


- Home
- volume 18 (2015)
- number 2-4
- Revision of the genus Frechastraea Scrutton, 1968 (Rugosa) in the Upper Frasnian of Belgium


View(s): 1678 (36 ULiège)
Download(s): 370 (10 ULiège)

Revision of the genus Frechastraea Scrutton, 1968 (Rugosa) in the Upper Frasnian of Belgium

Abstract
Frechastraea pentagona (Goldfuss, 1826), type species of Frechastraea Scrutton, 1968, F. minima (Rozkowska, 1953), F. micrommata (Roemer, 1852), F. kaisini (Tsien, 1978), F. cf. tungkanlingensis (Yoh, 1937), F. crassiseptata (Tsien, 1978) and F. limitata (Milne-Edwards & Haime, 1851) occur mainly in the Upper Palmatolepis rhenana conodont Zone. Their stratigraphic distribution is discussed in the Upper Frasnian of Belgium and compared with other areas of Western and Eastern Europe. The Belgian material previously identified as Frechastraea micrastraea (Penecke, 1904) is now assigned to F. crassiseptata. The Champ Broquet Formation is proposed herein to include the Neuville and Les Valisettes Members which were formerly considered as two different formations. The red marble lenses of the Petit-Mont Member are developed at different levels of the Champ Broquet Formation, in the Dinant Synclinorium.
Table of content
1. Introduction
1Three species of the genus Frechastraea Scrutton, 1968, namely F. phillipsastraeiformis (Moenke, 1954), F. coeni Coen-Aubert, 2012 and F. glabra Coen-Aubert, 2012, have been described by Coen-Aubert (2012) at the base of the Upper Frasnian from Belgium. In continuation of this paper, six species and one form of Frechastraea are revised herein; these are F. pentagona (Goldfuss, 1826), type species of the genus, F. minima (Rozkowska, 1953), F. micrommata (Roemer, 1852), F. kaisini (Tsien, 1978), F. cf. tungkanlingensis (Yoh, 1937), F. crassiseptata (Tsien, 1978) and F. limitata (Milne-Edwards & Haime, 1851). Most of these occurrences characterize the Upper Palmatolepis rhenana conodont Zone. As mentioned by Coen-Aubert (2012), the base of the Upper Frasnian has been fixed by the Subcommission on Devonian Stratigraphy, at the entry of the conodont Palmatolepis semichatovae. According to Sandberg et al. (1992), the entry of P. semichatovae is observed within the Lower Palmatolepis rhenana Zone, together with the first occurrence of Ancyrognathus triangularis, in bed 150 lying 2.6 m above the base of the Neuville Formation as it is exposed along the southern access road to the Lion quarry at Frasnes, on the south side of the Dinant Synclinorium. Additionally, the massive rugose coral Frechastraea coeni was noted as an excellent marker for the base of the Upper Frasnian in Belgium by Coen-Aubert (2012).
2During the revision of the geological map of Wallonia, several Palaeozoic structural units have been redefined in Southern Belgium by Belanger et al. (2012). According to these authors and to Delmer (2004), there is no Namur Synclinorium as its two flanks do not belong to the same structural unit. Then Delmer (2004) suggested using the expression Namur Basin just as it is the case for the Campine Basin.
3The main part of the material investigated herein was collected by the author and Michel Coen in situ, during geological surveys made bed by bed in different localities from the southern part of Belgium (Fig. 1). This sampling is supplemented by a few older thin sections referred in this paper to the "Old collection from the Institut royal des Sciences naturelles de Belgique".
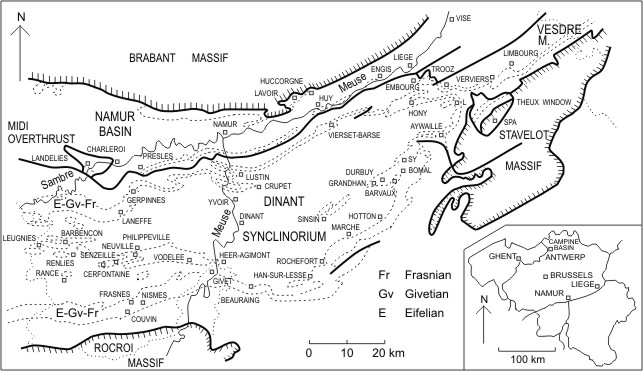
4Figure 1. Geological setting and locality map in the southern part of Belgium. The small outliers of Streupas and Kinkempois are situated close to Liège, at the western end of the Vesdre Massif and are located with precision by Coen-Aubert (1974b, fig. 2). L corresponds to Louveigné, on the northeastern side of the Dinant Synclinorium.
2. Geological setting and lithostratigraphy
5The Champ Broquet Formation (Fig. 2) is proposed herein, within the Dinant Synclinorium, to include the Neuville and Les Valisettes Members which were formerly considered as two different formations by Boulvain et al. (1993, 1999). The Neuville Formation was introduced by Tsien (1974, 1975) for a succession of four lithological units in the Philippeville Massif. It was then reduced to the first two lithological units of Tsien (1974, 1975) by Boulvain et al. (1993, 1999) who assigned its last two lithological units to the Les Valisettes Formation. Moreover, Boulvain et al. (1999) referred to the Petit-Mont Member the reefal lens of red limestone exposed in the Petit-Mont quarry at Vodelée which starts its development a few metres above the base of the Neuville Member and continues to grow up rather high, laterally to the Les Valisettes Member. In the Philippeville Massif, a similar situation is observed in the Haumont quarry also at Vodelée, the Beauchâteau quarry at Senzeille and the Tapoumont quarry at Neuville whereas the smaller red bioherm of Les Bulants quarry from the same locality is nearly limited to the Neuville Member. The stratotype of the Champ Broquet Formation is the same as those of the Neuville and Les Valisettes Members described by Boulvain et al. (1999). Champ Broquet is a place situated to the south of the two railway sections which constitute this stratotype; it can be seen on the map published by Boulvain et al. (1999, p. 72).
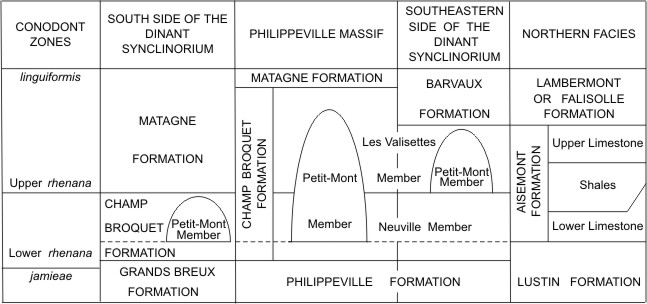
6Figure 2. Lithostratigraphy in the Upper Frasnian from the south side of the Dinant Synclinorium, the Philippeville Massif, the southeastern side of the Dinant Synclinorium and the northern facies. The latter characterize the north side of the Dinant Synclinorium, the south side of the Namur Basin and the Vesdre Massif. The broken line corresponds to the base of the Upper Frasnian.
7On the south side of the Dinant Synclinorium, the lenses of the Petit-Mont Member are often restricted to the Neuville Member whereas the Les Valisettes Member is poorly developed and overlain by a great thickness of black shales belonging to the Matagne Formation. In the Philippeville Massif, the Matagne Formation is much reduced and according to Boulvain et al. (1999), nodular limestones rich in massive rugose corals are locally present in the underlying Les Valisettes Member, in the vicinity of the big lenses of the Petit-Mont Member.
8As it is defined in this paper, the Champ Broquet Formation resembles more than before the Moulin Liénaux and Grand Breux Formations, mostly represented on the south side of the Dinant Synclinorium, where bioherms also occur. Indeed, the nodular limestones of the Neuville Member serve as basement for the lenses of the Petit-Mont Member just as it is the case for the Chalon Member at the base of the bioherms of the Arche Member in the Moulin Liénaux Formation and for the Bieumont Member at the base of the bioherms of the Lion Member in the Grand Breux Formation. The predominantly shaly sediments of the Les Valisettes Member are present laterally and above the lenses of the Petit-Mont Member. The same situation exists for the Ermitage Member in the Moulin Liénaux Formation and for the Boussu-en-Fagne Member in the Grand Breux Formation.
9However, there are differences with the reefal lenses of the Petit-Mont Member exposed along the southeastern side of the Dinant Synclinorium. Indeed, the red marble bioherms of Sy, Bomal, Durbuy, Barvaux and Grandhan investigated by Coen (1974) and Coen et al. (1977) start on a basement formed by two beds of limestones, which has been correlated by these authors with the base of the Matagne Formation, on the south side of the Dinant Synclinorium. That is to say that the lenses of the Petit-Mont Member from the southeast side of the Dinant Synclinorium are younger than the Neuville Member and entirely developed in the Les Valisettes Member which is capped in this area by the purple shales of the Barvaux Formation. Locally, massive rugose corals occur in the Barvaux Formation, for instance at Sinsin and Marche-en-Famenne.
10To the northwest of the Philippeville Massif, there are other small lenses of the Petit-Mont Member at Barbençon, Solre-Saint-Géry and Leugnies. These quarries studied by Dumoulin (2001) are once more lateral to the Les Valisettes Member which is succeeded in these localities by the Famenne Formation. This also the case in the Rance Anticline where a bioherm of the Petit-Mont Member is followed after an unconformity by Famennian shales of the Famenne Formation with conodonts of the Palmatolepis triangularis Zone. The data about Rance have been provided by Biron et al. (1983) and Muchez et al. (1996). So the position of the Petit-Mont Member is highly variable in the Champ Broquet Formation.
11Along the eastern side of the Dinant Synclinorium, the Champ Broquet Formation passes laterally into the Aisemont Formation which characterizes the north side of the same structural unit, the south side of the Namur Basin and the Vesdre Massif. The Aisemont Formation, which is also present at Huccorgne on the north side of the Namur Basin and in the Campine Basin, consists of two levels of limestone separated by shales. According to Coen (1974, pl. I) and Coen et al. (1977, fig. 1), the basement of the Petit-Mont Member along the southeastern border of the Dinant Synclinorium can be correlated with a bed of limestone occurring within the middle shaly part of the Aisemont Formation, at Comblain-la-Tour and Aywaille on its northeastern side. Above the Aisemont Formation, the base of the Lambermont Formation corresponds to the base of the Barvaux Formation for Boulvain et al. (1999). The presence of nodular limestones with massive rugose corals close to the base of the Lambermont Formation is one of the characteristics of this lithostratigraphic unit. As for the Falisolle Formation, it has been introduced by Delcambre & Pingot (2000, 2014); its stratotype is situated at Aisemont on the south side of the Namur Basin. The Falisolle Formation is mostly represented by shales between the top of the Aisemont Formation and the base of Middle Famennian sandstones; so it crosses the Frasnian-Famennian boundary. The Frasnian part of the Falisolle Formation is developed laterally to the main part of the Lambermont Formation. It has been used recently by Denayer & Poty (2010) and Denayer et al. (2012).
12For these northern areas, detailed geographic and stratigraphic information with maps showing the localities and outcrops is provided by:
13- Coen-Aubert (1974a, b), Coen (1974) and Coen-Aubert & Coen (1975) for the Vesdre Massif, the Streupas and Kinkempois outliers, the Theux Window and the north side of the Dinant Synclinorium;
14- Coen-Aubert & Lacroix (1979, 1985) and Coen (1976) for the Namur Basin and the La Tombe outlier (Landelies);
15- Poty (1982) for the Visé area;
16- Coen-Aubert (2014) for the Campine Basin (Booischot and Heibaart boreholes).
3. Systematic Palaeontology
17All the figured specimens are stored in the Collection of Palaeontology from the Institut royal des Sciences naturelles de Belgique at Brussels (IRScNB).
18Family Phillipsastreidae Roemer, 1883
19Genus Frechastraea Scrutton, 1968
20Type species. By original designation, Cyathophyllum pentagonum Goldfuss, 1826.
21Diagnosis. Massive rugose corals, pseudocerioid to astreoid at times with small corallites. Septa of two orders, carinate or non-carinate, dilated in the dissepimentarium and occasionally thicker at its inner border, thin in the tabularium. Major septa extending nearly to the axis of the corallites or restricted to the dissepimentarium. Dissepimentarium composed of several rows of small globose dissepiments arranged in horizontal layers with possible occurrence of a few horseshoe dissepiments at the border of the tabularium. Tabulae complete or incomplete.
22Frechastraea pentagona (Goldfuss, 1826)
23(Plate 1A-H)
24* v 1826 Cyathophyllum pentagonum nobis; Goldfuss : 60, pl. 19, fig. 3.
251851 Acervularia pentagona; Milne-Edwards & Haime : 418.
261851 Acervularia goldfussi; Milne- Edwards & Haime : 417.
271853 Acervularia pentagona; Milne-Edwards & Haime : 238, pl. 53, figs 5-5b.
281853 Acervularia goldfussi; Milne-Edwards & Haime : 236, pl. 53, figs 3-3a.
291881 Acervularia pentagona Goldf. sp.; Schlüter : 89, pl. 9, figs 4-5.
301883 Acervularia pentagona; Roemer : 352, fig. 70.
31p. 1885 Phillipsastrea pentagona Goldf. sp.; Frech : 54, pl. 3, figs 6?, 9-10 (non figs 7-8), pl. 8, fig. 3.
32non 1952 Phillipsastraea pentagona (Goldf.); Soshkina : 102, pl. 43, fig. 145.
33non 1952 Phillipsastraea pentagona Goldfuss; Ivania : p. 135, pl. 1, fig. 2.
34v ? 1953 Phillipsastraea pentagona Goldfuss; Rozkowska : 64, pl. 8, fig. 7.
35non 1955 Phillipsastraea pentagona (Goldfuss); Ivania : 223, pl. 43, fig. 2.
36non 1960 Phillipsastraea pentagona Goldfuss, 1826; Zheltonogova & Ivania : 381, pl. D-34, fig. 2.
37non 1965 Phillipsastraea pentagona Goldfuss; Ivania : 182, pl. 99, figs 423-424.
38v 1967 Hexagonaria pentagona (Goldfuss, 1826); Pickett : 60, fig. 17, pl. 3, fig. 10, pl. 4, fig. 13, pl. 7, fig. 30.
39p. 1967 Phillipsastraea pentagona (Goldfuss); Sorauf : 29, figs 13, 1a-b (non figs 6, 1a-g, figs 13, 1c-f).
40p. 1967 Phillipsastraea goldfussi (de Verneuil & Haime); Sorauf : 27, figs 10, 1a-f (non figs 11, 1a-b).
41v non 1968 Phillipsastraea pentagona (Goldfuss); Tsien : pl. 2, figs 3-4, pl. 3, fig. 11, pl. 5, fig. 1.
42p. 1968 Frechastraea pentagona pentagona (Goldfuss); Scrutton : 233, pl. 7, figs 2-5 (non pl. 6, fig. 5, pl. 7, fig. 1).
43non 1971 Phillipsastraea pentagona (Goldfuss); Tsien : figs 32, 2a-b.
44v p. 1974a Phillipsastrea pentagona pentagona (Goldfuss, G.A., 1826); Coen-Aubert : 10, pl. 1, figs 4-6 (non pl. 1, fig. 1).
45v 1974b Phillipsastraea pentagona (Goldfuss); Coen-Aubert : pl. 3, fig. 3.
46non 1975 Frechastraea pentagona (Goldfuss); Besprozvannykh et al. : pl. 16, fig. 2.
47v non 1975 Phillipsastrea pentagona; Tsien : fig. 26o.
48v non 1977 Phillipsastrea pentagona; Tsien : figs 7c-d.
49v 1977 Frechastraea pentagona pentagona; Coen et al. : 326.
50v non 1978 Phillipsastrea pentagona (Goldfuss, 1826); Tsien : 205, fig. 7, pl. 1, fig. 7.
51non 1978 Frechastraea pentagona pentagona (Goldfuss, 1826); Birenheide : 102, pl. 16, fig. 1.
52v 1981 Frechastraea pentagona (Goldfuss); Hill : F284, figs 182, 2a-c.
53? 1986 Frechastraea pentagona pentagona (Goldfuss); Sorauf & Pedder : pl. 5, figs 2, 5.
54v p. 1987 Frechastraea pentagona pentagona (Goldfuss, G.A., 1826); Coen-Aubert : 50.
55non 1992 Frechastraea pentagona (Goldfuss 1826); Hladil et al. : pl. 12, fig. 5.
561998 Frechastraea (Frechastraea) pentagona pentagona (Goldfuss 1826); Birenheide: 177, pl. 3, fig. 2.
57non 2003 Frechastraea pentagona (Goldfuss, 1826); Fedorowski : p. 121, pl. 62, figs 1-5.
58v 2014 Frechastraea pentagona (Goldfuss, 1826); Coen-Aubert : fig. 3C.
59Holotype. Specimen GMBo 206 stored in the Goldfuss collection from the Paläontologisches Institute of the University of Bonn in Germany. Limbourg in Belgium (Fig. 1), Transition limestone from the area of Namur (Frasnian?). This specimen has been chosen as lectotype by Pickett (1967). However as mentioned by Birenheide (1998), there was only one colony assigned to Frechastraea pentagona by Goldfuss (1826); so it is a holotype by monotypy. Pl. 19, fig. 3 in Goldfuss (1826), fig. 17, pl. 3, fig. 10, pl. 4, fig. 13 and pl. 7, fig. 30 in Pickett (1967), pl. 7, fig. 5 in Scrutton (1968) and fig. 182, 2a-b in Hill (1981).
60Material. 50 specimens with 97 thin sections. Personal sampling with that of M. Coen: Heibaart borehole at 1593 m; Chênée MC-1974-83-H58; Chênée MC-1974-84-L1; Verviers MC-1974-7-2, 16 and 18; Verviers MC-1974-56-402, 405 and 408; Verviers MC-1988-4-A684, A685, A686 and A688; Limbourg MC-1974-9-A87, A91 and A92; Limbourg MC-174-36-L43; Limbourg MC-1974-29-F76; Spa MC-1974-92-11 and 12; Esneux MC-1974-89-F84; Ferrières MC-1974-110- l. 12748 and l.12749; Aye MC-1974-112-5; Aye MC-114-D809, D810, D812, D814, D817, D818B and D820; Senzeille MC-1974-14-P33 and P36; Senzeille MC-1974-15-T887; Froidchapelle MC-71-C937, C938, C941, C943, C944 and C946; Froidchapelle MC-75-C992, C998, C999, D101, D105, D107 and D116; Surice MC-1986-5-B43 and B44; Rance MC-132-R3.
61Diagnosis. A pseudocerioid species of Frechastraea with 20 to 24 septa at tabularial diameters of 1.1 mm to 1.8 mm. Septa non-carinate and slightly dilated in the dissepimentarium with a short bulbous thickening at its inner border. Major septa reaching the axis of the corallites.
62Description. The material consists of platy, tabular and nodular colonies which are complete or fragmentary. Their height varies between 1 cm and 7.5 cm whereas the largest piece reaches an area of 15 cm x 8.5 cm. The corallites polygonal in shape are separated by a zigzagged or sometimes straight pseudotheca. In very few specimens, the outer zigzagged wall is locally weaker or even absent giving a more or less astreoid look. Some lateral and pericalicinal offsets have been observed.
63The septa are normally non-carinate; however, a few knobbly, spinose and yardarm carinae are present in some colonies. The septa are usually weakly dilated in the dissepimentarium and become thin in the tabularium or slightly beyond their entry into it. They are characterized by some thickening in a very narrow area at the border or the tabularium where an inner wall is occasionally seen in transverse or in longitudinal section.
64The major septa reach the axis of the corallites where they are often fusing partially or totally and forming pseudofossulae. In the centre of the tabularium, there are sometimes a thin pseudocolumella, an open space or a small and more or less complete pseudoaulos; in rare cases, the major septa are hardly extending into the tabularium. The minor septa traverse the entire dissepimentarium; occasionally, they are barely projecting into the tabularium.
65The dissepimentarium consists of 3 to 8 or even 2 to 10 rows of globose dissepiments which are arranged in horizontal layers. In a few colonies occur locally at the border of the tabularium small horseshoe and specialized dissepiments or one row of inclined dissepiments. The tabulae, which are frequently disrupted by axial ends of major septa, are incomplete and intersecting laterally with sometimes a flat-topped axial part; they are rarely horizontal, concave or convex.
66There are 18 to 26 or even 28 septa per corallite. The width of the tabularium ranges from 1 mm to 2.1 mm and even to 2.5 mm. The diameter of the corallites varies commonly between 3 mm and 5.5 mm and more generally between 2.5 mm and 6.3 mm, but it may reach 6.9 mm.
67Discussion. After this revision and the list of synonymy provided herein, it is clear that Frechastraea pentagona is not a very well understood species though its holotype has been figured several times. The type specimen has been collected at Limbourg in Belgium (Fig. 1) as read by Pickett (1967) on the label stored in Bonn. The name of Limbourg was already mentioned by Milne-Edwards & Haime (1851, 1853) whereas Roemer (1883) added that the colony described by Goldfuss (1826) comes from Verviers. Limbourg is built on the Famennian of the Vesdre Massif (Laloux et al., 1996), about 2 km to the northeast of the outcrop Limbourg MC-1974-9 at Les Surdents where I have collected three specimens of F. pentagona in the Upper Frasnian Lambermont Formation. And the small village Les Surdents is situated a few kilometres to the northeast of Verviers. Moreover, I have sampled several colonies of F. pentagona at Lambermont and Pepinster (Plate 1C) close to Verviers, also in the Lambermont Formation. All these localities have been described in detail by Coen-Aubert (1974b) and Birenheide et al. (1991).
68The holotype and the Belgian material of F. pentagona are characterized by septa slightly dilated in the dissepimentarium with a short bulbous thickening restricted to the border of the tabularium, normally without any trace of inner wall in both transverse and longitudinal sections. Some colonies identified as F. goldfussi (De Verneuil & Haime, 1850) by Milne-Edwards & Haime (1853, pl. 53, fig. 3) and Sorauf (1967, figs 10, 1a-f) are certainly representatives of F. pentagona. As mentioned by Coen-Aubert (2012), the holotype of F. goldfussi is lost and it is impossible to know what is the species in actuality. Specimens with an inner wall and very small corallites, which were assigned to F. pentagona by Scrutton (1968, pl. 6, fig. 5, pl. 7, fig. 1) and Hladil et al. (1992, pl. 12, fig. 5) seem to be very close to F. minima. The material from the Upper Frasnian of the Kuznetsk Basin in Russia referred to F. pentagona by Ivania (1952, 1955, 1965) and Zheltonogova & Ivania (1960) probably belongs to F. crassiseptata.
69Finally, F. phillipsastraeiformis from the Middle to the Upper Frasnian of the Holy Cross Mountains in Poland, which has been compared with F. pentagona by Coen-Aubert (2012), is distinguished by weakly carinate and spindle-shaped dilated septa and by slightly greater septal number and diameters of the tabularia and corallites.
70Distribution. Frechastraea pentagona occurs rather high in the Upper Frasnian from various areas of Belgium. The material collected by the author and M. Coen comes from:
71- the base of the Falisolle Formation in the Campine Basin;
72- the lower part of the Lambermont Formation in the Kinkempois outlier, the Vesdre Massif, the Theux Window and the northeast side of the Dinant Synclinorium;
73- the Les Valisettes Member in the Philippeville Massif at Neuville, Senzeille and Cerfontaine whereas F. pentagona has also been observed at the top of the Petit-Mont Member, in the Hautmont quarry at Vodelée and in the Renlies Anticline;
74- the Barvaux Formation at Marche-en-Famenne and Sinsin, on the southeast side of the Dinant Synclinorium.
75Outside Belgium, F. pentagona is known in the Upper Frasnian from South Devon in Great Britain as well as from the Harz Mountains, the Aachen Syncline and the Land North Rhine-Westphalia in Germany. It may be present in the Upper Frasnian from the Holy Cross Mountains in Poland.
76Frechastraea minima (Rozkowska, 1953)
77(Plate 2A-G)
78v 1953 Phillipsastraea pentagona Goldfuss var. minima nov.; Rozkowska : 66, pl. 8, fig. 9.
79v p. 1967 Phillipsastraea pentagona minima Rozkowska; Sorauf : 31, figs 14, 1a-b (non fig. 14, 1c).
801968 Frechastraea pentagona (Goldfuss) minima (Rozkowska); Scrutton : 236, pl. 8, figs 1-3.
81v p. 1968 Phillipsastraea pentagona (Goldfuss); Tsien : pl. 2, fig. 3, pl. 3, fig. 11, (non pl. 2, fig. 4, pl. 5, fig. 1).
82v 1974a Phillipsastrea pentagona minima Rozkowska, M., 1953; Coen-Aubert : 12, pl. 2, figs 1-3.
83v 1975 Phillipsastrea pentagona; Tsien : fig. 26o.
84v 1977 Phillipsastrea pentagona; Tsien : figs 7c-d.
85v 1978 Phillipsastrea pentagona (Goldfuss, 1826), var. minima Rozkowska, 1953; Tsien : 205, pl. 2, fig. 1.
86v 1978 Phillipsastrea frocapella nov. sp.; Tsien : 207, pl. 2, fig. 3.
87v 1978 Phillipsastrea pentagona (Goldfuss, 1826); Tsien : 205, fig. 7, pl. 1, fig. 7.
88v 1978 Phillipsastrea sp. 1; Tsien : 208, pl. 2, fig. 7.
89non 1978 Frechastraea pentagona minima (Rozkowska, 1953); Birenheide : pl. 16, fig. 2.
90non 1978 Frechastraea pentagona minima (Rozkowska); Kong & Huang : 87, pl. 30, fig. 3.
91v 1983 Frechastraea pentagona pentagona (Rozkowska, M., 1953); Biron et al. ; pl. 1, fig. 3.
92v 1987 Frechastraea pentagona minima (Rozkowska, M. 1953); Coen-Aubert : 50, pl. 2, figs 7-8.
932001 Frechastraea pentagona minima (Rozkowska 1953); Schröder : 67, pl. 3, fig. 13.
94v 2003 Scruttonia minima (Rozkowska, 1953); Fedorowski : 123, pl. 66, fig. 1.
95Holotype. Specimen A98 stored in the Department of Geology from the University of Poznan, Poland. Upper Frasnian of Psie Gorki at Kielce, Holy Cross Mountains, Poland. Pl. 8, fig. 9 in Rozkowska (1953) and pl. 66, fig. 1 in Fedorowski (2003).
96Material. 54 specimens with 95 thin sections. Personal sampling with that of M. Coen and F. Tourneur: Visé MC- 1977-1-D and E; Senzeille MC-1974-14-N74, P37, P50, P53, P54, P55, P57, T863, T864, T865 and T87; Senzeille MC-1974-15- B29, B30, B33, N54, T881, T882, T883, T885, T8810, T891, T892, T893 and T894; Froidchapelle MC-71-C939 and C949; Froidchapelle MC-75-C993B and D115; Surice MC-1986-5-A523, A524, B18, B40, B41 and B42; Rance MC-1978-17-1, 3, 4, 5, 6, V91, V92, V93, V94, V97A and V98; Rance MC-1978-18-W1, W2, W3, W6, W8, W10 and W12.
97Diagnosis. An astreoid to pseudocerioid species of Frechastraea with 16 to 22 septa at tabularial diameters of 0.7 mm to 1.3 mm. Major septa reaching the axis of the corallites. Inner wall systematically present at the border of the tabularium.
98Description. The material consists of complete and fragmentary colonies which are platy, sheet-like and rarely tabular or nodular. Their height varies between 1 cm and 4.5 cm and reaches 6 cm in one specimen whereas the largest piece has an area of 15 cm x 11 cm. On colony is encrusted by a laminar stromatoporoid and another one by an auloporid. The corallites not regularly polygonal in shape are separated by a zigzagged pseudotheca which is weak to more or less strong; occasionally, they are connected by confluent septa. So the colonies are astreoid to pseudocerioid or locally thamnasterioid. Among the material from Beauchâteau quarry at Senzeille and Cerfontaine, the look is more often pseudocerioid with sometimes a straight pseudotheca (Plate 2A-B). A few lateral, pericalicinal and axial offsets have been observed.
99The septa are usually non-carinate, slightly dilated in the dissepimentarium and thin in the tabularium. They are more thickened and contiguous at the very boundary between the dissepimentarium and the tabularium where an inner wall is formed; this continuous ring is present in both transverse and longitudinal sections. The major septa reach the axis of the corallites where they are fusing laterally, more or less completely axially or forming pseudofossulae. In the centre of the tabularium, there are occasionally a pseudocolumella, a pseudoaulos, a plane of bilateral symmetry or an open space; in rare cases, the major septa are hardly extending into the tabularium. The minor septa traverse all the dissepimentarium; sometimes, they are barely projecting into the tabularium.
100The dissepimentarium is composed of 3 to 6 or even 2 to 8 rows of globose and occasionally flattened dissepiments which are arranged in horizontal layers. At the border of the tabularium, the inner wall consists of a very narrow tight fan of septal trabeculae. The tabulae, which may be disrupted by axial ends of major septa, are incomplete and intersecting laterally with sometimes a flat-topped axial part; they are occasionally horizontal or concave and rarely convex.
101There are 14 to 24 septa per corallite. The width of the tabularium ranges from 0.6 mm to 1.4 mm. The diameter of the corallites varies commonly between 2 mm and 4 mm and more generally between 1.4 mm and 4.5 mm.
102Discussion. Frechastraea minima has been erected by Rozkowska (1953) as a subspecies of F. pentagona. It is separated from this taxon by astreoid colonies which are occasionally pseudocerioid or even locally thamnasterioid, by the systematic occurrence of an inner wall in both transverse and longitudinal sections and by slightly smaller septal number and diameters of the tabularia and corallites. Pseudocerioid colonies with an inner wall and rather small corallites such as that illustrated Plate 2A-B are now assigned to F. minima contrary to the opinion of Coen-Aubert (1987) who attributed them to F. pentagona. This also the case for some specimens identified by Tsien (1968, 1975, 1977, 1978) as F. pentagona. For the same reason, the colony described by Tsien (1978) as Phillipsastrea sp. 1 is referred to Frechastraea minima. F. frocapella (Tsien, 1978) was already considered as a synonym of F. minima by Coen-Aubert (1987). The pseudocerioid colony from the Upper Frasnian of the Holy Cross Mountains in Poland ascribed by Rozkowska (1953, pl. 8, fig. 8) to F. micrommata is probably a representative of F. minima, but it has not been figured in longitudinal section. The material illustrated by Birenheide (1978) and Kong & Huang (1978) as F. minima is different from this species, by several characters and more particularly by the absence of an inner wall.
103Distribution. The Belgian material has been collected by the author, M. Coen and F. Tourneur in the Upper Frasnian of the Visé area and the Philippeville Massif where it is present in the les Valisettes Member at Neuville, Senzeille, Cerfontaine and Vodelée. However, Frechastraea minima occurs also at the top of the red bioherms of the Petit-Mont Member in the Haumont quarry at Vodelée and in the Rance Anticline.
104Outside Belgium, the species is known in the Upper Frasnian from South Devon in Great Britain, the Aachen Syncline in Germany and the Holy Cross Mountains in Poland.
105Frechastraea micrommata (Roemer, 1852)
106Plate 3A-C
107*1852 Smithia micrommata n. sp.; Roemer : 197, pl. 51, figs 20 a-b.
108non 1885 Phillipsastrea pentagona Goldf. sp. var. micrommata Ferd. Roemer; Frech : 56, pl. 3, figs 11-13, pl. 8, fig. 1.
109non 1952 Phillipsastraea micrommata A. Röem.; Ivania : 134, pl. 1, fig. 1, pl. 4, fig. 3, pls. 7-8.
110non 1953 Phillipsastraea pentagona Goldfuss var. micrommata (F. Roemer); Rozkowska : 66, pl. 8, fig. 8.
111non 1955 Phillipsastraea micrommata (A. Roemer); Ivania : 223, pl. 43, fig. 1.
112non 1958 Phillipsastraea pentagona Goldf. var. micrommata (F. Roemer); Bulvanker : 123, pl. 60, figs 1-3, pl. 61, figs 4-5.
113non 1959 Phillipsastraea pentagona (Goldfuss) var. micrommata (F. Roemer); Middleton : 156, fig. 6f.
114non 1960 Phillipsastraea micrommata F. Roemer, 1883; Zheltonogova & Ivania : 382, pl. D-35, fig. 2.
115non 1965 Phillipsastraea micrommata F. Roemer; Ivania : 182, pl. 99, figs 421-422.
116non 1965 Phillipsastraea aff. pentagona Goldfuss var. micrommata (Roemer); Bulvanker : pl. 22, fig. 1.
117non 1967 Phillipsastraea pentagona micrommata (Roem.); Nikolaev & Rzonsnitskaya : pl. 7, fig. 13.
118v p. 1967 Phillipsastraea pentagona (Goldfuss); Sorauf : 29, figs 13, 1e-f (non figs 13, 1a-d).
1191968 Frechastraea micrommata (C.F. Roemer); Scrutton : 240, pl. 8, figs 4-5.
120v p. 1968 Phillipsastraea goldfussi (de Vern. et H .); Tsien : pl. 3, fig. 10 (non pl. 1, figs 3-4, pl. 4, fig. 6, pl. 5, fig. 2).
1211971 Phillipsastraea goldfussi (Edwards and Haime) var. minima; Tsien : pl. 32, fig. 1b.
122v 1974a Phillipsastrea pentagona micrommata (Roemer,C.F., 1852); Coen-Aubert : 16, pl. 2, figs 6-7.
123v 1975 Phillipsastrea micrommata; Tsien : fig. 26k.
124v 1977 Phillipsastrea micrommata; Tsien : figs 8a-b.
125v 1978 Phillipsastrea micrommata (Roemer, 1852); Tsien : 205, fig. 5.
1261978 Frechastraea micrommata (C.F. Roemer 1852); Birenheide : 103, fig. 54.
127Lectotype. Specimen n° 34 chosen by Scrutton (1968) and stored in the collections from the Paläontologisches Institute of the University of Bonn in Germany. Devonian, probably Frasnian of Ferques in the Boulonnais, France. Pl. 51, figs 20a-b in Roemer (1852), pl. 8, figs 4-5 in Scrutton (1968) and fig. 54 in Birenheide (1978).
128Material. 13 specimens with 22 thin sections. Personal sampling with that of H.H. Tsien: Senzeille MC-1974-14-N73; Froidchapelle MC-71-C950; Froidchapelle MC-75-C993A and C997; Beauchâteau quarry at Senzeille 6455. Old collection from the Institut royal des Sciences naturelles de Belgique: Senzeille 6816a, b-F2i -15682; Senzeille 6839-F2i-15760, 15761 and 15769; Senzeille 6840 (= Beauchâteau quarry)-F2i-15592; Senzeille 6854-F2i-15826; Saint-Georges 7006-F2IIIc-16801; Saint-Georges 7010-F2IIIc-16748.
129Diagnosis. A pseudocerioid species of Frechastraea with 20 to 22 septa at tabularial diameters of 1.1 mm to 1.5 mm. Septa non-carinate, rather slender and restricted to the dissepimentarium. Inner wall systematically present at the border of the tabularium.
130Description. The material consists of rather small colonies whose height varies between 1.5 cm and 4 cm. The corallites polygonal in shape are separated by a zigzagged or rarely straight pseudotheca. In a few specimens, the outer zigzagged wall is locally weaker giving a more or les astreoid look. Some lateral offsets have been observed.
131The septa are non-carinate and rather slender or slightly dilated in the dissepimentarium to which all of them are normally restricted. In a few colonies, some major septa are hardly projecting into the tabularium where they may be thinner; pseudofossulae are only occurring in some corallites of one specimen. A thin inner wall is nearly systematically present a the border of the tabularium and it is formed by the thickening of the inner septal ends and the inner row of dissepiments.
132The dissepimentarium consists of 3 to 6 or even 2 to 8 rows of globose and more or less flattened dissepiments which are arranged in horizontal layers. A thin inner wall is usually present at the border of the tabularium, without any trace of septal trabeculae. The tabulae are horizontal, concave or incomplete and intersecting laterally; they are rarely convex.
133There are 18 o 24 septa per corallite. The width of the tabularium ranges from 1 mm to 1.6 mm. The diameter of the corallites varies commonly between 3 mm and 6 mm and more generally between 2.8 mm and 6.8 mm, but it may reach 7.4 mm.
134Discussion. The lectotype of Frechastraea micrommata figured by Roemer (1852) and Scrutton (1968) is clearly characterized by septa restricted to the dissepimentarium and by the occurrence of an inner wall at the border of the tabularium. All the references that do not correspond to this situation, are excluded from the list of synonymy; in many of them, the major septa are reaching the axis of the corallites. This feature is also present in F. pentagona which differs additionally from F. micrommata by the lack of inner wall and flattened dissepiments. There are some similarities between F. micrommata and F. glabra from the base of the Belgian Upper Frasnian; however, the latter species is typically astreoid and once more has no inner wall.
135Distribution. The Belgian material collected by the author and H.H.Tsien comes from the Upper Frasnian Les Valisettes Member at Neuville, Senzeille and Cerfontaine, in the Philippeville Massif.
136Outside Belgium, it seems that the lectotype has been found in the Frasnian of the Boulonnais in France.
137Frechastraea kaisini (Tsien, 1978)
138(Plate 2H-I)
139v p. 1967 Phillipsastraea pentagona minima Rozkowska; Sorauf : 31, fig. 14, 1c (non figs 14, 1a-b).
140v p. 1968 Phillipsastraea pentagona (Goldfuss); Tsien : pl. 5, fig. 1 (non pl. 2, figs 3-4, pl. 3, fig. 11).
1411975 Phillipsastrea kaisini; Tsien : fig. 26l.
1421977 Phillipsastrea kaisini nov. sp.; Tsien : fig. 8c.
1431978 Phillipsastrea kaisini nov. sp.; Tsien : p. 206, fig. 8, pl. 2, figs 2a-b.
144Holotype. Specimen n° 6849 formerly stored in the Laboratory of Palaeontology from the Catholic University of Louvain at Louvain-la-Neuve, Belgium. One longitudinal thin section belonging probably to the same colony is now stored in the Collection of Palaeontology from the Institut royal des Sciences naturelles de Belgique at Brussels. Beauchâteau quarry at Senzeille in the Philippeville Massif, Belgium. Les Valisettes Member of the Champ Broquet Formation, Upper Frasnian. Pl. 5, fig. 1 in Tsien (1968), fig. 26l in Tsien (1975), fig. 8c in Tsien (1977) as well as fig. 8 and pl. 2, figs 2a-b in Tsien (1978).
145Material. 6 specimens with 9 thin sections. Personal sampling with that of H.H. Tsien: Senzeille MC-1974-14-T862; Froidchapelle MC-75-D118; Beauchâteau quarry at Senzeille 6849. Old collection from the Institut royal des Sciences naturelles de Belgique: Senzeille 6816a, b-F2i-15686; Senzeille 6840 (= Beauchâteau quarry)- F2i-15626; Senzeille 6854-F2i-15848.
146Diagnosis. An astreoid species of Frechastraea with 18 to 20 septa at tabularial diameters of 0.5 mm to 1 mm. Septa non-carinate and restricted to the dissepimentarium. Inner wall systematically present at the border of the tabularium.
147Description. The material consists of small platy and sheet-like colonies which are complete or fragmentary. Their height varies between 1 cm and 3.5 cm whereas the largest piece has a diameter of 5 cm. The corallites not regularly polygonal in shape are connected by a weak zigzagged pseudotheca or by confluent septa so that the colonies are sometimes thamnasterioid. Very few lateral offsets have been observed.
148The septa are non-carinate and dilated in the dissepimentarium to which all of them are restricted. They are more thickened and contiguous at the very boundary between the dissepimentarium and the tabularium where an inner wall is formed; this continuous ring is present in both transverse and longitudinal sections.
149The dissepimentarium consists of 2 to 5 rows of globose dissepiments which are arranged in horizontal layers. The tabulae are horizontal or concave; they are occasionally convex or incomplete and intersecting laterally.
150There are 18 to 20 septa per corallite. The width of the tabularium ranges from 0.5 mm to 0.7 mm, but it often reaches 0.9 mm to 1 mm in one colony. The diameter of the corallites varies between 1.5 mm and 2.6 mm or even 3 mm.
151Discussion. Frechastraea kaisini is related to F. minima from which it mainly differs by major septa restricted to the dissepimentarium. It is separated from F. micrommata by astreoid to thamnasterioid colonies, by septa typically dilated in the dissepimentarium and by smaller tabularia and corallites.
152Distribution. The species is only known in the Upper Frasnian of Belgium where it comes from the Les Valisettes Member in the Philippeville Massif.
153Frechastraea cf. tungkanlingensis (Yoh, 1937)
154(Plate 3D-F)
155cf. 1937 Prismatophyllum pentagonum var. nov. tungkanlingense; Yoh : 66, pl. 8,fig. 3.
156v 1985 Frechastraea pentagona tungkanlingense; Coen-Aubert & Lacroix : 125.
157Lectotype (here chosen, but not traced). Specimen figured by Yoh (1937, pl.8, fig. 3) and coming from the Givetian of the Tungkanling Hill near the town Hsiang Hsien, Guangxi in China.
158Material. 7 specimens with 11 thin sections. Personal sampling with that of M. Coen: Braives MC-1979-2-W51 and W51A; Fléron MC-1974-75-F75; Esneux MC-1974-89-F80 and F82; Rance MC-132-R2; Agimont MC-129-Heer 4.
159Diagnosis. A pseudocerioid species of Frechastraea with 26 to 28 septa at tabularial diameters of 2 mm to 2.5 mm. Septa poorly carinate and restricted to the dissepimentarium without any trace of inner wall.
160Description. The material consists of nodular and tabular colonies which are complete or fragmentary. Their height varies between 2.5 cm and 11 cm whereas the largest piece has an area of 12 cm x 8 cm. The corallites usually polygonal in shape are separated by a straight or zigzagged pseudotheca. A few pericalicinal offsets have been observed.
161The septa are non-carinate or bear a few spinose and knobbly carinae. They are rather slender or slightly dilated in the dissepimentarium to which all of them are normally restricted. Sometimes, the septa are more thickened at the border of the tabularium. In some specimens, a few major septa are hardly projecting in the tabularium where they often become thinner; rare pseudofossulae are also occurring.
162The dissepimentarium consists of 3 to 7 or even 2 to 9 rows of globose dissepiments which are arranged in horizontal layers. Occasionally, a few inclined or specialized dissepiments are present at the border of the tabularium. The tabulae are complete or incomplete and intersecting laterally with a frequent concave pattern; sometimes, they are also horizontal.
163There are 22 to 26 and even 20 to 28 septa per corallite. The width of the tabularium ranges from 1.3 mm to 1.9 mm and even from 1.1 mm to 2.1 mm. The diameter of the corallites varies commonly between 3 mm and 5.5 mm and more generally between 2.5 mm and 6.2 mm.
164Discussion. The Belgian material assigned to Frechastraea cf. tungkanlingensis is not very abundant and homogeneous. It is distinguished from the lectotype of the species by narrower tabularia, slightly fewer septa and the concave pattern of the tabulae. The Belgian colonies with septa thicker in the dissepimentarium (Plate 3D) resemble more the type specimen of Yoh (1937). F. cf. tungkanlingensis is separated from F. micrommata by the absence of an inner wall, a few carinae, somewhat larger tabularia and more closely spaced tabulae which are incomplete and concave. F. coeni from the base of the Upper Frasnian in Belgium is also related to F. cf. tungkanlingensis, but it is typically carinate and characterized by slightly smaller septal number and diameter of the tabularium. F. tungkanlingensis has been compared by Chen (1959) to F. regularis (Chen, 1959) from the Frasnian of Guizhou Province in China which is closer to F. coeni as discussed by Coen-Aubert (2012).
165Distribution. Frechastraea cf. tungkanlingensis has been collected by the author and M. Coen in different localities and deposits from the Upper Frasnian of Belgium:
166- in the upper limestone level of the Aisemont Formation at Huccorgne, on the north side of the Namur Basin and at Prayon, in the Vesdre Massif;
167- at the base of the Lambermont Formation in Hony, on the north side of the Dinant Synclinorium;
168- in the Petit-Mont Member at Heer-Agimont, in the Philippeville Massif and in the Renlies Anticline.
169Outside Belgium, the lectotype of F. tungkanlingensis comes from the Givetian of Guangxi in China.
170Frechastraea crassiseptata (Tsien, 1978)
171(Plate 3G-H; Plate 4A-F; Plate 5H)
172v p. 1974a Phillipsastrea pentagona pentagona (Goldfuss, G.A., 1826); Coen-Aubert : 10, pl. 1, fig. 1 (non pl. 1, figs 4-6).
173v 1974a Phillipsastrea cf. pentagona pentagona (Goldfuss, G.A., 1826); Coen-Aubert : pl. 1, figs 2-3.
1741975 Phillipsastrea senceliae; Tsien : fig. 26m.
1751975 Phillipsastrea bethunei; Tsien : fig. 26n.
176v 1977 Frechastraea pentagona micrastraea; Coen et al. : 326.
177v 1977 Frechastraea pentagona micrastraea (Penecke, 1904) ; Coen-Aubert : 53.
1781977 Phillipsastrea bethunei nov. sp.; Tsien : figs 8d-e.
1791977 Phillipsastrea senceliae nov. sp.; Tsien : figs 8f-g.
180* v 1978 Phillipsastrea crassiseptata nov. sp.; Tsien : 207, pl. 2, figs 4a-b.
1811978 Phillipsastrea senceliae nov. sp.; Tsien : 206,figs 11b-12a.
1821978 Phillipsastrea bethunei nov. sp.; Tsien : 207, figs 11a-12b.
183v 1982 Frechastraea pentagona micrastraea (Penecke, K.A.,1904); Coen-Aubert : 9.
184v 1991 Frechastraea pentagona micrastraea (Penecke,1904); Birenheide et al. : 4, 26.
185v 2012 Frechastraea micrastraea (Penecke, 1904); Coen-Aubert : 265-266, fig. 2.
186Holotype. Specimen n° 6847 and thin sections 6847IA and IB formerly stored in the Laboratory of Palaeontology from the Catholic University of Louvain at Louvain-la-Neuve, Belgium. Two thin sections belonging probably to the same colony are now stored in the Collection of Palaeontology from the Institut royal des Sciences naturelles de Belgique at Brussels. Beauchâteau quarry at Senzeille in the Philippeville Massif, Belgium. Upper part of the Petit-Mont Member from the Champ Broquet Formation, Upper Frasnian. Pl. 2, figs 4a-b in Tsien (1978).
187Material. 92 specimens with 170 thin sections. Personal sampling with that of M. Coen, E. Groessens and F. Tourneur: Saint-Georges MC-1978-5-V15, V17, V18, V20 and V22; Chênée MC-1974-3-L16 and L17; Chênée MC-1974-1-H92; Chênée MC-1974-82-M540; Chênée MC-1974-2-N28 and N29; Fléron MC-1974-74-H49; Fléron MC-1974-5-428; Verviers MC-1974-6-4; Verviers MC-1974-65-9, 10 and 209; Verviers MC-1974-56-483; Verviers MC-1988-4-A670, A673 and A679; Spa MC-1974-92-1; Yvoir MC-1974-130-P59; Louveigné MC-1974-90-N20 and N21; Louveigné MC-1974-10-l. 11971 and l. 11985; Hamoir MC-133-l. 8582 and l. 8583; Hamoir MC-120-D825, D846, D851, D857, D860, D861 and D862; Grandhan MC-39-B718, B720, C120 and C121; Walcourt MC-1976-5-U27; Beaumont MC-134-795, 796 and l. 14456; Grandrieu MC-1975-11-T7; Senzeille MC-1976-4-LB9 and LB10; Senzeille MC-1984-1-A512, A513, A514 and A516; Senzeille MC-1974-15-B1, B2, B7, B8A, B9, B10, B10A, B11, B13, B18, B18A, B23, B24, B25, B34, B35, T90, T91 and T92; Surice MC-1974-133-V14, V16, V17, V19, V24, V25 , V27 and V28; Surice MC-1986-5-B21, B23, B24, B25, B26, B27, B28, B31, B32, B33, B34 and B35; Agimont MC-129-Heer 1 and Heer 5.
188Diagnosis. A pseudocerioid to locally astreoid species of Frechastraea with 20 to 26 septa at tabularial diameters of 1.2 mm to 2.4 mm. Septa non-carinate and variably but typically dilated in the dissepimentarium. Major septa reaching the axis of the corallites. Possible occurrence of some horseshoe dissepiments.
189Description. The material consists of platy, tabular, nodular and massive colonies which are frequently fragmentary and which may reach large dimensions. Their height varies between 1 cm and 11 cm or even 14 cm whereas the largest piece has an area of 20 cm x 25 cm. The corallites polygonal in shape are separated by a straight or zigzagged pseudotheca. In several specimens, the outer zigzagged pseudotheca is locally weaker and rarely absent giving a more or less astreoid look. In one colony, the outer wall is often characterized by a dark median line. Some lateral and pericalicinal offsets have been observed.
190The septa are normally non-carinate; nevertheless, a few knobbly and more rarely yardarm carinae occur in some colonies. In the main part of the material, the septa are uniformly and typically dilated in the dissepimentarium (Plate 4A; Plate 5H) and they become thin in the tabularium or slightly beyond their entry into it. However, there are also septa more thickened or spindle-shaped thickened at the inner border of the dissepimentarium (Plate 4C) or even throughout its inner part. In very few specimens, the dilation of the septa is rather weak in the dissepimentarium. Moreover, some transverse sections from the Philippeville Massif are characterized by corallites with spindle-shaped thickened septa together with corallites whose septa are only slightly dilated in the dissepimentarium (Plate 4E).
191The major septa reach the axis of the corallites where there are frequently fusing partially or totally and forming pseudofossulae. In the centre of the tabularium, there are sometimes an open space, a pseudoaulos or a plane of bilateral symmetry; in rare cases, the major septa are hardly projecting into the tabularium. The minor septa traverse the entire dissepimentarium; they are rarely shorter, discontinuous or lacking.
192The dissepimentarium consists of 1 to 7 or even 10 rows of globose dissepiments which are arranged in horizontal layers with occasionally 1 or 2 series of inner inclined dissepiments. Small horseshoe and specialized dissepiments occur rather often at the border of the tabularium or close to it with locally 1 or 2 rows of inner dissepiments. In a few colonies, the narrow dissepimentarium of some corallites is restricted to 1 or 2 series with more or less flat, peneckielloid and horseshoe dissepiments (Plate 3H; Plate 4F). The tabulae, which are frequently disrupted by axial ends of major septa, are incomplete and intersecting laterally with sometimes a flat-topped axial part; they are rarely horizontal, concave or convex.
193There are 18 to 30 septa per corallite. The width of the tabularium ranges from 0.9 mm to 2.7 mm, but it may reach 2.8 mm to 3.3 mm in three colonies of Vodelée. The diameter of the corallites varies commonly between 2.5 mm and 5 mm and more generally between 2 mm and 5.5 mm or even 6.5 mm.
194Discussion. The Belgian material assigned herein to Frechastraea crassiseptata shows a wide variability. Colonies of this taxon are mostly pseudocerioid, but they may be locally astreoid. Very often, their septa are uniformly dilated throughout the dissepimentarium; sometimes, this thickening is weaker or more or less spindle-shaped at the border of the tabularium. Normally, the dissepiments are globose, but some small horseshoe dissepiments occur also. For this material, Tsien (1978) described and figured three new species: F. crassiseptata, F. bethunei (Tsien, 1978) and F. senceliae (Tsien, 1978) coming all three from the upper part of the Petit-Mont Member in the Beauchâteau quarry at Senzeille. The two latter taxa were illustrated in transverse section by Tsien (1975) and their holotypes were figured in transverse and longitudinal sections by (Tsien, 1977). Figures of the holotype of F. crassiseptata were only given by Tsien (1978). Additionally, there is some confusion between the illustrations of the holotypes of F. bethunei and F. senceliae in Tsien (1978) whose descriptions correspond clearly to the figures of their holotypes presented by Tsien (1977). That is to say that the holotype of F. bethunei corresponds to figs 11a and 12b of Tsien (1978) whereas the holotype of F. senceliae corresponds to figs 11b and 12a of the same paper.
195The species F. crassiseptata is used herein because there is no doubt about its holotype and its general look with septa uniformly dilated in the dissepimentarium and globose dissepiments (Plate 4A-B; Plate 5H) is more common among the Belgian material collected in different Upper Frasnian deposits and facies. F. senceliae is characterized by spindle-shaped dilated septa at the border of the tabularium (Plate 4C) and some small horseshoe dissepiments. As for F. bethunei, it has a narrow dissepimentarium with some horseshoe dissepiments associated with some more or less flat and peneckielloid dissepiments whereas its septa are faintly dilated in the dissepimentarium. As mentioned in the description, the characteristics of F. senceliae and F. bethunei can be observed in a same colony or together with corallites typical of F. crassiseptata (Plate 3G; Plate 4E). So it is not possible to make a clear distinction between the three synonymous species F. crassiseptata, F. senceliae and F. bethunei.
196The Belgian material ascribed herein to F. crassiseptata has been identified as F. micrastraea (Penecke, 1904) by Coen et al. (1977), Coen-Aubert (1977, 1982 and 2012) and Birenheide et al. (1991). F. micrastraea is only known by one specimen which comes from the Frasnian of Anti Taurus in Turkey and which has been illustrated by Penecke (1904, pl. 6, fig. 3, pl. 7, fig. 2; not traced). This colony has more the look of F. bethunei as it is characterized by a narrow dissepimentarium and a weak thickening of the septa within the dissepimentarium. From the same area in Turkey, Hubmann (1992) described one sample as Mixogonaria schafferi (Penecke, 1904) and compared it with Frechastraea bethunei; this specimen resembles F. crassiseptata though it has some small carinae. The original material of F. schafferi investigated by Penecke (1904) differs from F. crassiseptata by septa faintly carinate and dilated in the dissepimentarium and by larger tabularia and corallites. The relations between the genera Frechastraea and Mixogonaria Kong, 1978 have already been discussed by Coen-Aubert (2012).
197At the Belgian scale, Frechastraea crassiseptata is separated from F. pentagona by pseudocerioid to astreoid colonies, by septa more strongly thickened in the dissepimentarium with some spindle-shaped dilation at its inner border, by more frequent horseshoe dissepiments and by slightly greater diameters of the tabularia for corallites of same size. There are some affinities between F. crassiseptata and F. pollicaris McLean, 1994 from the Middle Frasnian of the Northwest Territories in Canada. In the latter species however, the septa are more coarsely dilated in the dissepimentarium with some small carinae and there is more often an open space in the centre of the tabularium.
198Despite the occurrence of more characteristic horseshoe dissepiments in parts of some Belgian colonies, F. crassiseptata is closely related to F. pentagona and must be assigned to Frechastraea due to its pseudocerioid aspect, its septa more or less uniformly thickened throughout the dissepimentarium and its small corallites. Phillipsastrea D’Orbigny, 1849 whose type species is P. hennahi (Lonsdale, 1840) from the Upper Givetian of the South Devon in Great Britain, is different as it is astreoid to thamnasterioid with septa spindle-shaped dilated in a wide zone of the inner dissepimentarium and more systematic horseshoe dissepiments. Diverse species of the genus Phillipsastrea from the Givetian of Spain, Morocco and Mauritania, which show these features, have been described recently by Coen-Aubert (2013).
199Distribution. The species is only known in the Upper Frasnian of Belgium where it has been collected in different coral deposits by the author, M. Coen, E. Groessens and F. Tourneur. It is already present in the lower limestone level of the Aisemont Formation, but it is more abundant in the upper one, on the south side of the Namur Basin, the Streupas outlier, the Vesdre Massif, the Theux Window as well as the north and east sides the Dinant Synclinorium. At Laneffe, on the north side of the same structural unit, one colony has been observed near the base of the Les Valisettes Member, according to the description given by Boulvain et al. (1999). F. crassiseptata is common in the red bioherms of the Petit-Mont Member from Solre-Saint-Géry and Leugnies, in the northwestern part of the Dinant Synclinorium and from Neuville, Senzeille, Vodelée and Heer-Agimont in the Philippeville Massif. The species is also widespread on the east side of the Dinant Synclinorium, in the red bioherms of Sy, Bomal and Grandhan.
200Frechastraea limitata (Milne-Edwards & Haime, 1851)
201(Plate 5A-G)
202 * 1851 Acervularia limitata; Milne-Edwards & Haime : 419.
2031853 Acervularia limitata; Milne-Edwards & Haime : 238, pl. 54, figs 1-1a.
204v non 1953 Phillipsastraea goldfussi Edwards & Haime; Rozkowska : 62, pl. 8, figs 5-6.
2051960 Phillipsastraea limitata (M. Edwards et J. Haime, 1851); Ermakova : 89, pl. 7, figs 8-9.
206non 1960 Phillipsastraea limitata M.-Edwards et Haime, 1851; Zheltonogova & Ivania : 382, pl. D-35, fig. 1.
207non 1965 Phillipsastraea limitata M.-Edwards et Haime; Ivania : 184, pl. 84, figs 363-364.
2081967 Haplothecia ogwellensis sp. nov.; Scrutton : 272, pl. 41, figs 3-6.
209non 1967 Phillipsastraea goldfussi (de Verneuil & Haime); Sorauf : 27, figs 10 a-f, 11 1a-b.
210v 1974a Phillipsastrea limitata (Edwards, H.M. et Haime, J., 1851); Coen-Aubert : 17, pl. 3, figs 1-4.
211v 1974b Phillipsastraea goldfussi (Verneuil et Haime); Coen-Aubert : pl. 3, fig. 4.
212non 2003 Frechastraea goldfussi (Verneuil et Haime, 1850); Fedorowski : 121, pl. 64, figs 1-5.
213? 2005 Frechastraea (Frechastraea) limitata (Milne-Edwards & Haime, 1850); Schröder : 80, pl. 12, figs 2-4.
214v 2014 Frechastraea limitata (Milne-Edwards & Haime, 1851); Coen-Aubert : fig. 3B.
215Remark. A more complete list of synonymy about the references before 1953 has been provided by Coen-Aubert (1974a).
216Lectotype. Specimen chosen by Soshkina (1951, p. 97) and coming from the Pengelly collection stored in the Geological Survey Museum of London in Great Britain; according to Scrutton (1968, p. 249), this specimen is lost. Devonian from Newton Quarry near Torquay, South Devon, Great Britain. Pl. 54, figs 1, 1a in Milne-Edwards & Haime (1853).
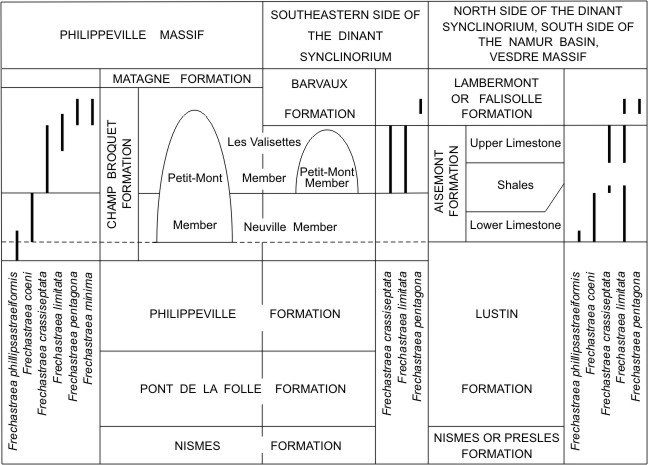
217Material. With the exception of Chaudfontaine L15 and Neuville 5 assigned to Frechastraea phillipsastraeiformis (Moenke, 1954) as well as of Prayon H49 assigned herein to F. crassiseptata, all the colonies mentioned by Coen-Aubert (1974a). In addition to this material, 26 specimens with 46 thin sections. Personal sampling with that of M. Coen and E. Groessens: Booischot borehole at 871 m; Braives MC-1979-2-W47; Fontaine l’Evêque MC-1974-137-L8; Huy MC-1977-8-U51 and U52; Saint-Georges MC-1978-5-V1, V81, V9 and V24; Chênée MC-1974-83-H59 and H68; Verviers MC-1974-56-403; Verviers MC-1988-4-A672, A674, A677, A678 and A681; Nannine MC-1974-115-R61; Hamoir MC-120-D842, D843 and D850; Barvaux l. 10582; Beaumont MC-1979-9-G9 and G13; Senzeille MC-1974-15-B22 and T888.
218Diagnosis. A pseudocerioid species of Frechastraea with 22 to 28 septa at tabularial diameters of 1.4 mm to 2.6 mm. Septa dilated and variably but typically carinate in the dissepimentarium. Major septa reaching the axis of the corallites.
219Description. The material consists of platy, tabular and nodular colonies which are frequently fragmentary. Their height varies between 1 cm and 9 cm whereas the largest piece reaches an area of 14 cm x 7 cm. The corallites usually polygonal in shape are separated by a strong zigzagged or sometimes straight pseudotheca. A few lateral and pericalicinal offsets have been observed.
220The septa bear small knobbly and yardarm carinae which are more or less numerous among the different corallites of a same colony. They are typically dilated in the dissepimentarium and become thin in the tabularium or slightly beyond their entry into it. In some specimens, a spindle-shaped thickening of the septa is locally present at the border of the tabularium.
221The major septa reach more or less the axis of the corallites. Their axial ends are often forming pseudofossulae or fusing axially, partially and even totally. In the centre of the tabularium are occasionally seen a small open space, a plane of bilateral symmetry, a few short fragments of septa and rarely an incomplete pseudoaulos. The minor septa traverse the entire dissepimentarium; in some corallites, they are hardly projecting into the tabularium.
222The dissepimentarium consists of 3 to 8 or even 2 to 10 rows of small globose dissepiments which are arranged in horizontal layers. In some colonies, inclined and very rarely specialized dissepiments occur locally at the border of the tabularium. Spots and more frequently broad asymmetrical fans of isolated or sometimes tight septal trabeculae are extending across all the dissepimentarium. The tabulae incomplete and intersecting laterally are rather often disrupted by axial ends of major septa; they may be also horizontal, concave, convex or characterized by a flat-topped axial part.
223There are 20 to 30 septa per corallite. The width of the tabularium ranges from 1.2 mm to 3 mm. The diameter of the corallites varies commonly between 3 mm and 6.5 mm and more generally between 2.5 mm and 8 mm.
224Discussion. The Belgian material described herein as Frechastraea limitata is very homogeneous and is clearly similar to the lectotype of Milne-Edwards & Haime (1853) and to the type specimens of F. ogwellensis (Scrutton, 1967) so that the two species already compared by Coen-Aubert (1974a) are now considered as synonyms. The colonies of F. ogwellensis investigated by Scrutton (1967) come from the Upper Frasnian of the road cutting, 36.5 m west of Ramsleigh Quarry entrance at East Ogwell near Newton Abbot, South Devon in Great Britain. In the old collection of the Institut royal des Sciences naturelles de Belgique, there is a transverse thin section from Ramsleigh (n° 329) labelled as F. limitata. So the Newton Quarry of Milne-Edwards & Haime (1853) for the lectotype of F. limitata could correspond to the Ramsleigh Quarry near Newton Abbot. The holotype of F. carinata Scrutton, 1968 has been collected in the same outcrop as that of F. ogwellensis. It differs from this species and from F. limitata by a weaker carination and by somewhat smaller septal number and diameter of the tabularium. As mentioned by Coen-Aubert (2012), the paratype of F. carinata figured by Scrutton (1968, pl. 9, fig. 2) is closer to F. coeni. This taxon is distinguished from F. limitata by major septa restricted to the dissepimentarium and by slightly fewer septa and narrower tabularia. At the Belgian scale, F. limitata is also related to F. crassiseptata from which it is separated by the systematic occurrence of carinae, the absence of horseshoe dissepiments and by slightly greater septal number and diameters of the tabularia and corallites.
225Concerning the list of synonymy, the specimen from the Upper Frasnian of the Kuznetsk Basin in Russia illustrated by Zheltonogova & Ivania (1960) and Ivania (1965) is different from F. limitata in often having a straight wall and inclined dissepiments whereas the material figured by Schröder (2005) seems to be transitional to F. phillipsastraeiformis as it is characterized by some spindle-shaped dilation of the septa.
226A few colonies with a zigzagged pseudotheca, carinate septa and small corallites recall the general look of F. limitata. These are the holotype of Haplothecia (Lanceothecia) punctata hexagona Yu & Jiang in Jiang (1982) from the Frasnian of Hunan in China, the holotype and one paratype of Kuangxiastraea mirabilis McLean (2010) figured by McLean (2010, pl. 43, figs 1-6) and coming from the Upper Frasnian of the British Columbia and the Northwest Territories in Canada, one paratype of Hexagonaria arietina McLean, 2010 figured by McLean (2010, pl. 36, figs 1-2) and coming from the Upper Frasnian of Alberta in Canada.
227Distribution. The species has been collected by the author, M. Coen and E. Groessens, in different coral deposits from the Upper Frasnian of Belgium. In the Campine Basin, the Namur Basin, the La Tombe and Kinkempois outliers, the Vesdre Massif as well as on the north and east sides of the Dinant Synclinorium, it is already present in the lower limestone level of the Aisemont Formation, but it is more abundant in the upper one; it occurs locally at the base of the overlying Lambermont Formation. Frechastraea limitata has also been sampled at the base of the Neuville Member from Barbençon, in the northwestern part of the Dinant Synclinorium. Additionally, it is widespread on the east side of the same structural unit, in the red bioherms of the Petit-Mont Member from Sy, Bomal, Durbuy and Barvaux. Only two colonies have been observed in the Philippeville Massif, from the Petit-Mont and Les Valisettes Members at Senzeille.
228Outside Belgium, F. limitata is known in the Upper Frasnian from South Devon in Great Britain, in the Frasnian from the Harz Mountains in Germany as well as from the Russian Platform and Timan in Russia. It may be present in the Upper Frasnian from the East Sauerland in Germany.
4. Stratigraphic and palaeobiogeographic conclusions
229The most frequent species of Frechastraea occurring in the Upper Frasnian of Belgium (Fig. 3) are F. coeni, F. crassiseptata, F. limitata, F. pentagona and F. minima. As mentioned by Coen-Aubert (2012), F. coeni is widespread in the lower limestone level of the Aisemont Formation from the Namur Basin, the Vesdre Massif and the north side of the Dinant Synclinorium. But the species is also recorded at the base of the Neuville Member in the northwestern and central parts of the same structural units and in the lower part of the bioherms from the Petit-Mont Member in the Philippeville Massif. In some localities, F. coeni is accompanied by F. glabra, F. limitata and F. phillipsastraeiformis. The latter taxon may be already present in the Philippeville Massif, at the very base of the Neuville Member, that is to say below the Middle-Upper Frasnian boundary, where it is associated with Scruttonia bowerbanki (Milne-Edwards & Haime, 1851) and the last representatives of the genus Hexagonaria Gürich, 1896.
230Figure 3. Stratigraphic distribution of the most frequent species of Frechastraea in the Upper Frasnian from the Philippeville Massif, the southeastern and north sides of the Dinant Synclinorium, the south side of the Namur Basin and the Vesdre Massif. The broken line corresponds to the base of the Upper Frasnian.
231Together with Frechastraea crassiseptata, F. limitata is much more abundant in the upper limestone level of the Aisemont Formation and in the red lenses of the Petit-Mont Member from the southeastern side of the Dinant Synclinorium. F. limitata is rare in the Philippeville Massif and in the lower part of the Lambermont Formation. On the contrary, F. crassiseptata is very common in the bioherms of the Petit-Mont Member from the Philippeville Massif and from Solre-Saint-Géry and Leugnies to the northeast of this structural unit. It is sometimes reported in the upper part of the lower limestone level of the Aisemont Formation when it is very thick. This is the case for sections such as Embourg at the western end of the Vesdre Massif and the nearby Streupas outlier. In the same way, F. crassiseptata is accompanied by F. glabra at the top of the lower limestone level of the Aisemont Formation, at Engis situated in the eastern part from the south side of the Namur Basin.
232F. pentagona is characteristic of the lower part of the Lambermont Formation, the Barvaux Formation and the Les Valisettes Member. In the Philippeville Massif, it is associated with F. minima and occasionally with F. micrommata and F. kaisini and it comes mostly from the accumulations of corals above the Petit-Mont Member. However, F. pentagona and F. minima have also been observed at the top of the Petit-Mont Member, in the Haumont quarry at Vodelée and in the Renlies Anticline. Moreover, F. minima and Phillipsastrea ranciae occur in the Petit-Mont Member at Rance, just below Famennian shales dated by conodonts (Biron et al., 1983).
233As for F. cf. tungkanlingensis, it has been found locally in different facies. Indeed, it is present in the upper limestone level of the Aisemont Formation and at the base of the overlying Lambermont Formation at Hony where it is probably reworked with colonies of F. limitata. But it has also been sampled in the Petit-Mont Member from Heer-Agimont with F. crassiseptata and from Renlies with F. pentagona.
234The stratigraphic succession of the three species F. coeni, F. crassiseptata and F. pentagona is remarkable and very useful for correlations within the Upper Frasnian of Belgium. Above the pure limestones of the Aisemont Formation and the various lenses of the Petit-Mont Member, the sedimentation is mostly argillaceous. However, massive rugose corals are still more or less numerous locally in the Lambermont Formation, the Barvaux Formation and the upper part of the Les Valisettes Member. All these corals seem to disappear before the end of the Frasnian and the Upper Kellwasser Event known in the Palmatolepis linguiformis conodont Zone. As proposed herein in the Dinant Synclinorium, the Champ Broquet Formation is still related to reefal ecosystems as it is the case for the Moulin Liénaux and Grand Breux Formations. At the top of the Frasnian, there are only fine shales without any corals. Maybe, the black shales of the Matagne Formation and the purple shales of the Barvaux Formation should be considered as particular members of a more extended argillaceous formation including Famennian shales. The introduction of the Falisolle Formation on the south side of the Namur Basin, which crosses the Frasnian-Famennian boundary, is a good step in this direction. Such discussions about the distribution of the facies and the lithostratigraphic units from the Upper Frasnian of Belgium are available in the papers of Mottequin (2008a, 2008b), Denayer & Poty (2010), Ernst et al. (2015) and Mottequin & Poty (2015).
235From a palaeobiogeographic point of view, there are clearly many similarities between the Upper Frasnian massive rugose corals from Belgium, South Devon in Great Britain, the Aachen Syncline, which is the continuation of the Vesdre Massif to the east, and the Harz Mountains in Germany and finally the Holy Cross Mountains in Poland. Concerning the species of Frechastraea, F. phillipsastraeiformis and F. minima have been erected in the Holy Cross Mountains. F. pentagona occurs in South Devon together with F. minima and F. limitata, in the Aachen Syncline together with F. minima and in the Harz Mountains together with F. limitata. The latter species has also been described in the Middle to Upper Frasnian of the Russian Platform and Timan in Russia. As noted in the taxonomic discussion of F. pentagona, F. crassiseptata may be present in the Upper Frasnian of the Kuznetsk Basin (Western Siberia) in Russia and F. minima in the Upper Frasnian of Moravia in the Czech Republic where I have in fact collected one colony of the species. The discovery of F. micrommata in the Frasnian of the Boulonnais in France made by Roemer (1852) has not been confirmed until now. The relations between F. cf. tungkanlingensis in Belgium and the Givetian material of Guangxi in China are not very clear. In conclusion, several species such as F. pentagona, F. minima, F. crassiseptata and F. limitata are good markers for the Upper Frasnian in different countries of Western and Eastern Europe.
236The well known Upper Frasnian Crisis has been recently discussed in detail by Mottequin & Poty (2015). As mentioned in this paper, the colonial and dissepimented solitary rugose corals disappeared progressively in the Upper Palmatolepis rhenana Zone, due to the widespread development of dysoxic and anoxic shaly facies; they are of course only present in reefal and coralliferous facies. At the top of the stage, the Upper Kellwasser Event is finally responsible for the last Frasnian extinctions of brachiopods, in the P. linguiformis Zone. Contrary to the opinion of Mottequin & Poty (2015), thamnoporids are still present in the Petit-Mont Member. More particularly, these small branches of tabulate corals are abundant in the red marble quarries of Rance investigated by Biron et al. (1983); the thamnoporids were called “rats’ tails” by the quarrymen and determine the decorative aspect of this ornamental stone used notably in the castle of Versailles, France.
5. Acknowledgements
237Several specimens were collected by Michel Coen, Eric Groessens and Francis Tourneur. The thin sections were made by Arthur Nijs, René Cremers and the Laboratory of Eddy Poty (Liège). Wilfried Miseur and Thierry Hubin helped me for the photographs of the corals. This paper contributes to IGCP 596 on Mid-Paleozoic climate and biodiversity. Ross McLean and Bernard Mottequin kindly reviewed the manuscript. I am most grateful to all these persons. However, I am only responsible for the opinions presented in this paper.
6. References
238Belanger, I., Delaby, S., Delcambre, B., Ghysel, P., Hennebert, M., Laloux, M., Marion, J.M., Mottequin, B. & Pingot, J.L., 2012. Redéfinition des unités structurales du front varisque utilisées dans le cadre de la nouvelle Carte géologique de Wallonie (Belgique). Geologica Belgica, 15, 169-175.
239Besprozvannykh, N.I., Dubatolov, V.N., Kravtsov, A.G., Latypov, Yu. Ya. & Spassky, N. Ya., 1975. Devonskie rugozy Taymyro-Kolymskoy provintsii. Trudy Instituta Geologii i Geofiziki, Akademiya Nauk SSSR, Sibirskoe Otdelenie, 228, 1-172. (In Russian).
240Birenheide, R., 1978. Rugose Korallen des Devon. In Krömmelbein, K. (ed.), Leitfossilien begründet von G. Gürich. 2., völlig neu bearbeitete Auflage, n 2. Gebrüder Borntraeger, Berlin-Stuttgart, 265 pp.
241Birenheide, R., 1998. Rugose und tabulate Korallen aus der Bohrung Viersen 1001. Fortschritte in der Geologie von Rheinland und Westfalen, 37, 161-213.
242Birenheide, R., Coen-Aubert, M., Lütte, B.P. & Tourneur, F., 1991. Excursion B1, Devonian coral bearing strata of the Eifel Hills and the Ardenne. In Lütte, B.P. (ed.), VI. International Symposium on Fossil Cnidaria including Archaeocyatha and Porifera, Excursion-Guidebook. Forschungsstelle für Korallenpaläozoologie, Münster, 113 pp.
243Biron, J. P., Coen‑Aubert, M., Dreesen, R., Ducarme, B., Groessens, E. & Tourneur, F., 1983. Le Trou de Versailles ou Carrière à Roc de Rance. Bulletin de la Société belge de Géologie, 92, 317‑336.
244Boulvain, F., Bultynck, P., Coen, M., Coen-Aubert, M., Lacroix, D., Laloux, M., Casier, J.G., Dejonghe, L., Dumoulin, V., Ghysel, P., Godefroid, J., Helsen, S., Mouravieff, N.A., Sartenaer, P., Tourneur, F. & Vanguestaine, M., 1999. Les formations du Frasnien de la Belgique. Memoirs of the Geological Survey of Belgium, 44, 1-125.
245Boulvain, F., Coen, M., Coen-Aubert, M., Bultynck, P., Casier, J.G., Dejonghe, L. & Tourneur, F., 1993. Les formations frasniennes du Massif de Philippeville. ServiceGéologique de Belgique, Professional Paper, 1993/1 (259), 1-37.
246Bulvanker, E.Z., 1958. Devonskie chetyrekhluchevye korally okrain Kuznetskogo basseyna. Vsesoyuznyi nauchno-issledovatelskiy Geologicheskiy Institut (VSEGEI), Leningrad, 212 pp. (In Russian).
247Bulvanker, E.Z., 1965. Pervye dannye o devonskikh rugozakh Severo-Vostoka SSSR. In Sokolov, B.S. & Ivanovskiy, A.B. (eds), Rugozy Paleozoya SSSR. Izdatel’stvo Nauka, Moskva, 54-58.
248Chen, M.S., 1959. Nekotorye siluriykie i devonskie stromatoporoidei i korally iz rayona Lushanya vostochnoy chasti provintsii Guychzhou. Acta Palaeontologica Sinica, 7, 285-318. (In Russian).
249Coen, M., 1974. Le Frasnien de la bordure orientale du Bassin de Dinant. Annales de la Société Géologique de Belgique, 97, 67-103.
250Coen, M., 1976. Le Frasnien du lambeau de poussée hercynien de la Tombe (Ardenne belge). Annales de la Société Géologique du Nord, 96, 69-71.
251Coen, M., Coen‑Aubert, M. & Cornet, P., 1977. Distribution et extension stratigraphique des récifs à "Phillipsastrea" dans le Frasnien de l'Ardenne. Annales de la Société Géologique du Nord, 96, 325‑331.
252Coen‑Aubert, M., 1974a. Représentants des genres Phillipsastrea D'Orbigny, A., 1849, Billingsastraea Grabau, A.W., 1917 et Iowaphyllum Stumm, E.C., 1949 du Frasnien du Massif de la Vesdre et de la bordure orientale du Bassin de Dinant. Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 49/8, 1‑38.
253Coen‑Aubert, M., 1974b. Le Givetien et le Frasnien du Massif de la Vesdre. Stratigraphie et paléogéographie. Mémoires in quarto de la Classe des Sciences de l'Académie Royale de Belgique, 2e série, 18/2, 1‑146.
254Coen‑Aubert, M., 1977. Distribution stratigraphique des Rugueux massifs du Givetien et du Frasnien de la Belgique. Annales de la Société Géologique du Nord, 97, 49‑56.
255Coen‑Aubert, M., 1982. Rugueux solitaires du Frasnien de la Belgique. Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 54/6, 1‑65.
256Coen‑Aubert, M., 1987. Nouvelles sous‑espèces de Phillipsastrea hennahi (Lonsdale, W., 1840) dans le Frasnien supérieur de la Belgique. Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 56, 45‑55.
257Coen-Aubert, M., 2012. New species of Frechastraea Scrutton, 1968 at the base of the Late Frasnian in Belgium. Geologica Belgica, 15, 265-272.
258Coen-Aubert, M., 2013. Phillipsastreids and Ptenophyllids (Rugosa) from the Givetian of Mauritania and Northwestern Spain. Geologica Belgica, 16, 164-178.
259Coen-Aubert, M., 2014. Revision of the Frasnian marine deposits from the Booischot borehole (Campine Basin, Belgium). Geologica Belgica, 17, 333-337.
260Coen‑Aubert, M. & Coen, M., 1975. Le Givetien et le Frasnien dans la vallée de la Meuse de Tailfer à Yvoir (bord nord du Bassin de Dinant). Annales de la Société Géologique de Belgique, 97, 499‑524.
261Coen‑Aubert, M. & Lacroix, D., 1979. Le Frasnien dans la partie orientale du bord sud du Synclinorium de Namur. Annales de la Société Géologique de Belgique, 101, 269‑279.
262Coen‑Aubert, M. & Lacroix, D., 1985. Le Frasnien dans la partie orientale du bord nord du Synclinorium de Namur. Bulletin de la Société belge de Géologie, 94, 117‑128.
263Delcambre, B. & Pingot, J.L., 2000. Carte géologique de Wallonie : Fontaine-l’Evêque – Charleroi 46/7-8. 1/25.000. Namur, Ministère de la Région wallonne, Direction générale des ressources naturelles et de l’environnement, avec une notice explicative de 114 p.
264Delcambre, B. & Pingot, J.L., 2014. Carte géologique de Wallonie : Tamines – Fosses-la-Ville 47/5-6. 1/25.000. Namur, Service Public de Wallonie, Direction générale opérationnelle de l'Agriculture, des Ressources naturelles et de l'Environnement, avec une notice explicative de 106 p.
265Delmer, A., 2004. Tectonique du front varisque en Hainaut et dans le Namurois. Memoirs of the Geological Survey of Belgium, 50, 1-61.
266Denayer, J. & Poty, E., 2010. Facies and palaeoecology of the upper member of the Aisemont Formation (Late Frasnian, S. Belgium): an unusual episode within the Late Frasnian Crisis. Geologica Belgica, 13, 197-212.
267Denayer, J., Poty, E., Marion, J.M. & Mottequin, B., 2012. Lower and Middle Famennian (Upper Devonian) rugose corals from southern Belgium and northern France. Geologica Belgica, 15, 273-283.
268De Verneuil, E. & Haime, J., 1850. Polypiers. In De Verneuil, E., Note sur les fossiles dévoniens du district de Sabero (Léon). Bulletin de la Société Géologique de France, 2e série, 7, 161-162.
269D'Orbigny, A., 1849. Note sur des polypiers fossiles. Victor Masson, Paris, 12 pp. (not seen).
270Dumoulin, V., 2001. Carte géologique de Wallonie : Grandrieu-Beaumont 52/5-6. 1/25.000. Namur, Ministère de la Région wallonne, Direction générale des ressources naturelles et de l’environnement, avec une notice explicative de 70 p.
271Ermakova, K.A., 1960. Nekotorye vidy kishechnopolostnykh devona tsentralnykh i vostochnykh oblastey Russkoy Platformy. Trudy Vsesoyuznyi Nauchno-Issledovatelskiy Geologo-Razvedochnyi Neftanoy Institut, 16 (Paleontologicheskiy Sbornik 3), 69-106. (In Russian).
272Ernst, A., Tolokonnikova, Z. & Denayer, J., 2015. Upper Frasnian (Upper Devonian) bryozoans in proximal facies of southern Belgium. Rivista Italiana di Paleontologia e Stratigrafia, 121, 3-20.
273Fedorowski, J., 2003. Typ Coelenterata, Gromada Anthozoa Ehrenberg, 1834, Podgromady: Rugosa Milne Edwards & Haime, 1850-Dividocorallia Fedorowski, 1991. In Malinowska, L. (ed.), Budowa Geologiczna Polski, tom 3, Atlas Skamienialosci przewodnich i charakterystycznych, czesc 1b-z.1, Dewon. Panstwowy Instytut Geologiczny, Warszawa, 49-124 and Atlas.
274Frech, F., 1885. Die Korallenfauna des Oberdevons in Deutschland. Zeitschrift der Deutschen geologischen Gesellschaft, 37, 21-130.
275Goldfuss, A., 1826. Petrefacta Germaniae 1, 1-76. Arnz & Comp., Düsseldorf.
276Gürich, G., 1896. Das Palaeozoicum im Polnischen Mittelgebirge. Verhandlungen der Russisch-kaiserlichen Mineralogischen Gesellschaft zu St. Petersburg, serie 2, 32, 1-539.
277Hill, D., 1981. Part F, Coelenterata, Supplement 1, Rugosa and Tabulata, 2 vols. In Teichert, C. (ed.), Treatise on Invertebrate Paleontology. The Geological Society of America, Inc. and The University of Kansas, Boulder, Colorado and Lawrence, Kansas, 762 pp.
278Hladil, J., Krejci, Z., Kalvoda, J., Ginter, M., Galle, A. & Berouzek,P., 1992. Carbonate ramp environment of Kellwasser time-interval (Lesni Lom, Moravia, Czechoslovakia). Bulletin de la Société belge de Géologie, 100, 57-119.
279Hubmann, B., 1992. Die Korallenfauna aus dem Devon von Feke (Antitaurus, SE-Türkei). II. Rugosa. Geologische und Paläontologische Mitteilungen Innsbruck, 18, 151-169.
280Ivania, V.A., 1952. Verkhnedevonskie korally Rugosa Glubokinskogo izvestnyaka s. Solomino (Kuzbass). Uchenye Zapiski Tomskogo Gosudarstvennogo Universiteta, 18, 125-164. (In Russian).
281Ivania, V.A., 1955. Tetrakorally verkhnego Devona. In Khalfin, L.L. (ed.), Atlas rukovodyashchikh form iskopaemykh fauny i flory zapadnoy Sibiri, t. 1. Gosgeoltekhizdat, Moskva, 219-228. (In Russian).
282Ivania, V.A., 1965. Devonskie korally Sayano-Altayskoy gornoy oblasti. Izd. Tomskogo Universiteta, Tomsk, 398 pp. (In Russian).
283Jiang, S.G., 1982. Anthozoa. In Geological Bureau of Hunan (ed.), The Palaeontological Atlas of Hunan. People's Republic of China, Ministry of Geology and Mineral Resources, Geological Memoirs, series 2, number 1. Geological Publishing House, Beijing, 81-162.
284Kong, L. & Huang, Y.M., 1978. Tetracoralla. In Guizhou Stratigraphy and Palaeontology Work Team (ed.), Palaeontological Atlas of Southwest China. Guizhou Volume. Part I, Cambrian-Devonian. Geological Publishing House, Beijing, 35-161.
285Laloux, M., Dejonghe, L., Geukens, F., Ghysel, P. & Hance, L, 1996. Carte géologique de Wallonie : Limbourg-Eupen 43/5-6. 1/25.000. Namur, Ministère de la Région wallonne, Direction générale des ressources naturelles et de l’environnement, avec une notice explicative de 192 p.
286Lonsdale, W., 1840. In Sedgwick, A. & Murchison, R.I., On the physical structure of Devonshire and on the subdivisions and geological relations of its older stratified deposits. Transactions of the Geological Society of London, series 2, 5, 697 (not seen).
287McLean, R.A., 1994. Frasnian rugose corals of western Canada. Part 3B: The massive Phillipsastreidae- Pachyphyllum, Smithicyathus, Frechastraea. Palaeontographica, Abteilung A, 230, 77-96.
288McLean, R.A., 2010. Frasnian (Upper Devonian) colonial disphyllid corals from Western Canada. NRC Research Press, Ottawa, Ontario, 189 pp.
289Middleton, G.V., 1959. Devonian tetracorals from South Devonshire, England. Journal of Paleontology, 33,138-160.
290Milne-Edwards, H. & Haime, J., 1851. Monographie des Polypiers fossiles des terrains paléozoïques. Archives du Muséum d'Histoire Naturelle, 5, 1-502.
291Milne-Edwards, H. & Haime, J., 1853. A Monograph of the British Fossil Corals. Part 4, Corals from the Devonian Formation. The Palaeontographical Society, London, 211-244.
292Moenke, M., 1954. Rodzaj Hexagonaria w dewonie Gor Swietokrzyskich. Acta Geologica Polonica, 4, 445-483.
293Mottequin, B., 2008a. New observations on Upper Devonian brachiopods from the Namur-Dinant Basin (Belgium). Geodiversitas, 30, 455-537.
294Mottequin, B., 2008b. Late Middle to Late Frasnian Atrypida, Pentamerida and Terebratulida (Brachiopoda) from the Namur-Dinant Basin (Belgium). Geobios, 41, 493-513.
295Mottequin, B. & Poty, E., 2015. Kellwasser horizons, sea-level changes and brachiopod-coral crisis during the late Frasnian in the Namur-Dinant Basin (southern Belgium): a synopsis. Geological Society, London, Special Publications, 423, online, http://doi.org/10.1144/SP423.6
296Muchez, P., Boulvain, F., Dreesen, R. & Hou, H.F., 1996. Sequence stratigraphy of the Frasnian-Famennian transitional strata: a comparison between South China and southern Belgium. Palaeogeography, Palaeoclimatology, Palaeoecology, 123, 289-296.
297Nikolaev, A.A. & Rzonsnitskaya, M.A., 1967. Devonian of Northeastern USSR. In Oswald, D.H. (ed.), International Symposium on the Devonian System, vol. 1. Alberta Society of Petroleum Geologists, Calgary, 483-502.
298Penecke, K.A., 1904. Das Sammelergebnis Dr. Franz Schaffer's aus dem Oberdevon von Hadschin im Antitaurus. Jahrbuch der Kaiserlich-Königlichen Geologischen Reichsanstalt, 53, 141-152.
299Pickett, J., 1967. Untersuchungen zur Familie Phillipsastreidae (Zoantharia rugosa). Senckenbergiana lethaea, 48, 1-89.
300Poty, E., 1982. Paléokarsts et brèches d’effondrement dans le Frasnien moyen des environ de Visé. Leur influence dans la paléogéographie dinantienne. Annales de la Société Géologique de Belgique, 105, 315-337.
301Roemer, C.F., 1852-1854. H.G. Bronn's Lethaea Geognostica, Erster Band, Theil 2, Kohlen-Periode (Silur-, Devon-, Kohlen- und Zechstein Formation). Stuttgart, 788 pp. and Atlas.
302Roemer, C.F., 1883. Lethaea geognostica. I. Theil, Lethaea palaeozoica. Zweite Lieferung. Stuttgart, 324-543 and Atlas.
303Rozkowska, M., 1953. Pachyphyllinae et Phillipsastrea du Frasnien de Pologne. Palaeontologia Polonica, 5, 1-89.
304Sandberg, C.A., Ziegler, W., Dreesen, R. & Butler, J.L., 1992. Conodont biochronology, biofacies, taxonomy and event stratigraphy around Middle Frasnian Lion Mudmound (F2h), Frasnes, Belgium. Courier Forschungsinstitut Senckenberg, 150, 1-87.
305Schlüter, C., 1881. Über einige Anthozoen des Devon. Zeitschrift der Deutschen Geologischen Gesellschaft, 33, 75-108.
306Schröder, S., 2001. Wenig bekannte rugose Korallen aus dem Mittel-und Ober-Devon der Aachener Mulde (Rheinisches Schiefergebirge). Geologica et Palaeontologica, 35, 63-79.
307Schröder, S., 2005. Stratigraphie und Systematik rugoser Korallen aus dem Givetium und Unter-Frasnium des Rheinischen Schiegergebirges (Sauerland/Bergisches Land). Zitteliana, B25, 39-116.
308Scrutton, C.T., 1967. Marisastridae (Rugosa) from south-east Devonshire, England. Palaeontology, 10, 266-279.
309Scrutton, C.T., 1968. Colonial Phillipsastraeidae from the Devonian of South-East Devon, England. Bulletin of the British Museum (Natural History), Geology, 15/5, 181-281.
310Sorauf, J.E., 1967. Massive Devonian Rugosa of Belgium. The University of Kansas Paleontological Contributions, 16, 1-41.
311Sorauf, J.E. & Pedder, A.E.H., 1986. Late Devonian rugose corals and the Frasnian-Famennian crisis. Canadian Journal of Earth Sciences, 23, 1265-1287.
312Soshkina, E.D., 1951. Pozdnedevonskie korally Rugosa, ikh sistematika i evolyutsiya. Trudy Paleontologicheskogo Instituta, 34, 1-122. (In Russian).
313Soshkina, E.D., 1952. Opredelitel devonskikh chetyrekhluchevykh korallov. Trudy Paleontologicheskogo Instituta, 39, 1-127. (in Russian).
314Tsien, H.H., 1968. Disphyllidae Hill, 1939 ou Phillipsastraeidae Roemer, 1883. Annales de la Société Géologique de Belgique, 91, 595-615.
315Tsien, H.H., 1971. The Middle and Upper Devonian reef-complexes of Belgium. Petroleum Geology of Taiwan, 8, 119-173.
316Tsien, H.H., 1974. Excursion J. In Bouckaert, J. & Streel, M. (eds), Guidebook of the International Symposium on Belgian micropaleontological limits from Emsian to Visean, Namur September 1st to 10th 1974. Service Géologique de Belgique, Bruxelles, 34 pp.
317Tsien, H.H., 1975. Introduction to the Devonian Reef development in Belgium. Livret-Guide, Excursion C (Nord de la France et de la Belgique), 2e Symposium International sur les Coraux et Récifs coralliens fossiles, Paris 1975. Bruxelles, pp. 3-43.
318Tsien, H.H., 1977. The sequence and distribution of Frasnian rugose corals fauna in Belgium. Mémoires du B.R.G.M., 89, 203-220.
319Tsien, H.H., 1978. Rugosa massifs du Dévonien de la Belgique. Mémoires de l'Institut Géologique de l'Université de Louvain, 29, 197-229.
320Yoh, S.S., 1937. Die Korallenfauna des Mitteldevons aus der Provinz Kwangsi, Südchina. Palaeontographica, Abteilung A, 87, 45-76.
321Zheltonogova , V.A. & Ivania, V.A., 1960. Podklass Tetracoralla (Rugosa). Tetracorally. In Khalfin, L.L. (ed.), Biostratigrafiya paleozoya Sayano-Altayskoy gornoy oblasti. Trudy Sibirskogo Nauchno-issledovatel'skogo Instituta Geologii, Geofiziki i Mineral'nogo Syr'ya, 20, 368-408. (In Russian).
322Plate 1. A-H: Frechastraea pentagona (Goldfuss, 1826). A-B: IRScNB a740, Senzeille MC-1974-14-P36 at Neuville; transverse and longitudinal sections. C: IRScNB a12968, Verviers MC-1974-7-2 at Pepinster; transverse section. D-E: IRScNB a12969, Froidchapelle MC-75-D107 at Cerfontaine; transverse and longitudinal sections. F: IRScNB a12970, Froidchapelle MC-71-C937 at Cerfontaine; transverse section. G-H: IRScNB a12971, Senzeille MC-1974-15-T887 at Senzeille; transverse and longitudinal sections. Magnification x 4.
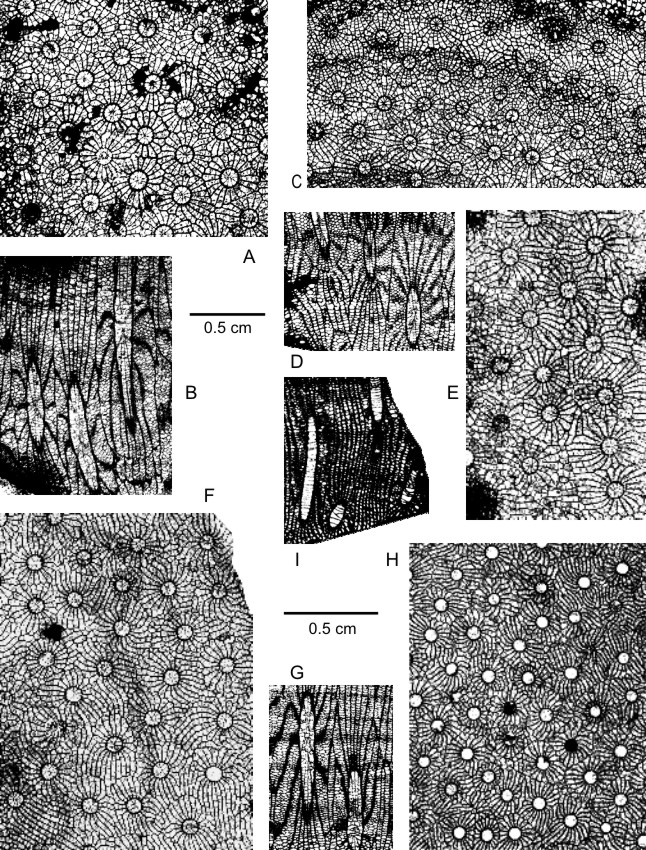
323Plate 2. A-G: Frechastraea minima (Rozkowska, 1953). A-B: IRScNB a12972, Senzeille MC-1974-15-T881 at Senzeille; transverse and longitudinal sections. C-D: IRScNB a12973, Froidchapelle MC-75-D115 at Cerfontaine; transverse and longitudinal sections. E: IRScNB a12974, SenzeilleMC-1974-15-T892 at Senzeille; transverse section. F-G: IRScNB a12975, Senzeille MC-1974-14-P55 at Neuville; transverse and longitudinal sections. H-I: Frechastraea kaisini (Tsien, 1978). IRScNB a12978, Froidchapelle MC-75-D118 at Cerfontaine; transverse and longitudinal sections. Magnification x 4 for figures A-D and x 5 for figures E-I.
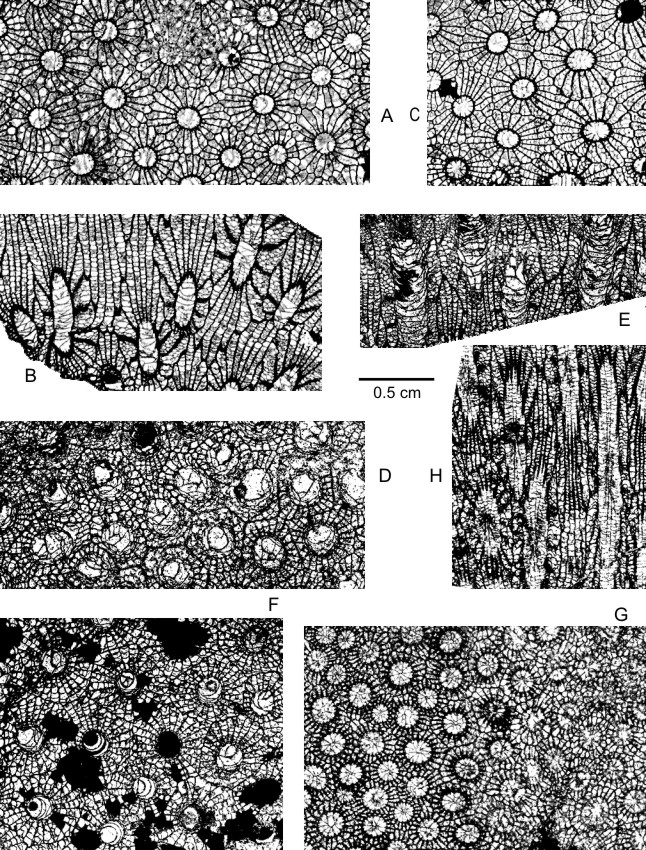
324Plate 3. A-C: Frechastraea micrommata (Roemer, 1852). A-B: IRScNB a12976, Froidchapelle MC-75-C993A at Cerfontaine; transverse and longitudinal sections. C: IRScNB a12977, Froidchapelle MC-71-C950 at Cerfontaine; transverse section. D-F: Frechastraea cf. tungkanlingensis (Yoh, 1937). D-E: IRScNB a12979, Braives MC-1979-2-W51 at Huccorgne; transverse section. F: IRScNB a12980, Esneux MC-1974-89-F80 at Hony; transverse section. G-H: Frechastraea crassiseptata (Tsien, 1978). IRScNB a12981, Surice MC-1974-133-V27 at Vodelée; transverse and longitudinal sections. Magnification x 4.
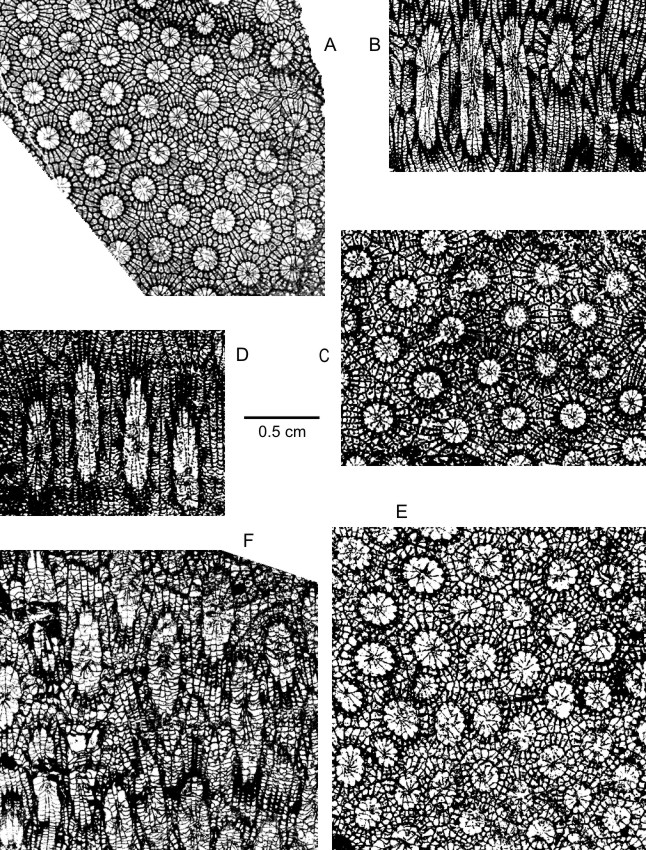
325Plate 4. A-E: Frechastraea crassiseptata (Tsien, 1978). A-B: IRScNB a12982, Hamoir MC-120-D851 at Bomal; transverse and longitudinal sections. C-D: IRSNB a12983, Hamoir MC-120-D825 at Bomal; transverse and longitudinal sections. E-F: IRScNB a12984, Senzeille MC-1984-1-A514 at Neuville; transverse and longitudinal sections. Magnification x 4.
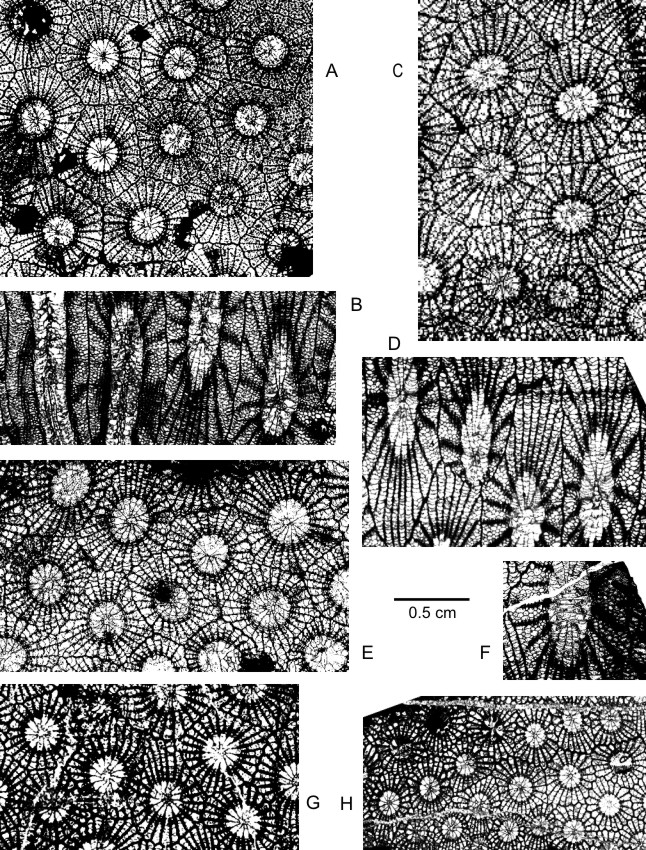
326Plate 5. A-G: Frechastraea limitata (Milne-Edwards & Haime, 1851). A-B: IRScNB a12986, Hamoir MC-120-D850 at Bomal; transverse and longitudinal sections. C-D: IRScNB a12987, Louveigné MC-135-183bis at Aywaille; transverse and longitudinal sections. E-F: IRScNB a749, Limbourg MC-1974-8-A17 at Les Surdents; transverse and longitudinal sections. G: IRScNB a12988, Verviers MC-1988-4-A677 at Lambermont; transverse section. H : Frechastraea crassiseptata (Tsien, 1978). IRScNB a12985, Verviers MC-1988-4-A679 at Lambermont; transverse section. Magnification x 4.
327Manuscript received 11.05.2015, accepted in revised form 26.08.2015, available on line 18.10.2015.
