


- Portada
- volume 18 (2015)
- number 2-4
- The Zemst borehole, first record of the EECO in the North Sea Basin and implications for Belgian Ypresian - Lutetian stratigraphy


Vista(s): 3002 (31 ULiège)
Descargar(s): 473 (10 ULiège)

The Zemst borehole, first record of the EECO in the North Sea Basin and implications for Belgian Ypresian - Lutetian stratigraphy

Abstract
Integration of new data in sedimentology, micropalaeontology and organic carbon isotopeanalysis of upper Ypresian strata in central Belgium (Zemst hole) enables differentiationof a series of biotic events and carbon isotope trends, which are believed to be associated with the 1.5-million-year-long period of global warming, known as the Early Eocene Climatic Optimum (EECO). The relatively low values in δ13Corg(-26.5‰ to -27.1‰) in the interval from the Panisel Sand to the Merelbeke Clay Members (upper NP12-lower NP13) are shown to be coinciding with a fairly high frequency in Apectodinium (>3%, up to 14%) and a Discoaster-bloom (16%-50%, essentially D. kuepperi). This is quite analogous, although less prominent, to what has been observed during the Paleocene-Eocene Thermal Maximum (PETM: Apectodinium abundance of 75% at middle and high latitudes and Discoaster blooms in tropical areas). The abrupt positive shift of up to 1‰ in δ13Corg values at the base of the overlying Pittem Clay Member (mid-NP13, mid-chron C22r), which is coincident with the virtual disappearances of Apectodinium (<0.1%) and Discoaster (<0.5%) seems to mark the end of the EECO in the southern North Sea Basin. The Zemst data allow the identification of the NP12/NP13 boundary, virtually coinciding with chron C23n/C22r boundary, at the depositional break between the Panisel Sand Member and the overlying Kwatrecht Member. The new data also allow to substantially refine the calcareous nannofossil stratigraphy during Biochron NP15 (mid-Lutetian) at middle latitudes. This is corroborated by additional data from Belgium, which furthermore reveal that the primary criterion for identifying the base of the Lutetian (LO of Blackites inflatus), as adopted in the Gorrondatxe GSSP (Spain), cannot be applied in the North Sea Basin s.s. (excluding the Paris Basin) and that there is an urgent need for defining appropriate substitutes for this boundary at these latitudes.
Tabla de contenidos
1. Introduction
1Several cored boreholes have recently been drilled in strategic areas in order to generate and update the geological maps of Belgium. Most of these wells were designed to elucidate regional stratigraphic problems.
2The Zemst borehole (BGD 73E359; x = 155.444, y = 187.591; GPS-coordinates 50°59’54.52”N, 4°26’46.50”E; Fig. 1) was executed in 2001 to evaluate the presence of the P1n clay in the area between Aalst and Mechelen. This P1n clay, which figures on the old geological maps of Belgium (Anonymous, 1893; Fig. 2), was introduced by Rutot (1890) as part of the Paniselian Stage (now upper part of Ypresian Stage; Steurbaut, 2006a,b) and termed P1(d) or the grey plastic clay (“argile grise plastique”), positioned in between the sandstone bearing P1d sands (“lower Paniselian”) and the overlying Aalter Sands (P2 or “upper Paniselian”). It clearly postdates the P1m clay (Anonymous, 1893), before known as the basal grey plastic clay (“argile grise plastique schistoïde de base”) or P1(a) (Rutot, 1890; Fig. 2), and today as Merelbeke Clay.
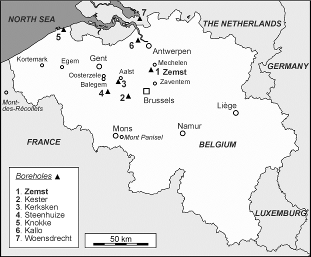
3Figure 1. Location of the Zemst borehole and additional borehole and outcrop sections mentioned in the text.
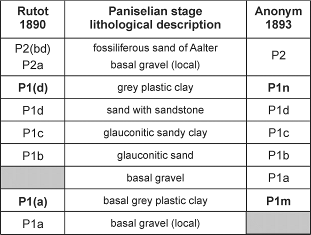
4Figure 2. Subdivision of the obsolete Paniselian Stage, used until the mid-1950 (e.g. Gulinck & Hacquaert, 1954) (grey shading = not identified).
5The present study aims at elucidating the stratigraphy of the upper Ypresian and the lower Lutetian in the Zemst area (Fig. 3), with special attention to the Ypresian-Lutetian transition. It essentially focuses on the regional lithology and sedimentology, the changes in calcareous nannofossil, microfossil and dinoflagellate cyst assemblages, the geochemistry (i.e. the organic carbon isotopes) and the general depositional conditions. It seeks at integrating these data into a sequence stratigraphic framework.
6A detailed palaeoenvironmental study of the upper Ypresian sediments of Belgium, including the Zemst borehole, is in progress. The palynofacies results of Zemst are summarised in Table 2, in order to support the interpretation of the organic carbon isotope data (chapter 4.2).
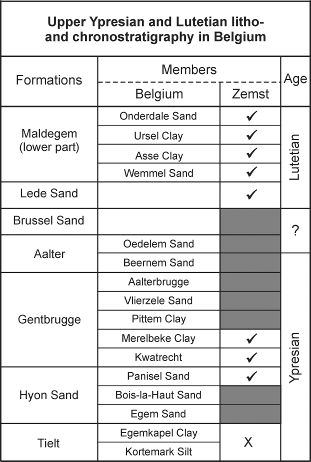
7Figure 3. Overview of the upper Ypresian and Lutetian lithostratigraphy of Belgium (after Steurbaut, 2006b), and its application in the Zemst borehole ( = present; grey-shaded = missing; X = Tielt Fm present, but member remains unclear).
2. Material and methods
8The Zemst borehole was logged by Steurbaut & King in August 2001 (Fig. 4). Grainsize analyses have been carried out on 53 samples in the lower half of the borehole (27.80 m - 54.75 m), at approximately 0.5 m spacing (Matthijs & Buffel, 2001, report) (Fig. 5). Gamma-ray measurements were performed by TNO - Delft (Fig. 5). Calcareous nannofossils and calcareous microfossils (essentially foraminifera and ostracods) have been studied at high resolution (at approximately 0.8 m spacing, increasing to 0.3 m in the upper Wemmel Sand Member), following the preparation and investigation procedures explained in Steurbaut & King (1994) and in Steurbaut (2011). Abbreviations used in the present paper: LO = lowest occurrence, LCO = lowest consistent occurrence (practically permanent presence with frequency >1% of total assemblage), HO = highest occurrence (the abbreviations FO and FCO, used by Agnini et al., 2006 refer to first occurrence and first common occurrence respectively). P is the percentage of planktonic foraminifera in the total foraminiferal population (see King, 1989). Nannofossil taxonomy is essentially from Perch-Nielsen (1985), taking into account subsequent modifications by Young & Bown (1997) and Aubry & Bord (2009). The nannofossil samples, residues and slides are stored in the collections of the RBINS (Brussels, Belgium). Six samples have been examined for dinoflagellate cysts and other organic-walled microfossils (positions shown on Fig. 4), following processing techniques described in Heilmann-Clausen (1985). These dinoflagellate cyst samples, residues and slides are kept (temporarily) at the Exxon Mobil Upstream Research Company, Texas (USA).
9Carbon isotope analyses were performed on 15 samples. Organic matter of the sediments has been isolated, following the procedure described in Storme et al. (2014). The δ13C results were normalized to theinternational PDB standard (VPDB, Vienna Peedee Belemnite).
3. Stratigraphic interpretation
3.1. Tielt Formation: 55 m - 54.65 m
10No calcareous fossils have been retrieved from the clayey silts at the base of the borehole. The borehole was not deep enough to allow a reliable gamma ray record (Fig. 5). The organic-walled microfossil assemblage from this interval (Z1 at 54.80 m, Fig. 4), essentially dinoflagellate cysts, is dominated by the Spiniferites spp. - Achomosphaera spp. group (~24%) and Homotryblium spp. (~13%). It is marked by high proportions of Lingulodinium machaerophorum (~6%), Charlesdowniea coleothrypta (~5%), Eatonicysta aff. ursulae (~5%) and Areoligera senonensis (~4%). Such a combination of taxa indicates marine conditions on the deeper parts of the inner shelf, possibly marked by abnormal (low or high) salinity conditions (Dybkjær, 2004; Sluijs et al., 2005).
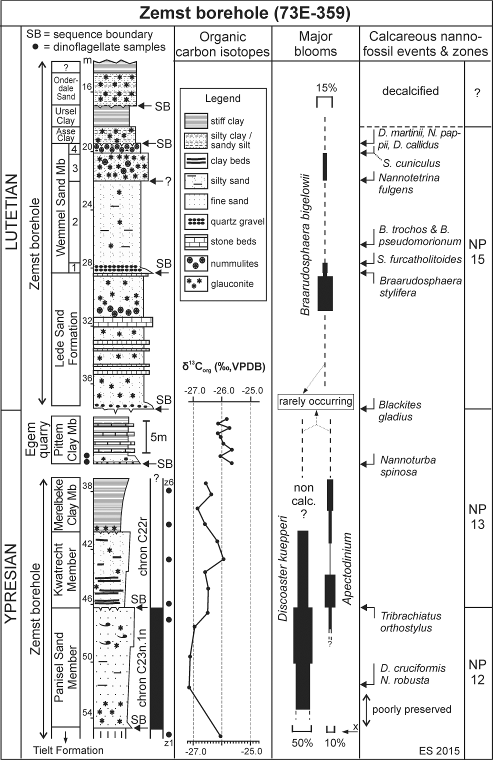
11Figure 4. Lithostratigraphic interpretation, organic carbon isotope evolution and calcareous nannofossil trends, events and zones in the Zemst borehole section and in the Pittem Clay Member at Egem, with position of the studied organic-walled microfossil samples. The magnetostratigraphic interpretation is a compilation of data from Belgium and Denmark, not from Zemst (the magnetostratigraphic position of the lowermost unit - Tielt Fm - remains unclear) (x = rare occurrence of Apectodinium).
12The assemblage is furthermore characterised by rare Cerebrocysta bartonensis, Cleistosphaeridium polypetellum, Homotryblium deconinckii, Ochetodinium romanum, Pentadinium laticinctum and Turbiosphaera galatea,and by the absence of the Charlesdowniea columna-group and Glaphyrocysta? spineta. Apectodinium is extremely rare (only a few specimens of A. homomorphum encountered). Because of the presence of Charlesdowniea coleothrypta and the absence of Areosphaeridium diktyoplokum it is attributable to the D8 dinoflagellate zone of Costa & Manum (1988), which is rather short-ranging within the middle of the Ypresian (52.1 Ma - 50.5 Ma, according to Köthe, 2012). Similar assemblages with few Apectodinium (1% or less), abundant Homotryblium spp. (over 10%) and Areoligera senonensis (over 5%) have been identified in the Steenhuize-Wijnhuize borehole (Vanhove & De Coninck, 1992: sample ST2 at 8.5 m depth) and at Kester (Steurbaut, unpubl. information). They were recorded in a clayey silt unit, which is part of the Tielt Formation. These similarities allow to conclude that this silt unit is also present at Zemst.
3.2. Panisel Sand Member (upper Hyon Sand Formation): 54.65 m - 46.25 m
3.2.1. Lithology
13This unit is separated from the underlying Tielt Formation by a deeply Thalassinoides-burrowed omission surface (contact Cz1, Table 1). It shows a coarsening-upward trend (Fig. 5), grading from glauconite-rich sandy silt, with coarser base and small siliceous concretions, to silty fine sand with sparsely dispersed molluscs and, further upward (49.0 m - 46.25 m), to silty fine to medium-grained sand with dispersed shell debris. It shows a progressive decrease in gamma ray count rate (from 70 to 40 cps) (Fig. 5). The lithofacies of this unit is comparable (glauconitic clayey to silty sand with siliceous concretions) to that of the Panisel Sand Member in its type locality (Geets, 1992), although presents a much higher silt fraction in its lower two thirds.
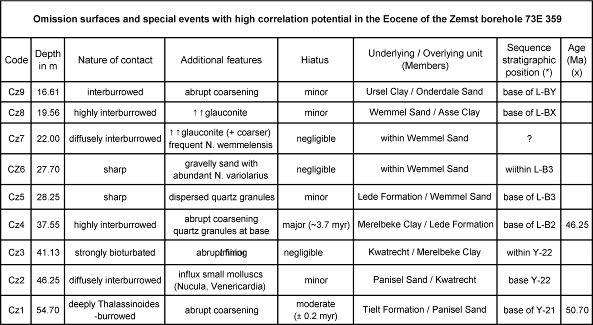
14Table 1. Characterization of the major breaks in sedimentation and associated events in the Zemst borehole section. (*) : the naming of the Lutetian sequences is tentative; (x) : the ages are according to the 2012 GTS.
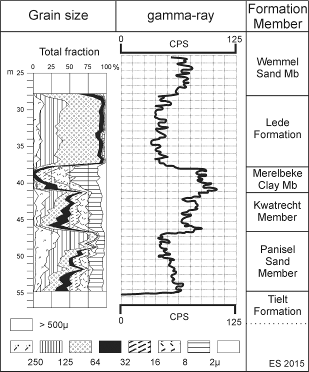
15Figure 5. Grain-size variation (Matthijs & Buffel, 2001) and gamma-ray signature of the Ypresian deposits in the Zemst borehole.
3.2.2. Calcareous microfauna and macrofauna
16The lowest part (up to ca. 54 m) is decalcified. Above this, benthic foraminifera are abundant, but of low diversity. The assemblage is dominated by Cibicidoides proprius and associated with Asterigerina bartoniana, Cancris subconicus, Elphidium laeve, Cribroelphidium hiltermanni, Nonion commune and polymorphinids. Planktonic foraminifera (Muricoglobigerina sp.) occur rarely but consistently, with a maximum abundance (P ca. 2%) in the lowest calcareous sample (53.5 m). Ostracods are relatively rare, including Cytheretta decipiens, Pterygocythereis cornuta and Leguminocythereis bicostata. Small molluscs are rather sparsely dispersed. Anomia cf. primaeva occurs consistently throughout. Callista sp., Lutetia sp. and Venericardia sp. are common at 46.5 m. Similar benthic foraminiferal assemblages are recorded from the middle and upper Ypresian poorly sorted glauconitic sands and sandy clays in western Belgium (Kaasschieter, 1961; King, 1990). Most of the ostracods have a similar distribution. L. bicostata has not previously been recorded in Belgium. It was described from the middle part of the London Clay Formation in England (Keen, 1978; King, 1991). The microfaunal and macrofaunal assemblages indicate an inner neritic environment.
3.2.3. Calcareous nannofossils
17Nannofossils are consistently present, except in the basal part (0.6 m), which is decalcified. They are only present in low numbers (generally <10 specimens/field of view at x1000) and are mostly poorly preserved. Species diversity is moderate (between 15 and 20 taxa), with a maximum of 30 in the uppermost sample (46.50 m). The assemblages are dominated by Discoaster kuepperi (between 38% and 50%). They also contain high numbers of small Prinsiaceae (10%-28%) and Pontosphaera pulchra (up to 10%), and in the uppermost sample (46.50 m)also common Micrantholithus spp. (13%). The composition of the assemblages points to inner neritic conditions, becoming coastal towards the top (abundance of Micrantholithus spp.). Small to medium-sized (5 to 8 µm) subcircular densely packed diamond-shaped calcitic bodies of unknown relationship (tentatively named “inc. sed. sp.”) are also frequently occurring (up to 5%). The assemblages are furthermore characterised by the co-occurrence of Tribrachiatus orthostylus, Discoaster lodoensis, Chiphragmalithus armatus, Discoaster cruciformis and Nannoturba robusta, indicating zone VIIIb of Steurbaut (1998), which corresponds to the topmost part of zone NP12 of Martini (1971). Similar assemblages have been recorded in the Panisel Sand Member in the Kester and the Kerksken boreholes (Steurbaut, unpubl. information).
3.2.4. Organic-walled microfossils
18Sample Z2 at the top of the unit (46.87 m) is also dominated by the Spiniferites spp. - Achomosphaera spp. group (~22%). Compared to Z1 it is much richer in Cleistosphaeridium diversispinosum (~9%) and Apectodinium homomorphum (3.4%), but poorer in Homotryblium spp. (~5%) and Areoligera senonensis (~2%). The increase in Apectodinium may indicate an increase in sea surface temperatures, the high numbers in C. diversispinosum a more open, less restricted sedimentation regime (Sluijs et al., 2005). The assemblage is marked by a series of first records, among which are these of Areosphaeridium diktyoplokum (very rare) and Impletosphaeridium kroemmelbeinii (rare), the presence of Eatonicysta ursulae and by the absence of Impletosphaeridium cracens and Phthanopteridinium comatum,supporting the presence of the Panisel Sand Member at Zemst (Steurbaut, Heilmann-Clausen & Van Simaeys, in prep.). On the basis of this dinoflagellate cyst combination it has to be classified into the D9a dinoflagellate zone of Köthe (2012), which is fairly long-ranging in the late Ypresian (50.5 Ma - 48.0 Ma, according to Köthe, 2012).
3.2.5. Organic carbon isotopes
19The δ13Corg values, which are quite negative (-27.1‰), remain very constant throughout most of the unit. There is a 0.5‰ return to more positive values in the topmost meter.
3.3. Kwatrecht Member (lower Gentbrugge Formation): 46.25 m - 41.13 m
3.3.1. Lithology
20This unit is separated from the underlying and overlying units by interburrowed omission surfaces (Fig. 4). It is marked by a coarsening-upward trend (Fig. 5), grading from glauconite-rich sandy silt, with frequent small molluscs, to a diffuse alternation of thin sandy silt and clay layers and lenses, disturbed by bioturbation, and further upward to bioturbated glauconitic silty sands. The gamma ray record is marked by small fluctuations, although remains fairly constant (between 70 and 80 cps). The lithofacies of this unit is comparable (especially the alternation of thin silty sand layers and clay layers) to that of the Kwatrecht Member in its type area Melle-Kwatrecht (De Moor & Geets, 1974) and in the Kerksken borehole (Steurbaut, unpubl. information).
3.3.2. Calcareous microfauna and macrofauna
21The upper part (above ca. 43 m) is decalcified. Benthic foraminifera are abundant, dominantly Anomalinoides nobilis, Cibicidoides proprius, Cibicidina mauricensis, C. tendami, Cribroelphidium hiltermanni, Spiroplectammina deperdita and polymorphinids. Rare planktonic foraminifera (Muricoglobigerina sp.) also occur (P<1%). Ostracods are uncommon, but there is a relatively diverse assemblage, including Cyamocytheridea mourloni, Eucytherura hyonensis, L. bicostata and P. cornuta. Spinarcella, a microfossil taxon of unknown systematic position (Bignot, 1989),is also present. Small molluscs are frequent, but partially decalcified, including Nucula sp., Lutetia sp., Venericardia sp., and Trigonodesma lissa. The bryozoan Lunulites sp. and the solitary corals Turbinolia sp. and Sphenotrochus sp. also occur.Similar benthic foraminiferal and ostracod assemblages are recorded from the mid and upper Ypresian poorly sorted glauconitic sands and sandy clays in western Belgium (Kaasschieter, 1961; King, 1990). The microfaunal and macrofaunal assemblages indicate an inner neritic environment.
3.3.3. Calcareous nannofossils
22Nannofossils are consistently present below 43.5 m, although in low numbers (generally ~5 specimens/field of view at x1000) and are poorly preserved. Species diversity is moderate (between 20 and 25 taxa). The assemblages are dominated by Discoaster kuepperi (29%-27%) and small Prinsiaceae (28%-14%), and in the lowermost sample (46.10 m) also by Micrantholithus spp. (20%). Inc. sed. sp. (see above) is also common (around 10%). The assemblages indicate a clear palaeoenvironmental shift from coastal (common Micrantholithus spp. at the base) to inner neritic conditions. They also present a high diversity in Discoaster taxa (up to 7 taxa) and are furthermore characterised by the co-occurrence of Discoaster lodoensis, Discoaster cruciformis, Imperiaster obscurus s.s. (2x3 long-rayed form) and Nannoturba robusta, and by the absence of Tribrachiatus orthostylus. This indicates zone IX of Steurbaut (unpubl. information) which corresponds to the basal part of zone NP13 of Martini (1971). Similar assemblages have been identified in the Kwatrecht Member in the Kerksken borehole (Steurbaut, unpubl. information).
3.3.4. Organic-walled microfossils
23Samples from the base (Z3 at 46.15 m) and the top of the unit (Z4 at 42.40 m) present similar dinoflagellate cyst assemblages, in terms of species diversity (Z4 somewhat less diverse) and occurrences, although differ in their relative proportions. Z3 is dominated by the Spiniferites spp. - Achomosphaera spp. group (~25%), Apectodinium spp. (14%), Homotryblium spp. (11%) and Cleistosphaeridium diversispinosum (~11%), whereas Z4 is dominated by C. diversispinosum (33%) and Homotryblium spp. (19%). The Spiniferites spp. - Achomosphaera spp. group is less abundant in Z4 (11%), while Apectodinium spp. reach minimum values (<1%), suggesting substantial palaeoenvironmental changes during the deposition of the unit. Areosphaeridium diktyoplokum is relatively frequent (up to 3%) in both assemblages, while E. ursulae is also persisting, indicating dinoflagellate zone D9a of Köthe (2012). The highest occurrence (HO) of Areoligera senonensis is in Z3. Impletosphaeridium cracens first occurs in Z4.Identical dinoflagellate cyst assemblages dominated by the Spiniferites spp. - Achomosphaera spp. group (14%) and Homotryblium spp. (~10%), and marked by frequent A. diktyoplokum (between 2% and 3%), the progressive disappearance of A. senonensis and the rare first records of I. cracens have been recorded in the Kwatrecht Member in the Melle-Kwatrecht area (Van Simaeys, unpubl. information).
3.3.5. Organic carbon isotopes
24The δ13Corg values are fluctuating between -26.6‰ and -26.0‰.
3.4. Merelbeke Clay Member (middle Gentbrugge Formation): 41.13 m - 37.55 m
3.4.1. Lithology
25This unit is separated from the underlying and overlying units by interburrowed omission surfaces (Fig. 4). It consists of clayey fine silt to silty clay (75%<16 µm; 15%<2 µm), glauconitic at the base, with an increased number of coarse silt partings and thin coarse silt lenses in the uppermost meter. Dispersed pyrite is present throughout. The highest gamma ray values have been observed in this unit (Fig. 5). They present a decreasing trend (between 100 and 80 cps), with a maximum of 106 cps in the lowermost 0.5 m. This fine-grained lithofacies-type has been associated with the Merelbeke Clay Member, although its clay content (fraction <2 µm) is much less than in the type area (De Moor & Geets, 1974).
3.4.2. Calcareous microfauna and macrofauna
26Samples from this unit contain pyritic moulds of formerly calcareous fossils. These comprise small benthic molluscs, planktonic molluscs (pteropods) and rare benthic foraminifera. The benthic molluscs are mostly specifically indeterminate, but include the bivalve Varicorbula globosa and the gastropods Adeorbis sp., Euspira sp., ? Ringicula sp. and a rissoid. The pteropods comprise Camptoceratops priscum, and two undescribed species of Limacina, a relatively globose and low-spired form (Limacina aff. taylori) and a higher-spired and more elongated form (Limacina aff. tutelina). C. priscumis a very short-ranging taxon, widespread in the southern North Sea Basin. It has also been recorded in the Aquitaine Basin and the USA Gulf Coast (Cahuzac & Janssen 2010), and in Uzbekistan (Janssen, King & Steurbaut, 2011) and Khazakstan (King et al., 2013), always within upper Zone NP12 and lower Zone NP13.
27A very similar faunal assemblage, with the same taxa of pteropods, has been recorded previously from the Merelbeke Clay in the Knokke and Kallo boreholes (King, 1990, with Limacina aff. taylori recorded as Spiratella sp. nov. A). The Merelbeke Clay is probably within the youngest part of pteropod zone 9. Pteropods are more common in this clay unit than at any other level in the Eocene of Belgium. These planktonic molluscs are characteristic of environments open to oceanic water circulation, but do not necessary indicate great water depths. It suggests a mid to outer neritic environment.
3.4.3. Calcareous nannofossils
28This unit contains no calcareous nannofossils because of decalcification, which probably took place during early diagenesis.
3.4.4. Organic-walled microfossils
29The dinoflagellate cyst assemblages from the lower part (Z5 at 40.15 m) and the top of the unit (Z6 at 39.90 m) are dominated by the same taxa, but show substantial differences in species diversity (52 taxa in Z5 versus 35 taxa in Z6). Both contain very high numbers of the freshwater green algal taxon Pediastrum sp. and the marginal marine dinoflagellate cyst taxon Homotryblium spp. (see Dybkjær, 2004; Sluijs et al., 2005 for palaeoenvironmental interpretations), but with different proportions (Pediastrum: 30% in Z5 and 53% in Z6; Homotryblium: 28% in Z5 and 21% in Z6; note that these are percentages of the total organic-walled microfossil assemblage and not of the total palynomorph assemblage, as shown in Table 2). This witnesses the transgressive character of the unit, through which lakes or marshes, were flooded and eroded updip, allowing their freshwater remnants to enter the marine deposition system. The highest occurrence of common Apectodinium spp. (essentially A. homomorphum) is recorded at the top of this unit (7.5%). The assemblages are furthermore marked by the presence of Impletosphaeridium cracens and the absence of Phthanopteridinium comatum. Similar assemblages are known from the type-locality of the Merelbeke Clay Member (Van Simaeys, unpubl. information) and in the interval 239 m - 237 m in the Kallo borehole, except for Pediastrum spp. which is less frequent (3%) in the latter (De Coninck, 1976).
3.4.5. Organic carbon isotope signature
30The δ13Corg values present a zigzag pattern, the endpoints of which are -27.0‰ in the middle of the unit and -26.4‰ slightly higher up.
3.5. Lede Sand Formation: 37.55 m - 28.45 m
3.5.1. Lithology
31This unit is also separated from the underlying and overlying units by interburrowed omission surfaces (Fig. 4, Table 1). It consists of well-sorted glauconitic silty very fine sand, which upward include six generally thin sandstone levels (one thicker of about 55 cm in the upper half) and locally contain mollusc moulds, oyster fragments, Nummulites variolarius (abundance peak between 31.5 m and 29 m) and plant debris. Frequent millimetric quartz granules are recorded at the base of the unit. The gamma ray values are quite low and rather constant, fluctuating between 37 and 50 cps (Fig. 5). This lithofacies of homogeneous fine sand with sandstone levels is very similar of that of the Lede Sand Formation in its type-locality at Balegem and in the area from Balegem through Lede to the north of Brussels (Fobe, 1986).
3.5.2. Calcareous microfauna and macrofauna
32Samples from the Lede Formation contain abundant and moderately diverse benthic foraminifera and ostracods, except in the basal part which is decalcified. These assemblages have been well-documented previously (Keij, 1957; Kaasschieter, 1961) and are not analysed here in detail. Nummulites variolarius is common to abundant throughout.
33In the lower part of the Lede Formation (samples between 37.0 m and 31.5 m) the foraminifera Asterigerina, Lobatula, Pararotalia and polymorphinids are common; miliolids occur in low numbers. A diverse ostracod assemblage is dominated by Schizocythere. In its highest part, in samples at 30.5 m to 29.5 m, there is a major increase in the proportion of miliolids, which comprise over 50% of the benthic foraminiferal population (excluding Nummulites). Nine genera of Miliolidae are represented. This influx is associated with the occurrence of the “larger foraminifera” Fabularia bella, Alveolina sp. and Orbitolites complanutus. Alveolina occurs only as rare abraded fragments; Orbitolites is uncommon, but well preserved; Fabularia is common.
34Planktonic foraminifera occur consistently but very rarely throughout the Lede Sand Formation, with P<2%. Pseudohastigerina is dominant. Hooyberghs (1984a) records 13 species of planktonic foraminifera from this unit at Balegem, but their relative abundance is not recorded. Here also, the dominance of Pseudohastigerina is noted. As noticed by King (1989) this indicates assignment to planktonic zone NSP7, characterised by the dominance of Pseudohastigerina. In Belgium, this zone is represented within the interval from the Aalter Sand Formation to the top of the Ursel Member (NP14–NP16) (King, 1989).
35Other faunal components include sporadic bryozoa (Lunulites) and occasional calcitic molluscs (ostreids and pectinids). The aragonitic molluscs have probably been destroyed by post-depositional dissolution, as is typical for the Lede Formation.
36The microfaunal assemblage indicates an inner neritic environment with low sedimentation rates. The high proportion of epifaunal foraminifera, including cibicidids, indicates extensive algal (seaweed) or seagrass (Thalassia) cover; the fine plant debris dispersed in some intervals may be seagrass remnants. The miliolid-rich foraminiferal population (e.g. Quinqueloculina carinata) in the upper Lede Formation indicates a very shallow, warm environment with somewhat lowered salinity (Murray et al., 1981, p. 244).
3.5.3. Calcareous nannofossils
37The associations are poorly preserved and strongly overgrown in the lowermost 3 m of the Lede Formation. Intact specimens are rare (2 to 4 per field of view at 1000x magnification). Species diversity is low (between 10 and 15 identifiable taxa). The assemblages are dominated by small and medium-sized Prinsiaceae (~55%), Coccolithus pelagicus (~16%)and Zyghrablithus bijugatus (~12%), keeping in mind that their composition is biased in favour of the most solid forms. Higher up preservation is progressively improving and from ~30.50 m upward the associations are fairly well preserved, with a normal species diversity (maximum between 30 to 35 taxa and ~10 specimens/field of view at 29.50 m). In this upper part of the Lede Formation the assemblages are dominated by small Prinsiaceae and by Braarudosphaeraceae, with almost equal proportions (~27%), and to a lesser degree by Lanternithus minutus (~15%) and Zyghrablithus bijugatus (~15%). Braarudosphaera bigelowii is the most common among the Braarudosphaeraceae (~15%). This change in nannofossil composition indicates a shallowing of the depositional system from inner neritic to almost coastal with probably hyposaline high-energy conditions (Giunta et al., 2003). Braarudosphaeraceae are again decreasing at the top of the unit (28.5 m) (11%, with B. bigelowii up to 4%), alongside with decreasing species diversity (20 to 25 taxa) and general abundance (5 to 10 specimens/field of view).
38The assemblages of the Lede Formation are marked by the co-occurrence of Blackites gladius, Braarudosphaera stylifera and an undescribed species of Nannoturba. This, in association with the absence of Discoaster sublodoensis, Blackites inflatus and Nannotetrina fulgens, indicates the lower part of nannofossil zone NP15, although not the very base. Identical associations have been recorded in the Lede Formation at its type-locality Balegem (Steurbaut, unpubl. information), Oosterzele (Smith et al., 2004), Mont-des-Récollets (Vandenberghe et al., 1998 and Steurbaut, unpubl. information) and in unit ZL2 of the Lede Formation at Zaventem (Herman et al., 2000) (Fig. 1).
3.6. Wemmel Sand Member (lower Maldegem Formation): 28.45 m - 19.56 m
3.6.1. Lithology
39Four subunits can be identified (Fig. 4). The lower, tentatively named Wemmel Sand W1 (28.45 m - ca. 27.68 m) consists of silty glauconitic very fine sands, rather poorly sorted, with thin layers of quartz granules at several levels (highest level at 27.72 m - 27.68 m) and frequent specimens of N. variolarius and of a flattened Nummulites sp. of the prestwichianus group. The base of this unit and, thus, of the Wemmel Sand Member, is marked by dispersed small quartz granules and concentrations of Nummulites variolarius, Ditrupa and fish otoliths. The second subunit (Wemmel Sand W2: 27.68 m - ca. 22 m) is represented by clayey silty fine sand, with common N. orbignyi and increasing clay content in its upper three meters, while the third (W3: 22 m - 20 m) is a darker coloured (dark green), somewhat coarser, clayey fine sand with abundant coarse glauconite and frequent N. orbignyi. The topmost 0.5 m (Wemmel Sand W4) is very heterogeneous, consisting of two distinct, partly calcite-cemented Nummulites beds, separated by clayey to silty sand.
3.6.2. Calcareous microfauna and macrofauna
40There are substantial differences in microfauna between the basal meter of the Wemmel Sand Member (W1) and its main body (W2-W4). In the lowermost meter, this fauna is characterised by abundant N. variolarius, associated with frequent specimens of a highly flattened Nummulites sp. of the prestwichianus group. The latter does not appear to have been recorded previously in Belgium. The foraminiferal taxon Fabularia is frequent, and occurs commonly associated with abraded fragments of Alveolina and Orbitolites in the highest quartz granule bed (27.70 m). It is probable that a portion of the microfauna, particularly the ’larger foraminifera’ has been reworked from the underlying Lede Sand Formation. Planktonic foraminifera occur in low abundance, but at 27.7 m index P is around 5%, the highest value recorded in the Wemmel Sand Member of Belgium.
41A diverse benthic microfauna has been observed in the main body of the unit (W2-W3). This has been well-documented previously from other sections in Belgium (Keij, 1957; Kaasschieter, 1961) and is not analysed in detail here. Nummulites orbignyi is common throughout. Anomalinoides affine, Asterigerina bartoniana, Cibicidoides proprius and polymorphinids are common. Planktonic foraminifera occur throughout, but in low abundance (P<2%). Pseudohastigerina is dominant, as noted previously by Hooyberghs (1984b). The ostracod assemblage is dominated by cytherideids, Leguminocythereis and Thracella. Molluscs are sparse and mostly partially decalcified. In W4 the microfauna seems to be similar, although difficult to identify due to secondary growth of calcite overgrowths.
42The microfauna of the Wemmel Sand Member, except for its basal meter, indicates an inner neritic environment, probably somewhat deeper than the Lede Sand Formation. The highest unit, the nummulite coquina, is indicative of an abrupt shallowing, as it is apparently a current-sorted accumulation.
3.6.3. Calcareous nannofossils
43The assemblages are poorly preserved and strongly overgrown in the lowermost m of the Wemmel Sand Member (W1). They are moderately rich (10 per field of view at 1000x magnification), and present a moderate to high species diversity (around 30 taxa). Rhabdosphaeraceae (24%), Prinsiaceae (22%) and Discoasteraceae (16%) are dominant. B. bigelowii is less frequent (4%). There is substantial reworking from the Cretaceous (4%) and to a much lesser degree from the Lede Formation (e.g. a few specimens of Braarudosphaera stylifera). The assemblages are marked by Blackites gladius, and the lowest occurrence of Sphenolithus furcatholitoides and Naninfula deflandrei.
44Subunit W2 shows an increase in nannofossils, in species diversity (between 30 and 35 taxa), as well as in number of specimens (15/field of view), which are furthermore much better preserved. The assemblage is dominated by Prinsiaceae (27%), Rhabdosphaeraceae (around 20%), L. minutus (around 15%) and Z. bijugatus (around 10%: co-occurrence of rectangular and triangular forms). Discoasteraceae are less well represented (<2%). It is furthermore marked by a series of first appearances, in ascending order these of Blackites trochos and Blackites pseudomorionum (from 27.60 m onward), followed by these of Reticulofenestra aff. umbilica (Dmax<13µm) and Pontosphaera formosa (both at 24.50 m).
45The assemblages in W3 are quite similar to these in the upper part of subunit W2, in terms of general nannofossil abundance, species diversity (30 to 35 taxa) and preservation. The most dominant groups throughout this unit are also Prinsiaceae, Rhabdosphaeraceae, Lanternithus minutus and Zyghrablithus bijugatus, although with slightly fluctuating proportions. B. bigelowii is fairly frequent (4%) throughout the unit. Nannotetrina fulgens and Dictyococcites onustus are first recorded in W3, although only very rarely. Similar assemblages are recorded in W4, although with a major decrease in Braarudosphaeraceae. The base of W4 is marked by the first record of Sphenolithus cuniculus. The co-occurrence of B. gladius and N. fulgens allow their attribution to the middle part of NP15.
3.7. Asse Clay Member (lower Maldegem Formation): 19.56 m - 18.00 m
3.7.1. Lithology
46The highly glauconitic and highly burrowed sandy silt and sandy clays, overlain by fossiliferous (essentially Lentipecten corneum) silty clays, between 19.56 m (diffuse junction marked by deep glauconite-filled burrows, which begin at 19.56 m and end at 19.45 m; Cz8 in Table 1) and 18.00 m depth are attributed to the Asse Clay Member.
3.7.2. Calcareous microfauna and macrofauna
47The diverse benthic foraminiferal assemblage includes common to abundant Asterigerina bartoniana, frequent Loxostomum teretum and Neoeponides karsteni, and rare Lenticulina gutticosta and Uvigerina farinose. There is an increase in the proportion of planktonic foraminifera at the base of the Asse Clay Member. P = ca. 3% at 19.30 m, increasing to ca. 25% at 18.7 m. Muricoglobigerina is dominant in this sample, but Pseudohastigerina is also common. The ostracod assemblage is similar to that in the Wemmel Sand Member. The pectinid bivalve L. corneum, characteristically abundant in the Asse Member at other localities, is here abundant between 18.8 m and 18.3 m, and occurs more rarely up to ca. 18.1 m.
48The benthic microfauna of the Asse Clay Member indicates somewhat deeper water than during the deposition of the Wemmel Sand Member. This is also indicated by the steadily increasing proportions of planktonic foraminifera. High proportions of planktonic foraminifera were also noted in the Asse Member in the Knokke borehole (King, 1990). This indicates an increase in water depths from the base of the Asse Clay Member.
3.7.3. Calcareous nannofossils
49The assemblages are only moderately preserved, but highly diverse (~35 taxa) and quantitatively rich (between 20 and 25 specimens/field of view), especially in the lowermost meter. They are dominated by Prinsiaceae (~50%). Rhabdosphaeraceae (~15%) and Coccolithus pelagicus (~13%) are also common; Pontosphaera spp. (~6%), L. minutus (~5%) and Z. bijugatus (~5%, exclusively triangular forms) are frequent. From 18.50 m and upwards the assemblages become progressively impoverished because of increasing decalcification.
50The assemblages of the Asse Clay Member are marked by a series of lowest occurrences, among which these of Nannotetrina pappii, Dictyococcites callidus and Discoaster martinii. This, in association with the presences of B. gladius and N. fulgens, indicates the upper part of NP15 (see Steurbaut, 1986 and Steurbaut in Schuler et al., 1992).
3.8. Remainder of the Maldegem Formation: 18.00 m - 11.66 m
51The Asse Clay Member is abruptly passing into the overlying Ursel Clay Member without any interruption in sedimentation. The latter, which consists of non-glauconitic silty clay, is completely decalcified in the Zemst hole. An omission surface is recorded at 16.61 m depth, separating the Ursel Clay Member from the overlying non-calcareous Onderdale Sand Member and the rest of the Maldegem Formation.
4. Discussion
4.1. The Zemst dataset and its impact on Belgian stratigraphy
52Due to its strategic position halfway between Brussels and Antwerp (Fig. 1), the Zemst borehole plays a crucial role in understanding the lateral distribution of the upper Ypresian lithostratigraphic units in Belgium, which were nearly all defined in areas westwards of Zemst (Steurbaut & Nolf, 1986; Steurbaut, 1998, 2006b). A substantial hiatus of at least 0.2 myr has been recorded between the Tielt Formation and the overlying Panisel Sand Member at Zemst (Cz1 in Table 1; Fig. 4), because of the absence of at least the Egem Sand Member and the Bois-la-Haut Sand Member (probably other units as well). Both the Egem Sand and the Bois-la-Haut Sand Members were deposited during chron C23n.1n (Steurbaut, 1998, fig. 4 and unpubl. information), of which the duration is about 0.21 myr (Vandenberghe et al., 2012, p. 899). This magnetostratigraphic interpretation is a re-calibration of data from Belgium (Ali et al., 1993), Denmark (Ali, 1988, unpubl. PhD) and North Sea borehole 81/46A (Steurbaut, 2011 and unpubl. information).
53Comparison with the Mont Panisel area (Steurbaut & King, 1994), about 70 km southwest of Zemst and with the Kester area (Steurbaut, unpubl. information) about midway between these two, witnesses the progressive thinning of the Panisel Sand Member in northeastern direction, from about 21 m at its type-locality to about 8 m at Kester. It also shows a similar thickness further northeast (e.g. ~8 m at Zemst) and northwest (e.g. ~9 m at Kerksken). The identification of the Kwatrecht Member at Zemst erases any doubts that might linger about its very existence. This is its first undoubted record in Belgium, besides its initial record at the type locality (De Moor & Geets, 1974). The Merelbeke Clay Member is widespread in central Belgium, and is the youngest upper Ypresian unit being deposited at Zemst and probably further eastwards. However, this unit is rapidly thinning from a maximum thickness of about 12 m (?15 m) in the Gent area (De Moor & Geets, 1974) to about 3.6 m at Zemst.
54This investigation throws also new light on the composition and distribution of the Lutetian units in Belgium. At Zemst the lowermost Lutetian unit is the Lede Formation, which is about 9 m thick. This is considerable (cf. ~6.80 m at Balegem: Nolf & Steurbaut, 1990), but not exceptional (Kaasschieter, 1961, map 15: thicknesses over 15 m in the Leuven area). The overlying Wemmel Sand Member is almost 9 m thick, and thus quite expanded. Its thickness generally fluctuates between 2 m to 6 m, east of Brussels, increasing to 14 m west of Brussels (Rutot, 1882). According to Kaasschieter (1961, map 5) its maximum thickness is recorded SW of Brussels, reaching up to 18 m, if this interval is correctly interpreted. At Zemst it is subdivided in 4 subunits, of which the lowermost consists of thin quartz gravels beds, with considerable reworking from the underlying Lede Sand Formation, and the two uppermost have never been recorded or investigated in detail in Belgium (one uncertain, although possible record by Rutot, 1887 at Ukkel: 2 m of oxidized and leached fine sand below the Asse Clay). The Asse Clay and Ursel Clay Members are both present in their typical form, although very reduced in thickness (both around 1.5 m thick). The transition between both units is sharp, without break in sedimentation, and results from a sudden deepening of the depositional system. Both the base of the Asse Clay and the top of the Ursel Clay are separated from adjacent units through interburrowed omission surfaces (Cz8 and Cz9 in Table 1 respectively).
55The Zemst data contribute much in the understanding of the nature and distribution of the third, fourth and fifth order depositional sequences in the upper Ypresian and lower and middle Lutetian in Belgium. Substantial evidence is available now to believe that the Panisel Sand Member encompasses the main body of a single depositional sequence, labelled Y-21, and that the Kwatrecht Member and the Merelbeke Clay Member, respectively represent the lowstand and highstand systems tract of an overlying sequence Y-22 (Steurbaut, unpubl. information). The transgressive systems tract of the latter, at the base of the Merelbeke Clay, is generally very condensed (10 cm of glauconitic silty fine sand, Fig. 4). The bases of the Lede Sand Formation, the Wemmel Sand Member, the Asse Clay Member and the Onderdale Sand Member are marked by omission surfaces, which all correspond to (probably third order) sequence boundaries (Fig. 4).
4.2. Organic carbon isotope evolution in the upper Ypresian of Belgium
56The upper Ypresian carbon isotope record, as presented in Vandenberghe et al. (GTS, 2012, fig. 28.11) is based on the compilation of data from Cramer et al. (2009), integrating a series of deep-sea cores from different latitudes and longitudes in world’s major oceans (essentially Atlantic, Pacific and high-latitude Southern oceans). Continuous high-resolution carbon isotope profiles from single localities or palaeoenvironmentally uniform areas (e.g. sub-basins such as the Belgian and the Paris Basins), and covering the entire upper Ypresian (52-47.8 Ma) are very scarce. The high-resolution study of the Demerara Rise stable isotopes (tropical western Atlantic), covering the entire lower and middle Eocene, is one of the few exceptions (Sexton et al., 2006). Mostly these records represent only parts of the Ypresian (Schmitz et al., 1996: Denmark, middle Ypresian to lower upper Ypresian; Arenillas et al., 1999: Possagno Italy, up to lower Ypresian; Cramer et al., 2003, Lourens et al., 2005, Stap et al., 2010, D’haenens et al., 2014: essentially Atlantic ODP holes, up to middle Ypresian; Coccioni et al., 2012: Gubbio, Italy, up to lowermost upper Ypresian; Zachos et al., 2010 and Westerhold et al., 2011: up to lowermost Ypresian from southern Atlantic and tropical Pacific respectively).
57Although the carbon isotope record of the Zemst borehole also covers only a small part of the upper Ypresian, it may help in understanding the organic carbon isotope evolution through the Early Eocene Climatic Optimum (EECO) in shallow marine settings at middle and high latitudes. The EECO is a 1.5-million-year-long global warming (from ca. 51.8 Ma to ca. 50.4 Ma), expressed by low δ18O values, which also seem to coincide with a substantial decrease in δ13C values (Zachos et al., 2001; 2008; Sexton et al., 2006, fig. 9). It has been identified in different environments, from continental (e.g. Smith et al., 2014, USA) through shallow marine (e.g. Deprez et al., 2015, Kazakhstan) to deep water settings (e.g. Sexton, 2006, Atlantic Ocean; Bijl et al., 2009, Pacific Ocean; Shamrock, 2010, eastern Indian Ocean; Luciani et al., 2015, Italy).
58The upper Ypresian in the Zemst borehole and in the Egem quarry (Fig. 4) consists of a series of units which were deposited in relatively marginal marine conditions, although representing various sedimentary facies. Palynofacies analysis reveals very low rates of reworking, minor amounts of amorphous organic matter (except for sample Z5 in the lower part of the Merelbeke Clay) and a fairly constant influx of terrestrial derived organic matter (around 55%, increasing to about 90% in the two uppermost samples) (Table 2). The palynofacies composition of the upper part of the Merelbeke Clay at Zemst (Z6) is very similar to that of the lower part of the Pittem Clay at Egem, although the organic carbon isotope values are quite different, while the two samples from the Merelbeke Clay, which show different palynofacies compositions present nearly identical organic carbon isotope values (Table 2). This suggests that the organic carbon isotope trends as recorded in the Zemst borehole and at Egem are not likely to be substantially biased by variation in types of organic matter.
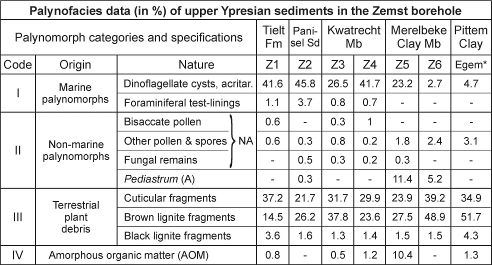
59Table 2. Palynofacies data (% of the total palynomorph assemblage) of the upper Ypresian sediments in the Zemst borehole and the Egem quarry (* = sample from 0.7 m above the top of the basal sandstone of the Pittem Clay) (the marine palynomorphs are dominated by dinoflagellate cysts, but also include rare acritarchs; NA = non-aquatic; A = aquatic).
60In the Zemst borehole there is a 1‰ negative shift in δ13Corg values from -26.1‰ at the top of the Tielt Formation to -27.1‰ near the middle of the overlying Panisel Sand Member (no data from the base of this member, Fig. 4). These organic carbon isotopevalues remain fairly negative(around -26.5‰) in the overlying Kwatrecht Member and the Merelbeke ClayMember (with a minimum of -27.0‰ in the middle of the Merelbeke Member). They are quite negative compared to overlying units (Noiret, unpubl. information), and consequently indicate that the interval from the Panisel Sand Member up to the Merelbeke Clay Member was deposited during the EECO. The deposition of the Merelbeke Clay, which is within lower NP13 and in early chron C22r (Steurbaut et al., submitted), probably took place during its final phase. The 0.5‰ to 1‰ positive shift at the base of the overlying Pittem Clay to about -26.0‰ (as in the Egem quarry, Fig. 4) marks the return to pre-EECO organic carbon isotope values (as recorded in the Kortemark quarry, Noiret, unpubl. information). A similar positive shift in δ13C values, coinciding with a positive shift in δ18O values, has been observed in deep-sea profiles in the tropical western Atlantic (Demerara Rise) at approximately 50.4 Ma, within the middle of chron C22r (top of event c of Sexton et al., 2006). It corresponds to the end of the EECO.
4.3. Dinoflagellate cyst taxon Apectodinium as proxy for global warming
61Since Crouch et al. (2001) presented evidence that the Apectodinium blooms during the PETM are expected to be associated with exceptionally high global sea-surface temperatures and/or an substantial increase in marginal-marine surface-water productivity (enhanced nutrient input), many additional records of these PETM blooms have been published from all over the globe, including the North Sea Basin (Heilmann-Clausen & Schmitz, 2000; Steurbaut et al., 2003) (see Sluijs et al., 2008a and b, for overviews). Since these blooms lasted many ten thousands of years, Sluijs et al. (2008a) suggested that, besides temperature and nutrient availability, a third yet unknown critical environmental factor, stimulating massive Apectodinium production had to be involved.
62If sea surface temperature was one of the driving factors behind the development of the Apectodinium blooms, these should also be observed during subsequent periods of global warming, such as the ETM-2 (Eocene Thermal Maximum 2), ETM-3 (Eocene Thermal Maximum 3) and the EECO. Apectodinium is generally absent during the ETM-2 within the Arctic region (except for a few specimens at the start), probably because sea surface salinity dropped below tolerable values (Sluijs et al., 2009). Agnini et al. (2009) recorded an Apectodinium bloom (20% to 50% of the total assemblage) within the ETM-3 event (also known as X-event) in the Farra d’Alpago section (NE Italy). Additional data on dinoflagellate cyst distribution during ETM-2 and ETM-3 are scarce because both periods of global warming have not been unequivocally identified in many areas.
63In the Wursterheide borehole (N Germany) Apectodinium (essentially A. homomorphum) is consistently represented with respectable percentages (1% to 4%) in the lower part of the Areosphaeridium diktyoplokum zone (Heilmann-Clausen & Costa, 1989), which according to its position should correspond to part of the EECO. The top of this Apectodinium interval is nearly coincident with the HO of Cordosphaeridium biarmatum, which in Belgium is located within the Merelbeke Clay (De Coninck, 1977). Apectodinium (essentially A. homomorphum and A. quinquelatum) is also present in SW Siberia (Iakovleva & Heilmann-Clausen, 2010), where it has been recorded in low numbers in the interval bracketed by the LO of the Charlesdowniea columna-group and the middle part of NP13, which, in Belgium, should approximately correlate to the interval from the base of the Egem Sand Member to the top of the Merelbeke Clay.
64In the Zemst borehole Apectodinium, merely represented by A. homomorphum and A. quinquelatum, is rather frequent from the top of the Panisel Sand Member (3.4%; the only dinoflagellate cyst sample from the Panisel Sand Member at Zemst) up to the top of the Merelbeke Clay (7.5%), with a maximum of 13.6% at the base of the Kwatrecht Member. Similar distribution patterns have been recorded in the Kallo borehole (De Coninck, 1976), with the start of a substantial increase in Apectodinium (0 to 3%) at 268 m depth and a maximum of 20% at the base of the Egem Sand Member (259 m). Moderately to high Apectodinium percentages (>3%) seem to last up to the top of the Egem Sand Member (242 m), followed by a decrease to 1% in the Merelbeke Clay Member. Comparable increased frequencies (13%) are known from isolated samples in the Panisel Sand Member at Steenhuize-Wijnhuize, 44 km WSW of Zemst (Vanhove & De Coninck, 1992). In the Woensdrecht borehole, located close to the Belgian border ca. 18 km north of Kallo, substantial amounts of Apectodinium (4%) have been recorded within the Merelbeke Clay (De Coninck, 1977). This is followed in the Woensdrecht borehole, as in all Belgian sections studied, by a major decrease to less than 0.1% (generally only a few specimens) from the base of the Pittem Clay onward (De Coninck, 1977 and many unpublished data from Steurbaut et al.).
4.4. Fine tuning of the upper Ypresian - Lutetian calcareous nannofossil zonation at mid-latitudes
65Belgian sections have proved to be crucial for establishing a high-resolution upper Ypresian to middle Lutetian calcareous nannofossil zonation for mid-latitudes (Steurbaut, 1986, 1988, 1990, 2011; Damblon & Steurbaut, 2000; Herman et al., 2001; Smith et al., 2004). The multidisciplinary investigation of the Kysing-4 borehole in Denmark did clarify how these nannofossils further evolved through the late Lutetian in these middle latitudinal areas (Thomsen et al., 2012).
66The nannofossil record of the Zemst borehole allows fine tuning of the distribution patterns of important index taxa, which are fundamentally altering or enhancing the resolution of the existing zonation. Among the most relevant conclusions are:
671. The NP12/NP13 boundary coincides with the boundary between the Panisel Sand Member and the overlying Kwatrecht Member.
682. At mid-latitudes the topmost part of NP12 is marked by the LO of Helicosphaera aff. lophota, as shown by its presence at Zemst (46.50 m), Aktulagay, Kazakhstan (18.05 m) and Albaekhoved, Denmark (Al-64 at 18.30 m) (for data other than Zemst, Steurbaut, unpubl. information).
693. The associations in the Panisel Sand Member and to a lesser degree the Kwatrecht Member at Zemst are dominated by Discoaster kuepperi (between 30% and 50% in the former, and 16% and 30% in the latter). “Inc. sed. sp.”, a small globular calcitic body of unknown origin (see chapter 3.2.3) is also frequent in that interval. In the North Sea Basin, the start of the acme of D. kuepperi seems to coincide or is very close to the LO of “Inc. sed. sp.” and the LO of Reticulofenesta spp. The latter, which is marked by the popping up of very small Reticulofenstra specimens, although initially in low numbers, defines the base of subzone VIb of Steurbaut (1991, fig. 3; 1998). The LO of Reticulofenestra spp. corresponds to the FO of Noelarhabdaceae in Agnini et al. (2006, fig. 3). According to these authors this FO slightly predates the first common occurrence of the taxon (FCO). There is a major decrease in Discoaster kuepperi to less than 0.5% at the base of the Pittem Clay Member (Steurbaut, unpubl. information).
704. The basal part of NP15 is missing in most parts of the Belgian Basin, including Zemst. It has only been identified at Zaventem (Herman et al., 2001), and is marked by the total range of Nannotetrina alata, in association with the presence of Blackites gladius.
715. The LO of Nannotetrina fulgens and the LO of N. pappii are not coincident in the Belgian Basin, as previously thought (at base of Asse Clay Member; Steurbaut, 1986). The Zemst data indicate for the first time that N. fulgens appears earlier (at the base of subunit W3 of the Wemmel Sand Member).
726. The data from Zemst and other Belgian localities (e.g. Zaventem, Oosterzele, Stekene, and many others) reveal that 1. Nannotetrina alata - Blackites gladius, 2. Nannotetrina quadrata, 3. Sphenolithus furcatholitoides, 4. Clausicoccus obrutus, 5. Blackites trochos - Blackites pseudomorionum, 6. Nannotetrina fulgens, 7. Sphenolithus cuniculus and 8. Nannotetrina pappii - Discoaster martinii - Dictyococcites callidus constitute a chronological sequence of appearances within the middle Lutetian nannofossil zone NP15. Further subdivision can be achieved using the HO of Braarudosphaera stylifera, which occurs between event 2 and 3, and the HO of Blackites gladius, which defines the upper boundary of zone NP15 (Fig. 4).
4.5. Reflection on the Ypresian-Lutetian transition in Belgium
73A close inspection of the Zemst borehole and many other relevant outcrop and borehole sections in Belgium (Steurbaut, 2006b) reveals that the Ypresian-Lutetian transition is very incomplete in large parts of the Belgian Basin. This is due to non-deposition or subsequent erosion of stratigraphic units, which in turn is controlled by the geological outline of the area, the morphology of the palaeocoastline and changes in eustatic sea-level and in local tectonic behaviour, e.g. uplift of Brabant Massif in response to remote plate tectonic movements. The most complete successions are recorded in the Flemish hills (such as the Cassel and Mont-des-Récollets hills), a series of geological outliers, extending from central Belgium to NW France (Fig. 1). These hills are capped by ferruginous sandstones, presumably of Late Eocene age (Houthuys, 2014), which protected the hill-zone from post-Eocene erosion and denudation, which in the adjacent areas exceeded 100 m (Nolf & Steurbaut, 1990; Steurbaut & Nolf, in prep.). At Zemst there is a major hiatus of about 3 to 4 myr between the top of the Ypresian and the base of the Lutetian (Cz4 in Table 1), as shown by the absence of the upper Gentbrugge Formation (from Pittem Clay to Aalterbrugge Members) and the entire Aalter Sand and Brussel Sand Formations (Fig. 2). Towards the east and southeast the stratigraphic gap between the Ypresian and Lutetian tends to decrease strongly as the up to 70 m thick Brussels Formation, the infill of a complex megachannel structure, is interposing between the Merelbeke Clay and the Lede Sand Formation (Houthuys, 1990, 2011; Damblon & Steurbaut, 2000). In westward direction (Gent and further NW to Aalter, Steurbaut & Nolf, 1989) the Gentbrugge and Aalter Formations are completely preserved, but only parts of the Brussel Formation (e.g. off shore borehole Vlakte van de Raan, Steurbaut, 2011) and locally the Lede Formation have been identified. This brings us to the main issue, the identification of the base of the Lutetian in the Belgian Basin.
74The base of the Lutetian has in the past been defined at the ‘first occurrence datum’ of the planktonic foraminiferal taxon Hantkeninaat ca.48.6 Ma (e.g. Luterbacher et al., 2004), corresponding approximately to the base of calcareous nannofossil zone NP14, but recent studies have shown this Hantkeninaevent to be significantly diachronous. In April 2011 the International Union of Geological Sciences ratified the proposal that the base of the Lutetian is defined by the Gorrondatxe GSSP (NW Spain) (Molina et al., 2011). This boundary is placed at the 167.85 m level, corresponding to the lowest occurrence (LO) of the nannofossil taxon Blackites inflatus. The latter is well known from many low and middle latitude outcrop and borehole sections worldwide (Southern USA: Siesser, 1983; Russia: Shcherbinina, 2000; off NW Australia: Shamrock, 2010, etc.), but seems to be missing at higher mid latitudes, such as the North Sea Basin (not mentioned by Perch-Nielsen, 1971; Bigg, 1982; Aubry, 1983; Steurbaut, 1988; Verbeek et al., 1988; Varol, 1998). The most northern occurrence of B. inflatus in NW Europe is the Paris Basin (Aubry, 1983), where it occurs in very low numbers in the middle and upper part of the “Glauconie grossière” (e.g. at Margival, Steurbaut, 1988). The identification of additional nannofossil events, coeval with the LO of B. inflatus, is part of ongoing work, aiming at the exact positioning of the base of the Lutetian in the North Sea Basin s.s. (excluding the Paris Basin, home of the historical Lutetian stratotype).
5. Conclusions
75High-resolution sedimentological, micropalaeontological and organic carbon isotope investigations at Zemst have enabled to gain new insight in the Ypresian-Lutetian transition in central Belgium. Although the upper Ypresian succession is very incomplete (upper half of the Gentbrugge Formation and the entire Aalter Formation are missing), the Zemst data, and associated cross-correlations, have allowed for the first time to identify the NP12/NP13 boundary, and the nearly coincident boundary between chron C23n and chron C22r, in the Belgian Basin. These boundaries, which are dated at 50.63 Ma, fall within the break in sedimentation between the Panisel Sand Member and the overlying Kwatrecht Member. In terms of dinoflagellate cyst distribution, they are bracketed between the LO of Impletosphaeridium kroemmelbeinii and the LCO of Areosphaeridium diktyoplokum below and the HO of Areoligera senonensis and the LO of Impletosphaeridium cracens above. This study also definitely proves that the term P1n clay, which was used until recently, corresponds to the Merelbeke Clay and that its introduction (Rutot, 1890) was erroneous, due to insufficient knowledge of the Belgian upper Ypresian stratigraphy at the end of the 19th century.
76The relatively low values in δ13Corg (-26.5‰to -27.1‰) in the interval from the Panisel Sand to the Merelbeke Clay Members (upper NP12-lower NP13) are believed to correspond to the 1.5-million-year-long negative carbon isotope interval, recorded globally between approximately 51.8-50.4 million years ago and associated with global warming, known as the Early Eocene Climatic Optimum (EECO). This is the first positive identification of the EECO in the North Sea Basin. The negative carbon isotope values, which, according to the palynofacies data, are not likely to be substantially biased by variation in types of organic matter, are shown to be coinciding with a fairly high frequency of Apectodinium (>3%, up to 14%) and a Discoaster-bloom (16%-50%, essentially D. kuepperi). This is quite analogous, although less strongly pronounced, to what has been observed during the Paleocene-Eocene Thermal Maximum (PETM), the most prominent global warming event of the Cenozoic (Apectodinium-abundance, up to 75% at mid and high latitudes, e.g. Steurbaut et al., 2003 and abundance peaks of Discoaster in tropical areas, e.g. Discoaster spp. up to 35% of total assemblage in Indian Ocean Site 213, Kahn & Aubry, 2004; D. salisburgensis, up to 12% in Tanzania, Bown & Pearson, 2009). The abrupt positive shift of up to 1‰ in δ13Corg values at the base of the Pittem Clay Member at Egem (mid-NP13, mid-chron C22r), which is coincident with the virtual disappearances of Apectodinium (<0.1%) and Discoaster (<0.5%) is suggested to mark the end of the EECO in the southern North Sea Basin.
77The absence of the Brussel Formation also reveals that the Lutetian is incomplete at Zemst. The Lede Sand Formation is quite expanded and marked by the co-occurrence of the calcareous nannofossil taxa Blackites gladius and Braarudosphaera stylifera, indicating lower NP15, although not its basal part. Four distinct units have been identified in the overlying Wemmel Sand Member (middle NP15), of which the upper two, which are marked by the LO of Nannotetrina fulgens and the absence of N. pappii, have not been recorded before in Belgium. The Asse Clay Member (upper NP15) and the overlying Ursel Member (decalcified), which are part of a single depositional sequence, are reduced in thickness (both around 1.5 m thick). The Zemst data and addition outcrop and borehole data from Belgium reveal that 1. Nannotetrina alata - Blackites gladius, 2. Nannotetrina quadrata, 3. Sphenolithus furcatholitoides, 4. Clausicoccus obrutus, 5. Blackites trochos - Blackites pseudomorionum, 6. Nannotetrina fulgens, 7. Sphenolithus cuniculus, and 8. Nannotetrina pappii - Discoaster martinii - Dictyococcites callidus constitute a chronological sequence of appearances within the middle Lutetian nannofossil zone NP15. They also indicate that the major criterion for identifying the base of the Lutetian (LO of Blackites inflatus), as designated at the Gorrondatxe GSSP, is not applicable in the North Sea Basin s.s. (excluding the Paris Basin) and that the search for appropriate substitutes associated with this boundary is needed to allow interbasinal stratigraphic correlation during the Eocene at middle and high latitudes.
6. Acknowledgements
78The authors are much obliged to Philip Buffel (Korbeek-Lo) and Marleen De Ceuckelaire (KBIN, Brussel) for having provided the samples from the Zemst borehole. They are grateful to Adriano Vandersypen (RBINS) for help in production of the figures. Dr. Claus Heilmann-Clausen (Arhus Universitet, Denmark) and Dr. Malcolm Hart (Plymouth University, UK) are thanked for their constructive comments and suggestions, improving the manuscript.
7. References
79Agnini, C., Macrí, P., Backman, J., Brinkhuis, H., Fornaciari, E., Giusberti, L., Luciani, V., Rio, D., Sluijs, A. & Speranza, F., 2009. An early Eocene carbon cycle perturbation at ~52.5 Ma in the Southern Alps: Chronology and biotic response. Paleoceanography, 24, PA2209, 14 p.
80Agnini, C., Muttoni, G., Kent, D.V. & Rio, D., 2006.Eocene biostratigraphy and magnetic stratigraphy from Possagno, Italy: The calcareous nannofossil response to climate variability. Earth and Planetary Science Letters, 241, 815-830.
81Ali, J.R., 1988. Magnetostratigraphy of Early Palaeogene sediments from N.W. Europe. Unpublished PhD Thesis, University of Southampton, UK, 235 p.
82Ali, J.R., King, C. & Hailwood, E.A., 1993. Magnetostratigraphic calibration of early Eocene depositional sequences in the southern North Sea Basin. In Hailwood, E.A. & Kidd, R.B. (eds), High Resolution Stratigraphy. Geological Society, Special Publication, 70, 99-125.
83Anonymous, 1893. Légende de la Carte Géologique dressée par ordre du Gouvernement à l’échelle du 40.000e. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 6 (1892), 217-229.
84Arenillas, I., Molina, E. & Schmitz, B., 1999. Planktic foraminiferal and δ13C isotopic changes across the Paleocene/Eocene boundary at Possagno (Italy). International Journal of Earth Sciences, 88, 352-364.
85Aubry, M.-P., 1983. Biostratigraphie du Paléogène épicontinental de l’Europe du Nord-Ouest. Etude fondée sur les nannofossiles calcaires. Documents des Laboratoires de Géologie de Lyon, 89, 317 p.
86Aubry, M.-P. & Bord, D., 2009. Reshuffling the cards in the photic zone at the Eocene: Oligocene boundary. In Koeberl, C. & Montanari, A. (eds), The Late Eocene Earth. The Geological Society of America, Special Paper, 452, 279-301.
87Bigg, P.J., 1982. Eocene planktonic foraminifera and calcareous nannoplankton of the Paris Basin and Belgium. Revue de Micropaléontologie, 25(2), 69-89.
88Bignot, G., 1989. Les Pseudarcellidés, groupe orginal de microfossiles incertae sedis du Paléogène. Remarques sur sa classification et description de trois espèces nouvelles. Revue de Micropaléontologie, 31(4), 211-224.
89Bijl, P.K., Schouten, S., Sluijs, A., Reichart, G.-J., Zachos, J.C. & Brinkhuis, H., 2009. Early Palaeogene temperature evolution of the southwest Pacific Ocean. Nature, 461, 776-779.
90Bown, P. & Pearson, P., 2009. Calcareous plankton evolution and the Paleocene/Eocene thermal maximum event:New evidence from Tanzania. Marine Micropaleontology, 71, 60-70.
91Cahuzac, B. & Janssen, A., 2010.Eocene to Miocene holoplanktonic Mollusca (Gastropoda) of the Aquitaine Basin, southwest France. Scripta Geologica, 141, 1-193.
92Coccioni, R., Bancalà, G., Cantanzarit, R., Fornaciari, E., Frontalini, F., Giusberti, L., Jovane, L., Luciani, V., Savian, J. & Sprovieri, M., 2012. An integrated stratigraphic record of the Palaeocene-lower Eocene at Gubbio (Italy): new insights into the early Palaeogene hyperthermals and carbon isotope excursions. Terra Nova, 24, 380-386.
93Costa, L.I. & Manum, S.B., 1988.The description of the interregional zonation of the Paleogene (D1-D15) and the Miocene (D16-D20). In Vinken, R. (comp.), The Northwest European Tertiary Basin. Geologisches Jahrbuch, A, 100, 321-332.
94Cramer, B.S., Wright, J.D., Kent, D.V. & Aubry, M.-P., 2003. Orbital climate forcing of δ13C excursions in the late Paleocene–early Eocene (chrons C24n–C25n). Paleoceanography, 18(4), 1097, 25 p.
95Cramer, B.S., Toggweiler, J.R., Wright, J.D., Katz, M.E. & Miller, K.G., 2009. Ocean overturning since the Late Cretaceous: Inferences from a new benthic foraminiferal isotope compilation. Paleoceanography, 24, PA4216, 14 p.
96Crouch, E.M., Heilmann-Clausen, C., Brinkhuis, H., Morgans, H.E.G., Rogers, K.M., Egger, H. & Schmitz, B., 2001. Global dinoflagellate event associated with the late Paleocene thermal maximum. Geology, 29, 315-318.
97Damblon, F. & Steurbaut, E., 2000. Gobertange: site géologique remarquable. In La Gobertange – Une Pierre, des hommes. ASBL Gobertange 2000, Cera Holding, 19-48.
98De Coninck, J., 1976. Microfossiles à paroi organique de l’Yprésien du Bassin belge. Professional Paper, BelgischeGeologische Dienst (1975), 12, 1-151.
99De Coninck, J., 1977. Organic walled microfossils from the Eocene of the Woensdrecht borehole, southern Netherlands. Mededelingen RijksGeologische Dienst, N.S. 28(3), 33-64.
100De Moor, G. & Geets, S., 1974. Sedimentologie en litostratigrafie van de Eocene afzettingen in het zuidoostelijk gedeelte van de Gentse agglomeratie. Natuurwetenschappelijk Tijdschrift, 55 (1973), 129-192.
101Deprez, A., Tesseur, S., Stassen, P., D’haenens, S., Steurbaut, E., King, C., Claeys, P. & Speijer, R.P., 2015. Early Eocene environmental development in the northern Peri-Tethys (Aktulagay, Kazakhstan) based on benthic foraminiferal assemblages and stable isotopes (O, C). Marine Micropaleontology, 115, 59-71.
102D’haenens, S., Bornemann, A., Claeys, P., Röhl, U., Steurbaut, E. & Speijer, R.P., 2014.A transient deep-sea circulation switch during Eocene Thermal Maximum 2. Paleoceanography, 29, 370-388.
103Dybkjær, K., 2004. Morphological and abundance variations in Homotryblium-cyst assemblages related to depositional environments; uppermost Oligocene–Lower Miocene, Jylland, Denmark. Palaeogeography, Palaeoclimatology, Palaeoecology, 206(1–2), 41–58.
104Fobe, B., 1986. Petrografisch onderzoek van de coherente gesteenten van het Eoceen van Laag- en Midden-België. Unpublished PhD, Rijksuniversiteit Gent, part I, 215 p., part II, 138 p.
105Geets, S., 1992. Sedimentological investigation of the Mont-Panisel boring. Bulletin de la Société belge de Géologie, 99(3-4) (1990), 315-323.
106Giunta, S., Negri, A., Morigi, C., Capotondi, L., Combourieu-Nebout, N., Emeis, K.C., Sangiorgi, F., & Vigliotti, L., 2003. Coccolithophorid ecostratigraphy and multi-proxy paleoceanographic reconstruction in the Southern Adriatic Sea during the last deglacial time (Core AD91-17). Palaeogeography, Palaeoclimatology, Palaeoecology, 190, 39-59.
107Gulinck, M. & Hacquaert, A., 1954. L’Eocène. In Fourmarier, P. (ed.), Prodrome d’une description géologique de la Belgique. Société géologique de Belgique, Liège, 451-493.
108Heilmann-Clausen, C., 1985. Dinoflagellate stratigraphy of the uppermost Danian to Yprsian in the Viborg 1 borehole, central Jylland, Denmark. Danmarks Geologiske Undersøgelse, A, 7, 1-69.
109Heilmann-Clausen, C. & Costa, L.I., 1989. Dinoflagellate zonation of the Uppermost Paleocene? to Lower Miocene in the Wursterheide research well, NW Germany. Geologisches Jahrbuch, A, 111, 431-521.
110Heilmann-Clausen, C. & Schmitz, B., 2000. The late Paleocene thermal maximum δ13C excursion in Denmark? GFF, 122, p.70.
111Herman, J., Steurbaut, E. & Vandenberghe, N., 2001. The boundary between the Middle Eocene Brussel Sand and the Lede Sand Formations in the Zaventem-Nederokkerzeel area (Northeast of Brussels, Belgium).Geologica Belgica, 3(3-4) (2000), 231-255.
112Hooyberghs, H.J.F., 1984a. Planktonic foraminifera from the Lede Sands Formation at Balegem. Tertiary Research, 6, 165-175.
113Hooyberghs, H.J.F., 1984b. Planktonic foraminifera from the Wemmel Sands Member of the Asse Formation at Strombeek Bever (Belgium). Tertiary Research, 6, 93-106.
114Houthuys, R., 1999.Vergelijkende studie van de afzettingsstruktuur van getijdenzanden uit het Eoceen en van de huidige Vlaamse Banken. Aardkundige Mededelingen, 5, 137 p.
115Houthuys, R., 2011.A sedimentary model of the Brussels Sands, Eocene, Belgium. Geologica Belgica, 14(1-2), 55-74.
116Houthuys, R., 2014.A reinterpretation of the Neogene emersion of central Belgium based on the sedimentary environment of the Diest Formation and the origin of the drainage pattern. Geologica Belgica, 17(3-4), 211-235.
117Iakovleva, A.I. & Heilmann-Clausen, C., 2001.Eocene dinoflagellate cyst biostratigraphy of research borehole 011-BP, Omsk Region, southwestern Siberia. Palynology, 34(2), 195-232.
118Janssen, A.W., King, C. & Steurbaut, E., 2011. Notes on the systematics, morphology and biostratigraphy of fossil holoplanktonic Mollusca, 21. Early and Middle Eocene (Ypresian-Lutetian) holoplanktonic Mollusca (Gastropoda) from Uzbekistan. Basteria, 75(4-6), 71-93.
119Kaasschieter, J.P.M., 1961. Foraminifera of the Eocene of Belgium. Mémoires de l’Institut Royal des Sciences naturelles de Belgique, 147, 1-271.
120Kahn, A. & Aubry, M.-P., 2004. Provincialism associated with the Paleocene/Eocene thermal maximum: temporal constraint. Marine Micropaleontology, 52, 117-131.
121Keen, M.C., 1978. The Tertiary: Palaeogene. In Bate, R. & Robinson, E. (eds), A stratigraphical index of British Ostracoda. Seel House Press, Liverpool, Geological Journal Special Issue, 385-450.
122Keij, A.J., 1957. Eocene and Oligocene Ostracoda of Belgium. Mémoires de l’Institut Royal des Sciences naturelles de Belgique, 136, 1-210.
123King, C., 1989. Cenozoic of the North Sea. In Jenkins, D. & Murray, J. (eds), Stratigraphical atlas of fossil foraminifera (2nd ed.). Ellis Horwood, Chichester, British Micropalaeontological Society Series, 418 - 489.
124King, C., 1990. Eocene stratigraphy of the Knokke borehole (Belgium). In Laga, P. & Vandenberghe, N. (eds), The Knokke well (11E/138) with a description of the Den Haan (22W/276) and Oostduinkerke (35E/142) wells. Mémoires pour servir à l’Explication des Cartes Géologiques et Minières de la Belgique, 29, 67-102.
125King, C., 1991. Stratigraphy of the Ieper Formation and Argile de Flandres (Early Eocene) in western Belgium and northern France. In Dupuis, C., De Coninck, J. & Steurbaut, E. (eds), The Ypresian stratotype. Bulletin de la Société belge de Géologie, 97(3-4) (1988), 349-372.
126King, C., Iakovleva, A., Steurbaut, E., Heilmann-Clausen, C. & Ward, D., 2013. The Aktulagay section, west Kazakhstan: a key site for northern mid-latitude Early Eocene stratigraphy. Stratigraphy, 10(3), 171-209.
127Köthe, A., 2012. A revised Cenozoic dinoflagellate cyst and calcareous nannoplankton zonation for the German sector of the southern North Sea Basin. Newsletters on Stratigraphy, 45(3), 189-220.
128Lourens, L.J., Sluijs, A., Kroon, D., Zachos, J.C., Thomas, E., Röhl, U., Bowles, J. & Raffi, I., 2005. Astronomical pacing of late Palaeocene to early Eocene global warming events. Nature, 435, 1083-1087.
129Luciani, V., Backman, J., Fornaciari, E., Giusberti, L., Agnini, C. & D’Onofrio, R., 2015. Massive and permanent decline of symbiont bearing morozovellids and δ13C perturbations across the Early Eocene Climatic Optimum at the Possagno section (Southern Alps of northeastern Italy). Climate of the Past, 11(2), 671-712.
130Luterbacher, H.-P., Ali, J.R., Brinkhuis, H., Gradstein, F.M., Hooker, J.J., Monechi, S., Ogg, J.G., Powell, J., Röhl, U., Sanfilippo, A. & Schmitz, B., 2004. The Paleogene period. In Gradstein, F.M., Ogg, J.G. & Smith, A. (eds), A geologic time scale 2004. Cambridge University Press, Cambridge, 384-408.
131Martini E., 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation. Proceedings 2nd Planktonic Conference(Roma, 1970), Ed. Technoscienza, 2, 739-785.
132Matthijs, J. & Buffel, P., 2001. Korrelgrootte analyse van de gekernde boring 73E359 te Zemst. Rapport voor Ministerie van de Vlaamse Gemeenschap (ANRE), 8 p.
133Molina, E., Alegret, L., Apellaniz, E., Bernaola, ,G., Caballero, F., Dinarès-Turell, J., Hardenbol, J., Heilmann-Clausen, C., Larrasoaña, J.C., Luterbacher, H., Monechi, S., Ortiz, S., Orue-Etxebarria, X., Payros, A., Pujalte, V., Rodríguez-Tovar, F.J., Tori, F., Tosquella, J. & Uchman, A., 2011. The Global Standard Stratotype Section and Point (GSSP) for the base of the Lutetian Stage at the Gorrondatxe section (Spain). Episodes, 34(2), 86-108.
134Murray, J.M., Curry, D., Haynes, J.R. & King, C., 1981. Palaeogene. In Jenkins, D.G. & Murray, J.W. (eds), Stratigraphical atlas of fossil foraminifera. Ellis Horwood, Chichester, British Micropalaeontological Society Series, 228-267.
135Nolf, D. & Steurbaut, E., 1990. Stratigraphie de l'Eocène en Flandre Occidentale et dans les régions limitrophes. Bulletin d'Information des Géologues du Bassin de Paris, 27(3), 9-36.
136Perch-Nielsen, K., 1971. Elektronenmikroskopische Untersuchungen an coccolithen und verwandten Formen aus dem Eozän von Dänemark. Det Kongelige Danske Videnskabernes Selskab Biologiske Skrifter, 18(3), 1-76.
137Perch-Nielsen, K., 1985. Cenozoic calcareous nannofossils. In Bolli, H.M., Saunders, J.B. & Perch-Nielsen, K. (eds), Plankton stratigraphy. Cambridge University Press, Cambridge, Cambridge earth science series, 427-554.
138Rutot, A., 1882. Résolution de la question du Tongrien et du Wemmelien. Création du Système Asschien. Annales de la Société royale malacologique de Belgique, 3e sér., 17, 181-185.
139Rutot, A., 1887. Course géologique à Calevoet, Uccle et St-Gilles. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 1, 118-126.
140Rutot, A., 1890. La constitution de l’étage Paniselien.Procès-Verbaux de la Société belge de Géologie, de Paléontologie et d’Hydrologie(Bruxelles), IV, 1, 252-255.
141Schmitz, B, Heilmann-Clausen, C., King, C., Steurbaut, E., Andreasson, F.P., Corfield, R.M. & Cartlidge, J.E. (1996). Stable isotope and biotic evolution in the North Sea during the early Eocene: the Albaek Hoved section, Denmark. InKnox, R.W.O’B., Corfield, R.M. & Dunay, R.E. (eds), Correlation of the Early Paleogene in Northwest Europe. Geological Society, Special Publication, 101, 275-306.
142Schuler, M., Cavelier, C., Dupuis, C., Steurbaut, E. & Vandenberghe, N. 1992. The Paleogene of the Paris and Belgian Basins. Standard-Stages and regional stratotypes. Cahiers de Micropaléontologie, N.S. 1992, 7(1/2), 29-92.
143Sexton, P.F., Wilson, P.A. & Norris, R.D., 2006. Testing the Cenozoic multisite δ18O and δ13C curves: New monospecific Eocene records from a single locality, Demerara Rise (Ocean Drilling Program Leg 207). Paleoceanography, 21, PA2019, 17 p.
144Shamrock, J.L., 2010. Eocene calcareous nannofossil biostratigraphy, paleoecology and biochronology of ODP Leg 122 hole 762C, eastern Indian Ocean (Exmouth Plateau). University of Nebraska – Lincoln, USA, Dissertations & Theses in Earth and Atmospheric Sciences, Paper42, I-V, 1- 152(http://digitalcommons.unl.edu/geoscidiss/42).
145Shcherbinina, E.A., 2000. Middle Eocene nannofossils and geological events of the northeastern peri-Tethys. GFF, 122, 143-145.
146Siesser, W.G., 1983. Paleogene calcareous nannoplankton biostratigraphy: Mississippi, Alabama and Tennessee. Bulletin of Mississippi Bureau of Geology, 125, 1-61.
147Sluijs, A., Pross, J. & Brinkhuis, H., 2005. From greenhouse to icehouse; organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene. Earth-Science Reviews, 68, 281-315.
148Sluijs, A., Röhl, U., Schouten, S., Brumsack, H.J., Sangiorgi, F., Sinninghe Damsté, J.S. & Brinkhuis, H., 2008a. Arctic late Paleocene–early Eocene paleoenvironments with special emphasis on the Paleocene–Eocene Thermal Maximum (Lomonosov Ridge, Integrated Ocean Drilling Program Expedition 302). Paleoceanography23, PA1S11, 17 p.
149Sluijs, A., Brinkhuis, H., Crouch, E.M., John, C.M., Handley, L., Munsterman, M., Bohaty, S.M., Zachos, J.C., Reichart, G.-J., Schouten, S., Pancost, R.D., Sinninghe Damsté, J.S., Welters, N.L.D., Lotter, A.F. & Dickens, G.R., 2008b. Eustatic variations during the Paleocene-Eocene greenhouse world. Paleoceanography, 23, PA4216, 18 p.
150Sluijs, A., Schouten, S., Donders, T.H., Schoon, P.L., Röhl, U., Reichart, G.-J., Sangiorgi, F., Kim, J.-H., Sinninghe Damsté, J.S. & Brinkhuis, H., 2009. Warm and wet conditions in the Arctic region during Eocene Thermal Maximum 2. Nature Geoscience, 2(11), 1-4.
151Smith, M.E., Carroll, A.R., Scott, J.J. & Singer, B.S., 2014. Early Eocene carbon isotope excursions and landscape destabilization at eccentricity minima: Green River Formation of Wyoming. Earth and Planetary Science Letters, 403, 393-406.
152Smith, T., De Wilde, B. & Steurbaut, E., 2004. Primitive Equoid and Tapiroid mammals: keys for interpreting the Ypresian-Lutetian transition in Belgium. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, 74-Suppl, 165-175.
153Stap, L., Lourens, L.J., Thomas, E., Sluijs, A., Bohaty, S. & Zachos, J.C., 2010.High-resolution deep-sea carbon and oxygen isotope records of Eocene Thermal Maximum 2 and H2. Geology, 38(7), 607-610.
154Steurbaut, E., 1986. Late Middle Eocene to Middle Oligocene calcareous nannoplankton from the Kallo well, some boreholes and exposures in Belgium, and a description of the Ruisbroek Sand Member. Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie, 23(2), 49-83.
155Steurbaut, E., 1988. New Early and Middle Eocene calcareous nannoplankton events and correlations in middle to high latitudes of the northern hemisphere. Newsletters on Stratigraphy, 18(2), 99-115.
156Steurbaut, E., 1990. Calcareous nannoplankton assemblages from the Tertiary in the Knokke borehole. In Laga, P. & Vandenberghe, N. (eds), The Knokke well (11E/138) with a description of the Den Haan (22W/276) and Oostduinkerke (35E/142) wells. Mémoires pour servir à l’Explication des Cartes Géologiques et Minières de la Belgique, 29, 47-62.
157Steurbaut, E., 1991. Ypresian calcareous nannoplankton biostratigraphy and palaeogeography of the Belgian Basin. In Dupuis, C., De Coninck, J. & Steurbaut, E. (eds), The Ypresian stratotype. Bulletin de la Société belge de Géologie, 97(3-4) (1988), 251-285.
158Steurbaut, E., 1998. High-resolution holostratigraphy of Middle Paleocene to Early Eocene strata of Belgium and adjacent areas. Palaeontographica, Abt. A, 247(5-6), 91-156.
159Steurbaut, E., 2006a. Paniselian. In De Geyter et al., Disused Paleogene regional stages from Belgium: Montian, Heersian,Landenian,Paniselian, Bruxellian, Laekenian, Ledian, Wemmelian and Tongrian. In Dejonghe, L. (ed.), Current status of chronostratigraphic units named from Belgiumand adjacent areas.Geologica Belgica, 9(1-2), 203-213.
160Steurbaut, E., 2006b. Ypresian. In Dejonghe, L. (ed.), Current status of chronostratigraphic units named from Belgiumand adjacent areas.Geologica Belgica, 9(1-2), 73-93.
161Steurbaut, E., 2011. New calcareous nannofossil taxa from the Ypresian (Early Eocene) of the North Sea Basin and the Turan Platform in West Kazakhstan. Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 81, 247-277.
162Steurbaut, E. & King, C., 1994. Integrated stratigraphy of the Mont-Panisel borehole section (151E340), Ypresian (Early Eocene) of the Mons Basin, SW Belgium. Bulletinvan de Belgische Vereniging voor Geologie, 102(1-2) (1993), 175-202.
163Steurbaut, E., Magioncalda, R., Dupuis, C., Van Simaeys, S., Roche, E. & Roche, M., 2003. Palynology, paleoenvironments and organic carbon isotope evolution in lagoonal Paleocene/Eocene boundary settings in North Belgium. In Wing, S.L., Gingerich, P.D., Schmitz, B. & Thomas E. (eds), Causes and consequences of Globally Warm Climates in the Early Paleogene. Geological Society of America Special Paper, 369, 291-317.
164Steurbaut, E. & Nolf, D., 1986. Revision of Ypresian stratigraphy of Belgium and Northern France. Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie, 23(4), 115-172.
165Steurbaut, E. & Nolf, D., 1989.The stratotype of the Aalter Sands (Eocene of NW Belgium): stratigraphy and calcareous nannoplankton. Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie, 26(1), 11-28.
166Storme, J.-Y., Steurbaut, E., Devleeshouwer, X., Dupuis, C., Iacumin, P., Rochez, G. & Yans, J., 2014.Integrated bio-chemostratigraphical correlations and climatic evolution across the Danian–Selandian boundary at low latitudes.Palaeogeography, Palaeoclimatology, Palaeoecology, 414, 212-224.
167Thomsen, E., Abrahamsen, N., Heilmann-Clausen, C., King, C. & Nielsen, O.B., 2012. Middle Eocene to earliest Oligocene development in the eastern North Sea Basin: Biostratigraphy, magnetostratigraphy and paleoenvironment of the Kysing-4 borehole, Denmark. Palaeogeography, Palaeoclimatology, Palaeoecology, 350–352, 212–235.
168Vandenberghe, N., Hilgen, F.J. & Speijer, R.P., 2012. The Paleogene period. In Gradstein, F.M., Ogg, J.G., Schmitz, M.D. & Ogg, G.M. (eds), The Geological Time Scale 2012, Vol. 2. Elsevier Science Ltd, Amsterdam,855-921.
169Vandenberghe, N., Laga, P., Steurbaut, E., Hardenbol, J. & Vail, P.R., 1998. Tertiary Sequence Stratigraphy at the southern border of the North Sea Basin in Belgium. In de Graciansky, P.-C., Hardenbol, J., Jacquin, T., & Vail, P.R. (eds), Mesozoic and Cenozoic Sequence Stratigraphy of European Basins. SEPM Special Publication, 60, 119-154.
170Vanhove, H. & De Coninck, J., 1992. Microfossiles à paroi organique de l’Ypresien à Steenhuize-Wijnhuize et Ronse-Waaienberghe. Bulletin de la Société belge de Géologie, 99(3-4) (1990), 355-360.
171Varol, O., 1998. Palaeogene. In Bown, P.R. (ed.), Calcareous nannofossil biostratigraphy. Chapman and Hall, Kluwer Academic, London, British Micropalaeontological Society Publications Series, 200-224.
172Verbeek, J., Steurbaut, E. & Moorkens, T. 1988. Belgium.In Vinken, R. (comp.), The Northwest European Tertiary Basin, I.G.C.P. Project 124. Geologisches Jahrbuch, A, 100, 267-273.
173Westerhold, T., Röhl, U., Donner, B., McCarren, H.K. & Zachos, J.C., 2011. A complete high‐resolution Paleocene benthic stable isotope record for the central Pacific (ODP Site 1209), Paleoceanography, 26, PA2216, 13 p.
174Young, J.R. & Bown, P.R., 1997. Cenozoic calcareous nannoplankton classification. Journal of NannoplanktonResearch, 19(1), 36-47.
175Zachos, J., Pagani, M., Sloan, L., Thomas, E. & Billups, K., 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science, 292, 686-693.
176Zachos, J.C., Dickens, G.R. & Zeebe, R.E., 2008. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature, 451, 279-283.
177Zachos, J.C., McCarren, H., Murphy, B., Röhl, U. & Westerhold, T., 2010. Tempo and scale of late Paleocene and early Eocene carbon isotope cycles: Implications for the origin of hyperthermals. Earth and Planetary Science Letters, 299, 242–249.
178Manuscript received 12.10.2015, accepted in revised form 16.11.2015, available on line 23.11.2015.
