
The Pliocene–Pleistocene transition in the subsurface of the Dutch-Belgian border region: insights from borehole Huijbergen
Abstract
Cross-border correlations of the Pliocene–Pleistocene successions in the southern Netherlands and northernmost Belgium are problematic, because biostratigraphic markers are often lacking. Correlation is further hampered by the poor age constraints of the Belgian Merksplas Formation. To address these issues, sedimentary, mollusc and dinoflagellate cyst analyses are combined to characterise the lithostratigraphic units in the Huijbergen borehole (The Netherlands) and to provide age estimates. Subsequently, the Huijbergen borehole was correlated with nearby boreholes in Essen and Kalmthout (Belgium). The Piacenzian intervals of the Dutch Oosterhout Formation can be correlated with the Belgian Lillo Formation, with the latter’s threefold borehole log signature appearing virtually continuous across the border between both countries. The Dutch Maassluis and Waalre formations are correlated with the shell-bearing lower part and the unfossiliferous higher part of the Merksplas Formation respectively. Although dinocysts are not age-diagnostic for the Maassluis and Waalre formations in borehole Huijbergen, characteristic interglacial marine shells provide a Gelasian age assessment for the Maassluis Formation. By correlation, this age estimate can also be applied to the lower part of the Merksplas Formation, thereby elucidating the Pliocene–Pleistocene transition near the Dutch-Belgian border.
1. Introduction
1The marine coastal Piacenzian–Gelasian succession in the subsurface of the Dutch-Belgian border region is poorly understood. In the Netherlands, this succession comprises the Oosterhout, Maassluis and Waalre formations, whereas in northern Belgium this interval is made up of the Lillo and Merksplas formations. The correlation between the Pliocene shell-rich, fine-grained, slightly glauconitic sand of the Dutch Oosterhout Formation and Belgian Lillo Formation has been well established (Doppert et al., 1979; Slupik et al., 2007; Wijnker et al., 2008; Munsterman et al., 2019; Wesselingh et al., 2020). However, the correlation of the overlying coarsening upward interval of the Maassluis Formation and the coarse-grained Waalre Formation with the Belgian Merksplas Formation remains unresolved. Moreover, the age of the Merksplas Formation is still disputed, whether it concerns a Pliocene or a Pleistocene unit (e.g. Buffel et al., 2001; Gullentops et al., 2001; Louwye et al., 2020). The Dutch Pliocene–Pleistocene successions have been the subject of several studies and several age indications are available (Meijer et al., 2006; Slupik et al., 2007; Noorbergen et al., 2015; Dearing Crampton-Flood et al., 2018, 2020). An attempt was made to correlate the abovementioned Dutch and Belgian units in cross-border projects (Vernes et al., 2018), but many questions remain due to the lack of biostratigraphic data in this area. In the present study, we document the Upper Pliocene and Lower Pleistocene succession in a borehole in the Dutch border village of Huijbergen (Fig. 1) by combining data from lithology, gamma-ray (GR) logs, molluscs and dinocysts, and compare the studied succession to more complete Lower Pleistocene successions further north. The Huijbergen borehole is then used to extend the correlation south of the border into northern Belgium. The aim of this study is to establish precise cross-border correlations of the Pliocene–Pleistocene units near Huijbergen, Essen and Kalmthout, and to obtain relative age estimates for the Belgian Merksplas Formation in the northernmost Campine area.
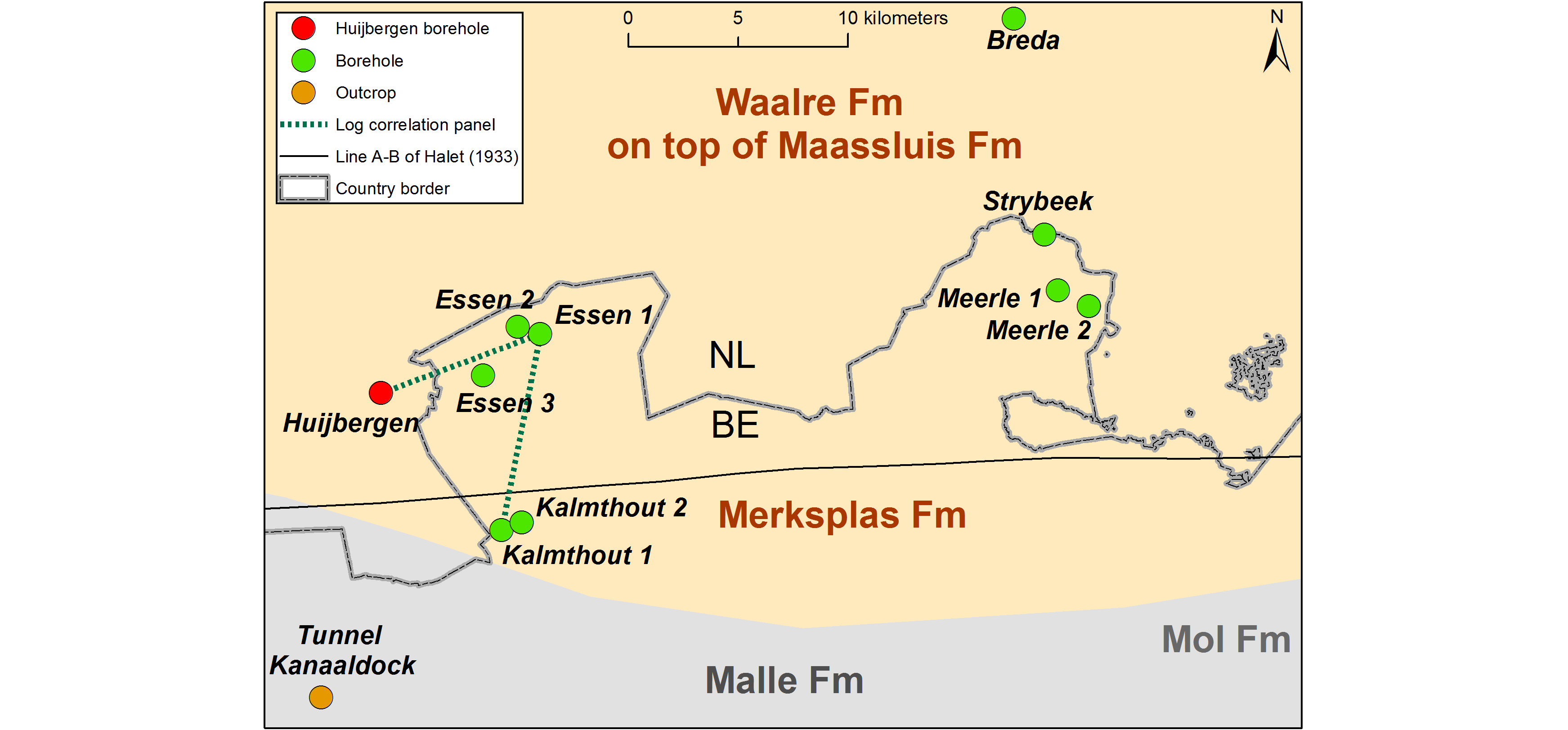
Figure 1. Location of the study area near the Dutch/Belgian border with the geographic extent of the main stratigraphic units and the relevant boreholes/outcrops (Table 1). The correlation panel of Fig. 4 is also indicated.
2. Geological background
2Pliocene–Pleistocene successions are present in the subsurface of northern Belgium (Flemish region), extending and thickening northwards into the Netherlands (Vinken, 1988; Westerhoff, 2009; Vernes et al., 2018). The Pliocene Lillo Formation is well studied in temporary outcrops in the Antwerp harbour area (e.g. de Heinzelin de Braucourt, 1950, 1955a; Deckers et al., 2020; Louwye et al., 2020). It was defined by De Meuter & Laga (1976) as “grey, grey-brown and light grey-brown shelly sand, clayey in the lower part and with several shell layers; in the upper part a gradual decrease of the clay content and the thick shell layers; in the uppermost part a gradual disappearance of the shells”. The Lillo Formation is slightly glauconitic and comprises the late Zanclean Luchtbal Member and the Piacenzian Oorderen, Kruisschans, Merksem and Zandvliet members (Louwye et al., 2020). Together with the underlying Zanclean Kattendijk Formation, the Lillo Formation can be correlated with the Dutch Oosterhout Formation (Doppert et al., 1979; Slupik et al., 2007; Wijnker et al., 2008; Munsterman et al., 2019; Wesselingh et al., 2020), which has similar lithological characteristics.
3North and northeast of the Antwerp harbour area, the Lillo Formation becomes covered by pale grey medium- to very coarse-grained quartz sand of the Merksplas Formation which can contain reworked shell layers at the base (Gulinck, 1962; Buffel et al., 2001). This unit was for the first time formally described by Gulinck (1962) as Sables grossiers de Merksplas. The latter author referred to the observations of Halet (1933), who attributed an Icenian age to the shell grit of this unit. The Icenian is a nowadays obsolete regional stage covering the Early Pleistocene (Gelasian) Norwich, Chillesford and Weybourne Crags in the United Kingdom (Harmer, 1920; Norton, 1967; Preece et al., 2020) and the Dutch Maassluis Formation (Doppert et al., 1975). No glauconite was mentioned by Halet (1933) from this shelly sand in boreholes near Essen and Meerle. Halet (1933) also noted that the “Icenian” shells disappeared southward from Essen, while the lithology of the sand remained similar. This probably corresponds to two facies types mentioned by Bogemans & Lanckacker (2014): when both are present, a non-calcareous sand facies with some thin silt/clay intercalations overlies a calcareous sand facies with few fine siliciclastic intercalations and shells. The upper part of the Merksplas Formation may contain some glauconite, clay laminae and wood fragments (Louwye et al., 2020). The palynology of the organic material was analysed by Vanhoorne (1962), who attributed it to the Upper Reuverian (Pliocene) due to its similarity with the lignite in the Mol Formation. Recently, Al-Silwadi (2017) suggested a mid- to late Pliocene age of the Merksplas Formation based on dinoflagellate cysts studied in the Rees borehole. However, the studied interval was before attributed to the Vosselaar Member (Malle Formation) by Buffel et al. (2001), casting doubt on its attribution to the Merksplas Formation. The Merksplas Formation is mostly covered by the Pleistocene Brasschaat Member (Malle Formation), but a precise distinction between both units can often not be made (Louwye et al., 2020).
4Halet (1933) already noted that coarse sand with “Icenian” shells also occurred across the Dutch border in Breda. The “Icenian deposits” of the Netherlands (e.g. Pannekoek, 1956; Zonneveld, 1958) are nowadays considered to be part of the Maassluis Formation (Doppert et al., 1975; Slupik et al., 2007). The Maassluis Formation was described by the former as marine, coarse to fine-grained sand with shells and intercalated sandy clay layers or clay lenses. Sporadically, peat remains, wood fragments and a little glauconite can occur (Vernes et al., 2018). The Maassluis Formation covers the largest part of the Netherlands and can reach a maximum thickness of 250 m towards the northwest (TNO-GSN, 2022a). In the south, clay intercalations are less common and the unit is dominated by sand (Vernes et al., 2018). The Maassluis Formation lies almost everywhere on top of the Oosterhout Formation. The boundary between both formations is not always clear, due to massive reworking (e.g. borehole Schelphoek; Slupik et al., 2007). Doppert et al. (1975) indicated that the Maassluis Formation has an Early Pleistocene age (Praetiglian–Tiglian). Meijer et al. (2006) presented a local zonation (molluscs, pollen and dinocysts included) for the Praetiglian Stage of the Maassluis Formation in the Noordwijk borehole, suggesting that cold and warm units within the Praetiglian could be correlated with Marine Isotope Stages (MIS) 100-96. Later, the stable isotope stratigraphy of a more than 100 m thick sequence within this Noordwijk borehole was studied by Noorbergen et al. (2015). These authors tuned their record to MIS 100-94 using the characteristic δ18O signature and complementary biostratigraphic data, slightly adjusting the attributions from Meijer et al. (2006) and Kuhlmann et al. (2006). The sedimentary facies and molluscs of seven sequences of the Maassluis Formation were studied in detail by Slupik et al. (2007) in borehole Schelphoek. Also in this borehole, the three lowermost sequences were assigned to the MIS 100, 98 and 96. The characteristic faunas described by e.g. Slupik et al. (2007) thus fall within the Gelasian, as its basal GSSP in Monte San Nicola (Sicily, Italy) corresponds to MIS 103 (Gibbard & Head, 2020). Only recently, the lectostratotype section for the Maassluis Formation in well B37D0228 (Maassluis), interval 103–206 m (cf. TNO-GSN, 2022a), was analysed for dinoflagellate cysts (Munsterman, 2021). Here, the base of the Maassluis Formation at 206 m is interpreted to coincide with MIS 100 (Gelasian) and is located just above the Pliocene–Quaternary boundary. Hence, the Maassluis Formation in the Netherlands is of an Early Pleistocene age.
5In the study area near the Belgian border, the Maassluis Formation is interfingering with the WA-2 subunit of the Waalre Formation, showing increasing marine influence west of the Roer Valley Graben (Westerhoff, 2009, fig. 2.10). The common occurrence of marine shells distinguishes the Maassluis Formation from the Waalre Formation. The latter consists of greyish-white, very fine to very coarse grained (micaceous) sand that may contain gravelly lag deposits and furthermore contains clay beds and laminae, silty to sandy, with sporadic peat and siderite (TNO-GSN, 2022b). The Waalre Formation represents Rhine-Meuse fluvial and coastal plain deposits and covers almost the entire Netherlands (Westerhoff, 2009; TNO-GSN, 2022b). Three subunits were recognised in the Waalre Formation (WA-1 – WA-3); the entire WA-2 subunit has an Early Pleistocene age (Westerhoff, 2009).
3. Material & methods
3.1. Borehole data
6Borehole Huijbergen (TNO-GSN B49G0204) is located in the SW Netherlands (WGS84 coordinates: 51.433338, 4.360121, surface at 18.2 m Normaal Amsterdams Peil), ca. 2.5 km west of the Belgian border (Fig. 1). It is a pulse borehole that was executed on January 10, 1990 and from which one-meter samples were obtained and stored at TNO-Geological Survey of the Netherlands (GSN). Each sample contains mixed sediment from an entire meter interval. The borehole penetrates Quaternary and Neogene sediments. Westerhoff (2009, fig. 2.5) provided a gamma-ray log next to data on grain size, the presence of mica flakes, glauconite content, CaCO3 %, organic matter % and indications of the presence of shells. In this study, we add additional lithological observations, dinocyst and mollusc analyses (Fig. 2). According to the “Ondergrondgegevens” (subsurface data, subsection “Geologisch Booronderzoek”) made accessible by DINOloket (https://www.dinoloket.nl/ondergrondgegevens), the Waalre Formation occurs from 4.2–55 m below surface (b.s.), the Maassluis Formation is present between 55–59 m b.s. and the Oosterhout Formation between 59–77 m b.s. Earlier, Westerhoff (2009) interpreted the 55–59 m interval as the top of the Oosterhout Formation, but this was formally reinterpreted as Maassluis Formation in the hypostratotype sheet of the Huijbergen borehole (TNO-GSN, 2012). The Huijbergen borehole is considered a hypostratotype for the Waalre Formation; its lower part (ca. 24–55 m) was attributed by Westerhoff (2009) to the WA-2 subunit. Borehole depths mentioned in this study are always measured in meters below surface (m b.s.).
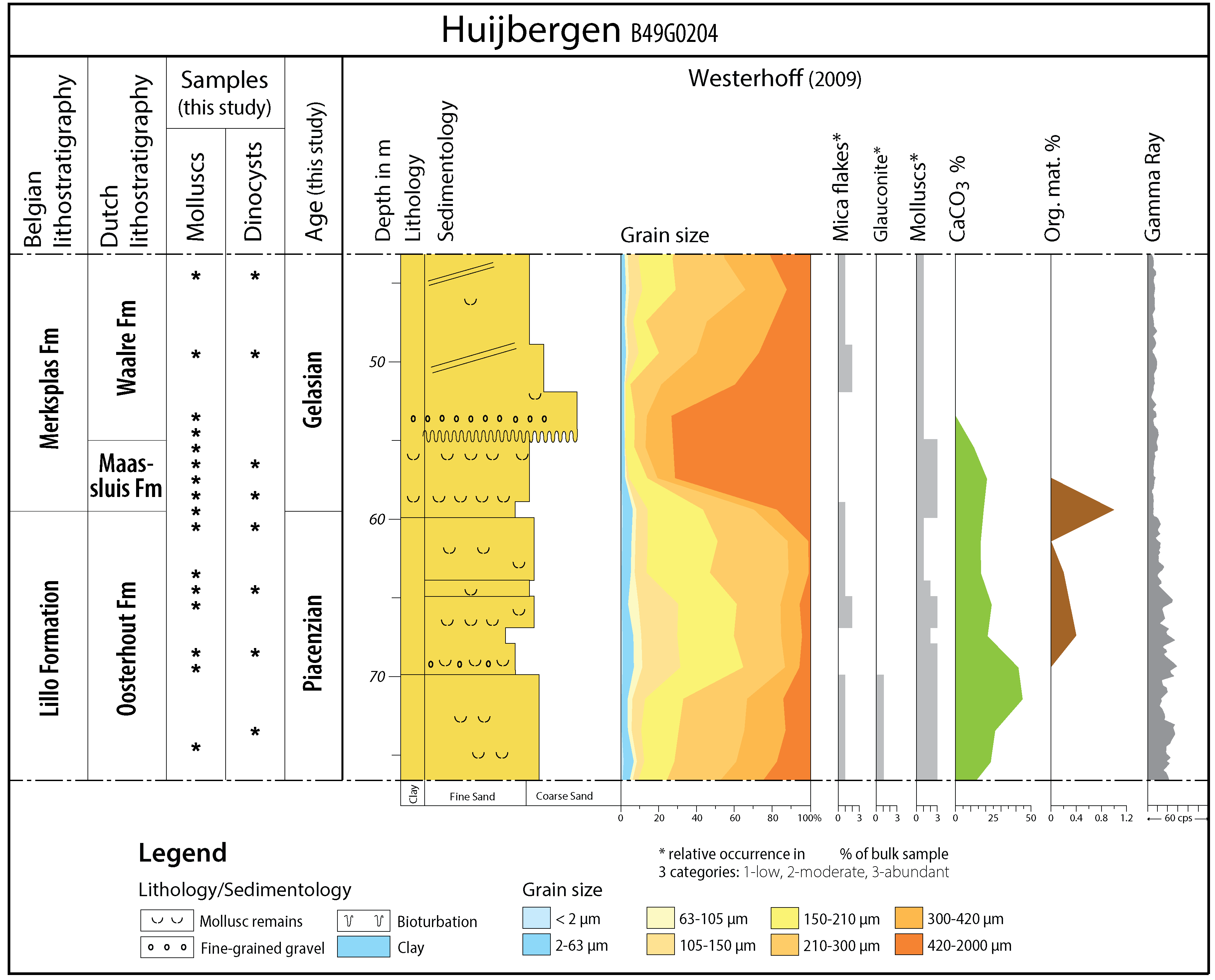
Figure 2. Borehole Huijbergen with lithostratigraphic interpretation (see 4.1.), dinocyst and mollusc sample locations and age assessments on the left side (this study). In the center and on the right side, a sketch of the original lithological description, results of grain-size analyses and data on mica flakes, glauconite content, relative abundance of molluscs, CaCO3 %, organic matter (%) and a gamma-ray log (data from Westerhoff, 2009).
3.2. Mollusc samples and analyses
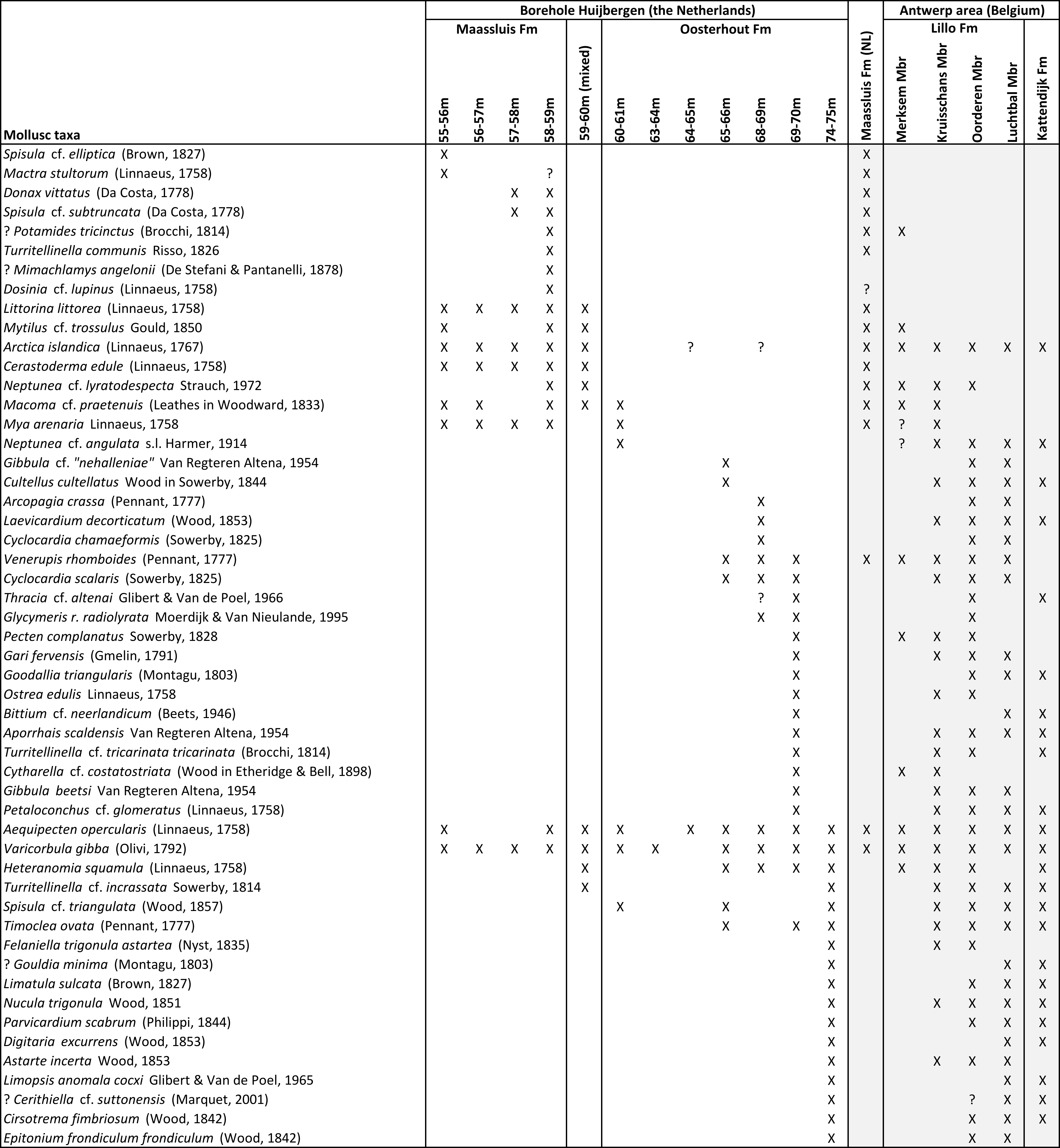
7Sixteen samples of sediment (each ca. 100–200 g) from the Oosterhout, Maassluis and Waalre formations in the Huijbergen borehole were analysed for molluscs (see Figs 2–3 for depth and stratigraphy). Samples were dry sieved with a mesh size of 1 mm. The obtained residues (Table 2) are stored in the Fossil Mollusc collections at Naturalis Biodiversity Center (Leiden, the Netherlands). Samples were inspected for preservation characteristics and sorted for fossil shells (Table 3). For the identification and/or stratigraphic distribution of molluscs we used Marquet (1993; 1998; 2001; 2002; 2004; 2005), Marquet & Landau (2006), Moerdijk et al. (2010), Vervoenen (1995), Vervoenen et al. (2000) and Wesselingh et al. (2020). Nomenclature follows Moerdijk et al. (2010), Pouwer & Wesselingh (2012), Wesselingh et al. (2012; 2014), Rijken & Pouwer (2014), Moerdijk & Janse (2015), Raad et al. (2016), Moerdijk et al. (2018), Menkhorst & Wesselingh (2018), Pouwer & Rijken (2022) and the World Register of Marine species (WoRMS) website. The attribution of the species formerly known as Turritella communis to Turritellinella tricarinata as proposed by the latter website is not followed here, as further research is required to elucidate the status of both species (viz. Moerdijk et al., 2018).
3.3. Dinoflagellate cysts samples and analyses
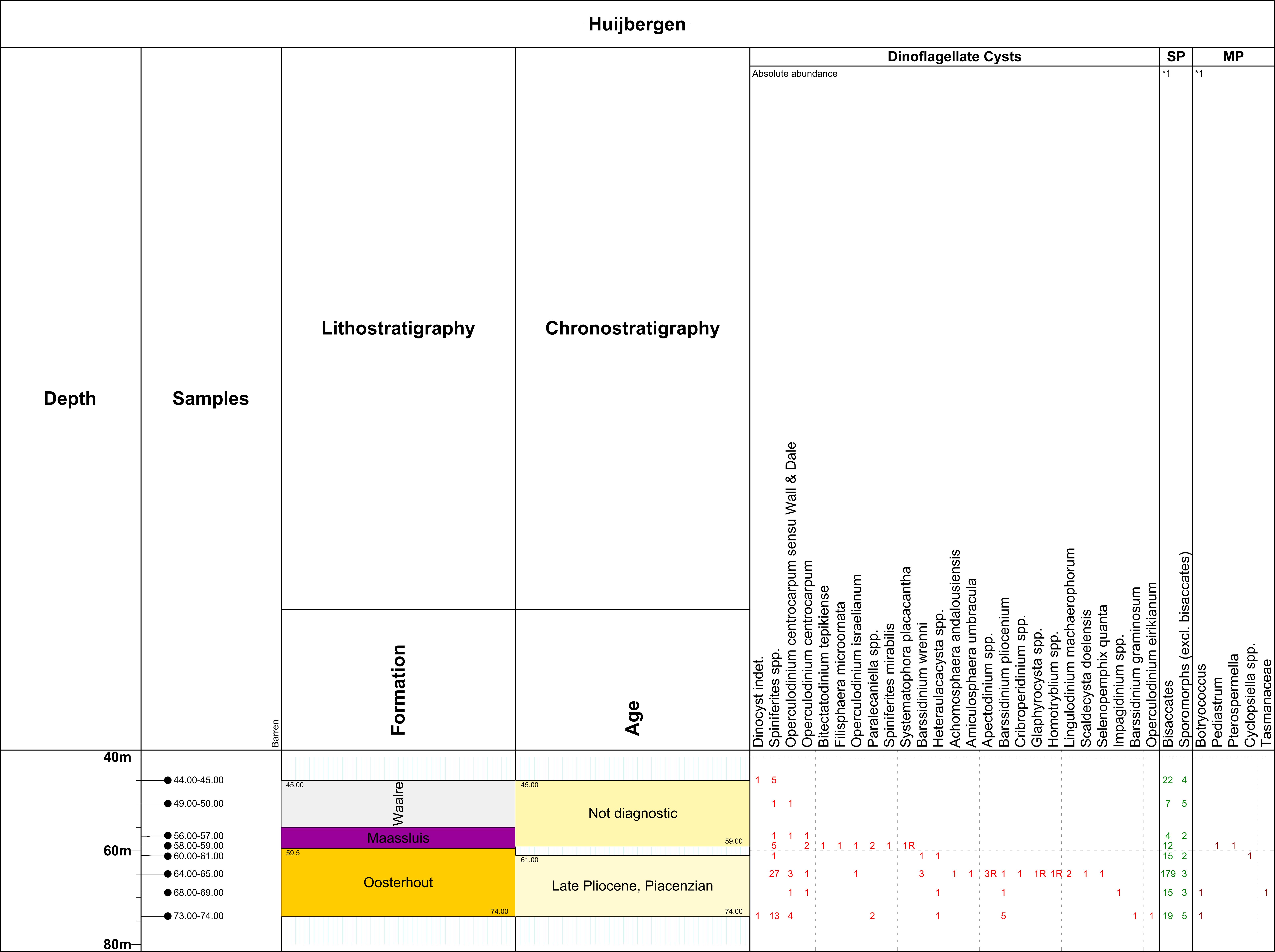
8Eight samples (each ca. 15–20 g) from the Oosterhout, Maassluis and Waalre formations in the Huijbergen borehole were analysed for dinocysts (Fig. 2). Standard palynological techniques, including HCl and HF digestion and 15 μm sieving, were applied following Janssen & Dammers (2008). No oxidation was used. The slides were mounted in glycerine jelly. One microscope slide per sample was counted until a minimum of 200 palynomorphs (spores, pollen and dinoflagellate cysts) had been identified (when present). The remainder of the slides were scanned for rare taxa. Miscellaneous fossils (e.g. green algae like Pediastrum, Botrycococcus) were also counted but kept outside the total sum of 200 specimens. The age interpretation is based on the Last Occurrence Datum (LOD) and First Occurrence Datum (FOD) of dinoflagellate cysts. Key references for the Pliocene–Pleistocene successions in the North Sea area are Dearing Crampton-Flood et al. (2020), De Schepper & Head (2008), De Schepper et al. (2009; 2015), Hennissen et al. (2017), Kuhlmann et al. (2006), Meijer et al. (2006) and Noorbergen et al. (2015). Dinoflagellate cyst taxonomy is according to that cited in Fensome et al. (2019). The slides are stored at TNO-Geological Survey of the Netherlands, in the archive of this borehole.
3.4. Regional correlation
9An overview of all sections mentioned in this study (both for correlation and their palaeontological content) is given in Table 1, with (digital) references to their original descriptions and interpretations. The Databank Ondergrond Vlaanderen (DOV) reference set can also be consulted online at https://www.dov.vlaanderen.be/data/opdracht/2023-033540.
10Louwye et al. (2020) performed correlations of the Belgian stratigraphic units in Belgian boreholes towards the Huijbergen borehole in the Netherlands. In this study, we use the northernmost Belgian boreholes to perform an updated correlation with the Huijbergen borehole. These Belgian boreholes are Essen 1 (BGD 001E0044, DOV B/1-1095, WGS84 coordinates: 51.4576, 4.4647) and Kalmthout 1 (BGD 006E0130, DOV kb7d6e-B155, WGS84 coordinates: 51.3772, 4.4391). Borehole (re)interpretations and correlations are based on existing borehole descriptions and log signatures.
Table 1. Overview of the boreholes together with the references to the archives of the geological surveys (BGD: Geological Survey of Belgium; DOV: Databank Ondergrond Vlaanderen; TNO–GSN: TNO–Geological Survey of the Netherlands).
|
Name |
TNO-GSN |
BGD |
DOV |
Reference |
|
Breda borehole |
B50B0296 |
/ |
/ |
Halet (1933) |
|
Essen 1 borehole |
/ |
Laga (1979) |
||
|
Essen 2 borehole |
/ |
Halet (1933) |
||
|
Essen 3 borehole |
/ |
Gulinck & Paepe (1969) |
||
|
Hank Borehole |
B44E0146 |
/ |
/ |
Dearing Crampton-Flood et al. (2018, 2020) |
|
Huijbergen borehole |
B49G0204 |
/ |
/ |
Westerhoff (2009) |
|
Kalmthout 1 borehole |
/ |
BGD (1980) |
||
|
Kalmthout 2 borehole |
/ |
Vanhoorne (1962) |
||
|
Maassluis borehole |
B37D0228 |
/ |
/ |
Munsterman (2021) |
|
Meerle 1 borehole |
/ |
Halet (1933) |
||
|
Meerle 2 borehole |
/ |
Laga & Hanssen (1976) |
||
|
Noordwijk borehole |
B30F0470 |
/ |
/ |
Noorbergen et al. (2015) |
|
Rees borehole |
/ |
Buffel et al. (2001) |
||
|
Schelphoek borehole |
B42G0022 |
/ |
/ |
Slupik et al. (2007) |
|
Strijbeek borehole |
/ |
Mourlon (1897) |
||
|
Tijsmanstunnel |
/ |
Laga (1972) |
4. Results
4.1. Lithostratigraphic units in borehole Huijbergen
11A coded lithological description is given by TNO-GSN (2012) for the Huijbergen borehole hypostratotype of the Waalre Formation. The lithostratigraphic interpretation by TNO-GSN (2012) is followed, only the boundary between the Maassluis and Oosterhout formations is slightly adjusted (59.5 m instead of 59 m b.s., see below). Descriptions of the sieve fractions above 1 mm are given for each sample in Table 2. Based on our observations of the samples, the different lithostratigraphic units in the studied interval (44–77 m b.s.) of borehole Huijbergen can be described as follows (Figs 2–3):
-
59.5–77 m b.s.: Oosterhout Formation - Characterised by grey to grey-green fine-grained quartz sand with variable but low amounts of glauconite and shells (Fig. 3). The gamma-ray signature shows medium values, probably related to the glauconite content, with higher values between 65–70 m, related to a relative increase in fine sand. Sample 59–60 m contains sediment and fossils characteristic of both the Oosterhout Formation and the overlying Maassluis Formation (see below) and may therefore be a mix of both units. Considering that the sample contains sediment from an entire one-meter interval, the actual boundary between both formations is likely located within this interval. For clarity, we use a depth of 59.5 m for the formation boundary throughout this work.
-
55–59.5 m b.s.: Maassluis Formation - Characterised by grey, very coarse grained quartz-rich sand with abundant worn shells and shell fragments (Figs 3–4). The gamma-ray values are distinctly lower than those of the underlying Oosterhout Formation, which is probably also partly related to the absence of glauconite.
-
44–55 m b.s.: Waalre Formation (WA-2 subunit) - Characterised by grey coarse-grained sand (Fig. 3). Only some dispersed and small decalcified shell fragments are present. The quartz shows a clear fining upward trend. In the lower part, some dispersed rounded greyish quartz pebbles are present. The gamma-ray values are similarly low as those of the underlying Maassluis Formation.
12An abrupt grain-size increase defines the boundary between the Oosterhout and Maassluis formations in borehole Huijbergen. Further north, the distinction between both formations often becomes more vague, due to finer grain sizes in the Maassluis Formation and massive reworking from the Oosterhout Formation (Slupik et al., 2007; Vernes et al., 2018). Next to coarser grain sizes, the disappearance of glauconite seems also a useful lithological criterion to distinguish the Maassluis Formation in Huijbergen. Although Westerhoff (2009) already indicated the absence of glauconite above 70 m, visual examination of the samples showed the presence of some glauconite until (mixed) sample 59–60 m. An upward decrease is observed in the studied part of the Oosterhout Formation. For instance, the glauconite content of sample 74–75 m is slightly higher than in the samples up to 65 m b.s. This decreases further to very low values in the upper 6 m of the Oosterhout Formation, presumably below the lower detection limit of Westerhoff’s analyses. Unfortunately, no new numerical analyses could be performed here. In general, the glauconite content of the upper part of the Oosterhout Formation is low, similar to that in the Belgian Lillo Formation.
Table 2. Description of the ≥1 mm fraction of the samples from the Huijbergen borehole.
|
Sample depth |
Description of the residue (≥1 mm) |
|
74–75 m |
Very little, fine shell grit. The shells all have the same whitish-yellowish creamy colour, the preservation ranges from extremely well-preserved to slightly abraded. The shells are often affected by bioerosion. Mainly small shell species, some Ditrupa fragments and echinid spines. |
|
69–70 m |
Abundant shell gravel, representing a varied fauna with mainly fragments of both thin and thicker shells (e.g. Ostrea edulis, Aequipecten opercularis, Aporrhais scaldensis). The shells are grey beige, with mixed types of conservation: bioerosion and slight abrasion are common. The surface is sometimes a little dissolved. Slightly clayey/silty sand sticks to the shells. Some Ditrupa fragments, echinoid spines, bryozoan fragments and barnacles are present. Some rounded quartz pebbles (<7 mm) and some black pebbles (<5 mm). |
|
68–69 m |
Abundant shell gravel, shell carbonate is slightly affected by both dissolution and abrasion. The colour of the shells is mainly creamy white, and a little blue grey. Large fragments of Aequipecten opercularis are abundant. Slightly clayey/silty sand sticks to the shells. Many barnacles, Ditrupa arietina, bryozoan fragments and large foraminifera. Some small pebbles. |
|
65–66 m |
Abundant fine shell gravel, fragments white to grey with a corroded surface. Strongly dominated by Varicorbula gibba. Fauna with low diversity. Rare fragments of the brachiopod Glottidia. Some very small red-brown clay pieces, often associated with Varicorbula gibba. |
|
64–65 m |
Large pieces of dried brown-red clay, with a few shell fragments enclosed. Only two abraded fragments of ?Arctica islandica and Aequipecten opercularis. |
|
63–64 m |
Large pieces of dried brown-red clay containing two decalcified fragments of Varicorbula gibba. |
|
60–61 m |
Small pieces of dried brown-red clay. Some worn Varicorbula gibba, one filled with clay. Rare shell gravel, consisting of mainly small and corroded fragments. One well-preserved small hinge fragment of a juvenile Mya arenaria. |
|
59–60 m |
Large pieces of dried brown-red clay, with a paired Varicorbula gibba enclosed. Some shell gravel is present with a mixed but mainly worn preservation. Moreover, some fragments show the imprints of coarse sand grains. |
|
58–59 m |
Very abundant shell gravel. Most shells are fragmented and show signs of abrasion/reworking and dissolution, the imprints of coarse quartz grains are often visible. The colour of the shells is predominantly white to light blue, but red-brown shells/fragments are also common. A varied assemblage of shell species is present including Arctica islandica, Mya arenaria and Littorina littorea. Barnacles, wood fragments, rare pebbles and coarse quartz grains occur. |
|
57–58 m |
Abundant rounded shell gravel, fewer large fragments than in the 58–59 m sample. Mixed colours. Shell surfaces with dissolution marks including sand grain imprints. Fauna similar but slightly less diverse than that of the underlying interval. |
|
56–57 m |
Abundant rounded shell gravel; shells and fragments with strongly dissolved surfaces including common sand grain imprints; abrasion common. Admixture of red-brown, grey and grey-blue colours. |
|
55–56 m |
Similar to 57–58 m, with fewer shell gravel and large fragments. Strong abrasion and dissolution of the shell surfaces. A decapod dactylus was recognised. |
|
54–55 m |
No shells, besides one unidentifiable decalcified splinter. Coarse quartz grains are abundant, slightly rounded but mostly still somewhat irregular, especially the larger grains. Rare large rounded quartz pebbles are also present (ca. 5 mm) |
|
53–54 m |
Idem to 54–55 m. Some rounded quartz pebbles (<6 mm) |
|
49–50 m |
Idem to 53–54 m. |
|
44–45 m |
No shells. Coarse quartz grains. |

Figure 3. Sediment samples of the Huijbergen borehole. (A) Sample 49–50 m (RGM.1365353) contains quartz grains and lacks calcareous fossils. It represents the Waalre Formation. (B) Samples 56–57 (RGM.1365354) and (C) 58–59 m (RGM.1365355) contain poorly sorted, mostly coarse-grained quartz sand with abundant, mostly abraded, shell fragments. Some claystone pebbles and gravel are found. This interval is attributed to the Maassluis Formation. (D) Sample 60–61 m (RGM.1365356) contains slightly oxidised quartz grains, (very) little glauconite, fine shell fragments and lithified brown-red clay nodules with small shell fragments. The sample is attributed to the Oosterhout Formation. (E) Samples 68–69 m (RGM.1365357) and (F) 74–75 m (RGM.1365358) are characterised by poorly sorted, fine-grained quartz sand rich in shells and shell fragments attributed to the Oosterhout Formation. Sediments in the Maassluis Formation samples are notably coarser grained than in the Oosterhout Formation samples, while they lack glauconite.
4.2. Molluscs in borehole Huijbergen
13Preservation and general characteristics of mollusc samples are provided in Table 2. A list of the identified species is given in Table 3. For each of the observed species, their known occurrences in Pliocene–Pleistocene lithostratigraphic units in Belgium and the Netherlands are detailed. Three associations are distinguished in the samples below 55 m b.s., differing in both species composition and preservation.
Table 3. Mollusc occurrences in borehole Huijbergen. The stratigraphical distributions of the different species in the Kattendijk Formation, Lillo Formation (Antwerp area) and the Maassluis Formation (the Netherlands) are listed.
4.2.1. Association I (65–75 m b.s.)
14Association I contains a relatively diverse fauna (Table 3). Sample 74–75 m contains 13 bivalve and four gastropod species. Taxa are known from the Lillo and Kattendijk formations of the Antwerp area. Most taxa indicate that the fauna corresponds to that of the Piacenzian Oorderen Member, but a number of taxa are known exclusively from the Zanclean Luchtbal Member and the Kattendijk Formation. The absence of Palliolum gerardi (Nyst, 1835) makes correlation with the Luchtbal Member of the Lillo Formation very unlikely, as it is the dominant species in this member (Wesselingh et al., 2020). The two samples representing the 68–70 m interval contain 16 bivalve and six gastropod species. Indicative species include Glycymeris radiolyrata radiolyrata, Ostrea edulis, Gari fervensis and Aporrhais scaldensis. Overall, the fauna shows large overlap with the fauna of the Piacenzian Oorderen Member (Lillo Formation). Sample 65–66 m contains only eight bivalve and one gastropod species. The fauna is dominated by Varicorbula gibba and contains common Venerupis rhomboides. It resembles the faunas reported from the Piacenzian Kruisschans Member in the Antwerp area (Marquet, 2004). Association I likely represents late Pliocene shallow marine (shoreface to offshore) warm-temperate settings. Abrasion seen on the shells implies physical reworking, but the ecological consistency of species implies local reworking in a high-energy environment. The fauna in sample 65–66 m is dominated by Varicorbula gibba. Varicorbula-dominated associations are well known from ecologically stressed seafloors (Aleffi et al., 2000) and are found today in the southern North Sea in fine-grained areas below the storm wave base, where summer hypoxia may occur.
4.2.2. Association II (60–65 m b.s.)
15Association II samples yield relatively little material and few species (Table 3) that provide little biostratigraphic information. Preservation is often poor due to prominent wear. Only Mya arenaria in sample 60–61 m has a well-preserved surface structure, other shells are abraded. Macoma cf. praetenuis appears for the first time in this association. The few species in assemblage II occur in the Piacenzian Kruisschans and Merksem members (Lillo Formation) in Antwerp and in the Gelasian Maassluis Formation of the Netherlands. The absence of Spisula inaequilatera is conspicuous, as it is dominating the Merksem faunas near Antwerp.
4.2.3. Association III (55–60 m b.s.)
16The samples are very rich in shells and especially fragments with a varied but mostly weathered and worn preservation. Only the 59–60 m sample (probably a mix of the under- and overlying units) contained little fine-grained shell grit. The overall fauna of Association III is dominated by species that are very characteristic of the Gelasian Maassluis and Norwich Crag formations of the southern North Sea basin (e.g. Littorina littorea, Cerastoderma edule, Mytilus cf. trossulus, Turitellinella communis, Spisula subtruncta, Donax vittatus) (Spaink, 1975; Boele, 2001; Meijer et al., 2006 (and references therein); Slupik et al., 2007). The recorded fauna differs considerably from the well-studied associations of the Pliocene Lillo Formation (Table 3). Only some reworked Neogene species were encountered, including one strongly worn fragment of a Pliocene Turitellinella incrassata in sample 59–60 m. A fragment of Mimachlamys angelonii in sample 58–59 m must have been reworked from Miocene deposits. The strong abrasion shows that most shells have been transported (Fig. 3). The species are marine and indicate temperate (interglacial) climatic conditions. Shallow depths are probable: Mya arenaria usually occurs above 20 m water depth, while Littorina littorea is a littoral species (Hayward & Ryland, 2017).
4.3. Dinocyst analyses of borehole Huijbergen
17Dinocyst, spore and pollen (palynomorph) data are shown in Table 4. The palynomorph yield is low, with the exception of sample 64–65 m. Age diagnostic dinoflagellate cyst species are only present in the interval 60–74 m. In this interval, typical palynomorphs were encountered as described from the stratotype of the Lillo Formation (Tunnel-Canal dock = Tijsmanstunnel section, Fig. 1; De Schepper et al., 2009). The last occurrence datum (LOD) of Barssidinium wrennii at 60–61 m shows that deposits at this depth and below have a Piacenzian or older age (Kuhlmann et al., 2006; De Schepper et al., 2009). The occurrence of B. pliocenicum between 64–74 m and B. graminosum in sample 73–74 m confirms a Piacenzian or older age. The genus Heteraulacacysta, also roughly indicative for this LOD, is recorded between 60–74 m b.s. The first occurrence datum (FOD) of dinoflagellate cyst Scaldecysta doelensis at sample depth 64–65 m is stratigraphically very significant. The stratigraphic range of this taxon is restricted to the upper Oorderen, Kruisschans and Merksem members (Piacenzian) of the Lillo Formation (De Schepper et al., 2009). The very rich 64–65 m sample is dominated by bisaccate pollen. De Schepper et al. (2009) noted similar dominance for the upper Oorderen, Kruisschans and Merksem members. Between 60–74 m, a marginal to shallow marine palaeoenvironment is inferred from increasingly diverse marine dinoflagellate cysts, including rare brackish algae (Botryococcus sp.). Based on the palynofacies in the overlying 44–59 m interval, continental to marginal marine conditions are interpreted.
Table 4. Results of the palynological analysis of the Huijbergen borehole (44–74 m interval).
5. Lithostratigraphic (re)interpretations and cross-border correlations
5.1. Reinterpretation of the Essen 1 borehole
18Borehole Essen 1 (BGD 001E0044, DOV B/1-1095) was described in detail and interpreted by Laga (1979). The latter interpreted the Merksplas Formation at 46–66 m b.s. and the Lillo Formation at 66–84.6 m b.s. A detailed lithological description was given (translated from Dutch):
-
46–54 m: “light grey and rusty coloured coarse angular quartz sand”
-
54–64 m: “the same sand, grey and with a little shell grit”
-
64–66 m: “idem, with a lot of shell grit and a glauconitic admixture”
-
66–70 m: “grey-brownish medium fine-grained glauconitic sand with a lot of shell grit”.
19The interpretations by Laga (1979) were reproduced in the correlation panel of Louwye et al. (2020, fig. 3). We follow the proposed lithological interpretation by the latter with the exception of the boundary between the Lillo and Merksplas formations (see below).
20To distinguish the Merksplas Formation, we maintain the original definition by Gulinck (1962), who mentioned coarse sand with clusters of reworked shells at the base. To characterize these shells, Gulinck (1962) explicitly referred to Halet (1933). The latter studied a borehole one km northwest of the Essen 1 borehole, here referred to as borehole Essen 2 (BGD 001E0033, DOV kb1d1e-B1). In that borehole, Halet (1933) identified a lithological boundary at 60 m b.s. Below, dark grey glauconitic sand was interpreted as Scaldisien (corresponding to the Lillo Formation; Laga et al., 2006). Above, coarse quartz sand with reworked shells of the “Icenian” was described, referred to by Gulinck as Sables grossiers de Merksplas. When these criteria are applied to Laga’s (1979) description of borehole Essen 1, the glauconitic interval with shells between 64–66 m would be best interpreted as the top of the Lillo Formation instead of the Merksplas Formation. Indeed, the gamma-ray values of the Lillo Formation remain high up to a depth of 64 m. Above 64 m, they sharply decline and resistivity values increase, indicative of a decreased glauconite content and/or increased grain sizes in the Merksplas Formation. This is similar to the boundary between the Oosterhout and Maassluis formations in the Huijbergen borehole. The overlying 54–64 m interval consists of coarse quartz sand with some shell grit and is attributed here to the lower part of the Merksplas Formation. This interval in borehole Essen 1 is about the same thickness and most probably corresponds to the shell-bearing interval (49–60 m) of borehole Essen 2.
5.2. (Re)interpretation of the Kalmthout 1 borehole
21The original interpretation (BGD, 1980) of the Kalmthout 1 borehole (BGD 006E0130, DOV kb7d6e-B155) positioned the boundary of the Merksplas and Lillo formations at 40 m below surface. Louwye et al. (2020, fig. 3) located this boundary a little higher (between 37–38 m) corresponding to an abrupt upward decrease in gamma-ray values. In the original description, the 36–40 m interval corresponds to “reduced, glauconitic, moderately clayey sand with some gravel” (translated from Dutch). In contrast, no glauconite is mentioned in the intervals above 36 m. Analogous to our reinterpretation of borehole Essen 1, we include the glauconitic intervals in the Lillo Formation. In contrast to borehole Essen 1, shells are lacking in the lower part of the Merksplas Formation in the Kalmthout 1 borehole. A small mismatch exists between the lithological boundary in the description of this counter-flush borehole and the transition observed on the GR log. However, this discrepancy may have been caused by the low sampling resolution (every 4 m): the transition on the log falls within the 36–40 m sampling interval.
5.3. Correlation of the Oosterhout and Lillo formations
22Both the Dutch Oosterhout Formation and the Belgian Lillo Formation comprise shelly, relatively fine-grained sand with low but variable amounts of glauconite and moderate gamma-ray values. Louwye et al. (2020) identified a threefold geophysical borehole log subdivision of the Lillo Formation in the Essen 1 and Kalmthout 1 boreholes, which they referred to from bottom to top as Li-A, Li-B and Li-C, in accordance with the geotechnical study of the Lillo Formation in the Port of Antwerp by Deckers et al. (2020). In the latter study, Li-A is defined as a sandy unit corresponding to the larger part of the Oorderen Member, while Li-B represents the more clayey sediments of the Kruisschans Member and/or the uppermost part of the Oorderen Member. The uppermost unit Li-C is again a sandy unit comprising the Merksem Member, but locally also comprising sandy upper parts of the Kruisschans Member. This threefold subdivision was also recognised by Louwye et al. (2020, fig. 3) in the Oosterhout Formation of the Huijbergen borehole; the three geophysical units Li-A, Li-B and Li-C were interpreted in the 70–77 m, 65–70 m, 55–65 m b.s. intervals respectively. However, we locate Li-C in the 59.5–65 m interval (see before), as the 55–59.5 m interval belongs to the Maassluis Formation due to its distinct lithology and abruptly lowered gamma-ray values (see 4.1.). The correlation with boreholes Essen 1 and Kalmthout 1 is given in Figure 4. Our analyses provide the following correlation between the Oosterhout Formation in Huijbergen and the units of the Belgian Lillo Formation:
-
Li-A (70–77 m b.s.): Lithology, mollusc and dinocyst species are consistent with the Piacenzian Oorderen Member in the Antwerp area.
-
Li-B (65–70 m b.s.): Lithology, mollusc and dinocyst species are consistent with the clayey part of the Oorderen Member and the Kruisschans Member in the Antwerp area.
-
Li-C (59.5–65 m b.s.): Lithology (fine slightly glauconitic sand with some shells and brown-red clay pieces) and dinocyst species are consistent with the Piacenzian Merksem Member of the Lillo Formation. This also largely applies to the molluscs, with the exception of the absence of Spisula inaequilatera. Only in mixed interval sample 59–60 m, a Maassluis Formation type of mollusc fauna was found in the sparse shell grit present, with some species absent in the Piacenzian Lillo/Oosterhout formations. In contrast, the dominant lithology of this sample corresponds more to the Oosterhout Formation and much less to the Maassluis Formation (TNO-GSN, 2012).
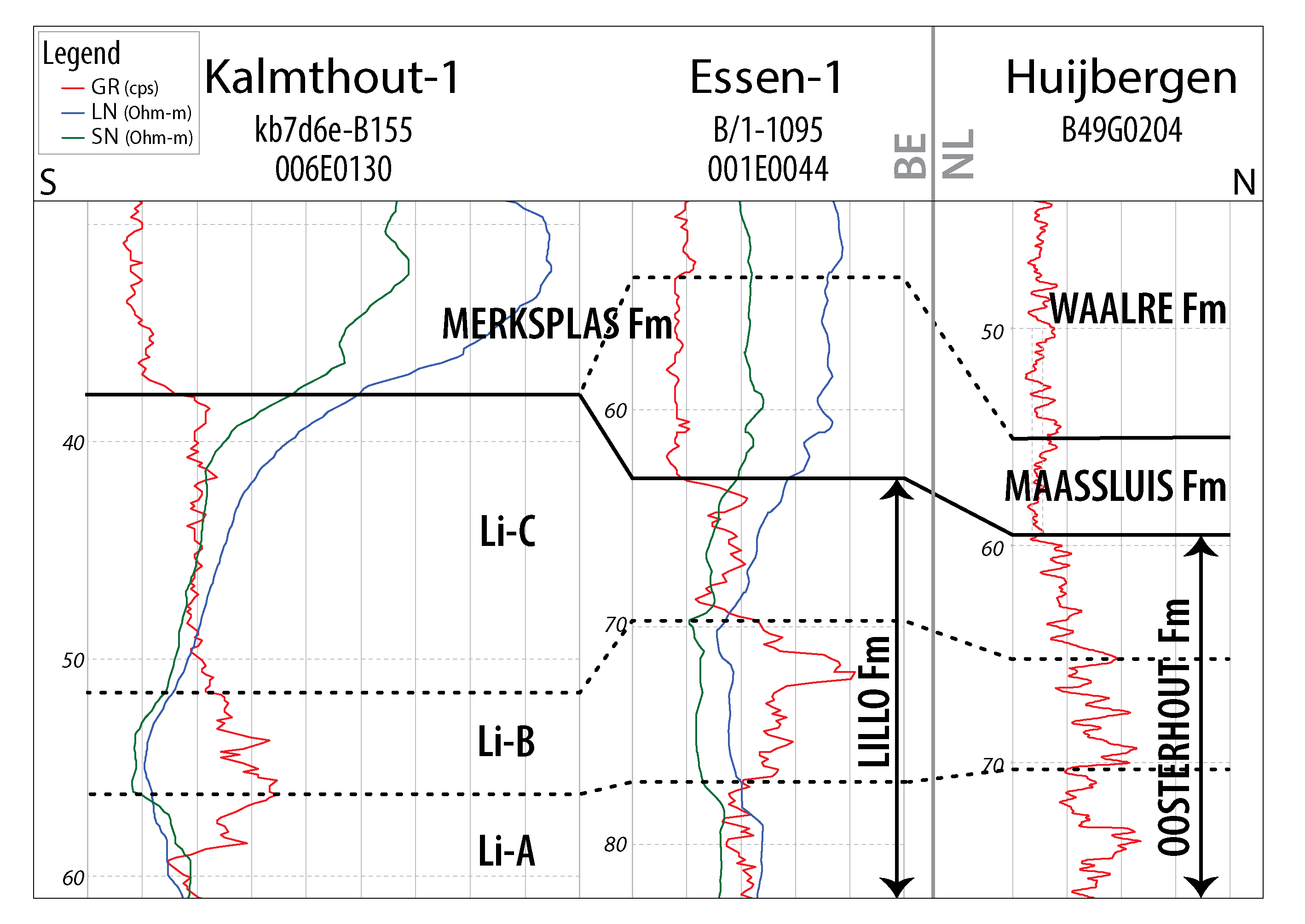
Figure 4. Borehole-log correlation panel for the Pliocene and Lower Pleistocene stratigraphic units along a N–S transect from northernmost Belgium across the border with the Netherlands (location of the profile: see Fig. 1). The original correlation panel of Louwye et al. (2020) is modified by this study. GR: gamma-ray. LN: long-normal (resistivity). SN: short-normal (resistivity).
5.4. Correlation of the Maassluis Formation with the shell-bearing lower part of the Belgian Merksplas Formation
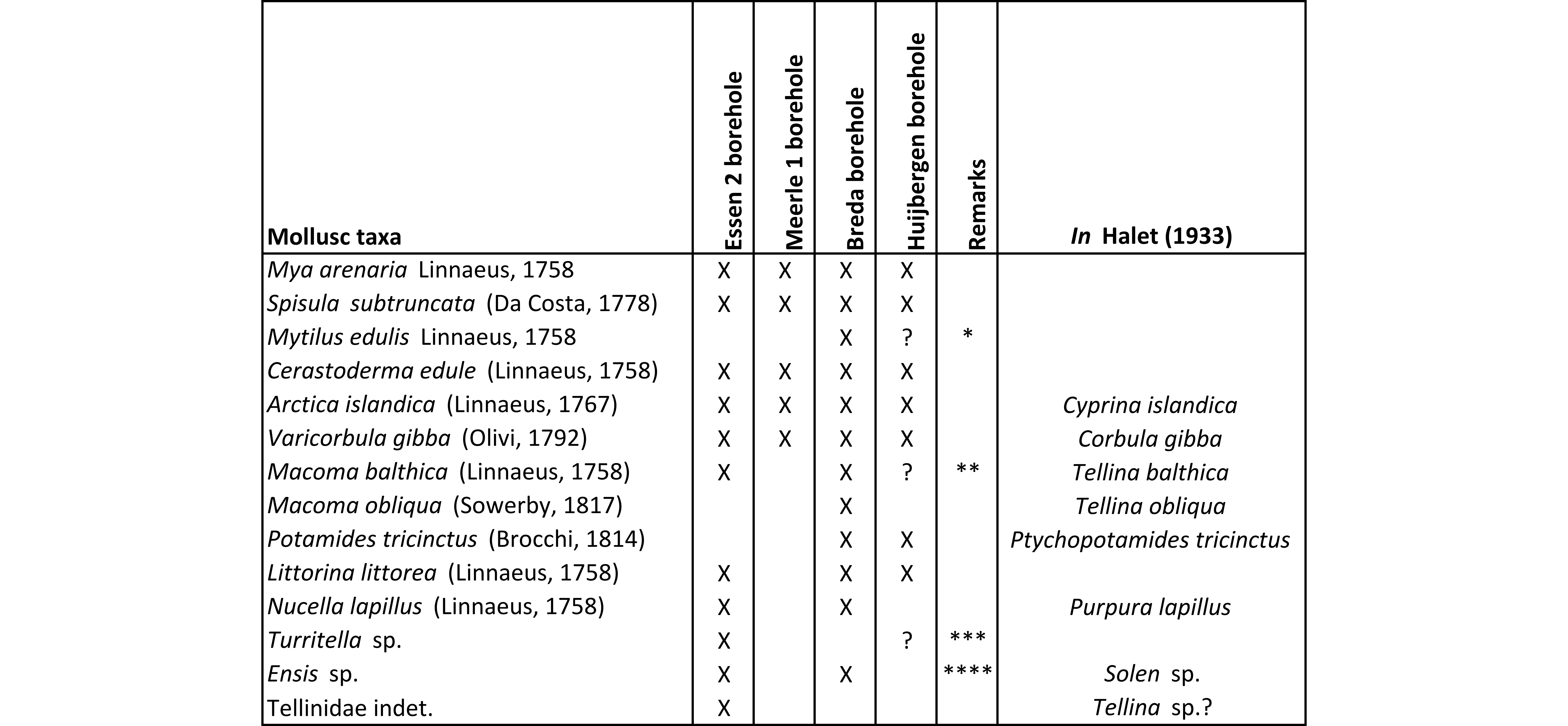
23Like the Maassluis Formation in the Huijbergen borehole, the lower part of the Merksplas Formation in the Essen area is coarse grained and contains reworked shell debris (Gulinck, 1962). Therefore, the Maassluis Formation as identified in Huijbergen (55–59.5 m b.s.) can be correlated with the shell-bearing lower part of the Merksplas Formation in the Essen 1 (54–64 m b.s.) and Essen 2 (49–60 m b.s.) boreholes. In the Kalmthout 1 borehole, shells are lacking in the base of the Merksplas Formation. Compared to the underlying glauconitic Oosterhout and Lillo formations, the Maassluis Formation and the lower part of the Merksplas Formation are very poor in, or lacking glauconite and are characterised by markedly lower, very uniform gamma-ray values (Figs 2 & 4). The Maassluis Formation shows a coarsening upward trend in the Huijbergen borehole, which is consistent with the increase in resistivity values in the Essen 1 borehole. Although the shell grit in borehole Essen 1 was not further characterised, Halet (1933) did study the “Icenian” fauna of the 49–60 m b.s. interval in the nearby borehole Essen 2 (Table 5). It is almost identical to that of the Maassluis Formation in Huijbergen (this study) (Table 5; Halet, 1933). These faunas strongly resemble Maassluis Formation faunas from other boreholes further north in the Netherlands (e.g. Slupik et al., 2007).
Table 5. Molluscs mentioned by Halet (1933) from the sables grossiers of the Merksplas Formation by locality (DOV & BGD references in Table 1). * Although possible, the occurrence of Mytilus edulis may be a misidentification of Mytilus trossulus, common in Huijbergen. ** Macoma balthica is only know from later Gelasian units, not from the study area (Preece et al., 2020). Hence this record likely represents misidentified M. praetenuis or M. obliqua. *** Probably Turritellinella communis. **** Probably Ensis, rather than Solen.
5.5. Correlation of the Waalre Formation with the upper part of the Merksplas Formation
24The Waalre Formation (WA-2 subunit) as identified in the Huijbergen borehole (44–55 m b.s.) correlates with the fossil-barren interval of the Belgian Merksplas Formation (46–54 m b.s.) in the Essen 1 borehole and, possibly, the entire Merksplas Formation in borehole Kalmthout 1 (Fig. 4). These intervals contain medium- to coarse-grained sand lacking shells. In the Huijbergen borehole, only a few unidentifiable, decalcified shell fragments were observed, precluding age assessments. Rare dinocyst species are chronostratigraphically indifferent and do not provide an age indication. In Huijbergen, the Waalre Formation shows a fining upward trend. The gamma-ray signature is almost identical to the underlying Maassluis Formation and the shell-bearing lower part of the Merksplas Formation in borehole Essen 1.
6. Discussion
6.1. Palaeontological aspects of the Piacenzian successions
25The biostratigraphic significance of Piacenzian molluscs must be further investigated in order to distinguish facies indicators and to improve the correlation of the various Piacenzian members of the Belgian Lillo Formation with intervals of the Dutch Oosterhout Formation. In borehole Huijbergen, several samples in the 69–75 m interval that are interpreted as Piacenzian contain some species known from older sediments in the Antwerp area. These include Digitaria excurrens, Limopsis anomala cockxi, Gouldia minima, Bittium cf. neerlandicum and Cerithiella cf. suttonensis, all known from the Zanclean Luchtbal Member. Yet the dominant Luchtbal fauna species, Palliolum gerardi, is entirely lacking within the Huijbergen borehole. To the north, the bioclastic Sprundel Member of the Oosterhout Formation is likely correlatable to the Oorderen Member in Antwerp. Facies-wise, however, the Sprundel Member (dominated by shelly bioclasts with abundant bryozoans) is very similar to the Belgian Luchtbal Member (and the eastern UK Coralline Crag Formation: Wesselingh et al., 2020). Similar to Huijbergen, Palliolum gerardi is absent in the Sprundel Member (unpublished data). Further research is ongoing on the Dutch Piacenzian successions that may provide detailed correlations with the Belgian units.
6.2. The Pliocene–Pleistocene boundary in the study area
26The precise location of the Pliocene–Pleistocene boundary in the southern North Sea Basin has been shown in boreholes in depocenters such as the offshore A15-03 borehole (Kuhlmann et al., 2006) and the onshore Hank borehole (TNO-GSN B44E0146; Dearing Crampton-Flood et al., 2018, 2020). Yet, their relationship with the exact boundary of the Oosterhout and Maassluis formations is currently subject to investigation, including the elucidation of formation boundary definitions. In general, the Oosterhout Formation in the Netherlands has been dated as Pliocene (e.g. Wijnker et al., 2008; Munsterman et al., 2019) and the Maassluis Formation as Gelasian (e.g. Meijer et al., 2006; Slupik et al., 2007; Noorbergen et al., 2015). The base of the Maassluis Formation in its lectostratotype section in Maassluis (TNO-GSN B37D0228) is interpreted to coincide with MIS 100 (Gelasian) (Munsterman, 2021). Within the study area, which was located in the southern basin margin, hiatuses and reworking are very commonly affecting the base of the Maassluis Formation (Slupik et al., 2007). Therefore, the Oosterhout/Maassluis and Lillo/Merksplas formation boundaries in the Dutch-Belgian border region likely represent a hiatus covering the Piacenzian–Gelasian boundary. The mollusc associations in the studied boreholes lack very cold faunal components (e.g. Serripes groenlandicus, Yoldia lanceolata, Portlandia arctica, Nuculana minuta) that are characteristic of cold phases in the basal (“Praetiglian”) part of the Maassluis Formation towards the north (Meijer et al., 2006; Slupik et al., 2007). Likely, marine conditions retreated during glacial time intervals from the proximally located study area. Our data do not differentiate whether the studied marine molluscs represent an interglacial highstand fauna from warm-temperate intervals within the Praetiglian or from the younger Tiglian (both Gelasian).
6.3. Palaeontological aspects of the Gelasian successions
27Lower Pleistocene shell-rich intervals in northern Campine boreholes (Essen, Meerle and Breda) were already documented by Halet (1933). These intervals are located in coarse quartz sand of the former “Icenian” regional stage that overlays the finer sand of the “Scaldisian” (Lillo and Oosterhout formations). Based on lithological characteristics, the sediments of the “Icenian” are now attributed to the Maassluis and Merksplas formations. A very similar fauna to that of boring Huijbergen was described by Halet (1933) from borehole Essen 2 and referred to as “Icenian”, including Mya arenaria, Arctica islandica, Spisula subtruncata, Cerastoderma edule and Littorina littorea. Nucella lapillus was recorded in borehole Essen 2 and not in borehole Huijbergen, but the species is known from the Maassluis Formation in borehole Schelphoek (Slupik et al., 2007). The same fauna (Table 5) was also observed in the Maassluis Formation (shells between 50–53, 65–68 & 78–90 m b.s.) of borehole Breda (TNO-GSN B50B0296) and the Merksplas Formation (40–60 m, most shells between 57.5–60 m b.s.) in borehole Meerle 1 (BGD 003W0057, DOV kb2d3w-B106) (Halet, 1933). Littorina littorea was not mentioned by Halet (1933) from borehole Meerle 1, but it was recognised by Laga & Hanssen (1976) between 54–57 m b.s. in the nearby borehole Meerle 2 (BGD 003W081, DOV kb2d3w-B81).
28Long after the work of Halet (1933), Littorina littorea was again noted in Essen by Gulinck & Paepe (1969) in borehole Essen 3 (BGD 006E0106; DOV kb7d6e-B132). These authors even separated the basal shell-rich unit of the Merksplas Formation as a distinct unit, which they referred to as “Old Pleistocene: estuarine facies of Strijbeek” (57–64 m). The samples were described as “pale grey, very coarse sand with shell grit (Littorina littorea)” (57–59 m) and “grey, very coarse sand, lots of shell grit (Littorina littorea)” (61–64 m) (translated from Dutch). The overlying parts of the Merksplas Formation (without shells) and younger units were referred to as “Old Pleistocene: continental facies”. It should be noted that also the age estimate for the underlying “Merksemian–Scaldisian” sediments (64–81 m) was “Old Pleistocene” at the time, while it is now attributed to the Piacenzian. The “estuarine facies of Strijbeek” with Littorina littorea is a reference to a Belgian borehole (BGD 003W001, DOV kb2d3w-B1) at the border near Strijbeek (NL), where the shell-rich basal part of the succession was previously noted by Mourlon (1897).
29The common occurrence of Littorina littorea (Fig. 5) in the Maassluis and Merksplas formations is a useful palaeontological indicator in the field, when describing and interpreting new drillings near the Dutch-Belgian border. In Belgium, it was neither reported from the Pliocene Lillo Formation by Glibert (1957, 1958) nor Marquet (1993; 1998) in their reference studies on Pliocene molluscs, nor it was found by private collectors in the Antwerp harbour (Van Nieulande & Vervoenen, pers. comm., 2023). Only de Heinzelin de Braucourt (1955b) mentions the species from the Belgian “Merkxemien” (obsolete regional stage corresponding to the Kruisschans and Merksem members), but no image nor repository was given. If present, the species must be very rare, given that the same author did not mention this species from his fieldwork in the “Merkxemien” of the Antwerp harbour (de Heinzelin de Braucourt, 1950, 1955a). The same applies to the UK: the oldest specimens (rare) were mentioned by Wood (1848) from the Newbourn and/or Butley members of the Red Crag Formation (latest Pliocene–earliest Pleistocene), but Littorina littorea only becomes abundant in the overlying Gelasian Norwich Crag Formation (ca. 2.2–2.45 Ma) (Leifsdóttir & Símonarson, 2002; Preece et al., 2020). According to Wood (1848), it is “the most characteristic shell” of this “Mammiliferous Crag” (former name of the Norwich Crag Formation). In the Netherlands, Littorina littorea is common in the Maassluis Formation in borehole Schelphoek (Slupik et al., 2007).

Figure 5. RGM.1365361. Littorina littorea Linnaeus, 1758 from sample 58–59 m, Maassluis Formation, borehole Huijbergen.
30In the study area, the Gelasian “Icenian” molluscs were only reported by Halet (1933) north of the line AB (Fig. 1). Although sediments of the Merksplas Formation appear similar north- and southward of this line, Quaternary marine species do not occur southward. There, only reworked Pliocene shells were reported at the base (Halet, 1933). This might indicate that the early Quaternary transgression (coming from the north) that created shallow marine conditions reached the AB line (Fig. 1). The coarse grain sizes of the Maassluis, Waalre and Merksplas formations with reworked shells testify to a high-energy depositional environment in the Huijbergen and Essen area. Such high-energy, marginal marine (possibly estuarine) settings provide adverse conditions for the preservation of dinocysts (oxidation), which explains why only a few and no age-diagnostic dinocyst species were found within these units in the Huijbergen borehole. The age of the unfossiliferous, upper interval of the Merksplas Formation is due to correlation with the Waalre Formation (WA-2) not older than Early Pleistocene (Gelasian) (e.g. Westerhoff, 2009).
6.4. Older age assessments of the Merksplas Formation
31The occurrence of characteristic mollusc associations in the lower part of the Merksplas Formation offers arguments for a relative dating as Gelasian (formerly “Icenian”). However, the age of the Merksplas Formation has long been debated. It has been attributed to both the Pleistocene (De Meuter & Laga, 1976), Pliocene (Gullentops et al., 2001) and the time interval of their transition (Kasse, 1988). The most recent review (Louwye et al., 2020) included the Merksplas Formation in the Pliocene, referring to Vanhoorne (1962). The latter analysed organic-rich samples from borehole Kalmthout 2 (BGD 006E0089; DOV kb7d6e-B115), most probably belonging to the Merksplas Formation (“pale grey very quartz-rich sand with lignite debris, 20 m below the clay”). The palynology appeared similar to that of the lignite in the Mol Formation, which was attributed to the Reuverian (upper Pliocene regional stage). However, the shells we used for the Gelasian dating showed clear traces of reworking. Therefore, reworking may also have affected the palynological associations of Vanhoorne (1962). In addition, Al-Silwadi (2017) found mid- to late Pliocene dinoflagellate cysts (3.7–2.72 Ma) in the 5–9 m interval of the Rees borehole (BGD 017E0399, DOV kb817e-B495), interpreted as Merksplas Formation. However, the same section was interpreted as the Vosselaar Member of the Malle Formation by Buffel et al. (2001), which raises questions on the correct stratigraphic attribution of this interval to the Merksplas Formation. The recorded Pliocene dinocysts (Capisocysta lyelli, Desotodinium wrennii, Geonettia waltonensis, Invertocysta lacrymosa, Pyxidinopsis cf. braboi) in the Rees borehole were furthermore not encountered in the Maassluis and Waalre formations of the Huijbergen borehole. Therefore, we refrain from extrapolating the conclusions of Al-Silwadi (2017) to our study area.
7. Conclusions
32The lithological and biostratigraphic aspects of the Huijbergen borehole in the southern Netherlands provide valuable insights on the Pliocene–Pleistocene successions near the Dutch-Belgian border:
-
The Dutch Oosterhout Formation can be correlated with the Belgian Lillo Formation. Based on dinoflagellate cysts and molluscs, a Piacenzian age is inferred for the studied succession in borehole Huijbergen. The geotechnical and geophysical threefold division of the Lillo Formation in Belgium can be recognised across the border into the Netherlands (Fig. 4). The dinocysts and molluscs found in these intervals closely resemble the associations and faunas known from the Li-A, Li-B and Li-C intervals of the Antwerp harbour area. Although the interval 69–75 m in Huijbergen can be correlated with the Belgian Oorderen Member (Piacenzian), some species are only known from the Zanclean Luchtbal Member in the Antwerp area, but these are considered facies indicators.
-
The Maassluis Formation can be correlated with the shell-bearing lower part of the Merksplas Formation. A Gelasian age is inferred based on the marine molluscs present in both formations. In more northern Dutch boreholes, similar associations are known from certain intervals within thick age-constrained successions (Meijer et al., 2006; Slupik et al., 2007; Noorbergen et al., 2015; unpublished data). The absence of very cold faunal components indicates that the Maassluis Formation in Huijbergen and the shelly intervals of the Merksplas Formation represent a warm-temperate interglacial highstand within the Praetiglian or Tiglian stages (both Gelasian). The studied fauna is well distinguished from the Pliocene faunas from the underlying Lillo and Oosterhout formations by the (common) appearance of e.g. Littorina littorea, Turritellinella communis, Mactra stultorum, Cerastoderma edule, Spisula subtruncata and Donax vittatus. This fauna disappears southward in the Merksplas Formation, which was noted by Halet (1933). The (very) coarse grain sizes and the abraded, often fragmented shells point to a high-energy environment, probably within marginal marine (possibly estuarine) depositional settings.
-
The Waalre Formation (WA-2 subunit) in Huijbergen can be correlated with the higher, unfossiliferous part of the Merksplas Formation. Because of the lack of diagnostic dinocysts or molluscs, the age of this unit could not further be established, other than being Gelasian or younger. In the Kalmthout 1 borehole, shells are absent in the lower part of the Merksplas Formation, where it completely correlates with the Waalre Formation.
-
In this study, the increase in grain sizes and the disappearance of glauconite were used to distinguish the Merksplas Formation from the underlying Lillo Formation. These are more reliable criteria than the disappearance of shells, given the local presence of shell grit in the lower part of the Merksplas Formation.
33The aim of this paper was to provide a relative age estimate of the Merksplas Formation by correlation with well-known lithostratigraphic units in the Netherlands. It does not resolve other long-standing discussions regarding the relationship of the Merksplas Formation with other units in the Campine region, such as the Malle Formation (including the Brasschaat Member) and the Mol Formation. However, it opens the opportunity to reconsider these issues in the future.
Acknowledgments
34Andre Slupik (TNO-GSN, Utrecht) helped by making the Huijbergen samples available for study. We would like to express our gratitude to Armin Menkovic (TNO-GSN) for the discussion on Dutch lithostratigraphy. Nico Janssen (TNO-GSN) is acknowledged for the dinoflagellate cyst preparation. Katleen van Baelen (VITO, Mol) is thanked for her excellent work on the figures. Pieter De Schutter (RBINS, Brussels) for making photographs of the sediment samples. SE thanks Marijn Roosen (NHM, Rotterdam), Tom Meijer (Alkmaar) and Peter Moerdijk (Middelburg) for sharing their opinions on certain molluscs. We also want to thank the reviewers for their helpful suggestions, improving our paper substantially: Noël Vandenberghe (KU Leuven), Stephen Louwye (UGent) and Stijn Goolaerts (RBINS).
Author contribution
35Jef Deckers conceptualised the research strategy and improved geophysical correlations. Stijn Everaert and Frank Wesselingh studied the molluscs. Dirk Munsterman studied the palynology. Stijn Everaert and Jef Deckers (re)interpreted some Belgian reference boreholes. All authors took part in the sampling and writing of the paper.
Data availability
36All sieving residues and molluscs of the Huijbergen borehole are stored at the Fossil Mollusc Collections of Naturalis Biodiversity Center (Leiden, the Netherlands), collection numbers are listed in the caption of Figure 3. The palynological slides are stored at TNO-GSN (Utrecht, the Netherlands) together with the other material of borehole Huijbergen.
References
37Aleffi, F. & Bettoso, N., 2000. Distribution of Corbula gibba (Bivalvia, Corbulidae) in the northern Adriatic Sea. Annales. Series historia naturalis, 10, 173–180.
38Al-Silwadi, S., 2017. Dinoflagellate cyst stratigraphy and paleoecology of the Upper Miocene and Pliocene, Rees Borehole, Northern Belgium. Unpublished MSc. Thesis Earth Science, Brock University, Ontario, Canada, 128 p.
39BGD, 1980. Boorverslag BGD 006E0130. Belgische Geologische Dienst. https://collections.naturalsciences.be/ssh-geology-archives/arch/006e/006e0130.txt, accessed 27/12/2022.
40Bogemans, F. & Lanckacker, T., 2014. Neogene – revised lithostratigraphy accepted 062014: 2.10. Merksplas Formation – Me. National Commission for Stratigraphy Belgium. https://ncs.naturalsciences.be/paleogene-neogene/210-merksplas-formation, accessed 28/12/2022.
41Boele, J., 2001. Mollusken en andere fossielen uit een boring in Sliedrecht. Afzettingen WTKG, 22, 34–36.
42Buffel, P., Vandenberghe, N., Goolaerts, S. & Laga, P., 2001. The Pliocene in four boreholes in the Turnhout area (North-Belgium): the relation with the Lillo and Mol Formations. Aardkundige Mededelingen, 11, 1–9.
43Dearing Crampton-Flood, E., Peterse, F., Munsterman, D. & Sinninghe Damsté, J.S., 2018. Using tetraether lipids archived in North Sea Basin sediments to extract North Western European Pliocene continental air temperatures. Earth and Planetary Science Letters, 490, 193–205. https://doi.org/10.1016/j.epsl.2018.03.030
44Dearing Crampton-Flood, E., Noorbergen, L., Smits, D., Boschman, C., Donders, T., Munsterman, D.K., Ten Veen, J., Peterse, F., Lourens, L. & Sinninghe Damsté, J.S., 2020. A new age model for the Pliocene of the southern North Sea basin: a multi-proxy climate reconstruction. Climate of the Past, 16, 523–541. https://doi.org/10.5194/cp-16-523-2020
45Deckers, J., Louwye, S. & Goolaerts, S., 2020. The internal division of the Pliocene Lillo Formation: correlation between Cone Penetration Tests and lithostratigraphic type sections. Geologica Belgica, 23/3-4, 333–343. https://doi.org/10.20341/gb.2020.027
46de Heinzelin de Braucourt, J., 1950. Stratigraphie pliocène et quaternaire observée au Kruisschans. I. Analyse stratigraphique & II. Conclusions. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 26/40-41, 1–38 & 1–22.
47de Heinzelin de Braucourt, J., 1955a. Deuxième série d’observations stratigraphiques au Kruisschans. Coupes de l’écluse Baudoin. I. Analyse stratigraphique & II. Conclusions. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 31/66-67, 1–29 & 1–14.
48de Heinzelin de Braucourt, J., 1955b. Considérations nouvelles sur le Néogène de l’Ouest de l’Europe. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 64/3, 463–476.
49De Meuter, F.J. & Laga, P.G., 1976. Lithostratigraphy and biostratigraphy based on benthonic foraminifera of the Neogene deposits of Northern Belgium. Bulletin de la Société belge de Géologie, 85/4, 133–152.
50De Schepper, S. & Head, M.J., 2008. Age calibration of dinoflagellate cyst and acritarch events in the Pliocene–Pleistocene of the eastern North Atlantic (DSDP Hole 610A). Stratigraphy, 5/2, 137–161.
51De Schepper, S., Head, M.J. & Louwye, S., 2009. Pliocene dinoflagellate cyst stratigraphy, palaeoecology and sequence stratigraphy of the Tunnel-Canal Dock, Belgium. Geological Magazine, 146/1, 92–112. https://doi.org/10.1017/S0016756808005438
52De Schepper, S., Schreck, M., Beck, K.M., Matthiessen, J., Fahl, K. & Mangerud, G., 2015. Early Pliocene onset of modern Nordic Seas circulation related to ocean gateway changes. Nature Communications, 6, 8659. https://doi.org/10.1038/ncomms9659
53Doppert, J.W.Chr., Ruegg, G.H.J., Van Staalduinen, C.J., Zagwijn, W.H. & Zandstra, J.G. 1975. Formaties van het Kwartair en Boven-Tertiair in Nederland. In Zagwijn, W.H. & Van Staalduinen, C.J. (eds), Toelichting bij geologische overzichtskaarten van Nederland. Rijks Geologische Dienst, Haarlem, 11–56.
54Doppert, J.W.C., Laga, P. & De Meuter, F., 1979. Correlation of the biostratigraphy of marine Neogene deposits, based on benthonic foraminifera, established in Belgium and The Netherlands. Mededelingen Rijks Geologische Dienst, 31, 1–8.
55Fensome, R.A., Williams, G.L. & MacRae, R.A., 2019. The Lentin and Williams Index of fossil dinoflagellates 2019 edition. AASP Contributions Series, 50, 1173 p.
56Gibbard, P.L. & Head, M.J., 2020. The Quaternary Period. In Gradstein, F.M., Ogg, J.G., Schmitz, M.D. & Ogg, G.M. (eds), Geological Time Scale 2020. Elsevier, Amsterdam, 1217–1255. https://doi.org/10.1016/B978-0-12-824360-2.00030-9
57Glibert, M., 1957. Gastropodes du Diestien, du Scaldisien et du Merkxemien de la Belgique. Première Note. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 33/36, 1–27.
58Glibert, M., 1958. Tableau stratigraphique des Mollusques du Néogène de la Belgique. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 34/32, 1–20.
59Gulinck, M., 1962. Essai d’une carte géologique de la Campine. Etat de nos connaissances sur la nature des terrains néogènes recoupés par sondages. Mémoires de la Société belge de Géologie, de Paléontologie et d’Hydrologie, série in-8°, 6, 30–39.
60Gulinck, M. & Paepe, R., 1969. Boorverslag BGD 006E0106. Aardkundige verklaring. Belgische Geologische Dienst. https://collections.naturalsciences.be/ssh-geology-archives/arch/006e?b_start:int=100, accessed 20/10/2022.
61Gullentops, F., Bogemans, F., De Moor, G., Paulissen, E. & Pissart, A., 2001. Quaternary lithostratigraphic units (Belgium). Geologica Belgica, 4, 153–164. https://doi.org/10.20341/gb.2014.051
62Hayward, P.J. & Ryland, J.S. (eds), 2017. Handbook of the Marine Fauna of North-West Europe. 2nd ed. Oxford University Press, Oxford, 785 p. https://doi.org/10.1093/acprof:oso/9780199549443.001.0001
63Halet, F., 1933. Observations nouvelles sur l’âge des dépôts dits amstéliens de la partie septentrionale de la Campine anversoise. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 43, 394–409.
64Harmer, F.W., 1920. The Pliocene Mollusca of Great Britain, vol. II, part I. London, Palaeontographical Society, 72, 485–652. https://doi.org/10.1080/02693445.1920.12035579
65Hennissen, J.A., Head, M.J., De Schepper, S. & Groeneveld, J., 2017. Dinoflagellate cyst paleoecology during the Pliocene–Pleistocene climatic transition in the North Atlantic. Palaeogeography, Palaeoclimatology, Palaeoecology, 470, 81–108. https://doi.org/10.1016/j.palaeo.2016.12.023
66Janssen, N. & Dammers, G., 2008. Sample processing for pre-Quaternary palynology. Internal TNO report, May 2008. Geological Survey of the Netherlands, 9 p.
67Kasse, C., 1988. Early-Pleistocene tidal and fluviatile environments in the southern Netherlands and northern Belgium. Ph.D. Thesis, Free University Press, Amsterdam, 190 p.
68Kuhlmann, G., Langereis, C., Munsterman, D.K., van Leeuwen, R.J., Verreussel, R., Meulenkamp, J. & Wong, T., 2006. Chronostratigraphy of Late Neogene sediments in the southern North Sea Basin and paleoenvironmental interpretations. Palaeogeography, Palaeoclimatology, Palaeoecology, 239, 426–455. https://doi.org/10.1016/j.palaeo.2006.02.004
69Laga, P., 1972. Stratigrafie van de mariene Plio-Pleistocene afzettingen uit de omgeving van Antwerpen met een bijzondere studie van de foraminiferen. Unpublished Ph.D. Thesis, KU Leuven, Leuven, 252 p.
70Laga, P., 1979. Boorverslag BGD 001E0044. Belgische Geologische Dienst. https://collections.naturalsciences.be/ssh-geology-archives/arch/001e/001e0044.txt, accessed 27/12/2022.
71Laga, P. & Hanssen, E., 1976. Boorverslag BGD 003W0081. Belgische Geologische Dienst. https://collections.naturalsciences.be/ssh-geology-archives/arch/003w, accessed 23/12/2022.
72Laga, P., Louwye, S. & Mostaert, F., 2006. Disused Neogene and Quaternary regional stages from Belgium: Bolderian, Houthalenian, Antwerpian, Diestian, Deurnian, Kasterlian, Kattendijkian, Scaldisian, Poederlian, Merksemian and Flandrian. Geologica Belgica, 9, 215–224.
73Leifsdóttir, Ó.E. & Símonarson, L.A., 2002. The mesogastropod Littorina littorea (Linné, 1758) in Iceland: palaeobiogeography and migration. Cainozoic Research, 1, 3–12.
74Louwye, S., Deckers, J. & Vandenberghe, N., 2020. The Pliocene Lillo, Poederlee, Merksplas, Mol and Kieseloolite Formations in northern Belgium: a synthesis. Geologica Belgica, 23/3-4, 297–313. https://doi.org/10.20341/gb.2020.016
75Marquet, R., 1993. The molluscan fauna of the Kruisschans Member (Lillo Formation, Late Pliocene) in the Antwerp Area (Belgium). Contributions to Tertiary and Quaternary Geology, 30, 83–103.
76Marquet, R., 1998. De Pliocene gastropodenfauna van Kallo (OostVlaanderen, België). Publicatie van de Belgische Vereniging voor Paleontologie vzw, 17, 246 p.
77Marquet, R., 2001. A study of some Neogene European species of Seilinae (Cerithiopsidae, Gastropoda). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 71, 195–208.
78Marquet, R., 2002. The Neogene Amphineura and Bivalvia (Protobranchia and Pteriomorphia) from Kallo and Doel (Oost–Vlaanderen, Belgium). Palaeontos, 2, 1–100.
79Marquet, R., 2004. Ecology and evolution of Pliocene bivalves from the Antwerp Basin. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 74 supplément, 205–212.
80Marquet, R., 2005. The Neogene Bivalvia (Heterodonta and Anomalodesmata) and Scaphopoda from Kallo and Doel (OostVlaanderen, Belgium). Palaeontos, 6, 1–142.
81Marquet, R. & Landau, B.M., 2006. The gastropod fauna of the Luchtbal Sand Member (Lillo Formation, Zanclean, Early Pliocene) of the Antwerp region (Belgium). Cainozoic Research, 5, 13–50.
82Meijer, T., Cleveringa, P., Munsterman, D.K. & Verreussel, R.M.C.H., 2006. The Early Pleistocene Praetiglian and Ludhamian pollen stages in the North Sea Basin and their relationship to the marine isotope record. Journal of Quaternary Science, 21, 307–310. https://doi.org/10.1002/jqs.956
83Menkhorst, H.P.M.G. & Wesselingh, F.P., 2018. De fossiele slakken van de Nederlandse kust II. Deel 12. Eulimacea, Cerithiopsacea, Triforacea. Spirula, 415, 38–51.
84Moerdijk, P.W. & Janse, A.C., 2015. De fossiele schelpen van de Nederlandse kust II, deel 9. Littorinidea. Spirula, 402, 20–24.
85Moerdijk, P.W., Janssen, A.W., Wesselingh, F.P., Peeters, G.A., Pouwer, R., Van Nieulande, F.A.D., Janse, A.C., Van der Slik, L., Meijer, T., Rijken, R., Cadée, G.C., Hoeksema, D., Doeksen, G., Bastemeijer, A., Strack, H., Vervoenen, M. & Ter Poorten, J.J., 2010. De fossiele schelpen van de Nederlandse kust. Nederlands Centrum voor Biodiversiteit Naturalis, Leiden, 294 p.
86Moerdijk, P.W., van Nieulande, F. & Pouwer, R., 2018. De fossiele schelpen van de Nederlandse kust II, deel 13. Turritellidae en Vermetidae. Spirula, 416, 25–36.
87Mourlon, M., 1897. La faune marine du quaternaire moséen révélée par les sondages de Strijbeek et de Wortel. Bulletin de l’Académie royale de Belgique, 3e série, 33, 778–780.
88Munsterman, D.K., 2021. Boring B37D0228: lectostratotype sectie van Formatie van Maassluis, interval 103–206 m. Internal TNO report 2021 R11284. Geological Survey of the Netherlands, 18 p.
89Munsterman, D.K., ten Veen, J.H, Menkovic, A., Deckers, J., Witmans, N., Verhaegen, J., Kerstholt–Boegehold S.J., van de Ven, T & Busschers, F.S., 2019. An updated and revised stratigraphic framework for the Miocene and earliest Pliocene strata of the Roer Valley Graben and adjacent blocks. Netherlands Journal of Geosciences, 98, e8. https://doi.org/10.1017/njg.2019.10
90Noorbergen, L.J., Lourens, L.J., Munsterman, D.K. & Verreussel, R.M.C.H., 2015. Stable isotope stratigraphy of the early Quaternary of borehole Noordwijk, southern North Sea. Quaternary International, 386, 148–157. https://doi.org/10.1016/j.quaint.2015.02.045
91Norton, P.E.P., 1967. Marine Molluscan assemblages in the Early Pleistocene of Sidestrand, Bramerton and the Royal Society Borehole at Ludham, Norfolk. Philosophical Transactions of the Royal Society of London B, 253, 161–200. https://doi.org/10.1098/rstb.1967.0034
92Pannekoek, A.J. (ed.) 1956. De Geologische geschiedenis van Nederland. Toelichting bij de Geologische overzichtskaart van Nederland op de schaal 1:200.000. Staatsdrukkerij en uitgeverijbedrijf, ’s-Gravenhage, 154 p.
93Pouwer, R. & Rijken, R., 2022. De fossiele schelpen van de Nederlandse kust II, deel 16. Naticidae. Spirula, 430, 30–38.
94Pouwer, R. & Wesselingh, F.W., 2012. De fossiele schelpen van de Nederlandse kust II, (deel 3). Trochidae, Solariellidae en Calliostomatidae. Spirula, 389, 151–164.
95Preece, R.C., Meijer, T., Penkman, K.E.H., Demarchi, D., Mayhew, D.F. & Parfitt, S.A., 2020. The palaeontology and dating of the ‘Weybourne Crag’, an important marker horizon in the Early Pleistocene of the southern North Sea Basin. Quaternary Science Reviews, 236, 106177. https://doi.org/10.1016/j.quascirev.2020.106177
96Raad, H., Rijken, R. & Pouwer, R., 2016. De fossiele schelpen van de Nederlandse kust II, deel 11. Conoidea. Spirula, 409, 28–49.
97Rijken, R. & Pouwer, R., 2014. De fossiele schelpen van de Nederlandse kust II, deel 7. Nassariidae. Spirula, 379, 46–55.
98Slupik, A.., Wesselingh, F.P. & Reumer, J.W.F., 2007. The stratigraphy of the Neogene–Quaternary succession in the southwest Netherlands from the Schelphoek borehole (42G4–11/42G0022) – a sequence-stratigraphic approach. Netherlands Journal of Geosciences – Geologie en Mijnbouw, 86, 317–332. https://doi.org/10.1017/S0016774600023556
99Spaink, G., 1975. Zonering van het mariene Onder–Pleistoceen en Plioceen op grond van mollusken fauna’s. In Zagwijn, W.H. & Van Staalduinen, C.J. (eds), Toelichting bij geologische overzichtskaarten van Nederland. Rijks Geologische Dienst, Haarlem, 118–122.
100TNO-GSN, 2012. Hypostratotype 49G0204. https://www.dinoloket.nl/sites/default/files/nomenclator/typelocaties/B49G0204.pdf, accessed 18/12/2022.
101TNO-GSN, 2022a. Maassluis Formation. In Stratigraphic Nomenclature of the Netherlands. TNO – Geological Survey of the Netherlands. https://www.dinoloket.nl/en/stratigraphic-nomenclature/maassluis-formation, accessed 7/10/2022.
102TNO-GSN, 2022b. Waalre Formation. In Stratigraphic Nomenclature of the Netherlands. TNO – Geological Survey of the Netherlands. http://www.dinoloket.nl/en/stratigraphic–nomenclature/waalre–formation, accessed 7/10/2022.
103Vanhoorne, R., 1962. La superposition des Sables de Mol et des Argiles de Campine. Mémoires de la Société belge de Géologie, de Paléontologie et d’Hydrologie, série in–8°, 6, 83–95.
104Vervoenen, M., 1995. Taphonomy of some Cenozoic Seabeds from the Flemish Region, Belgium. Belgische Geologische Dienst, Professional paper, 1994/5, 272, 115 p.
105Vervoenen, M., Wesselingh, F.P. & Van Nieulande, F.A.D., 2000. Mytilus antiquorum J. Sowerby, 1821 and other Pliocene mussels (Mollusca, Bivalvia) from the southern North Sea Basin. Contributions to Tertiary and Quaternary Geology, 37, 73–81.
106Vernes, R.W., Deckers, J., Bakker, M.A.J., Bogemans, F., De Ceukelaire, M., Doornenbal, J.C., den Dulk, M., Dusar, M., Van Haren, T.F.M., Heyvaert, V.M.A., Kiden, P., Kruisselbrink, A.F., Lanckacker, T., Menkovic, A., Meyvis, B., Munsterman, D.K., Reindersma, R., Rombaut, B., ten Veen, J.H., van de Ven, T.J.M., Walstra, J. & Witmans, N., 2018. Geologisch en hydrogeologisch 3D model van het Cenozoïcum van de Belgisch-Nederlandse grensstreek van Midden-Brabant / De Kempen (H3O – De Kempen). TNO–rapport TNO 2017 R11261 – VITO 2017/RMA/R/1348, 109 p. https://archief.onderzoek.omgeving.vlaanderen.be/Onderzoek–2314288, accessed 23/12/2022.
107Vinken, R. (ed.), 1988. The Northwest European Tertiary Basin. Geologisches Jahrbuch, Reihe A, 100, 508 p.
108Wesselingh, F.P., Rijken, R., van Nieulande, F., Janse, A.C. & Pouwer, R., 2012. De fossiele schelpen van de Nederlandse kust II. De Cerithium–achtigen (deel 2). Spirula, 385, 37–47.
109Wesselingh, F.P., van Nieulande, F., Fraussen, K., Moerdijk, P.W., Janssen, A.W., Pouwer, R. & Rijken, R., 2014. De fossiele schelpen van de Nederlandse kust II, deel 8. Buccinoidea. Spirula, 401, 194–205.
110Wesselingh, F.P., Busschers, F.S. & Goolaerts, S., 2020. Observations on the Pliocene sediments exposed at Antwerp International Airport (northern Belgium) constrain the stratigraphic position of the Broechem Fauna. Geologica Belgica, 23, 315–321. https://doi.org/10.20341/gb.2020.026
111Westerhoff, W.E., 2009. Stratigraphy and sedimentary evolution: The lower Rhine-Meuse system during the Late Pliocene and Early Pleistocene (southern North Sea Basin). Ph.D. Thesis, Vrije Universiteit Amsterdam. Published by TNO Built Environment and Geosciences – Geological Survey of the Netherlands, Amsterdam, 168 p.
112Wijnker, E., Bor, T.J., Wesselingh, F.P., Munsterman, D.K., Brinkhuis, H., Burger, A.W., Vonhof, H.B., Post, K., Hoedemakers, K., Janse, A.C. & Taverne, N., 2008. Neogene stratigraphy of the Langenboom locality (Noord–Brabant, the Netherlands). Netherlands Journal of Geosciences, 87, 165–180. https://doi.org/10.1017/S0016774600023209
113Wood, S.V., 1848. A monograph of the Crag Mollusca, or, description of shells from the Middle and Upper Tertiaries of the east of England, 1. Univalves. Monograph of the Palaeontographical Society of London, xii + 1–208.
114Zonneveld, J.I.S. 1958. Litho-stratigrafische eenheden in het Nederlandse Pleistoceen. Mededelingen van de Geologische Stichting, Nieuwe Serie, 12, 31–64.
115Manuscript received 21.10.2022, accepted in revised form 28.02.2023, available online 01.05.2023.
Pour citer cet article
