


- Portada
- Volume 19 (2016)
- number 1-2 - Devonian and Carboniferous research: ...
- Serpukhovian coral assemblages from Idmarrach and Tirhela Formations (Adarouch, Morocco)


Vista(s): 2356 (32 ULiège)
Descargar(s): 531 (12 ULiège)

Serpukhovian coral assemblages from Idmarrach and Tirhela Formations (Adarouch, Morocco)

Documento adjunto(s)
Anexidades
Abstract
The Serpukhovian coral assemblages from Idmarrach and Tirhela formations (Adarouch, Morocco) have been studied. They yielded quite diverse assemblages with a total of 32 rugose and 1 tabulate species. The distribution of corals in the sections Idmarrach 1, 2, 3, and 4 and Tirhela 1 and 2 has been established, which include Serpukhovian and Bashkirian rocks. The Serpukhovian assemblages are composed mostly of species that have their higher abundance in the upper Viséan. However, most of the recorded taxa in Adarouch have been already mentioned in Serpukhovian rocks from Britain, Moscow Basin, Urals, Donets Basin and other North African regions such as Tindouf and Béchar. Thus, their stratigraphic range is not expanded. The coral diversity is mainly concentrated in biostromes from the Idmarrach 1 section. However, the high total diversity is due to the combination of favourable depositional settings and a mixture in different beds of several ecological environments, such as coral shoals, protected lagoons and microbial mounds.
Most Serpukhovian species have been recorded in areas from the western Palaeotethys previously mentioned. The total assemblage can be considered as typical for the late Mississippian in the western Palaeotethys. However, a small degree of isolation is registered by the absence in the Serpukhovian from Adarouch of the genera Lonsdaleia, Actinocyathus, Tizraia and Kizilia that have been recorded in other North African basins. That fact may be explained by the incipient rising of some areas as ‘highs’ due to the start of the collision between Gondwana and Laurasia.
Tabla de contenidos
1. Introduction
1The Carboniferous stratigraphic successions in Adarouch (central Morocco) range in age from the middle Viséan up to the Bashkirian (Fig. 1). They are composed of both siliciclastic and carbonate rocks that occur in several faulted blocks (Berkhli, 1999; Vachard et al., 2006) and show excellent exposures that are prolific in diverse fossil groups such as foraminiferans, algae, brachiopods, conodonts, bryozoans, crinoids and corals. The description of the Serpukhovian coral assemblages from that area and their biostratigraphic, palaeogeographic and palaeoecologic implications is the main aim of this paper, which is presented as an homage to Eddy Poty, who studied the Mississippian corals during the last 40 years, providing much valuable data to our knowledge of both rugose corals and Carboniferous rocks.
2The first reference to the presence of rugose corals in the Adarouch area was provided by Termier & Termier (1950), who mentioned the Tizra outcrops at Adarouch and described Dibunophyllum vaughani from that locality. Termier et al. (1975) regarded the Adarouch outcrops to range in age from the Viséan to Serpukhovian. Later, Berkhli (1999) provided detailed stratigraphical and palaeontological data and defined three depositional sequences in the Carboniferous of Adarouch. He studied the Carboniferous stratigraphy of the Tizra Hills and defined the Oued Amhares Formation at the base, the Tizra and Mouarhaz formations in the middle and the Akerchi Formation at the top (Fig. 1). He also described the presence of laterally equivalent rocks to the Akerchi Formation in an eastern fault-bounded sheet, where he defined the Idmarrach and Tirhela formations. The continental siliciclastic rocks of the Migoumess Formation crop out further east (Berkhli & Vachard, 2001, 2002). The coral assemblages from the Tizra, Akerchi and Idmarrach formations were originally described by Said (2005) in an unpublished Ph.D. Thesis and later reported by Said et al. (2007). The aulophyllids from those formations were described by Said & Rodríguez (2008). The taphonomical and palaeoecological features of the assemblages from the Akerchi and Tizra formations and their environmental implications were the aims of two later papers (Said et al., 2010, 2011). Herbig & Aretz (2015) also described some tabulate corals from the Tizra Formation.
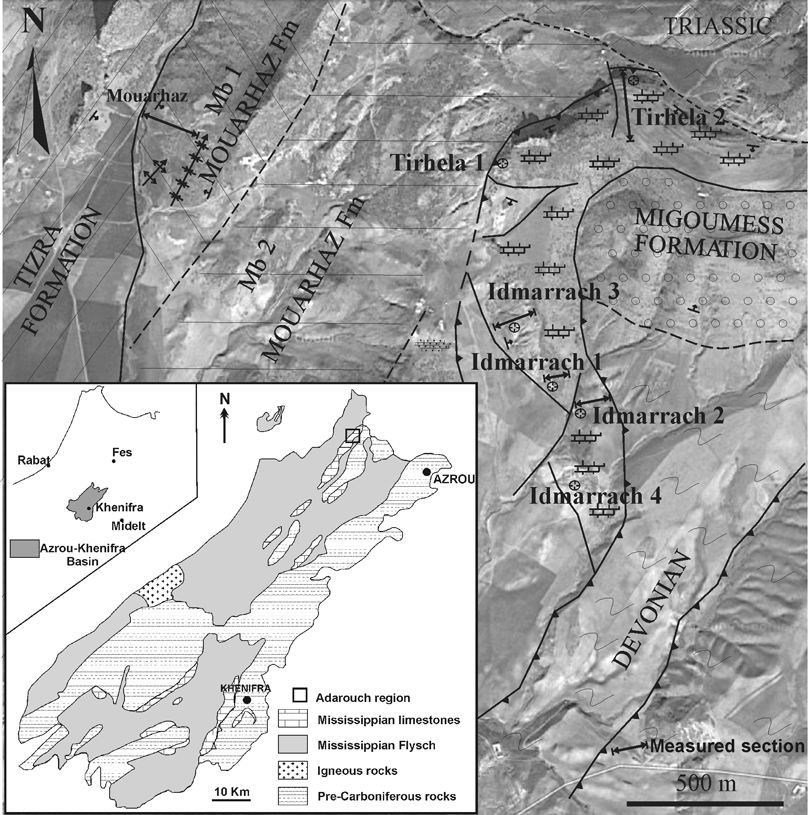
3Figure 1. Location of the Adarouch area (encased box) in the Moroccan Meseta. The photogeological map shows the location of the studied stratigraphic sections. Modified from Cózar et al. (2011).
4The Serpukhovian from the Adarouch region is mainly present in the western block, where the Tirhela and Idmarrach formations represent a time interval that comprises most of the Serpukhovian and the basal Bashkirian (Cózar et al., 2008, 2011). The Akerchi Formation contains also Serpukhovian sandy limestones that basically did not yield corals, apart from the biostrome near the base. The Idmarrach Formation shows predominantly fine-grained siliciclastic rocks (shales) interbedded with limestones in its lower part and well-bedded limestones in its upper part. The lower part of the Tirhela Formation is comparable to the upper part of the Idmarrach Formation, being composed mainly of limestones, but becomes more siliciclastic-rich in the upper part (Cózar et al., 2011).
5The coral assemblages from both formations are conspicuous; they occur mainly in biostromes occurring near the base and at the top of the Tirhela Formation and in a similar location in the upper limestones of the Idmarrach Formation. The lower biostromes are Serpukhovian in age, but the upper biostromes are lower Bashkirian in age (Cózar et al., 2011, Rodríguez et al., submitted). A first study of some assemblages from the Idmarrach Formation was undertaken by Said (2005, unpublished thesis) and later published in a brief compendium (Said et al., 2007). They assigned a late Viséan age to the Idmarrach coral assemblages, based on the data of Berkhli and Vachard (2001, 2002) and on the features of the coral assemblages that lack any indicator of a Serpukhovian age. However, Cózar et al. (2008, 2011) studied the foraminifers and conodonts from the Adarouch area and established that only the base of the Akerchi Formation is uppermost Viséan in age, and that the Idmarrach and Tirhela formations are mostly Serpukhovian, reaching the Bashkirian in their upper beds.
6The coral assemblages from some beds of the Idmarrach Formation (Idmarrach 1 and 2 sections) were listed in Said et al. (2007). New sampling campaigns have been undertaken since then, providing additional specimens from the Idmarrach 2 section and many specimens from a new section (Idmarrach 3), as well as from two isolated outcrops (Idmarrach 4 and Tirhela 1) and from the main section of Tirhela (Tirhela 2). The coral assemblages identified in the lower beds of both Idmarrach and Tirhela formations comprise typical late Viséan species (Said, 2005; Said et al., 2007).
2. Lithostratigraphic units
2.1. Idmarrach Formation
7Corals are abundant in some levels of the Idmarrach Formation. Three partial sections comprising the total stratigraphy of that formation (Idmarrach 1, 2 and 3, Figs 1 & 2) and an isolated outcrop (Idmarrach 4, Figs 1 & 2) have been studied (Cózar et al., 2011). The lowest beds containing corals occur in Idmarrach 3 section, which is close to the base of the Formation. They are well-bedded, dark-grey limestones interbedded with shales, which pass laterally to a microbial mound showing stromatactoid cavities. Corals are common in the bedded limestones and in the flanks of the microbial mound (Figs 3.1-2). Some colonies are in growth position, but they mostly accumulated in debris beds. Up to 19 species belonging to 14 genera have been recorded there (Table 1).
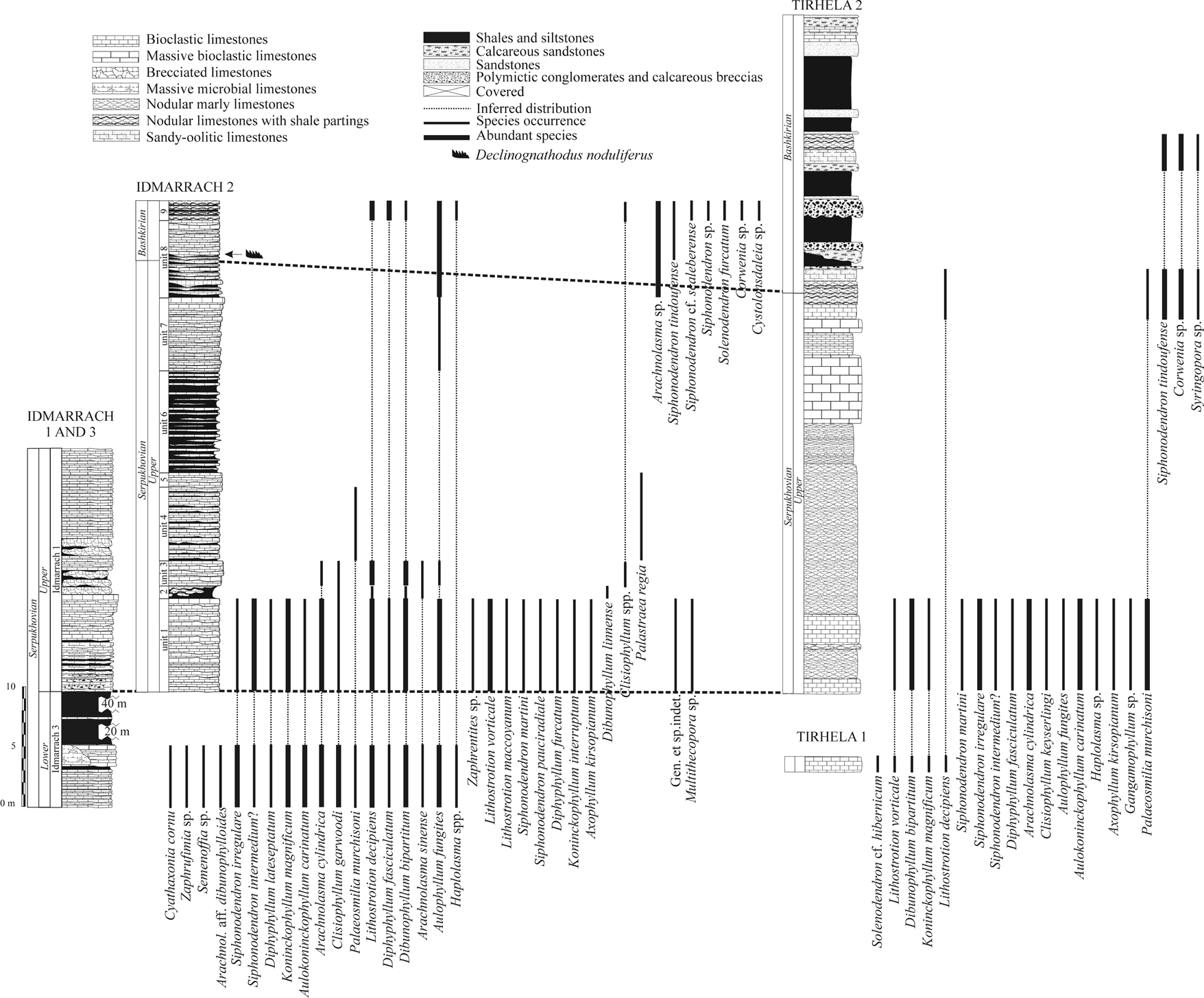
8Figure 2. Distribution of corals in the Idmarrach and Tirhela composite sections (adapted from Cózar et al., 2011).

9Figure 3. Serpukhovian outcrops that yielded corals in Idmarrach and Tirhela Formations. 1: Partial view of Idmarrach 1 section. 2: Detail of the beds containing corals in Idmarrach 1 section. 3: Detail of basal part of a biostrome, in the Idmarrach 2 section, composed of a pavement of gigantoproductid shells. Some coral colonies (dotted lines) are visible growing on the gigantoproductids. 4: Detail of a Siphonodendron colony in the biostromal beds in the Idmarrach 2 section (unit 1). 5: General view of the Idmarrach 3 section; up left (NW), massive microbial mound; right, well-bedded limestones. 6: Detail of the well-bedded limestones yielding corals. 7: general view of the Tirhela 2 section with steeply dipping beds to the south. 8: Detail of the beds yielding corals at the base of the Tirhela 2 section.
|
Species |
IDM1 |
IDM2 |
IDM3 |
IDM4 |
TIR1 |
TIR2 |
|
Haplolasma lamelliferum |
X |
X |
||||
|
Haplolasma aff. parvicarinatum |
O |
|||||
|
Aulophyllum fungites |
XO |
X |
X |
|||
|
Clisiophyllum garwoodi |
X |
X |
||||
|
Clisiophyllum keyserlingi |
X |
X |
X |
|||
|
Clisiophyllum sp. |
X |
|||||
|
Dibunophyllum bipartitum |
X |
XO |
X |
X |
X |
X |
|
Dibunophyllum linnense |
X |
|||||
|
Arachnolasma sinense |
X |
X |
||||
|
Arachnolasma cylindrica |
X |
X |
X |
|||
|
Arachnolasma sp. aff. dibunophylloides |
X |
|||||
|
Corwenia sp. |
O |
O |
||||
|
Koninckophyllum interruptum |
XO |
|||||
|
Koninckophyllum magnificum |
X |
X |
X |
X |
X |
|
|
Palaeosmilia murchisoni |
X |
X |
XO |
|||
|
Palastraea regia |
X |
|||||
|
Aulokoninckophyllum carinatum |
X |
X |
X |
|||
|
Semenoffia sp. |
X |
|||||
|
Lithostrotion decipiens |
X |
XO |
X |
X |
X |
O |
|
Lithostrotion vorticale |
X |
X |
X |
|||
|
Lithostrotion maccoyanum |
X |
|||||
|
Siphonodendron martini |
X |
X |
X |
|||
|
Siphonodendron pauciradiale |
X |
|||||
|
Siphonodendron irregulare |
X |
X |
X |
X |
||
|
Siphonodendron intermedium? |
X |
X |
X |
|||
|
Siphonodendron tindoufense |
O |
O |
||||
|
Siphonodendron cf. scaleberense |
O |
|||||
|
Diphyphyllum lateseptatum |
X |
X |
X |
|||
|
Diphyphyllum furcatum |
X |
X |
||||
|
Diphyphyllum fasciculatum |
XO |
X |
X |
X |
||
|
Solenodendron cf. hibernicum |
X |
|||||
|
Solenodendron furcatum |
O |
|||||
|
Axophyllum kirsopianum |
X |
X |
||||
|
Gangamophyllum sp. |
X |
|||||
|
Cystolonsdaleia? sp. |
O |
|||||
|
Cyathaxonia cornu |
X |
|||||
|
Zaphrentites sp. |
X |
|||||
|
Zaphrufimia sp. |
X |
|||||
|
Gen et sp. indet. |
X |
|||||
|
Multithecopora sp. |
X |
X |
||||
|
Syringopora sp. |
O |
|||||
|
Total Serpukhovian rugose taxa |
3 |
25 |
19 |
7 |
5 |
16 |
10Table 1. Species occurrence in the Idmarrach and Tirhela formations. X = Occurrence in Serpukhovian rocks. O = Occurrence in Bashkirian rocks. IDM1 = Idmarrach 1, IDM2 = Idmarrach 2 IDM3 = Idmarrach 3, IDM4 = Idmarrach 4, TIR1 = Tirhela 1, TIR2 = Tirhela 2.
11The Idmarrach 1 and Idmarrach 2 sections occur about 60 metres above Idmarrach 3. The Idmarrach 1 and 2 sections are composed of well-bedded grey limestones which near the base constitute biostromes that contain abundant colonial and solitary corals and gigantoproductid brachiopods (Figs 3.2-3). Most colonies are in growth position or only slightly tilted (Fig. 2.4). Corals occur also in the upper beds, but they are less common, except in the top beds of the Idmarrach 2 section, where another biostrome occurs (Idmarrach units 8 and 9, Fig. 2). That upper biostrome is Bashkirian in age and its coral assemblage is described in Rodríguez et al. (submitted). An isolated outcrop eastward from the previously mentioned sections has been called Idmarrach 4. The outcrop does not allow the measurement of a detailed log, but some additional corals have been collected there. Its precise stratigraphic position is unknown, but the lithologies and the coral assemblage indicate a possible correlation with Idmarrach 3 section.
12The assemblages from Idmarrach 1 and 2 were described by Said (2005) and listed by Said et al. (2007). Additional sampling and revision of some previous identifications in these sections, as well as sampling in Idmarrach 3 and 4 sections, provide new data on those assemblages. A total of 30 rugosan species belonging to 17 genera and one tabulate species have been recorded in the Serpukhovian part of all the Idmarrach sections. 25 species have been recorded from the Idmarrach 1 and 2 sections of which 18 occur in the basal biostromes. Details of comparative assemblages are illustrated in Figure 2 and Table 1.
2.2. Tirhela Formation
13The Tirhela Formation comprises about 40 metres of well-bedded and massive limestones that progressively pass upward to the siliciclastic deposits of the Migoumess Formation (Fig. 3.7). Below those rocks, there are some shales that are mainly covered. An isolated small outcrop about 50 metres below the main limestones, composed of sandy limestones, yielded some corals. The assemblage from that outcrop has been labelled as Tirhela 1. The main outcrop of the Tirhela Formation (Tirhela 2 section) shows a diverse assemblage of corals located in a biostrome near the base (Figs 2, 3.8). The corals are both, in growth position and reworked. The beds below and above the biostrome are rich in gigantoproductid brachiopods. The biostrome is regarded as equivalent to that near the base of Idmarrach 1 and 2. No corals have been collected in other beds of the Tirhela section, except in the transitional beds to the Migoumess Formation, where several biostromal beds yielded rugose corals of Bashkirian age, which have been described by Rodríguez et al. (submitted).
14A total of 18 rugose species belonging to 15 genera have been recorded in the Serpukhovian part of the Tirhela sections. The biostrome at the base yielded 85% of the recorded species. In spite of the coral abundance in the Bashkirian biostromes at the top of the section, the diversity there is very low; only 4 rugose species have been recorded and more than 90% of the collected specimens belong to only 2 species, Siphonodendron tindoufense Rodríguez et al., 2013b and Corwenia sp. (Rodríguez et al., submitted). The tabulate genus Syringopora is only recorded in Tirhela in the Bashkirian (Table 1).
15The complete assemblage of the Tizra Formation is shown in Figure 2 and listed in Table 1.
3. Coral assemblages
16More than 300 coral specimens have been collected in the Serpukhovian interval of the studied sections, 25 specimens from Idmarrach 1, 120 specimens from Idmarrach 2, 70 specimens from Idmarrach 3, 15 specimens from Idmarrach 4, 5 specimens not located in a section from Idmarrach area, 10 specimens from Tirhela 1 and 60 specimens from Tirhela 2. The specimens have been studied by means of about 400 thin sections.
1733 species (32 rugosans and 1 tabulate) have been identified in the Serpukhovian rocks (Fig. 2 and Table 1). The complete assemblages are shown in figures 4 to 7, following the systematic order proposed by Hill (1981).
18The oldest assemblages are located in the Idmarrach 3 and Tirhela 1 sections that have been dated as Pendleian (early Serpukhovian) with foraminifers and conodonts (Cózar et al., 2011). The corals are randomly distributed around a microbial mound. Most specimens are located in wackestones surrounding the mound, but some specimens are also located in packstones at the flank beds. They are mostly reworked and located with their larger dimensions parallel to the surface of the strata. However, some specimens are entire or almost, indicating a short transport. The assemblage is dominated by solitary corals, mostly belonging to the family Aulophyllidae. The most common species are Aulokoninckophyllum carinatum (Carruthers, 1909) (Figs 4.6a-c), Dibunophyllum bipartitum (McCoy, 1849) and several species of the genus Arachnolasma Grabau, 1922 (Figs 5.8-10). Three additional species are represented by at least three specimens: Clisiophyllum garwoodi (Salée, 1913), Aulophyllum fungites (Fleming, 1828) (Fig. 5.2) and Koninckophyllum magnificum (Thomson & Nicholson, 1876). All other solitary species (Table 1) are scarce. Colonial corals are less common but their diversity is not low. Six species have been recorded, all of them belonging to the family Lithostrotionidae. Only two of them are represented by three or more specimens in our collections, Lithostrotion decipiens (McCoy, 1849) and Siphonodendron irregulare (Phillips, 1836). Some solitary undissepimented corals have been recorded in marly beds interbedded with the limestones: Cyathaxonia cornu Michelin, 1847 (Fig. 6.9) and Zaphrufimia sp. (Fig. 6.8). The assemblage from Tirhela 1 is less diverse, composed only of 5 species and showing 3 species in common with Idmarrach 3. Only Lithostrotion vorticale (Parkinson, 1808) (Fig. 7.2) and Solenodendron cf. hibernicum (Clarke, 1966) (Fig. 6.4), represented by single specimens, have been not recorded in Idmarrach 3.

19Figure 4. Solitary and colonial rugose corals of the Idmarrach and Tirhela sections 1: Koninckophyllum interruptum Thomson & Nicholson, 1876, specimen IDM2/1-17, 1a: transverse section, 1b: longitudinal section. 2: Koninckophyllum magnificum (Thomson & Nicholson, 1876), specimen IDM2/1-30, 2a: transverse section, 2b: longitudinal section. 3: Palaeosmilia murchisoni Milne-Edwards & Haime, 1848, specimen IDM3/1-2, transverse section. 4: Palastraea regia (Phillips, 1836), specimen IDM2/4-2, transverse section. 5-6: Aulokoninckophyllum carinatum (Carruthers, 1909) 5: specimen IDM2/1-54, 5a: transverse section, 5b: longitudinal section. 6: specimen IDM3/1-15, 6a: transverse section, 6b: longitudinal section, 6c: enlarged view of 6a to show carinae in dissepimentarium. 7: Semenoffia sp., specimen IDM3/1-25, 7a: transverse section, 7b: longitudinal section. Scale bar 1 cm, same for all pictures except 6c.

20Figure 5. Solitary rugose corals of the Idmarrach and Tirhela sections 1: Haplolasma lamelliferum Semenoff-Tian-Chansky, 1974, specimen TIR2/1-15, 1a: transverse section. 1b: longitudinal section. 2: Aulophyllum fungites (Fleming, 1828), specimen IDM3/1-8, transverse section. 3: Clisiophyllum garwoodi (Salée, 1913), specimen IDM2/1-53, transverse section. 4: Clisiophyllum sp. specimen IDM2/1-56, transverse section. 5: Clisiophyllum keyserlingi (McCoy, 1849) specimen TIR2/1-3, 5a: transverse section. 5b: longitudinal section. 6: Dibunophyllum bipartitum (McCoy, 1849), specimen IDM2/2-15, transverse section. 7: Dibunophyllum linnense Hill, 1940, specimen IDM2/2-9, transverse section. 8: Arachnolasma sinense (Yabe & Hayasaka, 1920), specimen IDM3/1-34, transverse section. 9: Arachnolasma cylindrica Yü, 1933), specimen IDM3/1-8, transverse section. 10: Arachnolasma sp.-cf. dibunophylloides (Gorsky in Volkova, 1941), IDM3/1-35, 10a: transverse section. 10b: longitudinal section. Scale bar 1 cm, same for all pictures.
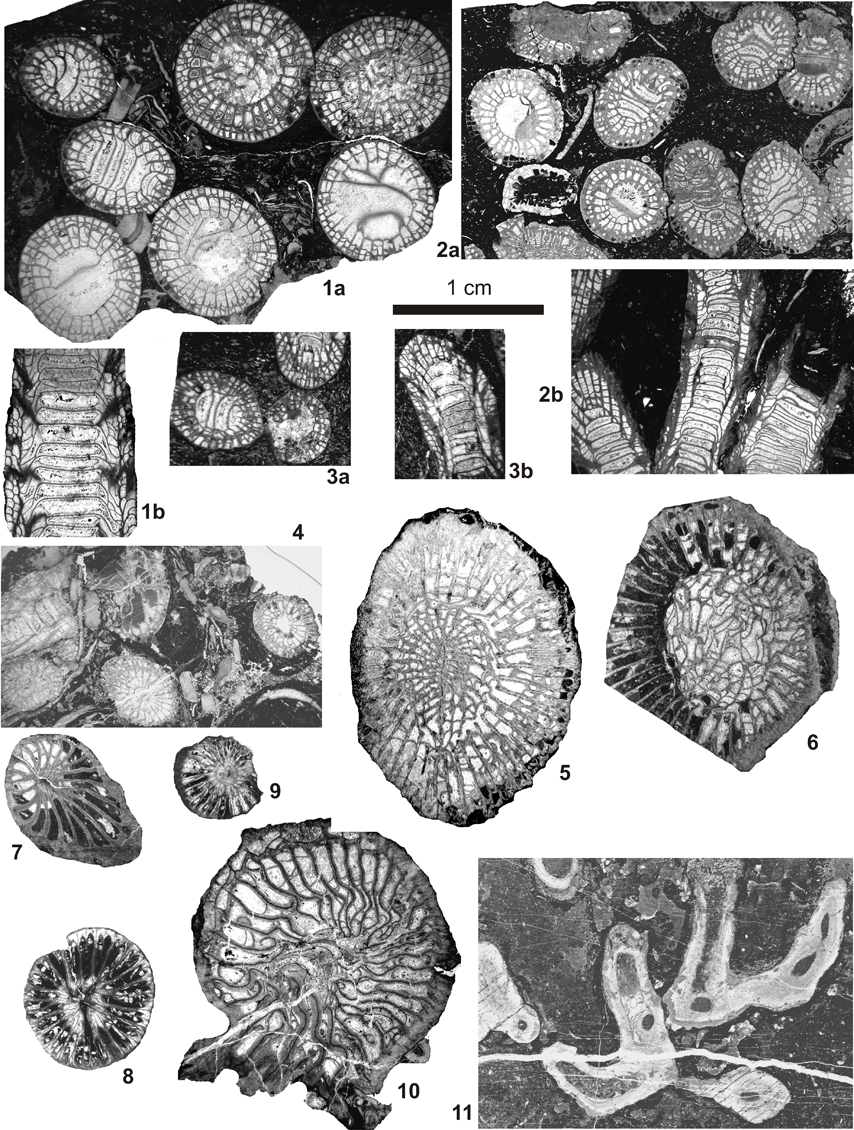
21Figure 6. Colonial and solitary corals (rugose and tabulate) of the Idmarrach and Tirhela sections. 1: Diphyphyllum lateseptatum (McCoy, 1849) specimen IDM2/1-13, 1a: transverse section. 1b: longitudinal section. 2: Diphyphyllum furcatum Hill, 1940, specimen IDM2/1-12, 2a: transverse section, 2b: longitudinal section. 3: Diphyphyllum fasciculatum (Fleming, 1828), specimen IDM2/1-24, 3a: transverse section, 3b: longitudinal section. 4: Solenodendron cf. hibernicum (Clarke, 1966) specimen TIR1/1-2, transverse and longitudinal sections. 5: Axophyllum kirsopianum (Thomson, 1880) specimen TIR2/1-19, transverse section. 6: Gangamophyllum sp., specimen TIR2/1-32, transverse section. 7: Zaphrentites sp., specimen IDM2/1-27, transverse section. 8: Zaphrufimia sp., specimen IDM3/1-50, transverse section. 9: Cyathaxonia cornu Michelin, 1846, specimen IDM3/1-51, transverse section. 10: Gen et sp. indet., specimen TIR2/1-34, transverse section. 11. Multithecopora sp., specimen IDM4/10, oblique section. Scale bar 1 cm, same for all pictures.
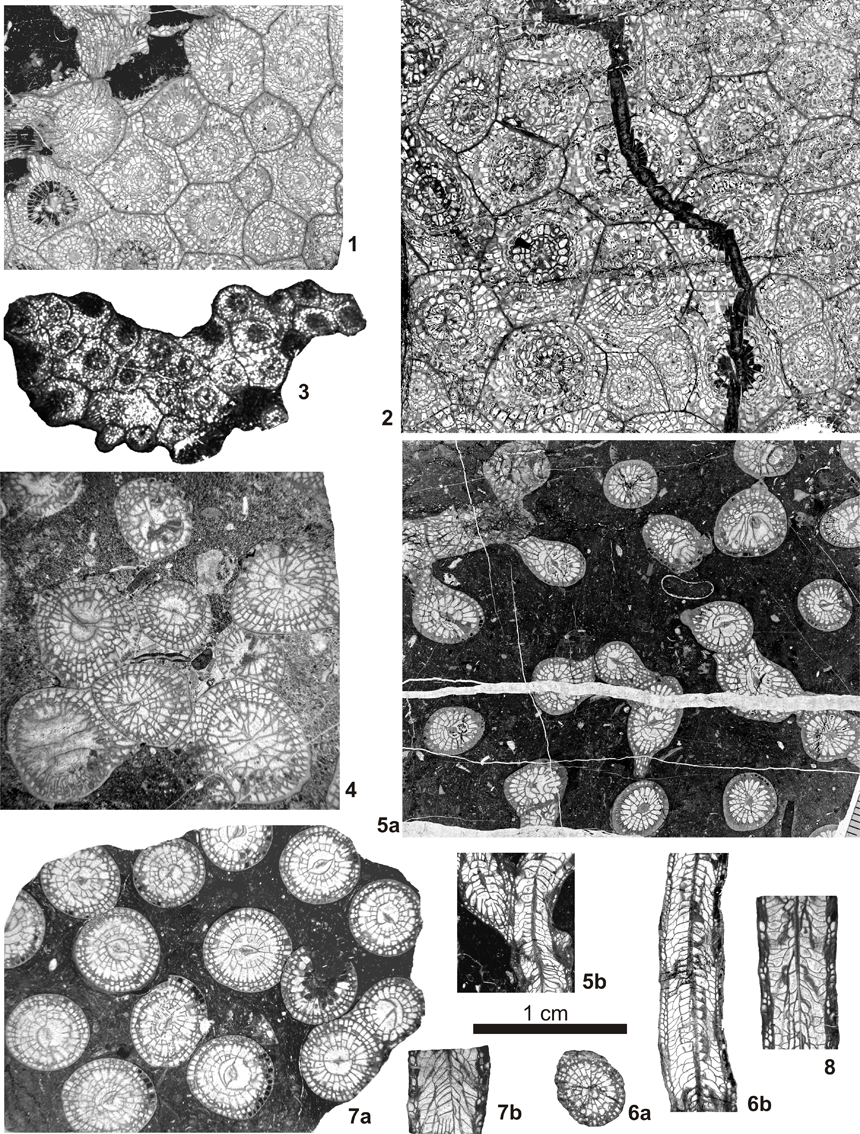
22Figure 7. Colonial rugose corals of the Idmarrach and Tirhela sections. 1: Lithostrotion decipiens (McCoy, 1849), specimen IDM2/1-5, transverse section. 2: Lithostrotion vorticale (Parkinson, 1808), specimen IDM2/1-36, transverse section. 3: Lithostrotion maccoyanum (Milne-Edwards & Haime, 1851), specimen IDM2/1-25, transverse section. 4: Siphonodendron martini (Milne-Edwards & Haime, 1851), specimen TIR2/1-27, transverse section. 5: Siphonodendron pauciradiale (McCoy, 1844), specimen IDM2/2-11, 5a: transverse section, 5b: longitudinal section. 6: Siphonodendron irregulare (Phillips, 1836) specimen IDM2/1-2, 6a: transverse section, 6b: longitudinal section. 7-8: Siphonodendron intermedium? Poty, 1981: 7: specimen IDM2/1-7, 7a: transverse section, 7b: longitudinal section, 8: specimen IDM2/1-10, longitudinal section. Scale bar 1 cm, same for all pictures.
23The assemblages from the unit 1 of the sections Idmarrach 1 and 2 occur in biostromal beds. The age of those biostromes, as well as those in the base of Tirhela 2 is Arnsbergian (late Serpukhovian) (Cózar et al., 2011). They are composed mainly of gigantoproductid brachiopods and colonial corals, but solitary dissepimented corals are common too. The outcrop at Idmarrach 2 shows a wide upper bedding surface where the colonies cover most of the exposed area. They are commonly cut in transverse sections. When lateral parts of the bed are seen in the eroded surface, the colonies show corallites oriented upwards (Fig. 3.4). If erosion of the strata is deeper, a pavement composed of gigantoproductid shells can be observed (Fig. 3.3). Some coral colonies are visible growing on the gigantoproductids. As most colonies seem to be in growth position, it is regarded as an autobiostrome (Kershaw, 1994).
24The coral assemblage is dominated by lithostrotionids (Table 1 and Fig. 2). The most abundant species are Siphonodendron intermedium? Poty, 1981 (Figs 7.7-8) and Lithostrotion decipiens (McCoy, 1849) (Fig. 7.1). The identification of the first species is given with a question mark, because the specimens show a conspicuous uniform size of corallites (tabularium 4.2-5.2 mm) and number of septa (19-23), that differs from the holotype by the presence of longer major septa, some of which reach the columella and a more complex tabularium. Other colonial species occurring in the same beds are Lithostrotion vorticale (Parkinson, 1808) (Fig. 7.2), Siphonodendron martini (Milne-Edwards & Haime, 1851) (Fig. 7.4), Siphonodendron pauciradiale (McCoy, 1844) (Fig. 7.5), Siphonodendron irregulare (Phillips, 1836) (Fig. 7.6a-b), Diphyphyllum lateseptatum (McCoy, 1849) (Fig. 6.1a-b), Diphyphyllum furcatum Hill, 1940 (Fig. 6.2a-b) and Diphyphyllum fasciculatum (Fleming, 1828) (Fig. 6.3a-b).
25Solitary dissepimented corals are also common and diverse in the biostromes of Idmarrach 1 and 2, and in interbedded marls, but they are usually reworked and rarely complete. Most species are represented by one or two specimens, only being quite abundant Dibunophyllum bipartitum (McCoy, 1849) (Fig. 4.6), Koninckophyllum interruptum Thomson & Nicholson, 1876 (Fig. 4.1a-b) and Koninckophyllum magnificum (Thomson & Nicholson, 1876) (Fig. 4.2a-b). A single specimen of the genus Zaphrentites Hudson, 1941 (Fig. 6.7) has been recorded in the interbedded marls.
26Corals in the upper beds of the Idmarrach 2 section are less abundant and diverse (Fig. 2). Only one fragmented colony of Palastraea regia (Phillips, 1836) (Fig. 4.4) and some specimens of Aulophyllum fungites (Fleming, 1828) and Palaeosmilia murchisoni Milne-Edwards & Haime, 1848 have been recorded.
27The biostromes at the base of the section Tirhela 2 show also a high abundance of colonial corals (Table 1, Fig. 2). In this case, part of the colonies are reworked. The assemblage is quite similar in its composition to that of Idmarrach 1 and 2, but the most abundant colonial coral species in Tirhela 2 is Siphonodendron irregulare (Phillips, 1836) and Dibunophyllum bipartitum (McCoy, 1849) and Axophyllum kirsopianum (Thomson, 1880) (Fig. 6.5) of the solitary rugosans. The diversity is somewhat lower than in Idmarrach 1 and 2, but in Tirhela occur some species that have been not recorded in Idmarrach, such as Gangamophyllum sp. (Fig. 6.6) and Haplolasma-lamelliferum Semenoff-Tian-Chansky, 1974, (Fig. 3.1a-b). With the exception of the biostromes at the base and at the top (the latter being Bashkirian in age) no other corals have been recorded in Tirhela 2 section.
4. Biostratigraphic and Palaeogeographic connotations
28The coral assemblages from Idmarrach and Tirhela formations show several peculiarities regarding their stratigraphical location. The assemblages are dominated by species whose distribution in the northern areas of the western Palaeotethys is mainly upper Viséan. Comparing with biostratigraphical charts from Britain (Mitchell, 1989), Belgium and northern France (Poty, 1981, 1985; Poty et al., 2006; Denayer et al., 2011) and south-west Spain (Rodríguez & Somerville, 2007), most species occur in the upper Viséan and do not reach the Serpukhovian in those regions. However, the main reason for that is because marine Serpukhovian rocks are scarce in the north-western Palaeotethys and the increasing tectonic movements produce mainly terrigenous facies that are not favourable for the development of corals (Fedorowski, 1981). In spite of that, Hill (1938-1941) mentions the persistence in the Namurian (Serpukhovian) from Britain of Aulophyllum fungites-(Fleming, 1828), Clisiophyllum keyserlingi McCoy, 1849, Dibunophyllum bipartitum-(McCoy, 1849), Koninckophyllum magnificum (Thomson & Nicholson, 1876), K. interruptum (Thomson & Nicholson, 1876), Diphyphyllum lateseptatum-(McCoy, 1849), Lithostrotion decipiens(McCoy, 1849), L. maccoyanum Milne-Edwards & Haime, 1851, Siphonodendron martini (Milne-Edwards & Haime, 1851) (with a question mark), S. pauciradiale-(McCoy, 1844), Palastraea regia (Phillips, 1836), and other species that do not occur in the Serpukhovian from Adarouch, belonging to the genera Lonsdaleia McCoy, 1849 and Actinocyathus D’Orbigny, 1849. She also indicates the presence of Cyathaxonia cornu-Michelin, 1847 and several species of Zaphrentis Rafinesque & Clifford, 1820 that now are included in the genera Zaphrentites Hudson, 1941 and Zaphrufimia Fedorowski, 2012.
29The analysis of coral assemblages in other regions of north-central Palaeotethys shows that some species recorded in the Serpukhovian from Adarouch were already identified in the Serpukhovian from Poland (Fedorowski, 1981; Khoa, 1977), Donets Basin (Vassiljuk, 1960), Voronezh anteclise (Kozyreva, 1973), Moscow Basin (Dobrolyubova, 1952, 1958, 1970), Urals (Degtyarev, 1973; Kachanov, 1965, 1975; Sayutina, 1973), Novaya Zemlya (Gorsky, 1938, 1951) and Kazakhstan (Bykova, 1966). Thus, all the genera (or their equivalents) and most species recorded in the Serpukhovian of Adarouch have been previously cited in the Serpukhovian from the north-central Palaeotethys region.
30 Studies in other areas from North Africa (Fig. 8) indicate that many species reach upper levels in different basins. Thus, Semenoff-Tian-Chansky (1974, 1985) demonstrates the presence of Dibunophyllum bipartitum-(McCoy, 1849), Palaeosmilia murchisoni Milne-Edwards & Haime, 1848, Koninckophyllum interruptum (Thomson & Nicholson, 1876), Aulokoninckophyllum carinatum (Carruthers, 1909), Clisiophyllum keyserlingi McCoy, 1849, Gangamophyllum Gorsky, 1938, etc. in the Serpukhovian from the Béchar Basin. However, none of the lithostrotionids that occur in Adarouch reach the Serpukhovian in Béchar, except perhaps Lithostrotion maccoyanum Milne-Edwards & Haime, 1851 and Diphyphyllum lateseptatum-(McCoy, 1849).
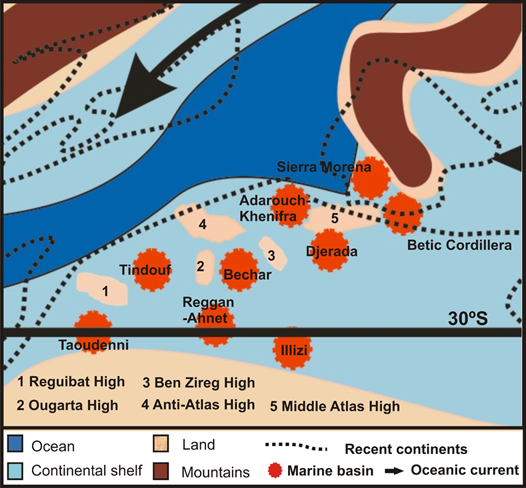
31Figure 8. Palaeogeography of northwestern Africa and southwestern Europe during the upper Mississippian, showing the main marine basins (modified from Coronado et al., 2015).
32Aretz (2011) adds the presence of Siphonophyllia samsonensis (Salée, 1913), Haplolasma paraarciferum Aretz, 2011, Clisiophyllum keyserlingi McCoy, 1849, Arachnolasma sp. and Lithostrotion decipiens (McCoy, 1849), in the Serpukhovian of the Illizi Basin and Palaeosmilia murchisoni Milne Edwards & Haime, 1848, Dibunophyllum bipartitum (McCoy 1849), Koninckophyllum magnificum Thomson & Nicholson, 1876, K. interruptum Thomson & Nicholson, 1876 and Lithostrotion vorticale (Parkinson, 1808) in the Serpukhovian of the Reggane Basin.
33Rodríguez et al.-(2013a) described coral assemblages from the north flank of the Tindouf Basin (south Morocco). Most coral species occur there later than in north-western Palaeotethys; the Serpukhovian assemblages contain many species in common with those from Adarouch: Arachnolasma cylindrica Yü, 1933, Palastraea regia (Phillips, 1836), Palaeosmilia murchisoni Milne Edwards & Haime, 1848, Koninckophyllum interruptum Thomson & Nicholson, 1876, Aulophyllum fungites Fleming, 1828, Dibunophyllum bipartitum (McCoy, 1849), Diphyphyllum furcatum Hill, 1940, D. lateseptatum-(McCoy, 1849), Lithostrotion decipiens (McCoy, 1849), L. vorticale (Parkinson, 1808), Siphonodendron irregulare (Phillips, 1836), S. pauciradiale (McCoy, 1849), S. martini (Milne-Edwards & Haime, 1851). However, some genera recorded in Tindouf are not present in the Serpukhovian from Adarouch, such as Lonsdaleia McCoy, 1849, Actinocyathus D’Orbigny, 1849, Kizilia Degtyarev, 1965 and Tizraia Said & Rodríguez, 2007 (Rodríguez et al., 2013a; Somerville et al., 2013). The last named genus occurs in the upper Viséan from the Tizra and Akerchi formations; Lonsdaleia McCoy, 1849, Actinocyathus D’Orbigny, 1849 and Kizilia Degtyarev, 1965 are not present in Adarouch (Said et al., 2013).
34No Serpukhovian rocks yielding corals have been recorded in the Jerada Basin (Aretz, 2010a), but upper Viséan assemblages show high similarities to those from Adarouch. A similar situation occurs in the Khenifra region, south of Adarouch and part of the same Azrou-Khenifra Basin, where only Asbian and Brigantian assemblages have been recorded (Aretz & Herbig, 2010; Somerville et al., 2012; Rodríguez et al., 2012; Said et al., 2013). As to be expected, those assemblages show high similarities with those from Adarouch.
35All data provided by the Serpukhovian assemblages in the western Palaeotethys indicate that no dramatic changes occurred in the transition from the Viséan to the Serpukhovian. Corals disappear only in those areas where the Variscan Orogeny affected the environment, reducing the epicontinental seas or providing large amounts of terrigenous material. Moreover, most species are very conservative, remaining unchanged during a long period that may be estimated between 12 and 15 million years. As stated in recent research of areas where the shallow-water carbonate platforms were preserved up to the Bashkirian (Kossovaya, 1996; Fedorowski et al., 2012; Rodríguez et al., 2013b, Cózar et al., 2014), some species such as Palaeosmilia murchisoni Milne-Edwards & Haime, 1848, Palastraea regia (Phillips, 1836), Dibunophyllum bipartitum (McCoy, 1849), remain unchanged and new species from other genera (Actinocyathus D’Orbigny, 1849, Arachnolasma-Grabau, 1922, Gangamophyllum-Gorsky, 1938, Axophyllum Milne-Edwards & Haime, 1851, Lithostrotion Fleming, 1828, Diphyphyllum Lonsdale, 1845, Siphonodendron-McCoy, 1849, etc.) still remain. Adarouch is an excellent example of that, because some of the species recorded in the Serpukhovian reach the Bashkirian (Rodríguez et al., submitted).
36In the same way, the high degree of similarity of the Serpukhovian coral assemblages from Adarouch to those from the northern Palaeotethys and to those from the Saharan Basins indicates that the routes for migration of corals (mainly by means of currents transporting their planulae) were still open during the Serpukhovian (Fig. 8). The massifs and highs that were beginning to develop in North Africa such as the Ougarta Shield, Reguibat Shield and multiple blocks in the Moroccan Meseta (Huvelin, 1967, 1969; Bouabdelli, 1989) were only partial barriers, which impeded the migration of some species/genera such as Lonsdaleia McCoy, 1849, Actinocyathus D’Orbigny, 1849, Kizilia Degtyarev, 1965, etc., but they represent a small percentage of the total assemblages, which show a very high degree of similarity and is typical for the Palaeotethys.
5. Palaeoecological inferences
37The assemblages from Idmarrach and Tirhela occur mainly in some concentrations that may be regarded as biostromes. The biostrome located at the base of Idmarrach 1 and 2 sections show most colonies in growth position, thus constituting an autobiostrome (Kershaw, 1994), but most solitary corals are reworked. On the other hand, some beds above it and the biostrome located at approximately the same stratigraphic level at the base of Tirhela, show all solitary corals and most colonies tilted or reworked (parabiostromes). Some beds in Idmarrach 3 section show also tilted and reworked colonies, but some other beds contain coral/brachiopods debris. Gigantoproductid brachiopods are abundant, both in the biostromal beds, and in beds above and below the biostromes. They are in growth position in some beds, mainly inverted in some other beds and reworked and fragmented in other instances. The rocks are well-bedded limestones with conspicuous bedding planes, which indicate notable interruptions in the sedimentation. All these features indicate a shallow-water platform that was colonized repeatedly by corals and gigantoproductid brachiopods. The periodic occurrence of storms produced the reworking of the coral and brachiopod assemblages in some instances or their preservation, when consolidation of the beds resulted before a storm occurred.
38The coral assemblages from the Serpukhovian biostromes can be compared to the Rugose Coral Association (RCA) 4 of Somerville & Rodríguez (2007). In spite of the age difference (the RCAs were defined based on upper Viséan assemblages), it is notable for the coincidence in terms of taxonomic diversity, development of growth form types, transport index, associated biotas and lithofacies. The main difference lies in the absence of massive cerioid colonies of Actinocyathus D’Orbigny, 1849, whose ecological niche is substituted by Lithostrotion Fleming, 1828 in Idmarrach and Tirhela. The biostromal assemblage can also be regarded as the C1-C3 type of Aretz (2010b), because some areas of the biostromes show the dominance of fasciculate colonies in growth position, but in other areas, there are important amounts of coral debris. Some intercalated beds of marly limestones that yielded common solitary and scarce colonial corals are included in the RCA 1 (Somerville & Rodríguez, 2007), with the most common genera, the same as in the upper Viséan assemblages: Aulophyllum Milne-Edwards & Haime, 1850 and Dibunophyllum Thomson & Nicholson, 1876. More sporadically, there are beds containing less abundant corals, mainly belonging to Siphonodendron McCoy, 1849, but also to solitary genera, such as Axophyllum Milne-Edwards & Haime, 1851, Arachnolasma Grabau, 1922, Clisiophyllum Dana, 1846 and Dibunophyllum Thomson & Nicholson, 1876. Those beds are regarded as RCA5. The combination of these three types of associations, allow the interpretation of the environment as dominated by coral shoals (RCA 4) that protect lagoons (RCA5 and RCA1) (Somerville & Rodríguez, 2007, fig. 3).
39At Idmarrach 3 section, a microbial mound is identified. The beds yielding corals are mainly located in the flanks of the mound and in well-bedded limestones with coral debris near the mound. They show a high diversity and are dominated by Siphonodendron irregulare, Lithostrotion decipiens, Koninckophyllum magnificum, Aulokoninckophyllum carinatum and Dibunophyllum bipartitum. Some undissepimented corals occur also there. They represent a mixture of corals developed near and on the mound flanks (RCA 4-6), small corals living in the deeper facies (RCA8) near the mounds, and coral debris removed from shallower zones (RCA5).
6. Conclusions
40The Serpukhovian assemblages from Idmarrach and Tirhela formations yielded quite diverse assemblages; 29 species of rugose corals in the four Idmarrach sections combined and 18 species in both Tirhela sections. A total of 32 rugose and 1 tabulate species have been recorded.
41Those Serpukhovian assemblages are composed mostly of species that have their higher abundance in the upper Viséan. However, most recorded taxa have been already documented in Serpukhovian rocks from different areas in the central and western Palaeotethys and their stratigraphic range is not expanded.
42The high diversity is due to the combination of favourable environments and a mixture of several ecological settings, such as coral shoals, protected lagoons and microbial mounds.
43Most species have been recorded in other areas from the western Palaeotethys. The total assemblage can be considered as typical for the late Mississippian from the Palaeotethys. However, a small degree of isolation is registered by the absence of some genera that have been recorded in other North African basins. That fact may be explained by the incipient rising of some massifs/highs due to the start of collision between Gondwana and Laurasia (Blakey, 2007, 2008).
7. Acknowledgements
44The present research has been carried out with the funds provided by the Research project CGL2012-30922BTE of the Spanish Ministry of Research and Innovation. This article is a contribution to the IGCP 596. The authors thank the cooperation of the Department of Mines and Development of the Ministère de l’Energie et des Mines of Morocco. The thin sections used for this study have been prepared by Isabel Díaz Mejías and some of the pictures were taken by Carlos Alonso. The authors wish to thank Jerzy Fedorowski, Olga Kossovaya and Xiang-dong Wang for their constructive comments which helped to improve the paper.
8. References
45Aretz, M., 2010a. Rugose corals from the upper Viséan (Carboniferous) of the Jerada Massif (NE Morocco): taxonomy, biostratigraphy, facies and palaeobiogeography. Palaeontologische Zeitschrift, 84(3), 323-344.
46Aretz, M., 2010b. Habitats of colonial rugose corals: the Mississippian (Carboniferous) of Western Europe as an example for a general classification. Lethaia, 43, 558-572.
47Aretz, M., 2011. Corals from the Carboniferous of central Sahara (Algeria): the collection "Marie Legrand-Blain". Geodiversitas, 33, 581-624.
48Aretz, M. & Herbig, H.G., 2010. Corals from the Upper Viséan of the southern Azrou-Khenifra Basin (Carboniferous, Central Moroccan Meseta). Palaeoworld, 19(3-4), 295-304.
49Berkhli, M., 1999. Sédimentologie, biostratigraphie et stratigraphie séquentielle du NE de la Méséta occidentale marocaine pendant le Carbonifère inférieur (Viséen-Serpoukhovien). Thèse État, Université Moulay Ismaïl de Meknès, 290 p. (Unpublished)
50Berkhli, M. & Vachard D., 2001. New biostratigraphical data from the Early Carboniferous sequences of the Adarouch area (NE Central Morocco). Newsletter on Stratigraphy, 39, 33-54.
51Berkhli, M. & Vachard D., 2002. Le Carbonifère du Maroc central: les formations de Migoumess, de Tirhela et d’Idmarrach. Lithologie, biostratigraphie et conséquences géodynamiques. Comptes Rendus Geoscience, 334, 67-72.
52Blakey, R.C., 2007. North America Palaeogeographic Maps (accessible online: http://cpgeosystems.com/namkeypaleogeography.html).
53Blakey, R.C., 2008.-Gondwana paleogeography from assembly to breakup; a 500 m.y. odyssey. Geological Society of America Special Paper, 441, 1–28.
54Bouabdelli, M., 1989. Tectonique et sédimentation dans un basin orogénique: le sillon viséen D’Azrou-Khenifra (Est du Massif hercynien central du Maroc). Thèse d’Etat es Sciences de l’Université de Strasbourg, 262 p. (Unpublished).
55Bykova, M.S., 1966. Niznekamennougolnye korally vostocnogo kazakstana [Lower Carboniferous corals from Eastern Kazakhstan]. Trudy Instituta Geologicheski Nauk Kazakstan. Institut Geologicheski Kazakskoi SSR, 1-214. [In Russian].
56Carruthers, R.G., 1909. In Lee, G.W., A Carboniferous fauna from Nowaja Zemlja, collected by Dr. W.S. Bruce. Transactions of the Royal Society of Edinburgh, 47(1), 146-188.
57Clarke, M.J., 1966. A new species of fasciculate Aulina from Ireland. Proceedings of Royal Dublin Society. Series A, 2, 221-227.
58Coronado, I, Fernández-Martínez, E., Rodríguez, S & Tourneur, F. 2015 Reconstructing a Carboniferous coral-alcyonarian association using a biomineralogical approach. Geobiology, 13(4), 340-356.
59Cózar, P., Vachard, D., Somerville, I.D., Berkhli, M., Medina-Varea, P., Rodríguez, S. & Said, I., 2008. Late Viséan-Serpukhovian foraminiferans and calcareous algae from the Adarouch region (central Morocco), North Africa. Geological Journal, 43, 463-485.
60Cózar, P., Said, I., Somerville, I.D., Vachard, D., Medina-Varea, P., Rodríguez, S. & Berkhli, M., 2011. Potential foraminiferal markers for the Viséan-Serpukhovian and Serpukhovian-Bashkirian boundaries – a case study from Central Morocco. Journal of Paleontology, 85, 1105–1127.
61Cózar, P., Vachard, D., Somerville, I.D., Medina-Varea, P., Rodríguez, S. & Said, I., 2014. The Tindouf Basin, a marine refuge during the Serpukhovian (Carboniferous) mass extinction in the northwestern Gondwana platform. Palaeogeography, Palaeoclimatology, Palaeoecology, 394, 12-28.
62Dana, J.D. 1846. Genera of fossil corals of the family Cyathophyllidae. American Journal of Science, 1, 181-184.
63Denayer, J., Poty, E. & Aretz, M., 2011. Uppermost Devonian and Dinantian rugose corals from Southern Belgium and surrounding areas. In Aretz, M. & Poty, E. (eds), Field Guides of 11th International Symposium on Fossil Cnidaria and Porifera, Liège 2011. Kölner Forum für Geologie und Paläontologie, 20, 151-201.
64Degtyarev, D.D., 1965. Novnie rugozi iz niznego karbona shoznogo Urala. [New rugose corals from the lower Carboniferous of the South Urals]. Paleontologicheski Zhurnal, 1, 48-53. [In Russian].
65Degtyarev, D.D., 1973. Raspredelenie korallov v razveze kamennogolnykh otlozheniy Urala [Distribution of corals in a section through the Carboniferous deposits of the Urals]. Akademiya Nauk SSSR Uralskii Nauchnyi Tsentr Institut Geolgii i Geokhimimii im Akademiya Trudy, 82, 206-230. [In Russian].
66Dobrolyubova, T.A., 1952. Korally roda Gangamophyllum iz nizhego karbona Podmoskovnoy kotloviny [Corals of the genus Gangamophyllum in the Lower Carboniferous of the Moscow depression]. Akademiya Nauk SSSR Paleontologicheskii Institut Trudy, 40, 71-84. [In Russian].
67Dobrolyubova, T.A., 1958.[Lower Carboniferous colonial tetracorals from the Russian Platform]. Akademiya Nauk SSSR Paleontologicheskii Institut Trudy, 70, 1-226. [In Russian].
68Dobrolyubova, T.A., 1970. Novye odinochyne rugosy iz nizhnego karbona Russkov platform. [New solitary Rugosa fron the Lower Carboniferous of the Russian Platform]. In Astrova, G.G. & Chidonova, I.I. (eds), Novye vidy paleozoyskikh mshanok i korallov. [New species of Paleozoic bryozoans and corals]. Nauka, Moscow, 121-134. [In Russian].
69Fedorowski, J., 1981. Carboniferous corals: distribution and sequence. Acta Palaeontologica Polonica, 26, 87-160.
70Fedorowski, J. 2012. The new Upper Serpukhovian genus Zaphrufimia and homeomorphism in some rugose corals. Palaeontographica Abt. A., 296, 109-161.
71Fedorowski, J., Bamber, E.W. & Baranova, D.V., 2012. An unusual occurrence of Bashkirian (Pennsylvanian) rugose corals from the Sverdrup Basin, Arctic Canada. Journal of Paleontology, 86, 979-995.
72Fleming, J., 1828. A history of British animals. Bell & Bradfute, Edinburgh, 565 p.
73Gorsky, I.I., 1938. Carboniferous corals of Novaya Zemlya. [Kammennougol’nyi korally Novoy Zemli]. In Nalivkin, D.V. (ed.), Paleontologiya Sovetskoy Arktiki. Bessoyunzii Arkticheskii Institut, 93(2), 1-221. [In Russian].
74Gorsky, I.I., 1951. Kammennougol’nye i permskie korally Novoy Zemli. [Carboniferous and Permian corals of Novaya Zemlya]. Trudy Nauchno-Issledovatel’skogo instituta geologii Arktiki, 32, 1-168. [In Russian].
75Grabau, A. W. 1922. Palaeozoic corals of China. Part 1. Tetraseptata. Palaeontologica Sinica Series B, 2, 1-76.
76Herbig, H.G. & Aretz, M., 2015. Late Viséan (Brigantian) micheliniid corals (Tabulata) from the eastern Moroccan Meseta. In Bernecker, M. (ed.), 12th International Symposium on Fossil Cnidaria and Porifera, Muscat 2015. Abstract volume, GUtech Geoscience Conference Publication 2, 13.
77Hill, D., 1938-1941. A monograph on the Carboniferous rugose corals of Scotland. Palaeontological Society of London Monograph, Pt. 1 (1-78), Pt. 2 (79-114), Pt. 3 (115-204).
78Hill, D., 1981. Rugosa and Tabulata. In Teichert, C. (ed.), Treatise on Invertebrate Paleontology Part F (Supplement 1). Geological Society of America and University of Kansas Press, Boulder Colorado and Lawrence, Kansas, 762 p.
79Hudson, R. G. 1941. On the Carboniferous corals: Zaphrentis carruthersi sp.nov. from the Mirk Fell beds and its relation to the Z. delanouei species group. Proceedings of the Yorkshire Geological Society, 24, 290-311.
80Huvelin, P. 1967. Nappe de glissement précoce hercynienne dans les Jebilet (Maroc). Comptes Rendus de l’Académie des Sciences de Paris, série D, 265, 1039-l 042.
81Huvelin, P., 1969. Mouvements hercyniens précoces et structure du Jebel Hadid près de Khenifra (Maroc). Comptes Rendus de l’Académie des Sciences de Paris, série D, 269, 2305-2308.
82Kachanov, E.I., 1965. Podi korallov v razrabotke biostratigraficheskoi sjemi nizhnekamennougolnij otlozhenii vostochnogo sklona Yuzhnogo Urala [The coral’s role in the development of the Lower Carboniferous biostratigraphic scheme of the Southern Urals Eastern slope]. Tabulatomorfnie Koralli Devona i Karbona SSSR: 91-98. Moscow. [In Russian].
83Kachanov, E.I., 1975. O svizi periodichnosti razvitiya rannekamennougolniy korallov s osnovnimi podrazdeleleniyami nizhnego otdela karbona Urala [On the relationship between the devepopment periodicity of early Carboniferous corals with the main divisions of the Urals Lower Carboniferous]. In Kamennougolnie otlozhenitzi na Urale. 94-109, Sverdlovsk. [In Russian].
84Kershaw, S., 1994. Classification and geological significance of biostromes. Facies, 31, 81-92.
85Khoa, N.D., 1977. Carboniferous rugosa and heterocorallia from boreholes in the Lublin region (Poland). Acta Palaeontologica Polonica, 22(4), 301-404.
86Kossovaya, O.L., 1996. The mid-Carboniferous rugose coral recovery. Geological Society, London, Special Publications, 102, 187-199.
87Kozyreva, T.A., 1973. Novyy rod Opiphyllum (Rugosa) iz bashkirskogo yarusa Voronezhskoy anteklizy [A new genus Opiphyllum from the Bashkirian Stage of the Voronezh anteclise]. Akademiya Nauk SSSR, Paleontologicheskii Zhurnal, 1973(3), 129-132.
88Lonsdale, W., 1845. Description of some characteristic Palaeozoic corals of Russia. In Murchison, R.I., de Verneuil, E. & von Keyserling, A. The geology of Russia in Europe and the Ural Mountains. 1. Geology. John Murray, London, 591-634.
89McCoy, F., 1844. A synopsis of the characters of the Carboniferous limestone fossils of Ireland. Printed-at-the-University-Press by M.-H.-Gill, Dublin, 207 p.
90McCoy, F., 1849. On some new genera and species of Palaeozoic Corals and Foraminifera. Annals and Magazine of Natural History, 2(3), 1–20.
91Michelin, H., 1847. Iconographie zoophytologique, description par localités et terrains des polypiers fossiles de France et pays environnants. P. Bertrand, Paris, 348 p.
92Milne-Edwards, H. & Haime, J., 1848. Recherches sur les polypiers. Premier mémoire : observations sur la structure et le développement des polypiers en général. Annales des Sciences Naturelles, 9, 37-89.
93Milne-Edwards, H. & Haime, J., 1851. Monographie des polypiers fossiles des terrains paléozoiques, précédée d’un tableau général de la classification des Polypes. Archives du Museum d’Histoire Naturelle, 5, 1-502.
94Mitchell, M., 1989. Biostratigraphy of Viséan (Dinantian) rugose coral faunas from Britain. Proceedings of the Yorkshire Geological Society, 47, 233–247.
95Orbigny, A. d’ 1849. Note sur les polypiers fossiles. Victor Masson, Paris, 1-12.
96Parkinson, J., 1808. Organic remains of a former world, volume 2: the fossil zoophytes. C. Whittingham London, 286 p.
97Phillips, J., 1836. Illustrations of the geology of Yorkshire, Part 2: The Mountain Limestone district. John Murray, London, 253 p.
98Poty, E., 1981. Recherches sur les tétracoralliaires et les hétérocoralliaires du Viséen de la Belgique. Mededelingen rijks geologische dienst, 35(1), 1-161.
99Poty, E., 1985. A rugose coral biozonation for the Dinantian of Belgium as a basis for a coral biozonation of the Dinantian of Eurasia. In X Congrès International de Stratigraphie et de Géologie du Carbonifère, Madrid, 1983. Compte Rendu. Instituto Geológico i Minero de España, 4, 29-31
100Poty, E., Devuyst, F.X. & Hance, L., 2006. Upper Devonian and Mississippian foraminiferal and rugose coral zonations of Belgium and northern France: A tool for Eurasian correlations. Geological Magazine, 143, 829-857.
101Rafinesque, C.S. & Clifford, J.D. 1820. Prodrome d'une monographie des turbinolies fossiles du Kentuky (dans l'Amérique septentrionale). Annales générales des sciences physiques, Bruxelles, 5, 231-235.
102Rodríguez, S. & Somerville, I.D., 2007. Comparisons of rugose corals from the Upper Viséan of SW Spain and Ireland: implications for improved resolutions in late Mississippian biostratigraphy. In Hubmann, B. & Piller, W.E. (eds), Fossil Corals and Sponges: Proceedings of the 9th International Symposium on Fossil Cnidaria and Porifera, Graz, 2003. Austrian Academy of Sciences, Schriftenreihe der Erdwissenschaftlichen Kommissionen,17, 275-305.
103Rodríguez, S., Somerville, I.D., Said, I. & Cózar, P., 2012. Late Viséan coral fringing reef at Tiouinine (Morocco): implications for the role of rugose corals as building organisms in the Mississippian. Geological Journal, 47, 462-476.
104Rodríguez, S., Somerville, I.D., Said, I. & Cózar, P., 2013a. An upper Viséan (Asbian-Brigantian) and Serpukhovian coral succession at Djebel Ouarkziz (Northern Tindouf Basin, Southern Morocco). Rivista Italiana di Paleontologia e Stratigrafia, 119(1), 3-17.
105Rodríguez, S., Somerville, I.D., Said, I. & Cózar, P., 2013b. Mississippian-like rugose corals from a Bashkirian biostrome in the Tindouf Basin, S. Morocco. Spanish Journal of Palaeontology, 28(2), 253-282.
106Rodríguez, S., Said, I., Somerville, I.D., Cózar, P. & Coronado, I., (submitted). Coral assemblages of the Serpukhovian- Bashkirian transition from Adarouch (Morocco). Proceedings of the 12th International Symposium on Fossil Cnidaria and Porifera.
107Said, I., 2005. Estudio de los corales rugosos con disepimentos del Mississippiense del NE de la meseta marroquí (sectores de Adarouch y Agouaraï) (unpublished). PhD thesis, Universidad Complutense de Madrid, 240p.
108Said, I & Rodríguez, S. 2007. A new genus of coral (Rugosa), from Adarouch Area (Brigantian, NE Central Morocco) Coloquios de Paleontología , 57, 23-35.
109Said, I., Berkhli, M. & Rodríguez, S., 2007. Preliminary data on the coral distribution in the Upper Viséan (Mississippian) succession from Adarouch area (NE Central Morocco). In Hubmann, B. & Piller, W. (eds), Fossil Corals and Sponges: Proceedings of the 9th International Symposium on Fossil Cnidaria and Porifera. Schriftenreihe der Erdwissenschaftlichen Kommission 17, 353–364.
110Said, I. & Rodríguez, S., 2008. Descripción de los corales aulophyllidos del Viseense y Serpujoviense del área de Adarouch (Marruecos). Coloquios de Paleontología, 58, 13-40.
111Said, I., Rodríguez, S., Berkhli, M., Cózar, P. & Gómez-Herguedas, A., 2010. Environmental parameters of a coral assemblage from the Akerchi Formation (Carboniferous), Adarouch Area, central Morocco. Journal of Iberian Geology, 36(1), 7-19.
112Said, I., Rodríguez, S., Somerville, I.D. & Cózar, P., 2011. Environmental study of coral assemblages from the upper Viséan Tizra Formation (Adarouch area, Morocco): implications for Western Palaeotethys biogeography. Neues Jahrbuch für Geologie und Palaeontologie, Abhandlungen, 260(1), 101-118.
113Said, I., Somerville, I.D., Rodríguez, S. & Cózar, P., 2013. Mississippian coral assemblages from the Khenifra area, Central Morocco: biostratigraphy, biofacies, palaeoecology and palaeobiogeography. Gondwana Research, 23, 367-379.
114Salée, A., 1913. Contribution à l'étude des polypiers du calcaire carbonifère de la Belgique II. Le groupe des Clisiophyllides. Mémoires de l'Institut Géologique de l'Université de Louvain, 1, 179-293.
115Sayutina, T.A., 1973. Nizhnekamennougolnye korally Severnego Urala: podotryad Acrophyllina [Lower Carboniferous corals of the northern Urals; Suborder Acrophyllina]. Akadademiya Nauk SSSR, Paleontologicheskii Institut Trudy, 140, 1-168. [In Russian].
116Semenoff-Tian-Chansky, P., 1974. Recherches sur les Tétracoralliaires du Carbonifère du Sahara Occidental. Mémoires du Museum national d’histoire naturelle, série C, Sciences de la Terre, 30, 1-316.
117Semenoff-Tian-Chansky, P. 1985. Corals. In Wagner, R.H., Winkler-Prins, C.F., Granados, L.F. (eds), The Carboniferous of the World, II, Australia, Indian Subcontinent, South Africa, South America and North Africa. IUGS Publication, 20, 374-381.
118Somerville, I.D. & Rodríguez, S., 2007. Rugose coral associations from the Upper Viséan of Ireland, Britain and SW Spain. In Hubmann, B. & Piller, W.E. (eds), Fossil Corals and Sponges: Proceedings of the 9th International Symposium on Fossil Cnidaria and Porifera, Graz, 2003. Austrian Academy of Sciences, Schriftenreihe der Erdwissenschaftlichen Kommissionen,-17, 329-351.
119Somerville, I.D., Rodríguez, S., Said, I. & Cózar, P., 2012. Mississippian coral assemblages from Tabainout mud-mound complex, Khenifra area, Central Morocco. In Denayer, J., Aretz, M. & Poty, E. (eds), Proceedings of the 11thInternational Symposium on Fossil Cnidaria and Porifera, Liège, Belgium, 2011. Geologica Belgica, 15(4), 308-316.
120Somerville, I.D., Cózar, P., Said, I., Vachard D., Medina-Varea, P. & Rodríguez, S., 2013. Palaeobiogeographical constraints on the distribution of foraminifers and rugose corals in the Carboniferous Tindouf Basin, S. Morocco. Journal of Palaeogeography, 2, 1–18.
121Termier, G. & Termier, H., 1950. Paléontologie marocaine II. Invertébrés de l'Ere Primaire. Foraminifères, spongaires et coelentérés. Notes du service géologique du Maroc, 73, 1-220.
122Termier, G., Termier, H., & Vachard, D., 1975. Recherches micropaléontologiques dans le Paléozoïque du Maroc central. Cahiers de Micropaléontologie, 4, 1-99.
123Thomson, J., 1880. Contributions to our knowledge of the rugose corals from the Carboniferous Limestone of Scotland. Proceedings of the Royal Philosophical Society of Glasgow, 12, 225-261.
124Thomson, J. & Nicholson, H.A., 1876. Contributions to the study of the chief generic types of the Palaeozoic corals. Annals and Magazine of Natural History, 17(2), 123–128; 17(4), 290–305; 17(6), 451–461; 18(7), 68–73.
125Vachard, D., Orberger, B., Rividi, N., Pille, L. & Berkhli M., 2006. New Late Asbian/early Brigantian (Late Viséan, Mississippian) dates in the Mouchenkour Formation (central Morocco): palaeogeographical consequences. Comptes Rendus Palevol, 5, 769–777.
126Vassiljuk, N.P., 1960. Nizhnekamennougolnye korally Donetskogo basseyna [Lower Carboniferous corals of the Donets Basin]. Trudy Instituta geologicheskich nauk, Akademiya Nauk Ukrainskoy SSR, seriya stratigrafii i paleontologii, Kiev, 13, 178 p. [In Russian].
127Volkova, M. S., 1941. Nizhnekamenougolnye korally Centralnogo Kazakstana [Lower Carboniferous corals from Central Kazakhstan]. Materialy po Geologii i Tsentralnogo Kazakstana, 11, 119 p. [In Russian].
128Yabe, H. & Hayasaka, I., 1920. Geographical research in China 1911-1916. Palaeontology of Southern China, Atlas of fossils. Tokyo Geographical Society, Tokyo, 221 p.
129Yü, C.C., 1933. Lower Carboniferous corals of China. Palaeontologica Sinica, 12(3), 3-133.
130Manuscript received 26.06.2015, accepted in revised form 25.11.2015, available on line 05.07.2016.
