonomic revisions of the Neogene marine palynomorphs Cyclopsiella granosa (Matsuoka) and Batiacasphaera minuta (Matsuoka), and a new species of Pyxidinopsis Habib (Dinophyceae) from the Miocene of the Labrador Sea. In Head, M. & Wrenn, J. (eds), Neogene and Quaternary Dinoflagellate Cysts and Acritarchs. American Association of Stratigraphic Palynologists Foundation, Dallas, 165–180.
40Menkovic, A. & Westerhoff, W., 2010. Formatie van Inden. Lithostratigrafische Nomenclator van de Ondiepe Ondergrond. https://www.dinoloket.nl/formatie-van-inden, accessed 25/05/2020.
41Munsterman, D.K. & Brinkhuis, H., 2004. A southern North Sea Miocene dinoflagellate cyst zonation. Netherlands Journal of Geoscience/Geologie en Mijnbouw, 83, 267–285. https://doi.org/10.1017/S0016774600020369
42Munsterman, D.K., ten Veen, J.H., Menkovic, A, Deckers, J, Witmans, N, Verhaegen, J, Kerstholt-Boegehold, S.J., van de Ven, T. & Busschers, F.S., 2019. An updated and revised stratigraphic framework for the Miocene and earliest Pliocene strata of the Roer Valley Graben and adjacent blocks. Netherlands Journal of Geosciences, 98, e8. https://doi.org/10.1017/njg.2019.10
43Schäfer, A. & Utescher, T., 2014. Origin, sediment fill, and sequence stratigraphy of the Cenozoic lower Rhine Basin (Germany) interpreted from well logs. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 165/2, 287–314. https://doi.org/10.1127/1860-1804/2014/0062
44Strauss, C., Lund, J.J. & Lund-Christensen, J., 2001. Miocene dinoflagellate cyst stratigraphy of the Nieder Ochtenhausen research borehole (NW Germany). Geologisches Jahrbuch, Reihe A, 152, 395–447.
45Utescher, T., Ashraf, A., Kern, A.K. & Mosbrugger, V., 2020. Diversity patterns in microfloras recovered from Miocene brown coals of the lower Rhine Basin reveal distinct coupling of the structure of the peat-forming vegetation and continental climate variability. Geological Journal, https://doi.org/10.1002/gj.3801
46Vandenberghe, N., Laga, P., Louwye, S., Vanhoorne, R., Marquet, R., De Meuter, F., Wouters, K. & Hagemann, H.W., 2005. Stratigraphic interpretation of the Neogene marine‐continental record in the Maaseik well (49W0220) in the Roer valley Graben, NE Belgium. Memoirs of the Geological Survey of Belgium, 52, 39 p.
47Vandenberghe, N., Harris, W.B., Wampler, J.M., Houthuys, R., Louwye, S., Adriaens, R., Vos, K., Lanckacker, T., Matthijs, J., Deckers, J., Verhaegen, J., Laga, P., Westerhoff, W. & Munsterman, D. 2014. The implications of K‐Ar glauconite dating of the Diest Formation on the paleogeography of the Upper Miocene in Belgium. Geologica Belgica, 17, 161–174.
48Vandenberghe, N., Wouters, L., Schiltz, M., Beerten, K., Berwouts, I., Vos, K., Houthuys, R., Deckers, J., Louwye, S., Laga, P., Verhaegen, J., Adriaens, R. & Dusar, M., 2020. The Kasterlee Formation and its relation with the Diest and Mol Formations in the Belgian Campine. Geologica Belgica, 23/3-4, this volume. https://doi.org/10.20341/gb.2020.014
49Verhaegen, J., 2020. Stratigraphic discriminatory potential of heavy mineral analysis for the Neogene sediments of Belgium. Geologica Belgica, 23/3-4, this volume. https://doi.org/10.20341/gb.2020.003
50Vernes, R.W., Deckers, J., Bakker, M.A.J., Bogemans, F., De Ceukelaire, M., Doornenbal, J.C., den Dulk, M., Dusar, M., Van Haren, T.F.M., Heyvaert, V.M.A., Kiden, P., Kruisselbrink, A.F., Lanckacker, T., Menkovic, A., Meyvis, B., Munsterman, D.K., Reindersma, R., Rombaut, B., ten Veen, J.H., van de Ven, T.J.M., Walstra, J. & Witmans, N., 2018. Geologisch en hydrogeologisch 3D model van het Cenozoïcum van de Belgisch-Nederlandse grensstreek van Midden-Brabant / De Kempen (H3O – De Kempen). TNO-rapport TNO 2017 R11261 – VITO 2017/RMA/R/1348, 109 p.
51Westerhoff, W., 2003. Waubach Member. Stratigraphic Nomenclature of the Netherlands, TNO – Geological Survey of the Netherlands. http://acc.dinoloket.nl/en/stratigraphic-nomenclature/waubach-member, accessed on 30/05/2020.
52Westerhoff, W., 2009. Stratigraphy and sedimentary evolution: the lower Rhine-Meuse system during the Late Pliocene and Early Pleistocene (southern North Sea basin). TNO Geological Survey of the Netherlands, 168 p.
53Wong, T., de Lugt, I., Kuhlmann, G. & Overeem, I., 2007. Tertiary. In Wong, T., Batjes, D. & de Jager, J. (eds), Geology of the Netherlands. Royal Netherlands Academy of Arts and Sciences, Amsterdam, 151–171.
54Williams, G.L., Fensome, R.A. & MacRae, R.A., 2017. The Lentin and Williams index of fossil dinoflagellates 2017 edition. American Association of Stratigraphic Palynologists Contributions Series, 48, 1097 p.
55Manuscript received 05.06.2020, accepted in revised form 10.08.2020, available online 16.10.2020.
Om dit artikel te citeren:
