
A re-investigation of the Rhabdinopora flabelliformis fauna from the early Tremadocian ‘Dictyonema Shale’ in Belgium
Abstract
During a period of almost 150 years, the diverse Rhabdinopora flabelliformis fauna from the Tremadocian (Early Ordovician) of Belgium has been collected from both the Brabant Massif and the Stavelot and Rocroi Inliers (‘Ardenne Massifs’), and in a single borehole from the Condroz Inlier. In the present study, the large collections of the Royal Belgian Institute of Natural Sciences (Brussels) are revised and re-described in the light of present-day graptolite taxonomy. The taxa Rhabdinopora praeparabola, Rhabdinopora flabelliformis flabelliformis, R. f. anglica, R. f. socialis, R. f. norvegica and R. f. parabola have been identified. Based on our taxonomical revision, the early Tremadocian Rhabdinopora praeparabola, R. f. parabola and R. f. anglica graptolite biozones are recognized in the Brabant Massif, allowing correlation with other graptolitic successions worldwide. At the level of regional biostratigraphy, the boundary between the Mousty Formation and the Chevlipont Formation in the Brabant Massif can now be precisely located in the R. praeparabola graptolite Biozone, while the boundary between the Solwaster Member (Sm1a) and the Spa Member (Sm1b) of the Jalhay Formation in the Stavelot Inlier (Ardenne) is located within the R. f. anglica Biozone. All the Rhabdinopora flabelliformis fauna levels can be attributed to Time-Slice 1a of the lower Tremadocian.
1. Introduction
1The Dictyonema Shale (‘Dictyonema oil shale’, ‘Dictyonema argillite’, etc.) is a largely used term, representing Tremadocian (the lowest stage of the Ordovician) shales that are rich in organic matter and generally contain graptolitic fossils (the Dictyonema flabelliforme fauna) (Veski & Palu, 2003). The name ‘Dictyonema’ derived from the graptolite species Dictyonema flabelliforme (Eichwald, 1840) (now Rhabdinopora flabelliformis) which was originally thought to be a benthonic root-bearing taxon, but was later considered to be a planktonic nema-bearing graptolite, reattributed to Rhabdinopora Eichwald 1855 (Erdtmann, 1982). Currently, the taxa that have been placed in the original graptolite genus Dictyonema are considered as either of benthonic type (Dictyonema s.s.) or of planktonic type (Rhabdinopora) (Erdtmann, 1982). Therefore most of the formations attributed to the ‘Dictyonema shales’ do actually not display any real Dictyonema, but contain the planktonic genus Rhabdinopora instead. However, the name has been retained because for more than one century and a half the term of ‘Dictyonema Shale’ has been largely used and is well known as a source rock for uranium and some other heavy metals (Althausen, 1992).
2Since the 1980s, much research was focused on the precise position of the Cambrian-Ordovician boundary (which is also the base of the Tremadocian stage), resulting in the detailed investigation of graptolites and conodonts from the ‘Dictyonema shales’ of many localities, including the potential candidate sections for the Global Stratigraphy Section and Point (GSSP) for the base of the Ordovician System (Cooper et al., 2001). Many lower Tremadocian successions, such as the Green Point section in Newfoundland, Canada, the Nærsnes section in Oslo, Norway, and the Dayangcha section in Jilin Province, North China, have been studied in detail (e.g., Bruton et al., 1982; Cooper et al., 1998; Wang et al., 1998).
3In Belgium, graptolites from the ‘Dictyonema shale’ have been recorded in many localities since the late 19th century from the Ardenne Inliers (Stavelot and Rocroi ‘massifs’) (e.g., Malaise, 1874a, 1874b, 1881; Bulman, 1950a, 1970; Geukens, 1950, 1954; Erdtmann, 1986), and subsequently in the 1940’s also from the Brabant Massif (e.g., Lecompte, 1948, 1949) and the Wépion borehole in the Condroz Inlier (Graulich, 1961, 1963). The early works enabled a rough correlation between the Belgian Rhabdinopora flabelliformis fauna and the British R. socialis – flabelliformis biozones that is used up to date (e.g., Verniers et al., 2001; Herbosch & Verniers, 2013). However, it was so far difficult to conduct a precise correlation between the lower Tremadocian graptolite biozones from Belgium and those from other sections worldwide because the Belgian biostratigraphy has not been revised since more than half a century. Only the fossil assemblage of a single locality (Solwaster) in the Stavelot Massif was described by Erdtmann (1986) almost thirty years ago. This latter research was conducted in the frame of the search for potential candidate sections of the base of the Ordovician (Erdtmann, 1986).
4The largest collections of graptolite specimens from the ‘Dictyonema shales’ of Belgium are deposited in the Royal Belgian Institute of Natural Sciences, Brussels. The Rhabdinopora flabelliformis faunas of these collections have not been re-investigated since the 1970s. It appears therefore useful to re-analyze these faunas and to publish the revised lists of these collections in the light of present-day graptolite taxonomy. In addition, the Rhabdinopora-bearing strata of the borehole of Wépion, that displays the Early Ordovician of the Condroz Inlier, deposited in the collections of the Geological Survey of Belgium, has also been re-analyzed.
5Up to date taxonomic methods are used for the classification of the faunal assemblages. Regional and global correlations are possible following the recognition of stratigraphically important taxa such as R. praeparabola, R. f. parabola and R. f. anglica. The aim of the present work is not only the taxonomical re-investigation of the graptolite fauna, but also a more precise understanding of the lowest Tremadocian graptolite stratigraphy in Belgium. In addition, the present work also allows a somewhat finer subdivision of the lower Tremadocian graptolite zones in Avalonia.
2. Material and methods
6In Belgium, Rhabdinopora specimens have been collected and studied from two major areas, the Brabant Massif and the Ardenne Inliers (or ‘massifs’), of which the ‘Stavelot Massif’ in the eastern part of Belgium is the most significant in size (Fig. 1 A). An additional finding of a few specimens comes from the narrow Condroz Inlier (‘Bande de Sambre-et-Meuse’), located between the Brabant and the Ardennes.
7The discoveries of ‘Dictyonema’ assemblages from the Ardennes date back to the 1870’s with descriptions of species of Rhabdinopora (historically attributed to ‘Dictyonema’) that have been reported from numerous localities in the Salm Group of the Stavelot Inlier (‘Salmien’), including Sart, Spa, Verviers, La Gleize, Harzé, Jalhay-Gileppe and Solwaster-Gospinal (e.g., Malaise 1874a, 1881, 1888; Geukens, 1950, 1954; Bulman, 1970; Erdtmann, 1986 and references therein). The same levels were also recovered from the Rocroi Inlier in southern Belgium (but not from the Givonne and Serpont inliers), with discoveries of ‘Dictyonema sociale’ as early as 1874 in the ‘Revin slates’ near the old mill of Mairus by Malaise (1874b).
8The discoveries of ‘Dictyonema’ assemblages from the Brabant Massif are mostly from the Thyle River section (Fig. 1 B) from the lower Mousty Formation (now the Mousty Fm. and the Chevlipont Fm.) of localities including La Roche-Faux, Chastres and Genappe (e.g., Lecompte, 1948, 1949).
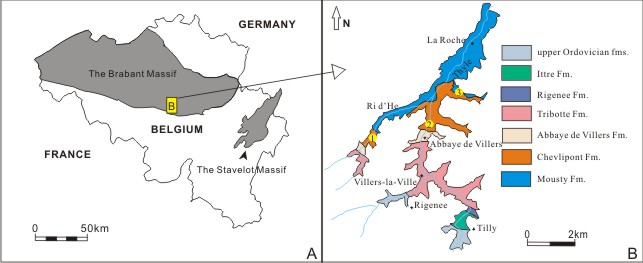
9Figure 1. A. Simplified map of the Stavelot Massif and the Brabant Massif of Belgium. B. The localities of specimens collected by Lecompte (1949) in the Brabant Massif, according to Lecompte (1949) and Herbosch & Verniers (2013). 1. Locality Genappe 26; 2. Locality Genappe 78; 3. Locality Chastre 95.
10The lowest Ordovician of the Condroz Inlier (‘Bande de Sambre et Meuse’) is not present in outcrops, but was only recovered from a single deep geological drilling, the Wépion borehole (Graulich, 1961).
11In the present study, most of the material that is re-described was originally collected by Maillieux in the year 1925 and Lecompte (1948, 1949) and investigated in terms of graptolite taxonomy by Bulman (1950a, 1970). Another portion of the material present in the Museum at Brussels, especially the collections from the Ardenne inliers, was collected by numerous anonymous collectors. The details of the localities, collectors, horizons and numbers of the re-investigated specimens are listed in Table 1. Among all the specimens, those collected by Lecompte (1949) can be clearly traced on their original sections and some of the outcrops are still visible (Herbosch and Verniers, personal comm.). They are therefore of high biostratigraphical potential and can be used to recognize internationally used graptolite biozones.
3. Results
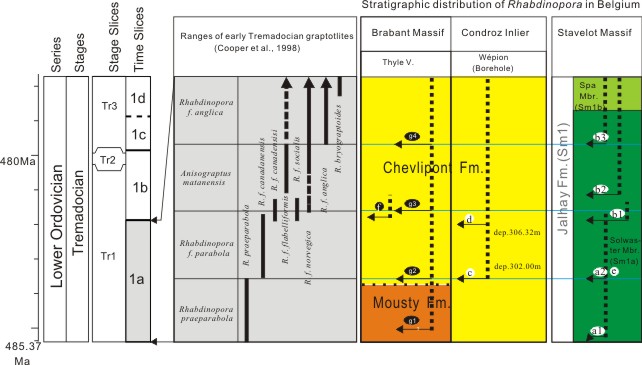
12In the present study, all Rhabdinopora collections from the Royal Belgian Institute of Natural Sciences, Brussels (Brabant Massif and Ardenne inliers) and from the Belgian Geological Survey (Wépion borehole in the Condroz Inlier) have been re-investigated. Table 1 includes a list of the important species for which the new identifications differ from the original names provided by previous authors. A selection of the investigated specimens is illustrated in Figures 4-6. For more details on taxonomy, the reader is referred to the Systematic Palaeontology section. In the Stavelot Massif, the specimens from the different localities can be attributed to the following taxa (as represented in Fig. 2): R. praeparabola, R. f. parabola (by Erdtmann, 1986), R. f. flabelliformis; R. f. socialis, and R. f. anglica (by other anonymous authors).
|
Outcrop area; formation; locality |
Original study |
Number of specimens |
Original identification |
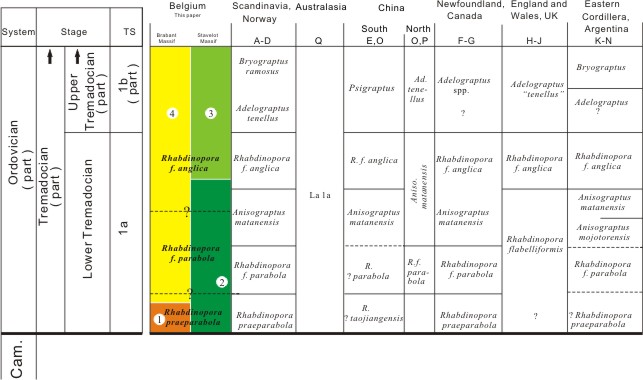
Identification in the present study |
|
Condroz Inlier; Chevlipont Fm.; Wépion borehole |
Graulich (1961) |
3 |
R. f. aff. norvegica |
R. f. norvegica |
|
Condroz Inlier; Chevlipont Fm.; Wépion borehole |
Graulich (1961) |
4 |
Not recognized |
R. f. parabola |
|
Brabant Massif; Chevlipont Fm.; Moulin de Chevlipont = Genappe 78 |
Lecompte (1949) |
27 |
Dictyonema D. f. norvegica D. f. sociale R. f. flabelliformis |
R. f. parabola R. f. flabelliformis R. praeparabola R. f. norvegica R. f. socialis |
|
Brabant Massif; Chevlipont Fm.; La Roche = Chastres 95 |
Lecompte (1949) |
20 |
D. f. typica D. f. sociale |
R. f. parabola R. praeparabola R. f. socialis R. f. flabelliformis |
|
Brabant Massif; Chevlipont Fm.; Genappe 26 |
Lecompte (1949) |
2 |
D. f. flabelliformis D. f. anglica D. f. norvegica |
R. f. anglica R. f. norvegica |
|
Brabant Massif; Chevlipont Fm.; Chastres 106 |
Lecompte (1949) |
4 |
Dictyonema |
R. f. norvegica R. f. flabelliformis |
|
Stavelot Massif; formation not specified; Lorce |
Lecompte (1948) |
2 |
D. f. sociale |
R. f. flabelliformis R. f. norvegica R. f. parabola |
|
Stavelot Massif; formation not specified; Spa |
Lecompte (1948) |
1 |
D. f. sociale |
R. f. flabelliformis R. f. parabola |
|
Stavelot Massif; Vielsalm Fm.; Lierneux, roadcut section Stoumont to Stavelot |
Lecompte (1948) |
1 |
D. f. sociale |
R. f. norvegica |
|
Stavelot Massif; formation not specified; Rahier |
Lecompte (1949) |
2 |
D. f. norvegica |
R. f. anglica |
|
Stavelot Massif; Vielsalm Fm.; Lierneux, roadcut section Sart à Spa-Stavelot |
Maillieux In 1925 |
16 |
D. flabelliforme |
R. f. anglica/R. f. flabelliformis |
|
Stavelot Massif; formation not specified; Rid’Hez |
Lecompte (1948) |
2 |
D. flabelliforme |
R. f. anglica/R. f. flabelliformis |
|
Stavelot Massif; Vielsalm Fm.; La Gleize |
Lecompte (1948) |
3 |
D. f. flabelliforme |
R. f. socialis |
|
Stavelot Massif; Tm IIIa; La Gileppe |
Bulman (1949) |
9 |
D. f. sociale |
R. f. socialis |
13Table 1. Revision of selected Tremadocian graptolites deposited in the Royal Belgian Institute of Natural Sciences, Brussels. Species in bold represent newly identified taxa.
14In the Wépion borehole of the Condroz Inlier, only very few specimens were recovered and identified by Graulich (1961). They appear at depths between 302.00 and 306.32 m and can be attributed to R. f. norvegica and R. f. parabola.
15In the light of modern graptolite taxonomy, the specimens from the locality La Roche in the Thyle River valley (Brabant Massif) can be attributed to R. f. parabola, R. praeparabola, R. f. socialis and R. f. flabelliformis; the outcrops at Genappe provide specimens attributed to R. praeparabola, R. f. parabola, R. f. flabelliformis, R. f. norvegica, R. f. anglica and R. flabelliformis; R. flabelliformis norvegica and R. f. flabelliformis are present at the locality Chastres. The reader is referred to Figure 2 and to the Systematic Palaeontology section for further detail.
4. Local and regional stratigraphical implication
16In the section for the Global Stratigraphy Section and Point (GSSP) for the base of the Tremadocian stage (base of the Ordovician System) at Green Point, Newfoundland (Cooper, 1999; Cooper et al., 2001), four lower Tremadocian graptolite biozones have been defined, in ascending order the R. praeparabola, the R. f. parabola, the A. matanensis and the R. f. anglica biozones. Cooper et al. (1998, figs 3, 4) and Cooper (1999, fig. 1) presented the ranges of the Tremadocian graptolite species from many sequences around the world and suggested an evolutionary transition with a sequence of the following taxa from bottom to top of R. praeparabola - R. f. parabola - R. f. canadensis - R. f. flabelliformis - R. f. anglica - R. f. bryograptides in slope environments (Erdtmann, 1982, 1986; Williams & Stevens, 1991; Cooper et al. 1998). The succession of early Tremadocian planktic graptolites thus provides a standard for comparison with the early Tremadocian sequence from Belgium (Fig. 2).
4.1. Stavelot Massif
17R. f. flabelliformis [originally identified as R. f. typica] and R. f. socialis are found in the lower part of the Solwaster Member (Salmian 1a = Sm1a), while R. f. anglica and R. f. norvegica do occur in the upper part of the Solwaster Member (Sm1a) and in the Spa Member (Sm1b). These results confirm the study by Lecompte (1948) in which he suggested that in the Stavelot Massif, the Rhabdinopora flabelliformis fauna appeared in strata in the following ascending order: (1) a: R. f. socialis Zone; (1) b: R. f. typica Zone and (2) c: R. f. anglica Zone. Erdtmann (1986) described R. ?praeparabola and R. parabola [now R. f. parabola] from a small section along the road from Gospinal to Solwaster in the Stavelot Massif. The appearance of R. ?praeparabola suggests that the lowest part of the Ordovician is present in this section near Solwaster and that these strata are stratigraphically even lower than those in the lower and upper parts of the Solwaster Mbr. (Sm1a) (Fig. 2).
18Figure 2. Left in grey: Composite ranges of Tremadocian graptolite species from selected sequences around the world (modified after Cooper et al., 1998, fig. 3). Right in yellow and green: Stratigraphic distribution of Rhabdinopora in Belgium. a1. Rhabdinopora praeparabola, a2, R. f. parabola by Erdrmann (1986); b1, R. f. flabelliformis; b2, R. f. socialis; b3, R. f. anglica by Lecompte (1948), Graulich (1963) & Geukens (1950, 1954); c. R. f. norvegica by Graulich (1961); d, R. f. parabola in this paper; e, R. f. belgica [=R. f. parabola] by Bulman (1970); f, R. f. socialis by Bulman (1950a); g1, R. f. parabola, ?R. praeparabola, R. f. socialis and R. f. flabelliformis in locality La Roche; g2, R. praeparabola, R. f. parabola, R. f. flabelliformis, D. f. norvegica and D. f. socialis in locality Genappe 78; g3, R. f. norvegica, R. f. flabelliformis in locality Chastres 106; g4., R. f. anglica and R. f. norvegica in Loc. Genappe 26 by Lecompte (1949). The Time-Slices (TS) by Webby et al. (2004) are also listed in the correlation charts for comparison.
4.2. Brabant Massif
19All the specimens from the three fossil-yielding localities in the Brabant Massif recorded by Lecompte (1949) are re-investigated in the present study. At the locality La Roche, the association of graptolites includes R. praeparabola, R. f. parabola, R. f. socialis and R. f. flabelliformis. This assemblage does not contribute to a more precise biostratigraphy. At the locality Genappe 78 (near the Moulin of Chevlipont), R. praeparabola, R. f. parabola, R. f. flabelliformis, R. f. norvegica and R. f. socialis are identified, suggesting that these strata are comparable with the levels of the R. f. parabola Biozone plus the Anisograptus matanensis Biozone. At the locality Chastres 106, R. f. norvegica, R. f. flabelliformis are collected, which suggests levels around the A. matanensis Biozone in comparison to the stratotype section in Newfoundland. At the locality Genappe 26, the graptolite association of R. f. anglica and R. f. norvegica suggests that this part of the Chevlipont Formation can be roughly correlated with the upper part of the Solwaster Member (Sm1a) and the lower part of the Spa Member (Sm1b) in the Stavelot Massif (Fig. 2).
4.3. Condroz Inlier
20The Tremadocian graptolites recorded by Graulich (1961) in the Condroz Inlier range from depth 301.65 m to 307.50 m in the Chevlipont Formation in the Wépion borehole. At the level 306.32 m, R. f. norvegica (original fossil number 133, original identification as R. f. aff. norvegica) was recorded and this R. f. norvegica-bearing level was compared to the upper sub-zone of the British Dictyonema Zone by Graulich (1961). The careful re-investigation of the borehole indicates another Rhabdinopora occurrence at the level 302.00 m. These latter specimens clearly belong to R. f. parabola. The existence of this taxon suggests that this interval is most comparable with that of the R. f. parabola-bearing strata in the Solwaster section examined by Erdtmann (1986). In the Wépion borehole, the fact that specimens of R. f. parabola appear in a horizon (302.00 m) higher than that of R. f.norvegica (306.32 m) confirms the interpretation of an inverted sequence of strata in this part of the borehole caused by Variscan tectonic movements.
21The present re-investigations of the fossil records of the Rhabdinopora flabelliformis faunas of Belgium therefore show that three early Tremadocian graptolite biozones can roughly be recognized, in ascending order the R. praeparabola, the R. f. parabola and the R. f. anglica biozones. Although the exact boundaries between each biozones are still unclear, their relationships with rock formations are clear now. In the Brabant Massif, the boundary between the Mousty Formation and the Chevlipont Formation is located in the R. praeparabola Biozone. In the Stavelot Inlier, the boundary between the Solwaster Member (Sm1a) and the Spa Member (Sm1b) of the Jalhay Formation is located within the R. f. anglica Biozone (Fig. 2).
5. Stratigraphical distribution of the early Tremadocian Rhabdinopora flabelliformis fauna and their international correlation
22After acquiring the skills for flotation, the Rhabdinopora flabelliformis fauna soon expanded worldwide in lower Tremadocian strata. This fauna is widely considered as the first appearance of the earliest anisograptids (R. wutingshanensis Mu, 1955). Discovered even earlier in the uppermost Cambrian in North China, the fauna is thought to be the earliest occurrence of planktic dictyonemids, although these uppermost Cambrian findings have not been confirmed from other localities outside China so far. The Rhabdinopora flabelliformis fauna is very useful in terms of biostratigraphy because its appearance is considered to be particularly useful for recognizing the base of the Ordovician System at a global level. The two subspecies, R. f. parabola and Rhabdinopora flabelliformis anglica have been adopted as the index taxa in two nominative graptolite biozones of the early Tremadocian. R. f. anglica is an unrestricted outer-shelf to slope form while R. f. parabola is a restricted slope form, according to Cooper et al. (1998) and Cooper (1999). Another Rhabdinopora species, R. praeparabola, one of the earliest planktic graptolite species, is also the index fossil of the lowest Tremadocian graptolite biozone (Fig. 3).
23Figure 3. Occurrences of the Rhabdinopora flabelliformis fauna in selected key sections. A. Egenhoff et al., 2004; B. Lindholm, 1991; C. Maletz et al., 2010; D. Maletz & Egenhoff, 2001; E. Wang & Wang, 2001; F. Williams & Stevens, 1991; G. Cooper et al., 1998; H. Bulman & Rushton, 1973; I. Cooper et al., 1995; J. Loydell, 2012; K. Zeballo et al., 2008; L. Ortega & Albanesi, 2005; M. Gutiérrez-Marco & Esteban, 2005; N. Monteros & Moya, 2011. O. Zhang et al., 2005; P. Wang & Erdtmann, 1987; Q. Egenhoff et al., 2004; 1, (orange) the Mousty Formation; 2, (dark green) the Solvaster Member (Sm1a); 3, (light green) the Spa Member (Sm1b); 4. (yellow) the Chevlipont Formation. TS: Time-Slices following Webby et al. (2004).
24In addition to the GSSP at Green Point, Newfoundland, several other sections can be considered as key strata for international correlation, because these sections allow a graptolite biozonation of the earliest part of the Ordovician (Fig. 3).
25For example, Bulman (1950b) reported D. f. canadense, D. clarki, D. rusticum, D. cristatum, D. cyathiforme and D. lapworthi from collections in the fossiliferous Dictyonema Shales of Matane, and from the south shore of the St. Lawrence River, both in Québec, Canada. The association of Rhabdinopora with Anisograptus and Staurograptus suggested a Tremadocian age, but no precise indication of age could be carried out at that time.
26In Newfoundland, Canada, four lower Tremadocian graptolite biozones have been set up in the Green Point section, they are in ascending order: the R. praeparabola, R. f. parabola, A. matanensis and R. f. anglica zones (Cooper, 1999; Cooper et al., 2001). In the R. praeparabola Biozone, only the nominative species and Staurograptus dichotomus are found ranging through the zone. The R. f. parabola zone is characterized by the widespread appearance of the index taxa. Near the top of the zone, R. f. canadensis and R. f. socialis appear. In the A. matanensis Biozone, R. f. canadensis was soon replaced by R. f. flabelliformis. Near the top of the biozone, R. f. norvegica is quite abundant and it ranges into the following R. f. anglica Biozone.
27In the Nærsnes section, south of Oslo, Norway, R. praeparabola, R. f. socialis, R. f. parabola and R. f. flabelliformis successively appear in the early Tremadocian (Bruton et al., 1982).
28In Bolivia, the R. flabelliformis Biozone represents a broad interval, although R. f. parabola and R. f. ssp. can be identified, but no finer subdivision is possible. This R. flabelliformis Biozone is correlated with the four lower Tremadocian biozones in Eastern North America and Scandinavia (Egenhoff et al., 2004; Maletz et al., 2010).
29In the Deanshanger borehole near Buckingham, central England, several subspecies of R. flabelliformis, including R. f. patula, were found below levels with the genus Adelograptus. This suggests a Tremadocian age (Bulman & Rushton, 1973). These R. flabelliformis-bearing rocks are broadly correlated with R. f. parabola, A. matanensis and R. f. anglica biozones in Baltica by Loydell (2012). In Australia, the Lancefieldian 1a Zone includes R. scitulum, R.? enigma and two other species in the Anisograptus Biozone. They can be correlated roughly to lower Tremadocian biozones in North America and Scandinavia (Cooper & Stewart, 1979; Cooper, 1999; Egenhoff et al., 2004).
30In the Dayangcha section of Jilin Province, North China, the earliest Ordovician planktonic graptolites, R. f. parabola, R. flabelliformis, R. f. socialis are found together with some quadriradiate and triradiate taxa in Staurograptus and Anisograptus (Lin in Chen et al., 1986). Later, Wang & Erdtmann (1987) reported the association of graptolites R. parabola, R. flabelliformis s.l. (Eichwald) and R. socialis in the same section and recognized the R. parabola Zone. Wang et al. (1998) restudied the earliest Ordovician graptolites from the same section and made some taxonomic revisions of the species that were previously studied by Chen et al. (1986). In the latter study, R. cf. praeparabola and R. f. parabola were described and a population study of R. f. parabola was carried out. In South China, early planktic graptolites such as R. praeparabola, R. f. parabola, R. f. flabelliformis, R. ganbeiensis and R. f. anglica have been reported in several localities in Jiangxi, Anhui and Hunan Province, resulting in the recognition of the R. f. parabola and R. f. anglica biozones (Wang & Wang, 2001; Zhang et al., 2005; Wang et al., 2014; Wang & Muir, 2015).
31In Argentina, three early Tremadocian graptolite chronozones have been recognized from the Bordo Atravesado Formation in the Famatina System, NW Argentina, in ascending order the R. f. parabola, theAnisograptus matanensis and the R. f. anglica chronozones. Early planktonic graptolite taxa such as R. f. flabelliformis, R. f. anglica, R. f. socialis, R. f. ?bryograptoides, R. cf. enigma, Anisograptusmatanensis, ?Bryograptus sp. and R. f. famatinensis are reported, together with the newly recovered Rhabdinopora turneriand R. flabelliformis acenolazai (Zeballo et al., 2005; Ortega & Albanesi, 2005; Gutiérrez-Marco & Esteban, 2005).
32Figure 3 summarizes the biozonation schemes of the different parts of the world. Based on international correlations, it appears now possible to attribute the lowermost part of the Belgian Rhabdinopora flabelliformis faunas (R. praeparabola) to the earliest Tremadocian (earliest part of Time-Slice 1a, Webby et al., 2004). These levels are present both in the Brabant Massif and in the Stavelot Inlier. The Chevlipont Formation of both the Condroz Inlier and the Brabant Massif spans Time-Slice 1a (but not the lowermost part) and also Time-Slice 1b. The Solwaster Member is attributed to Time-Slice 1a, while the Spa Member is corresponding to the upper part of Time-Slice 1a and Time-Slice 1b.
6. Systematic Palaeontology
33High-level taxonomy herein follows Maletz et al. (2009) and Maletz (2014). Open nomenclature is used following the recommendations of Matthews (1973) and Bengtson (1988). The format of the synonymy list follows the recommendations of Matthews (1973).
34The following abbreviations are used in the synonymy lists:
35?: determination questionable;
36.: determination correct and is accepted in this study;
37non: determination not accepted;
38*: valid under the term of ICZN;
39v: vidimus;
40p: partim;
411999 (Year in italics): this publication mentions a species, but without description or illustration (impossible to control the determination);
421999 (Year in Roman): this work contributes to our knowledge of this species.
43Phylum Hemichordata Bateson, 1885
44Class Pterobranchia Lankester, 1877
45Subclass Graptolithina Bronn, 1849
46Order Graptoloidea Lapworth in Hopkinson & Lapworth, 1875
47Suborder Graptodendroidina Mu & Lin in Lin, 1981
48Family Anisograptidae Bulman, 1950b
49Genus Rhabdinopora Eichwald, 1855
50Type species: Rhabdinopora flabelliformis (Eichwald, 1840)
51Remark: Hall (1851) erected the genus Dictyonema and designated Gorgonia retiformis Hall, 1843 as the lectotype (hypotype). This proximally incomplete specimen from the Rochester Shale of the Wenlock (Silurian) was hypothetically suggested as a benthonic root-bearing taxon by judging from the associated graptolites. In the following one and a half centuries, more than 200 graptolite taxa have been assigned to the genus Dictyonema s.l. making it a largely used genus. Due to the relatively large size, most of the meshworked Dictyonema specimens are incomplete and thus increase the difficulty of identification. Westergård (1909) and Bulman (1927) well perceived a separation of sessile and planktic dictyonemids based on the proximal structure. Finally, Erdtmann (1982) suggested that the planktic ‘Dictyonema’ with definite nema or buoyancy structures should be split from the benthic dendroids and regarded as early taxa of the Anisograptidea Bulman, 1950b. In his study, Erdtmann (1982) transferred 28 coni-siculate planktic ‘Dictyonema’ taxa to the planktonic nema-bearing genus Rhabdinopora Eichwald, 1855 with Rhabdinopora flabelliforme (Eichwald, 1840) as the type species. According to ICZN rules, Gorgonia retiformis Hall, 1843 remains in the benthic Dictyonema s.s. Questions addressing this taxonomic splitting rise mainly from the hypothetical benthonic ecology of the type species in Dictyonema s.l. Legrand (1985) insisted that siculate planktic ‘dictyonemids’ can be separated from Dictyonema s.l. only after well-preserved supplementary materials of the type species, Gorgonia retiformis Hall, 1843 have been collected from the exact layer in the type locality. This interpretation was accepted by several Chinese colleagues, who prefer to retain Dictyonema s.l. before detailed examinations are carried out on Gorgonia retiformis (Mu et al., 2002).
52More than 140 graptolite species and subspecies are definitely rooted benthic ‘dictyonemids’ and the remaining approximately 80 taxa are siculate planktic ‘dictyonemids’ (Erdtmann, 1982). The latter mainly range from the earliest Tremadocian to the early Ordovician. Erdtmann (1982) assigned more than twenty species and subspecies to Rhabdinopora. If one includes into this genus also the two recently described species from Argentina (R. turneri Gutierrez-Marco & Esteban, 2005 and R. f. acenolazai Gutierrez-Marco & Esteban, 2005), the Rhabdinopora flabelliformis cluster would occupy more than 40% of the Rhabdinopora taxa. These are mainly of an early Tremadocian age. Although disputes of the genus name for siculate planktic ‘dictyonemids’ rose from time to time, most researchers after Erdtmann (1982) accepted Rhabdinopora as the genus name for these characteristic graptolite species. In the present paper, we also use this generic classification method for the Belgian graptolite material.
53The number of primary stipes in Rhabdinopora has long been a matter of dispute. This is because the details of primary stipes could be concealed by one another if the specimens are preserved in lateral view. Presence of a quadriradiate proximal end has been confirmed in isolated specimens (Legrand, 1974). R. praeparapola, R. parabola, R. rustica, R. f. flabelliformis, R. f. parabola, R. f. canadensis and R. f. anglica have been confirmed display a quadriradiate proximal part (Erdtmann, 1988; Cooper et al., 1998). Cooper et al. (1998) suggested this genus to be characterized by four primary stipes and having a quadriradiate proximal development pattern as illustrated by Maletz (1992). Generally, the species in this genus can be recognized by their characteristic appearance of a meshwork and the siculate proximal end.
54Rhabdinopora praeparabola Erdtmann, 1982
55(Fig. 4 A-C)
56?1977 Dictyonema taojiangense Jin & Wang: 78-79, pl.16, figs 12-13, text-fig. 3, 1-3.
57*.1982 Rhabdinopora? praeparabola Erdtmann: 136-138, pl. I, figs 6, 8-10; pl. II, fig. 2?
58(See more synonyms of Rhabdinopora?praeparabola before 1982 in Erdtmann, 1982)
591985 Dictyonema proparabola Lin in Chen et al.: 238-240, pl. 62, figs 1-8; pl. 65, fig. 11, text-fig. 95.
601985 Dictyograptus flabelliformis cf. anglicus Bulman; Yu et al.: 22, pl. III, figs 4, 5.
61.1986 ?Rhabdinopora praeparabola Erdtmann; Erdtmann: 78, pl. 1, fig. 1, text-fig.1.
62.1988 ?Rhabdinopora praeparabola Erdtmann; Erdtmann: 335-338, figs 6A-D, 7A-D.
63.1998 Rhabdinopora praeparabola Erdtmann; Cooper et al.: 10-12, figs 8, 9a-f, 10b, c, e, f, 16a.
64cf.1998 Rhabdinopora cf. praeparabola Erdtmann; Wang et al.: 223, pl. I, figs 1-3.
651999 Rhabdinopora praeparabola Erdtmann; Feng & Erdtmann: 21-24.
66Material. Four specimens. Two of them are recovered from the ‘Chemin dans le bois de l’Ermitage vers le Moulin de Chevlipont’ (path in the ‘Ermitage’ wood towards the Chevlipont Mill) in the Brabant Massif; another two are from the locality ‘La Roche-Brabant, ‘Chemin de la carrière du vallon du ruisseau de Sainte Gertrude’ (the quarry path in the valley of the brook St. Gertrude) of La Roche in the Brabant Massif.
67Description. The relatively small rhabdosome is pendent to declined, measuring 18-21 mm in length and around 16-19 mm in width. There are four primary stipes, each are loosely flexuous and sinuous, they are relatively well spaced, with an average width of 0.4 mm in lateral aspect, separated by interspaces far wider than the stipes, which number generically 1-12 in 10 mm. The shape of the meshes is variable and irregularly spaced, for scattered dissepiments are generally present in the distal part of specimens. The observed siculae in the specimens are straight, of about 1.1 mm long and 0.2 mm wide (Fig. 4 C).
68Discussion. Cooper et al. (1998) noted that R. praeparabola is a species that contains variable forms but they all share some key characters, such as a relatively small rhabdosome, few dissepiments or none at all and loose flexuous stipes. Early planktonic Rhabdinopora with these characters can be attributed to this species. Gutiérrez-Marco & Esteban (2005) reported R. turneri from the A. matanensis Biozone in Argentina. R. turneri shows some similarity with R. praeparabola (sparse stipes and dissepiments) and is regarded as the reminiscent of R. praeparabola. R. turneri differs from R. praeparabola in having a pendent rhabdosome and wider stipes. R. praeparabola has incomplete dissepimental notches or no dissepiments which can be confused with mature Staurograptus dichotomous specimens. R. praeparabola can be separated from the latter by its pendent, robust sinuous stipes and relatively longer sicula.

69Figure 4. A, B, C, Rhabdinopora praeparabola: narrow-cone specimens displaying 4 primary stipes, A, IRSNB a12903, B, C, IRSNB a12904; D, F, Rhabdinopora flabelliformis norvegica: semi-discal specimens characterized by the coarse meshwork, D, IRSNB a12905, F, IRSNB a12907; E, G, H, Rhabdinopora flabelliformis flabelliformis: Polygon in H shows magnificated part in Figure 4 J, E, IRSNB a12906, G, IRSNB a12908, H, IRSNB a12909; I, Rhabdinopora flabelliformis parabola: specimen with divided nema/fibre structure, I, IRSNB a12910; J. magnification of Fig. 4 H: tri-radiate incomplete dissepimental notches from a stipe, IRSNB a12908. Scale bars in C, D and F represent 5 mm, scale bar in J represents 1 mm, others represent 10 mm.
70Rhabdinopora flabelliformis flabelliformis (Eichwald, 1840)
71(Fig. 4 E, G, H)
721840 Gorgonia flabelliforme Eichwald: 207.
73*1882 Dictyonema flabelliforme forma typica, Brögger: 31-36, 12, figs 17-19.
741904 Dictyonema flabelliforme (Eichwald); Ruedemann: 599, pl. 1, figs 1-22, text-figs 26, 27.
751927 Dictyonema flabelliforme (Eichwald); Bulman: 12-16, figs 6c-d, 12-14. pl. 1, figs 2, 4; pl. 2, figs 3-4.
76.1954 Dictyonema flabelliforme flabelliforme (Eichwald); Bulman: 14-17, pl. 2, figs 1-5, text-figs 4a-d.
771966a Dictyonema flabelliformis (Eichwald); Bulman: 296, text-fig. 4.
78.1966b Dictyonema flabelliformis (Eichwald); Bulman: 409, text-fig. 2a, b?.
79.1971 Dictyonema flabelliforme flabelliforme (Eichwald); Bulman & Størmer: fig. 1 A-D; fig. 2; fig. 5A-D.
80aff. 1973 Dictyonema flabelliforme aff. flabelliforme (Eichwald); Bulman & Rushton: 18-20, pl. 1, figs 1-4, pl. 2, fig. 4.
811982 Rhabdinopora flabelliformis (Eichwald); Erdtmann: 129-136, pl. 2, figs 4-6, 9.
821983 Dictyonema flabelliforme flabelliforme (Eichwald); Zhao: 374-375; pl. 1, figs 6-8.
83.1985 Dictyonema flabelliforme (Eichwald); Lin in Chen et al.: 57, pl. 3, fig. 8; pl. 4, fig. 3.
84.1986 Dictyonema flabelliforme (Eichwald); Lin in Chen et al.: 232, pl. 57, figs 1-7; pl. 58, figs 1-5; pl. 59, fig. 1; text-figs 92, 93.
851998 Rhabdinopora flabelliformis flabelliformis (Eichwald); Cooper et al.: 12-13, figs 6c, 7.
86.1999 Rhabdinopora flabelliformis flabelliformis (Eichwald); Feng & Erdtmann: 21-24.
87.2001 Rhabdinopora flabelliformis flabelliformis (Eichwald); Wang & Wang: 25, fig. 8a.
88.2003 Rhabdinopora flabelliformis flabelliformis (Eichwald); Gutiérrez-Marco & Esteban: figs 2y, 3f.
89.2005 Rhabdinopora flabelliformis flabelliformis (Eichwald); Gutiérrez-Marco & Esteban: 78-80, figs 4a-b, 14a-c.
90Material. 14 specimens, mostly from the Brabant Massif: one specimen is from La Roche, trench between 36.0 and 36.3 km along the railway line from Ottignies to Charleroi; seven specimens come from the path in the ‘Ermitage’ wood towards the Chevlipont Mill; four specimens from the locality La Roche, new road towards the West (‘Chastres 106’). The material includes two specimens from the Stavelot Inlier, locality Lorce, roadcut from Naze to Chession, 600 m south of the Amblève River.
91Description. Narrow to broad conical rhabdosome, with a divergence angle near the apex ranging from 50 to 105°. The rhabdosome measures 6-30 mm in length and around 4-25 mm in width. The meshwork generally possesses 7 to 9 stipes in an interval of 10 mm, with a mean value of 8 stipes. In some well-preserved specimens the autothecae, bithecae and stoloenthecae can also be observed. The stipes range from 0.4 to 0.5 mm in lateral width and they are generally straight to moderate in sinuosity. The dissepiments number 3-7 in 10 mm. The dissepiments are horizontal or moderately inclined and widely spaced in the proximal part of the rhabdosome and more closely spaced in the mature part of rhabdosome. The dissepiments are usually delicate, the bases pass gradually into the stipe walls.
92Remarks. Eichwald (1840) erected Gorgonia flabelliformis based on an incomplete specimen from the Tonschiefer of Baltischport in Estonia. The description is brief and not diagnostic, no particular specimens were identified and no illustration was given. Considerable confusion was caused by the vagueness of this description. The specimen figured by Brögger (1882) which was regarded as the forma typical of the species was generally accepted (see more discussion about the history of this species in Cooper et al., 1998, p. 12).
93Several subspecies have been regarded as the synonyms of R. f. flabelliformis. R. f. graptolithina (Kjerulf, 1865) and R. f. norvegica (Kjerulf, 1865). They were treated as junior synonyms of R. flabelliformis (Eichwald) by Erdtmann (1982). Cooper et al. (1998) preferred to maintain the name of R. f. norvegica which was used for over a century. R. f. rustica which has a greater width is provisionally included as the immature growth stage of R. f. flabelliformis. In the present study, R. f. norvegica is retained. R. f. flabelliformis can be distinguished from R. f. patula (Bulman & Rushton, 1973) by its smaller size and loosely spaced dissepiments. Cooper et al. (1998) suggested that R. f. flabelliformis can be distinguished from R. f. anglica by its slightly dense stipe spacing. The boundary between the two subspecies could be 7 stipes in 10 mm. The former has more stipes than 7, while the latter has less stipes.
94Rhabdinopora flabelliformis anglica (Bulman, 1927)
95(Fig. 6 F)
96* 1927 Dictyonema flabelliforme anglica Bulman: text-figs 16, pl. 1, figs 1, 3, 5-8, pl. 2, figs 5-8.
97. 1954 Dictyonema flabelliforme anglicum Bulman; Bulman: pl. 4, figs 3, 5, text-fig. 5a.
98.1971 Dictyonema flabelliforme anglica Bulman; Bulman & Størmer: fig. 3A-C.
99. 1982 Rhabdinopora flabelliforme anglica (Bulman); Erdtmann: 129.
1001983 Dictyonema flabelliforme anglica (Bulman); Yang et al.: 355, pl. 134, fig. 6.
101v. 1984 Dictyograptus taoyuanensis Bulman; Liu: pl. II, fig. 1a-c, fig. 2.
102non 1984 Dictyograptus flabelliforme anglica (Bulman); Liu: 39, pl. II, fig. 5, text-fig. 1.
103. 1998 Rhabdinopora flabelliforme anglica (Bulman); Cooper et al.: figs 5, 6d, 19, 20, 21e.
104. 2001 Rhabdinopora flabelliforme anglica (Bulman); Wang & Wang: fig. 7.
1052003 Rhabdinopora flabelliformis anglica (Bulman); Gutiérrez-Marco & Esteban: fig. 2.
106.2005 Rhabdinopora flabelliformis anglica (Bulman); Gutiérrez-Marco & Esteban: figs 14h-j.
107Material. Eight specimens. One specimen comes from the Brabant Massif, locality ‘Rid’Hez, Chemin à l’Est du ruisseau Rid’Hez ou Trou du Bois, talus S (Genappe 26)’; seven specimens from the Stavelot Massif; four specimens are from Spa, road to Sart (Stavelot Inlier); one specimen is from the ‘north shore of Lake Gileppe between the first 2 East Points (Pl. Limbourg)’; two specimens are from Lorce, roadcut from Naze to Chession, 600 m south of the Amblève River.
108Description. The rhabdosome is narrowly conical. The branches are relatively well spaced, stipes have an average width of 0.7 mm in lateral aspect, separated by interspaces far wider than the stipes, which number only 7-8 in 10 mm. The dissepiments are extremely irregular and rather infrequent, they are slender and straight, normally perpendicular to the stipes, and with an average width of 0.1 mm in lateral view, usually numbering only 3-4 in 10 mm. The bases of the dissepiments pass gradually into the stipe walls, these dissepiments appear darker in comparison with adjacent stipes. The sparse dissepiments and widely spaced branches make the appearance of meshwork in R. f. anglica characteristically open.
109Rhabdinopora flabelliformis parabola (Bulman 1954)
110(Fig. 4 I; Fig. 5 E, G, H; Fig. 6 C, D)
111*.1954 Dictyonema flabelliforme parabolus Bulman: 25-27, pl. 1, figs 1-5, text-fig. 9.
1121970 Dictyonema flabelliforme belgica Bulman: 215-218, pl. 1, figs 2-4, 6, 7; text-figs 1-5.
113.1982 Rhabdinopora parabola (Bulman); Erdtmann: pl. 2, figs 1-2.
1141983 Dictyonema flabelliforme belgicum Bulman; Zhao: 375, pl. 2, figs 2-7.
115.1985 Dictyonema flabelliforme parabolus Bulman; Lin in Chen et al.: 58, 59, pl. 1, figs 1, 4; pl. 3, figs 1, 6, 7; pl. 4, figs 1, 2, 4; pl. 7, figs 1, 2.
116.1985 Dictyograptus flabelliformis parabolus (Bulman); Yu et al.: 21, 22, pl. 3, figs 10, 11.
117.1985 Dictyograptus flabelliformis belgicus (Bulman); Yu et al.: 28, pl. 3, figs 1, 2.
118.1986 Rhabdinopora parabola (Bulman); Erdtmann: 80-82, pl. 1, figs 2-5, text-fig. 2.
119.1988 Rhabdinopora parabola (Bulman); Erdtmann: 338-339, figs 7E-F.
1201988 Dictyonema flabelliformis parabola-belgicum; Lin: 321-322, pl. I, figs 1, 2, 4-6; pl. II, fig. 2-4, 6; pl. III, figs 1, 3, 5, 6; pl. IV, figs 1, 3; text-fig. 3a, b, c; text-figs 4a, c, d, h-k; text-figs 8a, b; text-figs 9a, b.
121.1998 Rhabdinopora flabelliformis parabola (Bulman); Wang et al.: 223-224, pl. I, fig. 5-8; pl. II, fig. 1-6; pl. III, fig. 1-3.
122.1998 Rhabdinopora flabelliformis parabola (Bulman); Cooper et al.: 14-15, figs 6a, 9g-p, 10a, d, g, 11-15, 16b-o, 25g, h, j.
123(See more synonyms list of R. f. parabola in Cooper et al., 1998, p. 14)
1241999 Rhabdinopora flabelliformis parabola (Bulman); Feng & Erdtmann: 21-24.
125.2000 Rhabdinopora flabelliformis parabola (Bulman); Landing et al.: 488, fig. 3.
126.2001 Rhabdinopora flabelliformis parabola (Bulman); Cooper et al.: 25, fig. 7l; fig. 8b, c
127.2001 Rhabdinopora flabelliformis parabola (Bulman); Wang & Wang: fig. 3A, B.
128Material. 68 specimens. 50 specimens come from the path in the ‘Ermitage’ wood towards the Chevlipont Mill (Brabant Massif); three specimens are from the locality ‘La Gleize 6’, 650 meters west of the Church and 15 specimens are from the surrounding areas of Spa (both in the Stavelot Inlier).
129Description. Rhabdosome with a parabolic outline. The divergence angle near the apex is around 90-145°. The rhabdosome length spans from 14 to 71 mm and the width varies from around 14 to 47 mm. The stipe width ranges from 0.35 to 0.6 mm in dorsal and lateral view and they are generally straight to moderate in sinuosity. Interspaces are around twice the stipe width. The stipe spacing ranges from 9 to 12 in 10 mm with a mean value of 10. Dissepiments are horizontal to moderately inclined and close to very closely spaced in the mature part of the rhabdosome. The dissepiments are extremely irregular and rather frequent, they are slender and straight, normally perpendicular to the stipes, and with an average width of 0.1 to 0.2 mm, usually numbering 6-8 in 10 mm (Fig. 5 E). Nematic structures like buoyancy structures are present (Fig. 4 I, Fig. 6 D).
130Remarks. Bulman (1954) noted that the variances between different specimens of the earliest planktic R. flabelliformis clusters are quite bigger due to different preservation conditions. Thus, population studies of these earliest planktonic graptolites would provide more comprehensive understandings (Cooper et al., 1998; Wang et al., 1998). This species could be distinguished from other species by its parabolic overall shape, a relatively smaller individual, irregular meshwork and well-developed fibres. R. f. belgica (Bulman, 1950a) was erected for its characteristic bundle of fibres at the apex of the rhabdosome in place of a single nema. In the Dayangcha section, some specimens from the R. f. parabola population also have sheafed nematic fibres and bundles around the sicula (Lin, 1988; Wang et al., 1998). Thus, the difference of fibres is of limited diagnostic value and R. f. belgica is here considered to be junior synonym of R. f. parabola.

131Figure 5.A, Rhabdinopora flabelliformis norvegica-flabelliformis transients, widely to moderately spaced dissepiments inserted by rounded, coarse meshwork caused by cortical thickening, A, IRSNB a12911; B, C, D, Rhabdinopora flabelliformis norvegica: meshwork formed by dissepiments and adjacent stipes; B, D: rounded, coarse meshwork caused by cortical thickening ; C: deformed meshwork showing tilted dissepiments and narrow intervals between adjacent stipes, B, IRSNB a12912, C, IRSNB a12913, D, IRSNB Vb 155W0273-306.32; F, Rhabdinopora flabelliformis socialis: note oblique and densely set dissepiments connecting adjacent stipes, F, IRSNB a12915; E, G, H, Rhabdinopora flabelliformis parabola: E: adjacent dissepiments cohere to each other laterally and form a membrane-like structure. G: moderately inclined and widely to moderately spaced dissepiments; H: juvenile rhabdosome with a parabolic outline, E, IRSNB a12914, G, IRSNB a12916, H, IRSNB a12917; scale bars in B, C, D and E represent 5 mm, scale bars in F represents 1 mm, others represent 10 mm.
132Rhabdinopora flabelliformis socialis (Salter, 1858)
133(Fig. 5 F; Fig. 6 A, E)
134*1858 Graptopora (Fenestella) sociale Salter: 65.
1351866 Dictyonema sociale (Salter); Salter: 331, figs 1a-c.
136.1927 Dictyonema flabelliformis sociale Salter; Bulman: 26-28, pl. 2, figs 1, 2.
1371940 Dictyonema flabelliformis sociale Salter; Størmer: 163, figs 6-8.
138.1954 Dictyonema flabelliforme sociale Salter; Bulman: 27-29, pl. 1, figs 1, 2; text-fig. 10a.
1391962 Dictyonema flabelliforme sociale Salter; Li & Chen: 11, pl. 1, figs 1, 2.
140aff. 1973 Dictyonema flabelliforme aff. Sociale Salter; Bulman& Rushton: 20, fig. 2k.
1411977 Dictyonema flabelliforme sociale Salter; Jin & Wang: 77, 78, pl. 16, figs 1-11.
1421983 Dictyonema flabelliforme sociale Salter; Zhao: 374, pl. 1, fig. 1a-c, fig. 5.
1431983 Dictyonema flabelliforme sociale Salter; Yang et al.: 355-356, pl. 131, fig. 12.
1441985 Dictyonema flabelliforme sociale Salter; Lin in Chen et al.: 59-60, pl. 3, figs 2, 3; pl. 5, figs 1-3; pl. 8, fig. 2.
1451988 Dictyonema flabelliformis socialis Salter; Lin: 233, pl. I, figs 3, 7; pl. II, figs 5, 7; pl. IV, fig. 6; text-fig. 7d, e; text-fig. 9c.
1461998 Rhabdinopora flabelliformis socialis (Salter); Cooperet al.: 18.
147cf. 1999 Rhabdinopora flabelliformis cf. socialis (Salter); Tortello & Esteban: 383-385, figs 6A-6C, 6E.
1482003 Rhabdinopora flabelliformis socialis (Salter);Gutiérrez-Marco & Esteban: 40, fig. 2.
1492005 Rhabdinopora flabelliformis socialis (Salter);Gutiérrez-Marco & Esteban: 85, fig. 14d-e.
150Material. 16 specimens. Most are from the Stavelot Inlier: two specimens are from the Gileppe dam; two specimens come from the locality La Gleize; nine specimens are from the north shore of Lake Gileppe between the first 2 East Points ‘(Pl. Limbourg)’; two specimens are from Lorce, roadcut from Naze to Chession, 600 m south of the Amblève River. Only one specimen comes from the Brabant Massif, from the railway trench between 36.0 and 36.3 km along the railway line from Ottignies to Charleroi.
151Description. The rhabdosome is broadly to narrowly conical in outline. The divergence angle near the apex is around 60-105°. The rhabdosome is about 33.1-76 mm in length and around 18-33.6 mm in width. The ratio between length and width is around 2. Stipes branch dichotomously, producing 17 stipes at 10 mm from the sicula and more than 35 at the distal limit. Stipe dorsal width ranges from 0.3 to 0.4 mm and stipes are generally straight to moderate in sinuosity. The mesh pattern is characterised by a quite regular distributionof stipes and dissepiments, the stipes space closely and parallel to each other with a number of 10-13 in 10 mm. The dissepiments are extremely densely spaced as they number 15-20 in 10 mm. The dissepiments are slender and delicate, ca. 0.1 mm wide and 0.7-0.8 mm long, mostly straight, and perpendicular to the stipes. The thickness of the dissepiments seems to be regular. The dissepiments appear darker in comparison with adjacent stipeswhich may reflect a primary structural/compositional differentiation.
152Remarks. This species is characterized by the closely spaced stipes (usually 10-13 in 10 mm) and the fine densely packed dissepiments (15-20 in 10 mm). In the Green Point section, this species first appeared at about the same layer as R. f. canadensis, in the uppermost R. f. parabola Zone (Cooper et al., 1998). R. f. canadensis differs from Dictyonema flabelliformis socialis in having apertural spines of the autothecae, its spines have forked, Y-shaped terminations.

153Figure 6. A,E, Rhabdinopora flabelliformis socialis: mature specimens with densely distributed dissepiments and stipes, A, IRSNB a12918, E, IRSNB a12915 ; B, Rhabdinopora flabelliformis norvegica: characterized by the rounded, coarse meshwork caused by cortical thickening, B, IRSNB a12919; C, D, Rhabdinopora flabelliformis parabola: specimen with divided nema/fibre structure, C, IRSNB a12920, D, IRSNB a12921; F, Rhabdinopora flabelliformis anglica: mature specimens with loosely distributed dissepiments and stipes, F, IRSNB a12922. All scale bars represent 10 mm.
154Rhabdinopora flabelliformis norvegica (Kjerulf, 1865)
155(Fig. 4 D, F; Fig. 5 B-D; Fig. 6 B)
156*1865 Dictyonema norvegicum Kjerulf: 1, fig. 1a, b (non. figs 2, 3).
1571882 Dictyonema flabelliformis mut. norvegicum Kjerulf; Brӧgger: 36.
1581909 Dictyonema flabelliformis norvegicum Kjerulf; Westergård: 60, pl. III, figs 8, 9.
1591912 Dictyonema flabelliformis norvegicum Kjerulf; Hahn: 139, pl. XX.
160.1954 Dictyonema flabelliformis norvegicum Kjerulf; Bulman: 23-25, pl. 3, fig. 1-3, ?5; text-fig. 8a-d, ?e.
1611966a Dictyonema norvegicum Kjerulf; Bulman: 290-291, 294, text-fig. 1a-d, text-fig. 2, text-fig. 3.
162.1971 Dictyonema flabelliforme norvegica (Kjerulf); Bulman & Størmer: fig. 4A-C.
163.1998 Rhabdinopora flabelliformis norvegica (Kjerulf); Cooper et al.: 18.
164Material.19 specimens. 14 specimens come from the path in the ‘Ermitage’ wood towards the Chevlipont Mill (Brabant Massif); five specimens are from Lorce, road from Naze to Chession, 600 m south of the Amblève River (Stavelot Inlier).
165Description. The largest reminiscent of the rhabdosome suggests that the total length exceeded at least 77.9 mm long with a distal width of 56.7 mm. The length-width ratio is commonly higher than 1.3, with a rapid initial dichotomous branching. The mesh pattern is characterised by a quite regular distribution of stipes and dissepiments giving rise to sub-rounded fenestrulae. The meshwork generallypossesses 9-13 stipes in 10 mm. Dissepimentsjoin the stipes and show a characteristic pouch-like “balconies”morphology. The stipes are slightly undulating. The lateral stipe width ranges from 0.3 to 0.6 mm. The dissepiments are variable, sometimes 8 to 12 in 10 mm, rarely angular. Cortical overgrowth happened in the rhabdosome and dissepiments make the appearance of the meshwork in this species characteristically coarse and robust (Fig. 5 B, D).
166Remarks.This species is characterized by its rounded, coarse meshwork caused by cortical thickening. Even broken specimens can be identified according to the coarse meshwork.
7. Acknowledgements
167This work was financially supported by the National Natural Science Foundation of China (No. 41402010, 41372017), the Fundamental Research Funds for the Central Universities (No. 20620140384) and China Postdoctoral Science Foundation (No. 2014M550313). WWH acknowledges the palaeontology research team of the CNRS research unit Evo-Eco-Paléo (UMR 8198) that financed a post-doctoral stay at the University of Lille. This is a contribution to the French ANR ‘RALI’ project and to IGCP 591. Annelise Folie and Alain Drèze, from the Royal Belgian Institute of Natural Sciences (Brussels), are thanked for their assistance in the museum collection. Alain Herbosch (Université Libre de Bruxelles) and Jacques Verniers (Ghent University) are thanked for providing useful information on literature, regional geology and stratigraphy. Two reviewers, Jörg Maletz (Berlin) and Jacques Verniers (Ghent University) are thanked for their useful comments that greatly improved the manuscript.
8. References
168Althausen, M., 1992. Lower Paleozoic (Riphean) metalliferous black shales. Oil Shale, 9 (3), 194-207.
169Bateson, W., 1885. The later stages in the development of Balanoglossus kowalevskii, with a suggestion as to the affinities of the Enteropneusta. Quarterly Journal of Microscopical Science, 25, 81-122.
170Bengtson, P., 1988. Open nomenclature. Palaeontology, 31(1), 223-227.
171Bronn, H.G., 1849. Index Palaeontogicus B, Enumerator Palaeontologicus. E. Schweizerbart’sche, Stuttgart, 980 pp.
172Brögger, W.C., 1882. Die Silurischen Etagen 2 und 3 im Kristianiagebiet und auf Eker. Universitäts program, Kristiania, 1-376.
173Bruton, D. L., Erdtmann, B.-D. & Koch, L., 1982. The Nærsnes section, Oslo Region, Norway: a candidate for the Cambrian-Ordovician boundary stratotype at the base of the Tremadoc Series. In Bassett, M. G. & Dean, W.T. (eds), The Cambrian-Ordovician boundary: sections, fossil distributions, and correlations. Cardiff: National Museum of Wales, Geological Series No. 3, 61-69.
174Bulman, O.M.B., 1927 (for 1925). A monograph of British dendroid graptolites. Part I. Palaeontographical Society Monograph, 1-28, plates I, II.
175Bulman, O.M.B., 1950a. On some Ordovician graptolite assemblages in Belgium. Bulletin de l’Institut Royal des Sciences naturelles de Belgique, 26 (5), 1-8.
176Bulman, O.M.B., 1950b. Graptolites from the Dictyonema shales of Quebec. Quarterly Journal of the Geological Society of London, 106, 63-99.
177Bulman, O.M.B., 1954. The graptolite fauna of the Dictyonema Shales of the Oslo region. Norsk Geologisk Tidsskrift, 33, 1-40.
178Bulman, O.M.B., 1966a. On the type material of Dictyonema norvegicum Kjerulf. Norsk Geologisk Tidsskrift, 46, 289-298.
179Bulman, O.M.B., 1966b. Dictyonema from the Tremadocian of Estonia and Norway. Geological Magazine, 103, 407-413.
180Bulman, O.M.B., 1970. A new Dictyonema fauna from the Salmien of the Stavelot Massif. Bulletin de la Société Belge de Géologie, de Paléontologie et d’Hydrologie,79 (3-4), 213-224.
181Bulman, O.M.B. & Størmer, L., 1971. Buoyancy structure in rhabdosomes of Dictyonema flabelliforme (Eichwald). Norsk Geologisk Tidsskrift, 51, 25-31.
182Bulman, O.M.B. & Rushton, A.W.A., 1973. Tremadoc Faunas from boreholes in Central England. Bulletin of the Geological Survey of Great Britain, 43, 1-40.
183Chen, J.Y., Qian, Y.Y., Lin, Y.K., Zhang, J.M., Wang, Z.H., Yin, L.M. & Erdtmann, B.D., 1985. Study on Cambrian-Ordovician boundary strata and its biota in Dayangcha, Hunjiang, Jilin, China, 1-138. China Prospect Publishing House, Beijing.
184Chen, J.Y, Erdtmann, B.D., Gong, W.I., Li, H.M, Lin, Y.K., Qian, Y.Y., Tao, X.C., Wang, Y.X., Wang, Z.Z., Yang, J.D, Ying, L.M. & Zhang, J.M., 1986. Aspects of Cambrian-Ordovician Boundary in Dayangcha, China. 1-410, pl. 1-98, China Prospect Publishing House, Beijing.
185Cooper, R.A., 1999. Ecostratigraphy, zonation and global correlation of earliest Ordovician planktic graptolites. Lethaia, 32, 1-16.
186Cooper, R.A. & Stewart, I.R., 1979. The Tremadoc graptolite sequence of Lancefield, Victoria. Palaeontology, 22, 767-797.
187Cooper, R.A., Nowlan, G.S. & Williams, S.H., 2001. Global stratotype section and point for the base of the Ordovician System. Episodes, 24, 19-28.
188Cooper, R.A., Maletz, J., Wang H.F. & Erdtmann, B.-D., 1998. Taxonomy and evolution of earliest Ordovician graptolites. Norsk Geologisk Tidsskrift, 78, 3-32.
189Cooper, R.A., Rushton, A.W.A., Molyneux, S.G., Hughes, R.A., Moore, R.M. & Webby, B.C., 1995. The stratigraphy, correlation, provenance and palaeogeography of the Skiddaw Group (Ordovician) in the English Lake District. Geological Magazine, 132, 185-211.
190Egenhoff, S., Maletz, J. & Erdtmann, B.-D., 2004. Lower Ordovician graptolite biozonation and lithofacies of southern Bolivia: relevance for palaeogeographic interpretations. Geological Magazine, 141, 287-299.
191Eichwald, E.J., 1840. Ueber das silurische Schichtensystem in Esthland. Zeitschrift für Natur. und hilk. dimediz. Akad. zu St. Petersburg, Hefte 1 and 2. 210 pp.
192Eichwald, E.J., 1855. Beitrag zur geographischen Verbreitung der fossilen Thiere Russlands. Alte Periode. Bulletin de la Société Imperiale des Naturalistes de Moscou, 28, 433-466.
193Erdtmann, B.-D., 1982. A reorganization and proposed phylogenetic classification of planktic Tremadoc (early Ordovician) dendroid graptolites. Norsk Geologisk Tidsskrift, 62, 121-145.
194Erdtmann, B.-D., 1986. Comments on some earliest Ordovician (Salmian) Graptolites from Solwaster, Massif de Stavelot, Belgian Ardennes. Aardkundige Mededelingen, 3, 75-88.
195Erdtmann, B.D., 1988. The earliest Ordovician nematophorid graptolites: taxonomy and correlation. Geological Magazine, 125(4), 327-348.
196Feng, H.Z. & Erdtmann, B.D., 1999. Early Tremadoc graptolite sequence in the Wuning area, South China and its international correlation. Acta University Carolinae (Geologica), 43 (1-2), 21-24.
197Geukens, F.R.M., 1950. Quelques nouveaux gîtes à Dictyonema flabelliforme dans le massif de Stavelot. Bulletin de la Société Belge de Géologie, de Paléontologie et d’Hydrologie, 59, 163-169.
198Geukens, F.R.M., 1954. Quelques remarques au sujet de la répartition des Dictyonema flabelliforme dans le massif cambrien de Stavelot (Belgique). Congrès géologique International, Alger. Sect. xiii, fasc. xv, 45-52.
199Graulich, J.M., 1961. Le sondage de Wépion. Memoires pour servir à l’explication des Cartes géologiques et minières de la Belgique, 2, 1-102.
200Graulich, J.M., 1963. Découverte du niveau à Dictyonema flabelliforme parabola dans le Salmien du massif de Stavelot (Belgique). Comptes rendus de l’Académie des Sciences, Paris, 256, 3327-3328.
201Gutiérrez-Marco, J.C. & Esteban, S.B., 2003. Reappraisal of the Tremadocian graptolite sequence of the Famatina System, NW Argentina. In Ortega G. & Aceñolaza G.F. (eds), Proceedings of the 7th International Graptolite Conference. Correlación Geológica, Tucumán, 18, 39-44.
202Gutiérrez-Marco, J.C. & Esteban, S.B., 2005. Graptolitos del Tremadociense (Ordovícico Inferior) de la Formación Volcancito, Sistema de Famatina (La Rioja, Argentina). Revista Española de Paleontología, 20, 65-118.
203Hall, J., 1843. Geology of New York. Report on Fourth geological District, 115-116.
204Hall, J., 1851. New Genera of fossil Corals etc. American Journal of Science, Series 2, 11, 401.
205Hahn, F. F., 1912. On the Dictyonema-fauna of Navy Island, New Brunswick. Annals of the New York Academy of Sciences, 22, 135-160.
206Herbosch, A. & Verniers, J., 2013. Stratigraphy of the Lower Palaeozoic of the Brabant Massif, Belgium. Part I: The Cambro-Ordovician from the Halle and Ottignies groups. Geologica Belgica, 16 (1-2), 49-65.
207Hopkinson, J. & Lapworth, C., 1875. Descriptions of the Graptolites of the Arenig and Llandeilo rocks of St. Davids. Quarterly Journal of the Geological Society of London, 31, 631-672.
208Jin, Y.Q. & Wang, X.F., 1977. The multi-ramous graptolites from Baishuixi Formation and Qiaotingzi Formation (Lower Ordovician), central Hunan. Selected Papers on Stratigraphy and Palaeontology, 3, 74-85.
209Kjerulf, T., 1865. Veiviser ved geologiske excursioner i Christiana omegn. Universitets program for andet halvaar, 1-43.
210Lankester, E.R., 1877. Notes on the embryology and classification of the animal kingdom: comprising a revision of speculations relative to the origin and significance of the germ-layers. Quarterly Journal of Microscopical Science, New series, 17, 399-454.
211Landing, E., Bowring, S. A., Davidek, K. L., Rushton, A. W. A., Fortey, R. A. & Wimbledon, W. A. P., 2000. Cambrian–Ordovician boundary age and duration of the lowest Ordovician Tremadoc Series based on U–Pb zircon dates from Avalonian Wales. Geological Magazine, 137 (5), 485-494.
212Lecompte, M., 1948. Existence du Trémadoc dans le Massif du Brabant. Bulletin de l’Académie Royale de Belgique, Classe des Sciences, 34, 677-687.
213Lecompte, M., 1949. Découverte de nouveaux gîtes à Dictyonema dans le Trémadocien du Massif du Brabant.Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 25, 1-8.
214Legrand, P., 1974. Development of rhabdosomes with four primary branches in the group Dictyonema flabelliforme (Eichwald). In Rickards, R.B., Jackson, D.E. & Hughes, C.P. (eds), Graptolite studies in honour of O.M.B. Bulman. Special Papers in Palaeontology, 13, 19-34.
215Legrand, P., 1985. Dictyonema-Rhabdinopora. Norsk Geologisk Tidsskrift, 65, 224-226.
216Li, J, J. & Chen X., 1962. Cambrian and Ordovician graptolites from Sandu, S. Guizhou (Kueichou). Acta Palaeontologica Sinica, 10 (1), 12-30.
217Lin, Y.K., 1981. New materials of graptodendroids with special reference to the classification of Graptodendroidea. Bulletin of Nanjing Institute of Geology and Palaeontology, Academia Sinica, 3, 242-262 [in Chinese].
218Lin, Y.K., 1988. On the proximal tufts of threads in Dictyonema. Acta Palaeontologica Sinica, 27 (2), 218-237.
219Lindholm, K., 1991. Ordovician graptolites from the early Hunneberg of southern Scandinavia. Palaeontology, 34, 283-327.
220Liu, C.L., 1984. Application of multi-analysis in the taxonomy of graptolites near the Cambrian–Ordovician boundary from Taoyuan area, Hunan Province. Master Thesis, Nanjing University, 103 pp. [in Chinese].
221Loydell, D.K., 2012. Graptolite biozone correlation charts. Geological Magazine, 149, 124-132.
222Malaise, C., 1874a. Sur l’âge de quelques couches du terrain ardennais des environs de Spa. Bulletin de l’Académie royale de Belgique, 37, 800-801.
223Malaise, C., 1874b. Découverte de Dictyonema sociale. Bulletin de l’Académie royale de Belgique, 38, 464.
224Malaise, C., 1881. Sur le Dictyonema sociale, Salt. Documents paléontologiques relatives au terrain Cambrien de l’Ardenne, Bruxelles.
225Malaise, C., 1888. Sur la présence du Dictyonema sociale à La Gleize. Annales de la Société Géologique de Belgique, 15, B76.
226Maletz, J., 1992. The proximal development in Anisograptids (Graptoloidea, Anisograptidae). Paläontologische Zeitschrift, 66, 297-309.
227Maletz, J., 2014. The classification of the Pterobranchia (Cephalodiscida and Graptolithina). Bulletin of Geosciences, 89 (3), 477-540.
228Maletz, J. & Egenhoff, S., 2001. Late Tremadoc to early Arenig graptolite faunas of southern Bolivia and their implications for a worldwide biozonation. Lethaia, 34, 47-62.
229Maletz, J., Carlucci, J. & Mitchell, C.E., 2009. Graptolite cladistics, taxonomy and phylogeny. Bulletin of Geosciences, 84, 7-19.
230Maletz, J., Egenhoff, S. & Alonso, R., 2010. The Upper Tremadocian (Ordovician) graptolite Bryograptus, taxonomy, biostratigraphy and biogeography. Palaeontology, 53, 59-75.
231Matthews, S. C., 1973, Notes on open nomenclature and on synonymy lists. Palaeontology, 16, 713-719.
232Monteros, J.A. & Moya, M.C., 2011. Anisograptus mojotorensis sp. nov. en el Tremadociano temprano del norte argentino. Revista Española de Paleontología, 26, 53-67.
233Mu, E.Z., 1955. The new materials of the dendroid graptolites. Palaeontologia Sinica, 138, 1-62.
234Mu, E.Z., Li, J.J., Ge, M.Y., Lin, Y.K. & Ni, Y.N. 2002. The fossil Graptolites of China. Science press, Beijing, i-xiv, 1-1205.
235Ortega, G. & Albanesi, G.L., 2005. Tremadocian graptolite-conodont biostratigraphy of the South American Gondwanan margin (Eastern Cordillera, NW Argentina). Geologica Acta, 3, 355-371.
236Ruedemann, R., 1904. Graptolites of New York, Part 1. Graptolites of the lower beds. New York State Museum, Memoir 7, 1-803.
237Salter, J. W., 1858. On Graptopora, a new genus of Polyzoa, allied to the graptolites. Proceedings of the American Association for the Advancement of Science for 1857, 11, 63-66.
238Salter, J. W., 1866. On the fossils of North Wales, In Ramsay, A.C., The geology of North Wales. Memoirs of the Geological Survey of Great Britain, 3, 239-381.
239Størmer, L., 1940. Dictyonema shalesoutside the Oslo Region. Norsk Geologisk Tidsskrift, 20, 161-169.
240Tortello, M.F. & Esteban, S.B., 1999. La transición Cámbrico-Ordovícico en la Formación Volcancito (sierra de Famatina, La Rioja, Argentina).Ameghiniana, 36, 371-387.
241Verniers, J., Herbosch, A., Vanguestaine, M., Geukens, F., Delcambre, B. Pingot, J.L., Belanger, I., Hennebert, M., Debacker, T., Sintubin, M. & De Vos, W., 2001. Cambrian-Ordovician-Silurian lithostratigraphic units (Belgium). Geologica Belgica, 4, 5-38.
242Veski, R. & Palu, V., 2003. Investigation of Dictyonema oil shale and its natural and artificial transformation products by a vankrevelenogram. Oil Shale, 20 (3), 265-281.
243Wang, X.F. & Erdtmann, B.-D., 1987. Zonation and correlation of the earliest Ordovician graptolites from Hunjiang, Jilin Province, China. Bulletin of Geological Society of Denmark, 35, 245-257.
244Wang, X.F. & Wang, C.S., 2001. Tremadocian (Ordovician) graptolite diversification events in China. Alcheringa, 25, 155-168.
245Wang, H.F., Cooper, R.A. & Erdtmann, B.D., 1998. Restudy of the Earliest Ordovician graptolites from the Dayangcha Section in Jilin. Acta Palaeontologica Sinica, 37 (2), 212-231.
246Wang, W.H., Muir, L.A., Botting, J.P., Feng, H.Z., Servais, T. & Li, L.X., 2014. A Tremadocian (Early Ordovician) palaeoscolecidan worm from graptolitic shales in Hunan Province, South China. Palaeontology, 57 (3), 657-671.
247Wang, W.H. & Muir, L.A., 2015. Taxonomic and biostratigraphic reappraisal of some early Tremadocian (Ordovician) graptolites from Changde, South China. Palaeoworld, 24, 86-99.
248Webby, B.D., Cooper, R.A., Bergström, S.M. & Paris, F., 2004. Stratigraphic framework and time slices. In: Webby, B.D., Paris, F., Droser, M. & Percival, I. (eds), The Great Ordovician Biodiversification Event. Columbia University Press, New York, 41-47.
249Westergård, A. H., 1909. Studier öfver Dictyograptus-skiffern och dess gränslager. Lunds Universitets Årsskrift, N.F., 5, 1-79.
250Williams, S.H. & Stevens, R.K., 1991. Late Tremadoc graptolites from western Newfoundland. Palaeontology, 34, 1-47.
251Yang, D.Q., Ni, Y.W., Li, J.J., Chen, X., Lin, Y.K., Yu, J.H., Xia, G.S., Jiao, S.D., Fang, Y.T., Ge, M.Y. & Mu, E.Z., 1983. Graptolithina. In: Nanjing Institute of Geology and Mineral Resources (ed.), Palaeontological Atlas of East China (1) Early Palaeozoic. Geological Publishing House, Beijing, 353-357. [in Chinese]
252Yu, J.H., Liu, H.B. & Fang, Y.T., 1985. Dendroids from the Xinchangian Stage (Early Ordovician) of the Xiushui Drainage Basin, Jiangxi Province. Acta Palaeontologica Sinica, 24 (1), 18-28.
253Zeballo, F.J., Albanesi, G.L. & Ortega, G., 2005. Conodontes y graptolitos de las formaciones Alfarcito y Rupasca (Tremadociano) en el área de Alfarcito, Tilcara, Cordillera Oriental de Jujuy, Argentina Parte 2: Paleontología sistemática. Ameghiniana, 42, 1, 47-66.
254Zeballo, F.J., Albanesi, G.L. & Ortega, G., 2008. New late Tremadocian (Early Ordovician) conodont and graptolite records from the southern South American Gondwana margin (Eastern Cordillera, Argentina). Geologica Acta, 6, 131-145.
255Zhang, Y.D., Wang Z.H., Feng, H.Z., Luo, T.T. & Erdtmann, B.-D., 2005. Tremadocian (Ordovician) graptolite biostratigraphy of China - a review. Journal of Stratigraphy, 29, 215-235.
256Zhao, X.L., 1983. Early Xinchangian (X1) dendroids from Benxi, Liaoning. Acta Palaeontologica Sinica, 22 (4), 369-385.
257Manuscript received 11.02.2015, accepted in revised form 17.04.2015, available on line 09.06.2015.
To cite this article
