


- Accueil
- Volume 25 (2022)
- number 1-2
- The highly diversified rugose coral fauna from the Lower Givetian Meerbüsch quarry in the Eifel Hills (Germany)


Visualisation(s): 4766 (52 ULiège)
Téléchargement(s): 1620 (17 ULiège)

The highly diversified rugose coral fauna from the Lower Givetian Meerbüsch quarry in the Eifel Hills (Germany)

Abstract
Twenty taxa of Lower Givetian rugose corals have been investigated in the Dreimühlen Formation from the Meerbüsch quarry, in the Eifel Hills. The fauna consists mostly of solitary coralla associated with some fasciculate specimens of Disphyllum caespitosum (Goldfuss, 1826) and with large massive colonies of Argutastrea briceae (Rohart, 1988). The genus Marennophyllum n. gen. is introduced with Cystiphylloides marennense Coen-Aubert, 2019 as type species to include the solitary species previously assigned to the fasciculate genus Cystiphylloides Chapman, 1893. Three species are new: Marennophyllum wenningi n. sp., Grypophyllum schroederi n. sp. and Macgeea mistiaeni n.sp. The stratigraphic distribution of the material observed in Meerbüsch quarry is compared with that of the Middle Devonian in the Eifel Hills as a whole, the southern part of Belgium, and other countries of Western Europe. Some species such as Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851), A. vermiculare (Goldfuss, 1826), Grypophyllum convolutum (Wedekind, 1925), Stringophyllum acanthicum (Frech, 1885) and Aristophyllum luetti Coen-Aubert, 1997 are also present at the northern margin of Gondwana (Ma’der in Morocco, Zemmour in Mauritania, and Alborz Mountains in Iran). The occurrence of Heliophyllum cf. cribellum Oliver & Sorauf, 2002 in Meerbüsch quarry suggests an Eastern North American influence.
Table des matières
1. Introduction
1My interest in the Givetian rugose corals of the Meerbüsch quarry in the Eifel Hills, Germany is due to the discovery made by Walter Wenning, a German collector from Düren, of a level very rich in large colonies of Argutastrea briceae (Rohart, 1988) and never mentioned in German publications. Therefore, I visited this outcrop in 1999–2000 with W. Wenning and Michel Coen who made a geological survey bed by bed of the succession which concerns nearly all the Dreimühlen Formation and the base of the Rodert Formation. Additionally, it appears that the solitary rugose corals of this quarry are highly diversified and associated with some fasciculate species.
2The Meerbüsch quarry is situated in the Hillesheim Syncline belonging to the southern part of the Eifel Hills (Fig. 1) where the Lower Givetian is successively represented by the upper part of the Ahbach Formation, the Loogh, Cürten, Dreimühlen, Rodert and Kerpen Formations (Weddige, 1996; Schröder, 1998). The Dreimühlen Formation is subdivided in ascending order, into the Binz, Ley and Galgenberg Members. In fact, the Meerbüsch quarry is not very well known in the literature. It is one of the localities sampled by Salerno (2008, p. 9, fig. 28), a specialist of Givetian stromatoporoids, who also provided a log of the Galgenberg Member and the base of the Rodert Formation. Nose et al. (2017) investigated the top of the Binz Member in the Meerbüsch quarry, for a detailed palaeoecological study. In addition, they figured two plates of solitary rugose corals collected in this level.
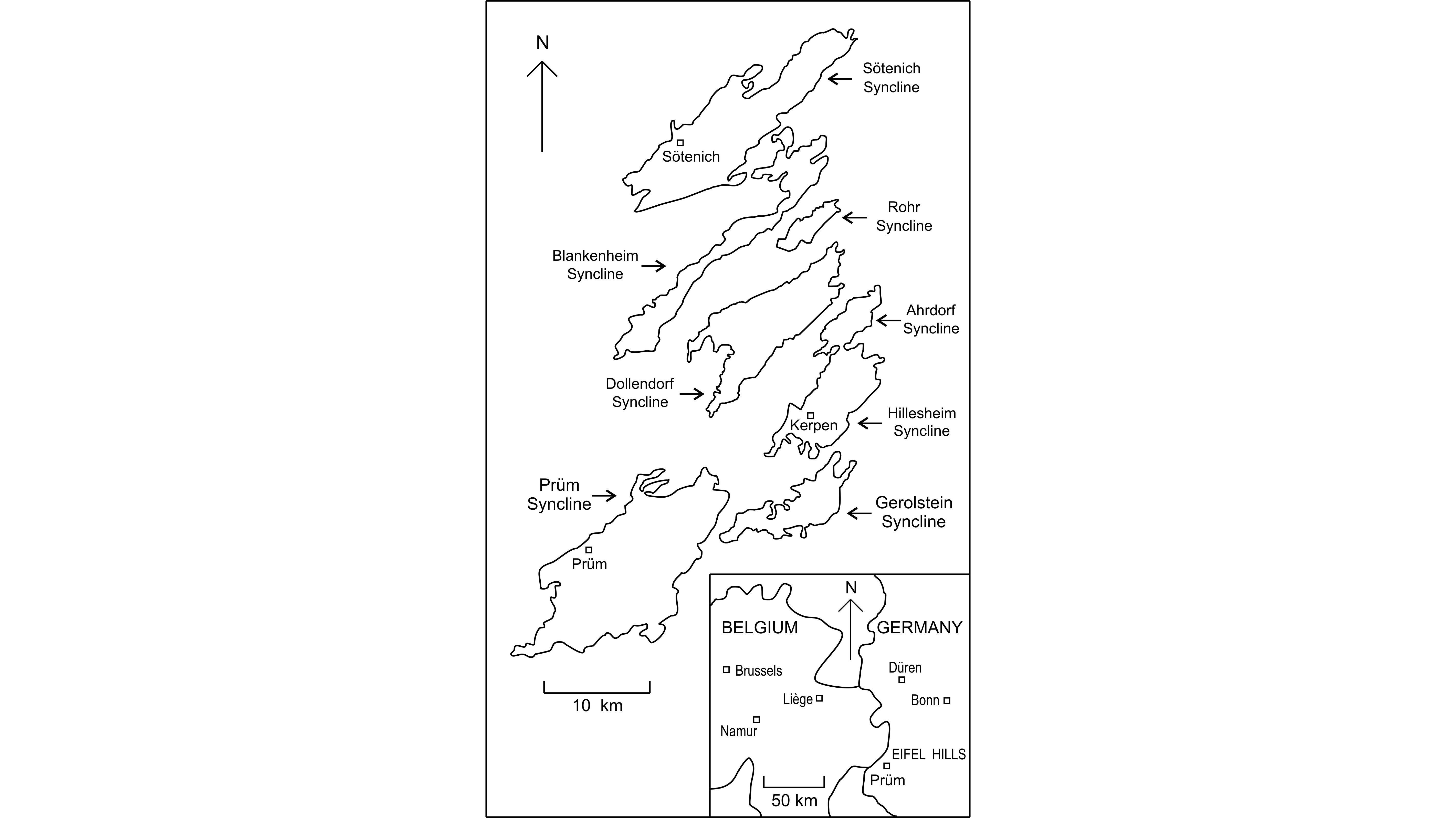
Figure 1. General location and map showing the Middle Devonian synclines of the Eifel Hills after Königshof et al. (2016, fig. 2).
2. Description of the Meerbüsch quarry
3The Meerbüsch quarry is situated in the Hillesheim Syncline, 2 km to the northeast of Kerpen, at the northeastern end of Meerbüsch Hill (Fig. 2).
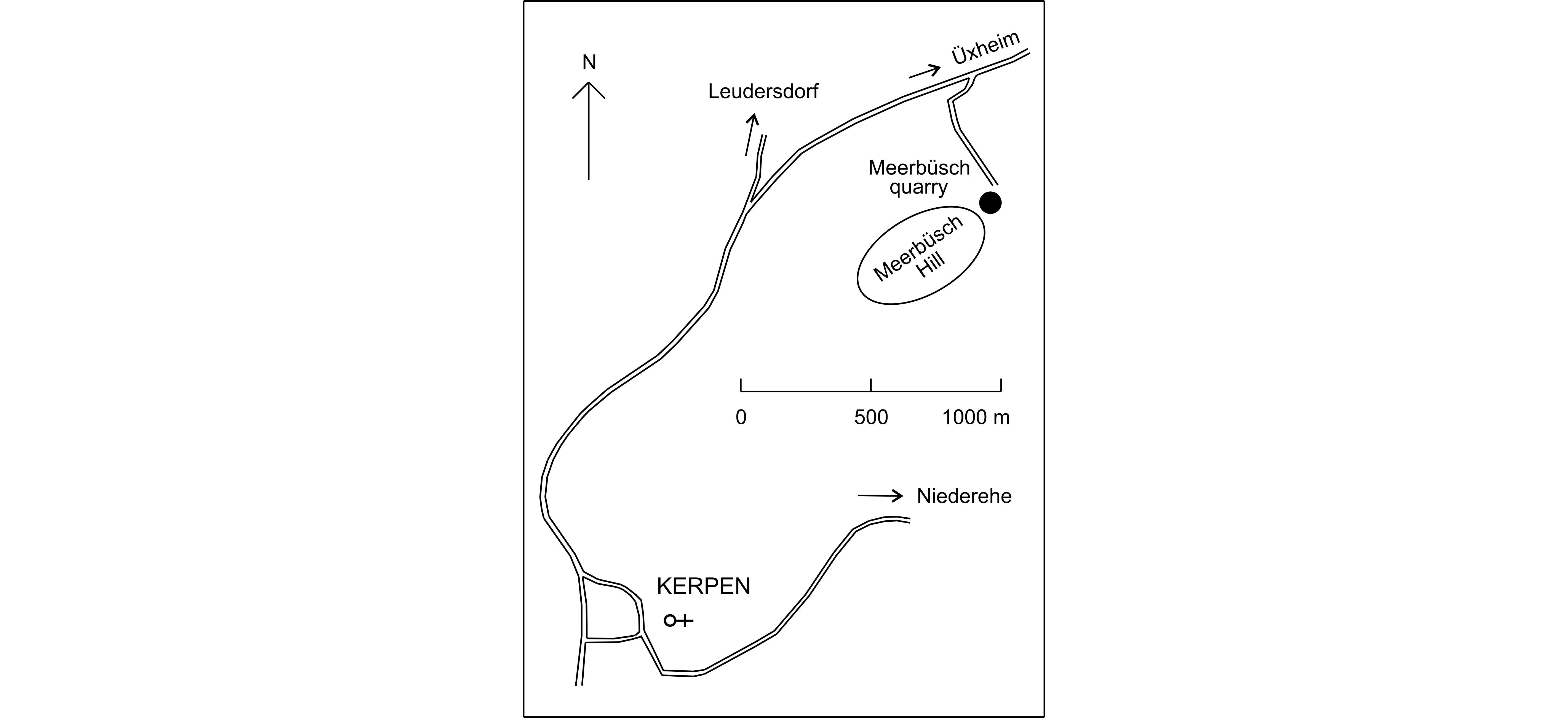
Figure 2. Location of the Meerbüsch quarry.
4At the northern border of the quarry, there are about 3 m of bluish-grey marls containing large solitary rugose corals associated with bushes of fasciculate rugose corals and thamnoporids. These layers belong to the top of the Binz Member, in the lower part of the Dreimühlen Formation (Fig. 3). The rugose corals are represented by calceolids, Marennophyllum praecipuum (Wedekind & Vollbrecht, 1931), M. wenningi n. sp., Marennophyllum sp.1, Atelophyllum leave (Schulz, 1883), Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851), A. vermiculare (Goldfuss, 1826), Disphyllum caespitosum (Goldfuss, 1826) and Aristophyllum luetti Coen-Aubert, 1997.
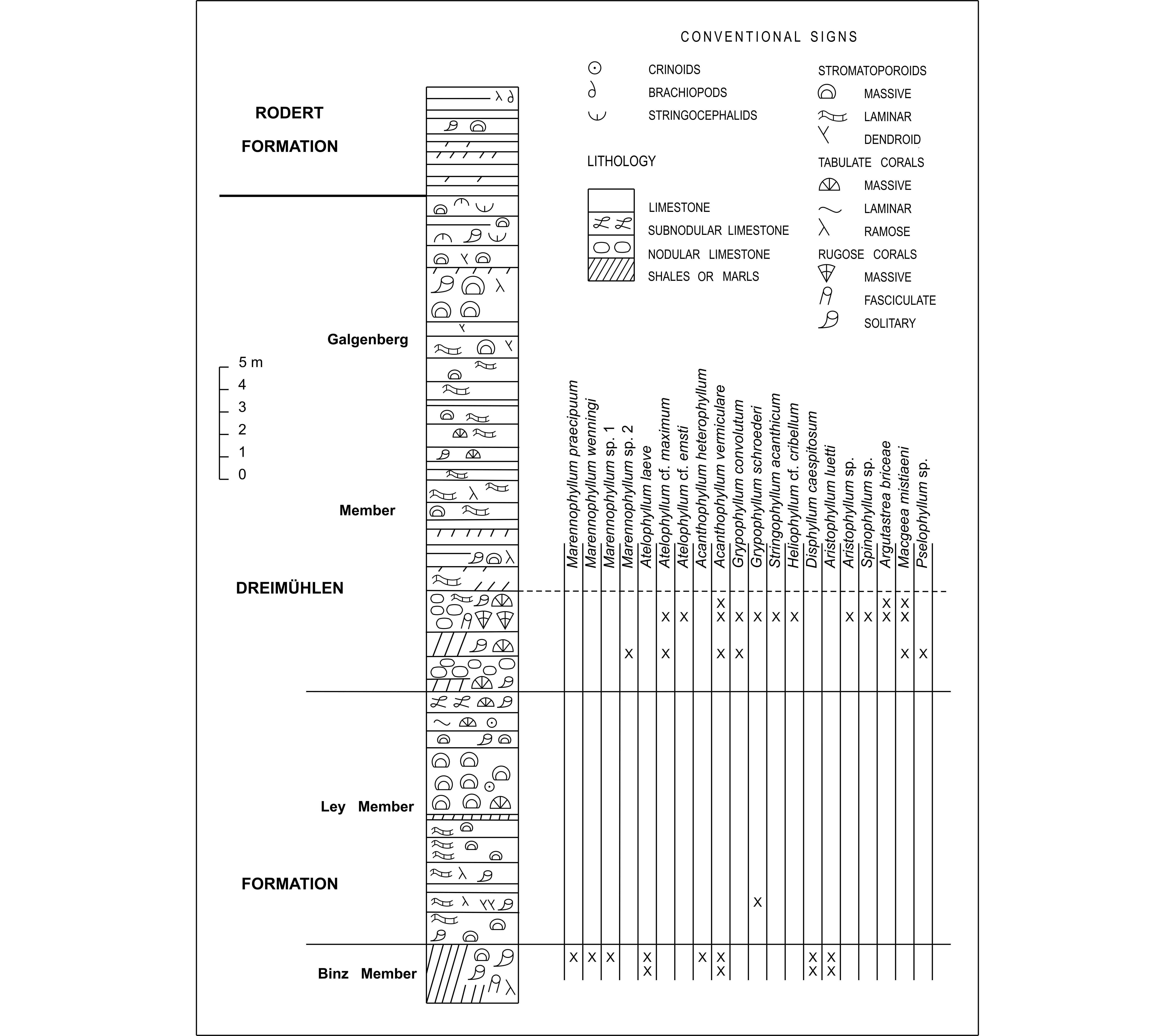
Figure 3. Log of the Meerbüsch quarry with the distribution of rugose corals.
5The Ley Member consists of:
-
5.8 m: limestone relatively rich in laminar stromatoporoids accompanied by massive stromatoporoids in the lower and upper parts and occurrence of thin beds of marls at the base and at the top. Some specimens of Grypophyllum schroederi n. sp. have been collected about 2 m above the base of this unit.
-
4.5 m: limestone with numerous massive stromatoporoids together with some alveolitids and favositids at the base and at the top; locally some crinoids and solitary rugose corals.
-
0.9 m: subnodular limestone with alveolitids and solitary rugose corals. There is a small fault at this level.
6The Galgenberg Member starts with:
-
2.6 m: yellow marls with an intercalation of nodular limestone in the middle; occurrence of solitary and fasciculate rugose corals, alveolitids and favositids in the marls.
-
1.9 m: nodular limestones with numerous large colonies of Argutastrea briceae in the lower 0.9 m. Additionally, abundant and various corals are represented in all these nodular limestones, by solitary and fasciculate rugose corals, favositids, alveolitids, thamnoporids, scolioporids and heliolitids. Appearance of laminar stromatoporoids at their top.
7Besides A. briceae, the following rugose corals have been collected in these two argillaceous units: Marennophyllum sp. 2, Atelophyllum cf. maximum (Schlüter, 1882), A. cf. emsti (Wedekind, 1922), Acanthophyllum vermiculare, Grypophyllum convolutum (Wedekind, 1925), G. schroederi, Stringophyllum acanthicum (Frech, 1885), Heliophyllum cf. cribellum Oliver & Sorauf, 2002, Aristophyllum sp., Spinophyllum sp., Macgeea mistiaeni n.sp. and Pselophyllum sp.
8The rest of the Galgenberg Member is characterized by pure and bedded limestones composed of:
-
3.2 m: limestone with some scattered laminar and massive stromatoporoids, ramose tabulate corals and solitary rugose corals; a few thin beds of shale.
-
8.85 m: limestone with rather frequent laminar stromatoporoids associated locally with massive and dendroid stromatoporoids, ramose tabulate corals, alveolitids, heliolitids and solitary rugose corals.
-
3.15 m: limestone rich in massive stromatoporoids with some dendroid stromatoporoids, ramose tabulate corals and solitary rugose corals.
-
2.4 m: two levels containing stringocephalids brachiopods and separated by fine-grained limestone; occurrence of some massive stromatoporoids and solitary rugose corals in the coquinas.
9Finally, the base of the Rodert Formation is exposed with a thickness of 4.8 m. It is composed of thin-bedded limestone with locally a few massive stromatoporoids, solitary rugose corals, ramose tabulate corals and brachiopods.
10Though my collection of rugose corals from the Meerbüsch quarry is not so large, it shows a wide variability at the generic and specific levels which is interesting to describe.
3. Systematic Palaeontology
11The types of the new species and the figured specimens are stored in the Collection of Palaeontology of the Institut royal des Sciences naturelles de Belgique at Brussels (prefixed IRScNB). With the exception of three colonies of Argutastrea briceae, all the material investigated herein has been collected by the author, in the Meerbüsch quarry (Eifel Hills, Germany).
12Family Cystiphyllidae Milne-Edwards & Haime, 1850
13Genus Marennophyllum n. gen.
14Type species. Cystiphylloides marennense Coen-Aubert, 2019.
15Derivation of name. After the well-known Marenne quarry situated on the south side of the Dinant Synclinorium in Belgium.
16Diagnosis. Solitary rugose corals. Interior of the corallum filled with vesicular dissepiments and tabellae badly differentiated. Small septal spines absent or rare to more numerous, mostly in the dissepimentarium. Conical stereoplasmic thickenings lacking or weakly to strongly developed, more frequent in the axial zone and present at all growth stages of the corallum. Successive septal cones locally fusing to fill varying amounts of the interior of the corallum. Peripheral globose dissepiments more or less inclined. Axial zone comprising globose tabellae, commonly somewhat larger than the dissepiments, and occasionally incomplete tabulae.
17Discussion. According to McLean (2021, p. 18, 22), the fasciculate genus Cystiphylloides Chapman, 1893, whose type species is C. aggregatum (Billings, 1859) probably from the Emsian of Ontario in Canada, is not appropriate for the solitary forms previously assigned to this taxon. Therefore, such solitary coralla are referred by McLean (2021) to the genus Nardophyllum Wedekind, 1925 whose type species is N. exzentricum Wedekind, 1925 from the Lower Givetian of the Eifel Hills. The holotype of this species is only represented by a transverse section figured among others by Wedekind (1925, pl. 9, fig. 59), without any longitudinal section. Additionally, N. exzentricum has been synonymized by Birenheide (1964, p. 33) with Cystiphyllum macrocystis Schlüter, 1889 whose lectotype from the base of the Givetian in the Eifel Hills is only known by a polished longitudinal section. For these reasons, it appears that Nardophyllum is badly defined.
18On the contrary, Cystiphylloides marennense Coen-Aubert, 2019, type species of Marennophyllum n. gen. and investigated by Coen-Aubert (2019), is well preserved, very abundant, and easy to collect in the Lower Givetian of the Belgian Marenne quarry. This species occurs close to the base of the Mont d’Haurs Formation, in a coralliferous level which is exposed in different parts of the quarry due to its large size and repetitions related to an important faulted zone. The numerous species listed with interesting details and assigned by McLean (2021, pp. 23–26) to Nardophyllum can be referred to Marennophyllum.
19Marennophyllum praecipuum (Wedekind & Vollbrecht, 1931)
20(Plate 1A–C)
21* 1931 Paralytophyllum praecipuum n. sp.; Wedekind & Vollbrecht, pl. 19, figs 1–12.
222017 Mesophyllum (Cystiphylloides) secundum pseudoseptatum (Schulz, 1883); Nose et al., pl. 2, fig. 5.
23Holotype. Specimen and thin sections WDKD 1849–1859 and 8125–8127 stored in the Forschungsinstitut Senckenberg at Frankfurt am Main, Germany. Lower Givetian Cürten Formation to the Binz Member of the Dreimühlen Formation at Niederehe, Hillesheim Syncline, Eifel Hills in Germany. Pl. 19, figs 1–12 in Wedekind & Vollbrecht (1931).
24Material. 2 specimens with 4 thin sections: Meerbüsch EIF346 and EIF395.
25Diagnosis. A species of Marennophyllum with a mean diameter of 25 mm to 42 mm. Corallum filled with numerous small dissepiments. Abundant and relatively thick septal spines with a few short septa in the dissepimentarium. Occurrence of a few septal cones in the tabularium.
26Description. The material consists of two fragmentary solitary rugose corals with growth lines, whose height varies between 4.5 cm and 8 cm; one of them is ceratoid. The outer wall is only preserved very locally.
27The corallum is completely filled with numerous small globose dissepiments. Septal spines are frequent, especially in the inner part of the dissepimentarium where they may be relatively long. Weak and more or less complete rings of stereoplasmic thickening appear in transverse section, in the inner dissepimentarium and in the tabularium. At the outer margin of the tabularium in one corallum, there are also a few short septa.
28The dissepimentarium consists of numerous rows of small globose dissepiments arranged in horizontal layers at the periphery and inclined in its inner part. The axial tabellae are typically vesicular and occasionally larger than the dissepiments; their overall pattern is horizontal or concave. Septal spines are observed in longitudinal section whereas septal cones are restricted to the tabularium.
29The diameter of the corallum ranges from 35 mm to 42 mm.
30Discussion. The material of Meerbüsch quarry represented by two specimens and the corallum figured by Nose et al. (2017) is similar to the holotype of Marennophyllum praecipuum which shows a variable development of thick septal spines throughout the dissepimentarium, in the different transverse sections of Wedekind & Vollbrecht (1931). According to Birenheide (1964, pp. 27–28; 1968, p. 21), M. praecipuum is a synonym of M. pseudoseptatum (Schulz, 1883) from the Lower Givetian Binz Member of the Dreimühlen Formation in the Hillesheim Syncline. The lectotype of this species has been chosen by Birenheide (1964) who gave only an external view of this specimen without investigating it in thin sections. The two coralla of M. pseudoseptatum illustrated by Birenheide (1964, 1978, fig. 114) are characterized by some discontinuous and thin septa in the dissepimentarium whereas the samples figured by Wedekind (1925, pl. 8, figs 49–50, 52) are more typical of M. praecipuum. This is also the case for the type specimens of M. tabulatum (Wedekind, 1925, pl. 7, figs 38–39) and M. striatum (Wedekind, 1925, pl. 7, fig. 40) which are much smaller than M. praecipuum; they have been collected in the Lower Givetian Loogh Formation of the Gerolstein Syncline, in the Eifel Hills, and once more synonymized by Birenheide (1968, pp. 27–28) with M. pseudoseptatum. Finally, it must be mentioned that the lectotype and two paratypes of M. corneolum (Wedekind & Vollbrecht, 1931) illustrated by Wedekind & Vollbrecht (1931, pl. 18, figs 8–9, pl. 20, figs 1–6) and coming from the same type locality and horizon as M. praecipuum are closely related to this species though they have rather short spines.
31Distribution. The species is only known in the Lower Givetian from the Hillesheim Syncline in the Eifel Hills. The material of Meerbüsch quarry comes from the top of the Binz Member in the lower part of the Dreimühlen Formation. The holotype of Marennophyllum praecipuum has been collected in the vicinity of this outcrop at Niederehe, in the Cürten Formation to the Binz Member of the Dreimühlen Formation.
32Marennophyllum wenningi n. sp.
33(Plate 1F–H)
34p. 1931 Lytophyllum corneolum n. sp.; Wedekind & Vollbrecht, pl. 21, figs 1–7, pl. 22, figs 1–8, pl. 23, figs 1–6 (non pl. 15, figs 4–6, pl. 18, figs 8–9, pl. 20, figs 1–6, pl. 31, fig. 6).
352017 Mesophyllum (Cystiphylloides) secundum secundum (Goldfuss, 1826); Nose et al., pl. 3, fig. 5.
36Derivation of name. The species is dedicated to Walter Wenning, a German collector from Düren who invited me to visit the Meerbüsch quarry.
37Holotype. IRScNB a13529 (= Plate 1F–G). Specimen EIF396 collected by M. Coen-Aubert in 2000, in the Lower Givetian of the Eifel Hills, Germany.
38Type locality and horizon. Meerbüsch quarry in the Hillesheim Syncline, Eifel Hills in Germany. Top of the Lower Givetian Binz Member in the lower part of the Dreimühlen Formation.
39Material. 3 specimens with 5 thin sections: Meerbüsch EIF396, EIF 397 and EIF468.
40Diagnosis. A species of Marenophyllum with a mean diameter of 18 mm to 32 mm. Corallum filled with relatively large dissepiments. Occurrence of some septal spines. Rare and weak septal cones in the tabularium.
41Description. The material consists of conical, trochoid, or ceratoid coralla which are often fragmentary and show sometimes growth lines; their height varies between 2.5 cm and 5.5 cm. One specimen is affected by rejuvenescence at the top. The outer wall is thin and locally preserved.
42The corallum is filled with rather large globose dissepiments and tabellae with a few septal spines. In transverse section, thin zones of stereoplasmic thickening are only observed locally at the border of the tabularium or in the dissepimentarium where thick spines or fragments of contiguous septa may be present.
43The dissepiments are inclined towards the axis of the corallum or occasionally arranged in horizontal layers at the periphery. The axial zone is narrow and occupied by vesicular tabellae with an overall horizontal or sometimes concave pattern. Weak septal cones occur locally in the tabularium.
44The diameter of the corallum ranges from 24 mm to 30 mm.
45Discussion. The new species has some affinities with Marennophyllum marennense which has a much greater diameter. It differs from M. praecipuum by fewer and small septal spines and by larger dissepiments arranged at the periphery in a few rows which are usually inclined. Marennophyllum wenningi is also represented by several paratypes of M. corneolum illustrated by Wedekind & Vollbrecht (1931). As mentioned above, the holotype and two paratypes of this taxon characterized by numerous and longer septal spines are closely related to M. praecipuum. Marennophyllum corneolum was also considered by Birenheide (1964, 1968) as a synonym of M. pseudoseptatum. Moreover the two coralla assigned to this species by Birenheide (1964) are without doubt separated from M. wenningi by the occurrence of some discontinuous and thin septa.
46Nose et al. (2017) identified as M. secundum (Goldfuss, 1826) one specimen of the new species collected at the top of the Binz Member in Meerbüsch quarry. Birenheide (1964, p. 25, pl. 27, fig. 130) has figured the holotype of this taxon without sectioning and studying it; his material from the Middle Devonian of the Eifel Hills is distinguished from M. wenningi by some segments of thin septa and by more numerous small dissepiments.
47Distribution. The species is only known in the Lower Givetian from the Hillesheim Syncline in the Eifel Hills. The material of Meerbüsch quarry comes from the top of the Binz Member in the lower part of the Dreimühlen Formation. The specimens figured by Wedekind & Vollbrecht (1931) have been collected in the vicinity of this outcrop at Niederehe, in the Cürten Formation to the Binz Member of the Dreimühlen Formation.
48Marennophyllum sp. 1
49(Plate 2A–B)
50Material. 1 specimen with 2 thin sections: Meerbüsch EIF463.
51Description. The material is restricted to a cylindrical corallum with growth rings, which is 7.5 cm high. The outer wall is thick and partially preserved.
52The corallum is filled with numerous small globose dissepiments. Septal spines are frequent in transverse section and more or less complete and weak septal cones are observed within the dissepimentarium and the tabularium. An eccentric patch of stereoplasma composed of coarse and contiguous septal trabeculae is present locally in the inner part of the dissepimentarium.
53The longitudinal section is asymmetric. On one side of the corallum, there are numerous rows of small dissepiments with small septal spines, arranged in horizontal layers at the periphery and inclined in the inner part dissepimentarium. On the other side of the specimen, there are only a few rows of small, inclined dissepiments and then a wide zone of compact stereoplasma obscuring the inner part of the dissepimentarium and partly the tabularium. This axial part of the corallum is occupied by small vesicular tabellae with some septal cones.
54The diameter of the corallum ranges from 35 mm to 40 mm.
55Discussion. The corallum of Meerbüsch quarry resembles one specimen figured by Birenheide (1964, 1978, figs 117c–d) as Marennophyllum macrocystis and coming from the Lower Givetian Loogh Formation of the Hillesheim Syncline, in the Eifel Hills. The lectotype of this species from the base of the Givetian in the Sötenich area (Fig. 1), also in the Eifel Hills, has been selected by Birenheide (1964, p. 34) who illustrated only a narrow polished longitudinal section of this corallum, without any stereoplasmic thickening. The same feature characterizes the small longitudinal section and only figure given by Schlüter (1889, pl. 3, fig. 10). Therefore, the interpretation of M. macrocystis is highly variable among the German authors. Marennophyllum crassicaulum (Wedekind & Vollbrecht, 1931, pl. 29, figs 1–5) from the Lower Givetian of the Cürten Formation to the Binz Member of the Dreimühlen Formation at Niederehe, in the Hillesheim Syncline, is related to Marennophyllum sp.1. However, this species differs from the specimen of Meerbüsch quarry by its greater size and by stronger eccentric stereoplasmic thickenings. Marennophyllum crassicaulum has been considered as synonym of M. pseudoseptatum by Birenheide (1964, 1968, p. 8) whereas it has been placed in synonymy with M. macrocystis by McLean (1976, p. 8; 2021, p.24).
56Distribution. The corallum of Meerbüsch quarry comes from the top of the Lower Givetian Binz Member, in the lower part of the Dreimühlen Formation.
57Marennophyllum sp. 2
58(Plate 1D–E)
59Material. 1 specimen with 2 thin sections: Meerbüsch EIF403.
60Description. The material is restricted to a fragmentary cylindrical corallum with growth lines, which is 8 cm high. The outer wall is present locally and encrusted by a laminar alveolitid on one side.
61The corallum is filled with globose dissepiments which are relatively small at the periphery and larger in the tabularium. Some short discontinuous septa are present against the outer wall and mainly in the outer dissepimentarium. No septal cones are observed in transverse section.
62The dissepimentarium consists of numerous rows of small globose and mostly inclined dissepiments. The axial tabellae are typically vesicular with an overall concave pattern. A few zones of thin stereoplasmic thickening occur in the tabularium.
63The diameter of the corallum ranges from 49 mm to 54 mm.
64Discussion. Marennophyllum sp. 2 is distinguished from M. praecipuum by its greater diameter, mostly inclined dissepiments, less septal spines and the occurrence of short discontinuous septa against the outer wall and in the dissepimentarium. The last feature also characterizes M. lenzi (Pedder & McLean, 1982) from the Upper Pragian of the Yukon Territory in Canada. But in this species, the septa are often thick and occasionally contiguous at the periphery whereas the dissepimentarium consists of fewer dissepiments with some stereoplasmic thickenings. Taxa such as Marennophyllum sp. 2 and M. lenzi are in fact intermediate between Marennophyllum and Atelophyllum Wedekind, 1925. As mentioned by Coen-Aubert (2019, pp. 125–126), it is difficult to use for these forms the genus Zonophyllum Wedekind, 1924 as its type species Z. duplicatum Wedekind, 1924 from the Lower Eifelian of the Eifel Hills is badly known and defined. McLean (2021, pl. 8, figs 3, 6–7, 10, 13–14) has recently figured several transverse from the type material of Wedekind (1924), but no longitudinal section. With only one specimen of Marennophyllum sp. 2, it is not possible to discuss in detail its generic attribution.
65Distribution. The corallum of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation.
66Genus Atelophyllum Wedekind, 1925
67Type species. By original designation, Mesophyllum emsti Wedekind, 1922.
68Diagnosis. See Coen-Aubert (2019).
69Discussion. Coen-Aubert (2019) has preferred to use Atelophyllum rather than Lekanophyllum Wedekind, 1924. The two genera are considered as synonyms and this was confirmed by McLean (2021, p. 64). Their type species are respectively Atelophyllum emsti from the Lower Givetian of the Sauerland in Germany and Lekanophyllum punctatum Wedekind, 1924 from the Upper Eifelian of the Eifel Hills. The holotype of L. punctatum is only known in transverse sections refigured recently by McLean (2021, pl. 20, figs 7–9) whereas the same author has illustrated two transverse and two longitudinal sections (McLean, 2021, pl. 21, figs 1, 4–6) for the holotype of Atelophyllum emsti.
70Atelophyllum laeve (Schulz, 1883)
71(Plate 2D–G)
72* 1883 Actinocystis laevis Schulz; Schulz, p. 238, pl. 22, figs 1–2.
731956 Dialytophyllum laevis (Chulz); Ma, p. 73, pl. 26, fig. 2, pl. 38, fig. 1.
741964 Plasmophyllum (Mesophyllum) maximum laeve (Schulz 1883); Birenheide, p. 44, pl. 8, figs 33–35, pl. 12, fig. 55, pl. 15, fig. 75, pl. 16, fig. 76, pl. 17, fig. 78, pl. 18, fig. 86, pl. 27, figs 133–134, pl. 28, fig. 135.
751978 Mesophyllum (M.) maximum laeve (Schulz 1883); Birenheide, p. 168, fig. 106.
76non 1998 Mesophyllum (Mesophyllum) maximum laeve (Schulz 1883); Schröder, p. 66, pl. 18, fig. 106.
772017 Mesophyllum (Mesophyllum) maximum laeve (Schulz, 1883); Nose et al., pl. 2, fig. 4.
78Lectotype. Specimen 3b of the Schulz collection chosen by Birenheide (1964, p. 44) and stored in the Paläontologisches Institut from the University of Bonn in Germany. Lower Givetian Binz Member of the Dreimühlen Formation at Berndorf, Hillesheim Syncline, Eifel Hills, Germany. Pl. 22, figs 1–2 in Schulz (1883) and pl. 27, fig. 134 in Birenheide (1964).
79Material. 6 specimens with 10 thin sections: Meerbüsch EIF345, EIF347, EIF394, EIF451, EIF452 and EIF460.
80Diagnosis. A species of Atelophyllum with 92 to 106 septa at a diameter of 36 mm to 55 mm. Major and minor septa relatively long and well developed. Numerous rows of small dissepiments arranged in horizontal layers at the periphery.
81Description. The material consists of fragmentary ceratoid and cylindrical coralla often with growth lines, whose height varies between 4 cm and 10.5 cm. Two specimens are characterized by a broad axial depression bordered by a narrow peripheral platform whereas another one is affected by rejuvenescence at the top. The outer wall is rarely preserved and locally encrusted by auloporids in one sample.
82The septa are frequently thin and even discontinuous at the periphery where there are strong yardarm and spinose carinae in some coralla. Then the septa are non-carinate and variably dilated in the dissepimentarium. In some cases, this thickening affects more the major septa. A few dissepiments like abutments occur in the outer dissepimentarium of one specimen. The septa become thin in the tabularium or at least at their axial ends. But they may be also even or more and less dilated in this axial zone.
83When the major septa are thin in the tabularium, they reach the axis of the corallum or leave a small open space in its centre. If they are thicker, this open space is much wider. In both cases, the axial ends of the major septa are sometimes rhopaloid, discontinuous, curved or forming pseudofossulae; in one specimen, they are united in a partial ring within the tabularium. The minor septa traverse all, nearly all or half the dissepimentarium. In its inner part, they may be divided into segments or contratingent; they are rarely entering into the tabularium.
84The dissepimentarium consists of 13 to 20 or only 5 to 10 rows of small dissepiments usually arranged in horizontal layers at the periphery and inclined in its inner part. Coarse and isolated trabeculae are present in the outer dissepimentarium of some coralla. The tabularium is filled with vesicular tabellae occasionally disrupted by axial ends of thick and discontinuous septa.
85There are 88 to 110 septa per corallum. The diameter of the corallum ranges from 29 mm to 49 mm. The width of the tabularium varies between 11 mm and 21 mm.
86Discussion. The material of Meerbüsch quarry is similar to the German one investigated by Schulz (1883), Ma (1956) and Birenheide (1964, 1978). The specimen of Meerbüsch quarry illustrated by Nose et al. (2017) corresponds to a rather young stage whereas another one from the Upper Eifelian Junkerberg Formation of the Eifel Hills figured by Schröder (1998) is different by its smaller size, poorly developed septa and vesicular tabellae with a convex pattern. The Russian species Atelophyllum tenuiseptatum (Bulvanker, 1958, p. 74) from the Middle Givetian of the Kuznetsk Basin seems to be close to A. laeve though it may have slightly less septa.
87Distribution. The material of Meerbüsch quarry comes from the top of the Lower Givetian Binz Member, in the lower part of the Dreimühlen Formation. In the Eifel Hills, Atelophyllum laeve is mostly known in this lithostratigraphic unit as mentioned by Birenheide (1978).
88Atelophyllum cf. maximum (Schlüter, 1882)
89(Plate 3A–B)
90cf. 1889 Mesophyllum maximum Schlüt.; Schlüter, p.70, pl. 7, fig. 1.
91cf. 1926 Mochlophyllum maximum Schlüter; Vollbrecht, pl. 15, fig. 2.
92cf. 1964 Plasmophyllum (Mesophyllum) maximum maximum (Schlüter 1882); Birenheide, p. 43, pl. 7, figs 30–31.
93Lectotype. Specimen WDKD 2590 chosen by Birenheide (1964, p. 42) and stored in the Forschungsinstitut Senckenberg at Frankfurt am Main, Germany. Upper Eifelian Junkerberg Formation of Auburg in the Gerolstein Syncline, Eifel Hills in Germany. Pl. 15, fig. 2 in Vollbrecht (1926), pl. 7, fig. 30 in Birenheide (1964), fig. 105 in Birenheide (1978) and fig. 63, 2a in Hill (1981).
94Material. 3 specimens with 6 thin sections: Meerbüsch EIF404, EIF406 and EIF423.
95Diagnosis. A species of Atelophyllum with 96 to 122 septa at a diameter of 70 mm to 108 mm. Long major and minor septa relatively thin in the outer part of the dissepimentarium. Small open space in the centre of the tabularium.
96Description. The material consists of fragmentary, cylindrical, and ceratoid solitary rugose corals whose height varies between 4 cm and 10 cm. Two specimens are characterized by an elliptical transverse section whereas another one shows longitudinal ribs. The outer wall is rarely preserved and locally encrusted by a thin laminar stromatoporoid in one sample.
97The septa may be thin and discontinuous at the periphery with some strong yardarm carinae. Then the septa are non-carinate and dilated throughout their length. They are more thickened in the tabularium or only at its entry and at the inner border of the dissepimentarium. In a rather young stage, the septa are more discontinuous at the periphery and they are affected by a more important thickening in the rest of the transverse section.
98The major septa reach more or less the centre of the tabularium; their axial ends are sometimes rhopaloid or divided into segments. The minor septa traverse all or nearly all the dissepimentarium; occasionally, they are discontinuous in its inner part or they are hardly projecting into the tabularium. In the rather young stage, the minor septa may be shorter and more variable in length.
99The dissepimentarium consists of numerous rows of small, inclined dissepiments which are sometimes arranged in horizontal layers at the periphery. Long and isolated septal trabeculae may be present in the outer part of the dissepimentarium. The tabularium is narrow and occupied by vesicular tabellae intersected by axial ends of thick septa.
100There are 110 to 112 or even 134 septa per corallum. The diameter of the corallum ranges commonly from 45 mm to 55 mm and more generally from 33 mm to 67 mm. The width of the tabularium varies between 10 mm and 14 mm.
101Discussion. Atelophyllum cf. maximum shows some similarities with A. laeve from which it is separated by several features: a larger corallum with slightly more septa, a narrower tabularium, the weaker dilation of the septa though it is still present in the tabularium, smaller and more numerous dissepiments, commonly longer major and minor septa. Some specimens of A. laeve are closer to A. cf. maximum as their major septa reach the axis of the corallum, but they are thinning in the tabularium, what is not the case for the form described herein. A. laeve was considered by Birenheide (1964, 1978) as a subspecies of A. maximum which is characterized by a great size and many septa as A. cf. maximum. However, A. maximum is different in having more discontinuous septa at the periphery as well as less thickened septa throughout their length and leaving a small open space in the centre of the tabularium. It must also be mentioned that the type material of A. maximum from the Upper Eifelian of the Eifel Hills is only known in transverse sections. A few authors have assigned to A. maximum coralla very close to the form of Meerbüsch quarry in having septa relatively continuous at the periphery, evenly thickened throughout their length and reaching or not the centre of the tabularium. Such specimens have been figured from the Upper Givetian of the Guizhou Province in China, by Liao & Birenheide (1984, p. 6) and from the Middle Devonian of the Eifel Hills, by Scrutton (1985, pl. 3.3.13); the material from the Middle Givetian of Leon in Spain investigated by Birenheide & Soto (1981, p. 258) is also related to these coralla.
102Atelophyllum robertsense (Merriam, 1973, p. 27) coming from the Lower Givetian of Nevada in USA has some affinities with A. cf. maximum; however, it differs by thinner septa and a small open space in the centre of the tabularium. The septa are more dilated in A. alhambraense (Merriam, 1973, p. 27), from the Lower Givetian of the same area, which is distinguished from A. cf. maximum by more septa and a greater diameter of the corallum.
103Distribution. The questionable material of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation. In the Eifel Hills, Atelophyllum maximum is mainly represented in the Upper Eifelian Junkerberg Formation, according to Birenheide (1978, p. 168).
104Atelophyllum cf. emsti (Wedekind, 1922)
105(Plate 4A–B)
106cf. 1922a Mesophyllum emsti Wdkd.; Wedekind, p. 57, pl. 2, fig. 1.
107cf. 2019 Atelophyllum emsti (Wedekind, 1922); Coen-Aubert, p. 126, pl. 2A–C, pl. 3A–D).
108cf. 2021 Lekanophyllum emsti (Wedekind, 1922); McLean, pl. 21, figs 1, 4–6.
109Holotype. Specimen and thin sections WDKD 45034506 and 97299730 stored in the Forschungsinstitut Senckenberg at Frankfurt am Main, Germany. Lower Givetian Oberhonsel Formation at Emst near Hagen in North Sauerland, Germany. Pl. 2, fig. 1 in Wedekind (1922a), figs 64, 1g–h in Hill (1981) and pl. 21, figs 1, 4–6 in McLean (2021).
110Material. 1 specimen with 2 thin sections: Meerbüsch EIF431.
111Diagnosis. See Coen-Aubert (2019).
112Description. The material is restricted to a cylindrical corallum with longitudinal ribs, which is 6 cm high. The outer wall is very locally preserved.
113The septa are discontinuous in a wide peripheral zone with a few yardarm carinae. Then, they are dilated throughout their length or they are sometimes thinning at their axial ends. The major septa leave a small open space in the centre of the tabularium. The minor septa are reduced to short segments in the inner part of the dissepimentarium.
114The dissepimentarium consists of small, inclined dissepiments, occasionally arranged in horizontal layers at the periphery. Coarse and isolated trabeculae extend across the entire dissepimentarium. The axial tabellae are vesicular with an overall horizontal pattern. A few spots of thin stereoplasma occur at the inner border of the dissepimentarium.
115There are about 90 septa for a diameter of 39 mm to 42 mm whereas the width of the tabularium measures 17 mm to 23 mm.
116Discussion. The specimen of Meerbüsch quarry is related to Atelophyllum emsti whose septa are more strongly dilated in the dissepimentarium and the minor ones not so frequently represented in its inner part. The form also resembles A. amanshauseri (Wedekind, 1925) from the Givetian Oberhonsel or Schwelm Formation of North Sauerland in Germany which is characterized by a smaller septal number and diameter of the corallum.
117Distribution. The questionable corallum of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation. In Germany, Atelophyllum emsti is only known in the Lower Givetian Oberhonsel Formation from the Sauerland.
118In Belgium, the species has been investigated by Coen-Aubert (2019), in the Lower to Middle Givetian from the south side of the Dinant Synclinorium where it has been collected in the lower part of the Mont d’Haurs Formation.
119Family Ptenophyllidae Wedekind, 1923
120Genus Acanthophyllum Dybowski, 1873
121Type species. By subsequent designation of Schlüter (1889, p. 38), Cyathophyllum heterophyllum Milne-Edwards & Haime, 1851.
122Diagnosis. See Coen-Aubert (2017).
123Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851)
124(Plate 4C–D)
125v * 1851 Cyathophyllum heterophyllum; Milne-Edwards & Haime p. 367, pl. 10, figs 1, 1a–b.
126v 1997 Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851); Coen-Aubert, p. 11, pl. 1, figs 1–4, pl. 2, figs 57.
127v 2011 Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851); Coen-Aubert, p.36, pl. 2, fig. 6.
128v 2017 Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851); Coen-Aubert, p. 163, pl. 1A–B.
1292020 Acanthophyllum heterophyllum (Milne Edwards & Haime, 1851); Jamart & Denayer, figs 5E–F.
130Remarks. More complete lists of synonymy with the references before 1997, 2011 and 2017 have been provided by Coen-Aubert (1997, 2011, 2017).
131Holotype. See Coen-Aubert (2017).
132Material. 1 specimen with 2 thin sections: Meerbüsch EIF391.
133Diagnosis. See Cen-Aubert (2017).
134Description. The material is restricted to a fragmentary cylindrical corallum which is 3 cm high. The outer wall is preserved very locally.
135The septa are thin at the periphery and strongly dilated in the inner part of the dissepimentarium; this thickening affects more the major septa. Carinae are present at the border of the tabularium in which the septa are still carinate and relatively thick. The major septa reach the axis of the corallum and their axial ends are typically rhopaloid. The minor septa traverse the entire dissepimentarium or even enter into the tabularium where they are occasionally contratingent.
136The dissepimentarium consists of 8 to 14 rows of inclined dissepiments which are sometimes subhorizontal at the periphery. The tabulae are incomplete and intersecting laterally; they are disrupted by fragments of thick septa with spinose carinae.
137There are 76 septa for a diameter of 28 mm to 32 mm whereas the width of the tabularium measures 10 mm to 11.5 mm.
138Discussion. The specimen shows all the features characteristic of Acanthophyllum heterophyllum though it differs slightly by its septa only strongly dilated in the inner part of the dissepimentarium and slender in a rather wide zone at the periphery. Nevertheless, it falls within the variability of the species illustrated by the various German taxa of Wedekind (1924) considered as synonyms by Birenheide (1961, 1962) and also mentioned by Coen-Aubert (1997).
139Distribution. The corallum of Meerbüsch quarry comes from the top of the Lower Givetian Binz Member, in the lower part of the Dreimühlen Formation. For the rest, Acanthophyllum heterophyllum is widely distributed in the Eifelian and the Lower Givetian of the Eifel Hills.
140Outside Germany, the species is common in the Upper Eifelian and the Lower Givetian from the south side of the Dinant Synclinorium in Belgium and the North of France as well as from the Ma’der in Morocco. It is also known in the Upper Emsian of the Southeastern Armorican Massif in France, in the Lower Eifelian of the Holy Cross Mountains in Poland and in the lower part of the Givetian from the Zemmour in Mauritania.
141Acanthophyllum vermiculare (Goldfuss, 1826)
142(Plate 2C; Plate 3C–I)
143v * 1826 Cyathophyllum vermiculare nobis; Goldfuss, p. 58, pl.17, fig. 4.
144v 1997 Acanthophyllum vermiculare (Goldfuss, 1826); Coen-Aubert, p. 13, pl. 3, figs 1–8.
145v pp. 1997 Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851); Coen-Aubert, p.11 (non pl. 1, figs 1–4, pl. 2, figs 5–7).
146v 1998 Acanthophyllum vermiculare (Goldfuss, 1826); Coen-Aubert, pl. 2, fig. 1.
147v 2011 Acanthophyllum vermiculare (Goldfuss, 1826); Coen-Aubert, p. 37, pl. 2, figs 1–5, pl. 3, fig. 9.
148v 2016 Acanthophyllum vermiculare (Goldfuss, 1826); Coen-Aubert in Brice, fig. 109D.
1492017 Acanthophyllum concavum (Walther, 1928); Nose et al., pl. 3, fig. 3.
1502020 Acanthophyllum vermiculare (Goldfuss, 1826); Jamart & Denayer, figs 5A–C.
1512020 Acanthophyllum sp. 1; Jamart & Denayer, figs 5G–H.
152pp. 2020 Dohmophyllum sp.; Jamart & Denayer, fig. 9G (non fig. 9F).
153Remark. More complete lists of synonymy with the references before 1997 and 2011 have been provided by Coen-Aubert (1997, 2011).
154Holotype. Specimen GMBo 198 from the Goldfuss collection stored in the Paläontologisches Institut from the University of Bonn in Germany. Probably fields lying to the west of the church of Nohn in the Hillesheim Syncline, Eifel Hills, Germany. Probably Müllert Member of the Ahbach Formation at the base of the Givetian. Pl. 17, fig. 14 in Goldfuss (1826), pl. 3, figs 3–4 in Gräf (1958) and pl. 2, fig. 1 in Coen-Aubert (1998).
155Material. 19 specimens with 35 thin sections: Meerbüsch EIF349, EIF370, EIF372, EIF385, EIF386, EIF405, EIF407, EIF412, EIF414, EIF415, EIF428, EIF434, EIF437, EIF444, EIF453, EIF464, EIF465, EIF466 and EIF467.
156Diagnosis. A species of Acanthophyllum with 68 to 80 septa at a diameter of 20 mm to 35 mm. Septa thin to slightly dilated throughout their length. Dissepiments often inclined.
157Description. The material consists of complete and often fragmentary solitary rugose corals which are ceratoid and occasionally cylindrical, conical or rarely trochoid; their height varies between 4 cm or even 2 cm and 7 cm. Longitudinal ribs and growth lines are observed in some specimens whereas a few ones are affected by rejuvenescence. In some coralla, the thin outer wall is locally preserved and possibly encrusted by laminar stromatoporoids or syringoporids.
158The septa are rather faintly dilated throughout their length. This dilation is highly variable among the material, but also within a single specimen. Indeed, the septa are sometimes thin and locally discontinuous at the periphery of a few coralla. But the major septa may also be more thickened in all or part of the dissepimentarium or only at its inner border. A dark median line affects the septa in the dissepimentarium of a few specimens. In the tabularium, the major septa are occasionally thin, relatively thick or only dilated at their axial ends. In some cases and especially in rather young stages, a deposit of stereoplasma may be present on a layer of dissepiments, mostly in the inner dissepimentarium. Spinose carinae are rare in the dissepimentarium and more frequent in the tabularium.
159The major septa reach more or less the axis of the corallum. Their axial ends are sometimes divided into fragments, rhopaloid, bifurcated, slightly twisted, bent as a hook or united laterally to form pseudofossulae. The minor septa traverse the entire and wide dissepimentarium or even enter into the tabularium where they may be contratingent. The minor septa are rarely shorter or discontinuous at the periphery and in the dissepimentarium.
160The dissepimentarium consists of 10 to 16 or even 8 to 18 rows of inclined dissepiments which are occasionally arranged in horizontal layers at the periphery. Some spots of fine stereoplasma formed by thin and contiguous septal trabeculae occur in the dissepimentarium. The tabulae are closely spaced and incomplete to vesicular with a concave pattern; they are disrupted by thick septa and segments of them with spinose carinae.
161There are usually 70 to 84 septa per corallum, but this septal number reaches 90 to 96 in a few specimens. The diameter of the corallum ranges from 18 mm to 38 mm. The width of the tabularium varies commonly between 7 mm and 10.5 mm and more generally between 5.9 mm and 13.5 mm.
162Discussion. As a whole, the material of Meerbüsch quarry is similar to that from other localities of the Eifel Hills figured among others by Wedekind (1925), Birenheide (1961), Lütte (1984) and Coen-Aubert (1998) and to the Belgian one investigated by Coen-Aubert (1997) and Jamart & Denayer (2020). In fact, some specimens illustrated by these two authors as Dohmophyllum sp. and Acanthophyllum sp. 1 belong also to A. vermiculare. Nevertheless, the samples of Meerbüsch quarry slightly differ from the latter species by septa more thickened in some specimens (Plate 3D). These specimens recall Acanthophyllum heterophyllum where however the major septa are more strongly dilated than the minor ones in the main part of the dissepimentarium. The coralla of Meerbüsch quarry with relatively more thickened septa can be compared with A gerolsteinense (Wedekind, 1924), A. gerolsteinense crassum (Wedekind, 1924), A. intortum (Wedekind, 1923) and A. praematurum (Wedekind, 1923) from the Eifelian of the Eifel Hills. Birenheide (1962) placed these four taxa in synonymy with A. torquatum (Schlüter, 1884) from the Eifelian of the same area. As discussed by Schröder & Kazmierczak (1999, p. 103) and Coen-Aubert (2011, p. 38), the lectotype of A. torquatum figured by Schlüter (1889) is probably conspecific with A. vermiculare. Therefore, the material of Meerbüsch quarry is very interesting in showing the wide variability that affects the thickness of the septa in the species A. vermiculare. It must be added that such variability is also present among the Belgian specimens (Plate 2C) so that two of them have been assigned to A. heterophyllum by Coen-Aubert (1997, p. 11).
163Nose et al. (2017) referred to A. concavum (Walther, 1929) a small corallum of A. vermiculare collected at the top of the Binz Member, in the Meerbüsch quarry. Acanthophyllum concavum has been synonymized by Coen-Aubert (2000) with A. simplex (Walther, 1929) and both taxa come from the Givetian of the Bergisches Land in Germany. Acanthophyllum simplex is separated from A. vermiculare by smaller septal number and diameter of the corallum and by septa more dilated in the entire dissepimentarium.
164Distribution. The material of Meerbüsch quarry comes from the top of the Binz Member and from the base of the Galgenberg Member, in the Lower Givetian Dreimühlen Formation. In the Eifel Hills, Acanthophyllum vermiculare is common from the Upper Eifelian Junkerberg Formation to the Lower Givetian Kerpen Formation.
165Outside Germany, the species is abundant on the south side of the Dinant Synclinorium, in Belgium and the North of France, where it occurs close to the Eifelian–Givetian boundary, in the Hanonet Formation and at the base of the Trois-Fontaine Formation. Acanthophyllum vermiculare has also been observed in the Middle Devonian part of the Chalonnes Formation from the Southeastern Armorican Massif in France and in the Lower Givetian from the Ma’der in Morocco.
166Genus Grypophyllum Wedekind, 1922
167Type species. By original designation, Grypophyllum denckmanni Wedekind, 1922.
1681922b Grypophyllum Wedekind, p. 13.
1691951 Hooeiphyllum Taylor, p. 173.
170Diagnosis. Solitary rugose corals bordered by a thin or relatively thick wall. Septa of two orders, non-carinate, thin or slightly dilated throughout their length, occasionally discontinuous or disrupted by lonsdaleoid dissepiments at the periphery. Major septa extending to or nearly to the axis of the corallum. Minor septa variable in length and development. Dissepimentarium moderately wide with several rows of inclined dissepiments. Tabulae incomplete and closely spaced, commonly forming concave floors.
171Grypophyllum convolutum (Wedekind, 1925)
172(Plate 4E–G)
173* 1925 Stenophyllum convolutum Amanshauser; Wedekind, p. 10, pl. 2, fig. 8.
174non 1952 Stringophyllum convolutum (Amanshauser); Soshkina, p. 95, pl. 32, fig. 114.
175non 1959 Stenophyllum convolutum (Amanshauser), 1920; Spassky, p. 38, fig. 15.
1761962 Stenophyllum convolutum Wedekind 1925; Birenheide, p. 104.
177non 1969 Grypophyllum convolutum (Amanshauser), in Wedekind, 1925; Tsien, p. 122, pl. 47, fig. 2.
178pp. 1972 Grypophyllum convolutum (Wedekind 1925); Birenheide, p. 418, pl. 3, fig. 11, pl. 4, figs 12–14 (non pl. 5, fig. 15).
1791978 Grypophyllum convolutum (Wedekind 1925); Birenheide, p. 137, fig. 81.
180non 1983 Leptoinophyllum convolutum (Wedekind); Cao et al., p. 144, pl. 51, fig. 4.
1811985 Grypophyllum convolutum (Wedekind); Scrutton, p. 23, pl. 3.3.41.
182non 1998 Grypophyllum convolutum Wedekind 1925; Schröder, p. 49, pl. 8, figs 56–57.
1831999 Grypophyllum convolutum (Wedekind, 1925); Schröder & Kazmierczak, p. 101, pl. 2, fig. 11.
184non 2001 Grypophyllum sp. ex gr. Gruppe convolutum/denckmanni; Schröder & Salerno, pl. 3, figs 25–26.
185Lectotype. Specimen and thin sections WDKD 4237-4238 chosen by Birenheide (1962, p. 104) and stored in the Forschungsinstitut Senckenberg at Frankfurt am Main, Germany. Lower Givetian Oberhonsel Formation at Heggen-Sange, Attendorner Mulde, Sauerland, Germany. Pl. 2, fig. 8 in Wedekind (1925) and pl. 4, fig. 13 in Birenheide (1972).
186Material. 2 specimens with 4 thin sections: Meerbüsch EIF416 and EIF433.
187Diagnosis. A species of Grypophyllum with 62 to 78 septa at a diameter of 27 mm to 36 mm. Septa slender throughout their length. Minor septa variable, but rather long. Numerous rows of inclined dissepiments.
188Description. The material consists of two fragmentary, cylindrical and ceratoid solitary coralla with a height between 4.5 cm and 5 cm. The outer wall is thin and relatively well preserved; it is encrusted by laminar stromatoporoids and auloporids.
189The septa are non-carinate. They are occasionally discontinuous at the periphery with a few presepiments. The septa are slender or slightly dilated throughout their length. They are locally affected by a dark median line in the dissepimentarium of one specimen.
190The major septa reach the axis of the corallum or leave a small open space in the centre of the tabularium; there axial ends are more or less twisted in a whorl. The minor septa are variably developed, traversing all or part of the dissepimentarium or hardly entering into the tabularium where they are sometimes contratingent. They may also be lacking, reduced to spines or discontinuous at the periphery and in the dissepimentarium.
191The dissepimentarium consists of 6 to 12 rows or relatively large dissepiments which are variably inclined. The tabulae are incomplete and intersecting laterally; they are disrupted by axial ends of major septa.
192There are 72 to 76 septa per corallum. The diameter of the corallum ranges from 24 mm to 30 mm whereas the width of the tabularium varies between 8.6 mm and 14.5 mm.
193Discussion. The two specimens of Meerbüsch differ slightly by minor septa variably developed. They are similar to the lectotype of the species and to the German material described by Birenheide (1972, 1978). As explained by Coen-Aubert (2011, p. 38) however, one corallum of Birenheide (1972, pl. 5, fig. 15) belongs in fact to Acanthophyllum vermiculare praecursor (Frech, 1886) and it is mainly distinguished from Grypophyllum convolutum by continuous and systematically long minor septa. The samples of Schröder (1998) and Schröder & Salerno (2001) from the Lower Givetian of the Eifel Hills are excluded from the synonymy list due to their smaller diameter and septal number and to their relatively thick outer wall.
194Other references to G. convolutum are different from the species for various reasons. The transverse section from the Givetian of Armenia illustrated by Soshkina (1952) is rather large with more septa whereas the material of Cao et al. (1983) from the Devonian of northwest China has a smaller diameter. The corallum from the Givetian of the Urals in Russia figured by Spassky (1959) lacks minor septa. Finally, Tsien (1969) assigned to G. convolutum one transverse section from the base of the Givetian in southern Belgium with less septa which are irregular at the periphery and slightly dilated in the main part of the dissepimentarium.
195Distribution. The material of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation. In Germany, Grypophyllum convolutum is also known in the Givetian Loogh and Cürten Formations from the Eifel Hills and in the Lower Givetian Oberhonsel Formation from the Sauerland in Germany.
196Outside Germany, the species occurs in the Givetian of South Devon in Great Britain and in the Lower Givetian of the Ma’der in Morocco.
197Grypophyllum schroederi n.sp.
198(Plate 4H–J; Plate 5A–B)
199Derivation of name. The species is dedicated to Stefan Schröder, a distinguished German specialist of Devonian rugose corals.
200Holotype. IRScNB a 13545 (= Plate 4I–J). Specimen EIF449 collected by M. Coen-Aubert in 2000, in the Lower Givetian of the Eifel Hills, Germany.
201Type locality and horizon. Meerbüsch quarry in the Hillesheim Syncline, Eifel Hills in Germany. Base of the Lower Givetian Ley Member, in the middle part of the Dreimühlen Formation.
202Material. 6 specimens with 8 thin sections: Meerbüsch EIF436, EIF446, EIF447, EIF448, EIF449 and EIF450.
203Diagnosis. A species of Grypophyllum with 56 to 62 septa at a diameter of 15 mm to 23 mm. Septa rather slender throughout their length. Minor septa variable, but relatively short.
204Description. The material consists of fragmentary, conical and trochoid coralla, often with longitudinal ribs, whose height varies between 1.5 cm and 4 cm. The outer wall is thick and usually continuous. Several specimens are encrusted and enveloped by skeletons of thick laminar stromatoporoids which may enclose small sections of rugose and tabulate corals.
205The septa are non-carinate, thin to slightly dilated throughout their length. Occasionally, they are discontinuous at the periphery or affected by a triangular thickening against the outer wall. The major septa reach the centre of the tabularium; their axial ends are sometimes twisted, weakly rhopaloid or forming pseudofossulae. The highly variable minor septa are lacking, short, reduced to spines, discontinuous, or more or less long in the dissepimentarium where they are sometimes contratingent.
206There are only two longitudinal sections available. The dissepimentarium consists of 5 to 10 rows of variably inclined dissepiments. The narrow tabularium is filled with incomplete tabulae intersected by axial ends of major septa.
207There are 56 to 62 or even 68 septa per corallum. The diameter of the corallum ranges from 14.5 mm to 26 mm. The width of the tabularium varies between 4.5 mm and 8.3 mm.
208Discussion. Grypophyllum schroederi is related to G. denckmanni type species of the genus which comes from the Givetian of the Bergisches Land in Germany and has been revised by Coen-Aubert (2000). Nevertheless, the latter species differs from the former by slightly less septa, rather long minor septa, the possible occurrence of a stereozone against the outer wall and by septa rarely discontinuous at the periphery. Grypophyllum schroederi is separated from G. convolutum by smaller septal number and diameter of the corallum and by shorter minor septa. There are many similarities between G. schroederi and the specimens collected in the Cürten to Dreimühlen Formations from the Eifel Hills which have been illustrated by Schröder & Salerno (2001, pl. 3, figs 23, 25–26) as G. sp. aff. wedekindi Middleton, 1959 and G. sp. ex Gruppe convolutum/denckmanni; however, these coralla have fewer septa for nearly the same diameter. Grypophyllum wedekindi is a replacement name for G. gracile Wedekind, 1925 from the Givetian of the Bergisches Land that is characterized by a smaller size and by minor septa reduced to spines or lacking. Grypophyllum arcticum Bulvanker in Bulvanker et al. (1968) from the Givetian of Novaya Zemlya in Russia is also close to G. schroederi, but it is characterized by better developed minor septa.
209Distribution. The new species is only known in Meerbüsch quarry where it has been collected at the bases of the Lower Givetian Ley and Galgenberg Members, in the middle part of the Dreimühlen Formation.
210Family Stringophyllidae Wedekind, 1922
211Genus Stringophyllum Wedekind, 1922
212Type species. By subsequent designation of Wedekind (1925, p. 64), Stringophyllum normale Wedekind, 1922.
213Diagnosis. Solitary rugose corals. Septa of two orders, relatively thick and consisting of coarse monacanthine trabeculae. Septa sometimes continuous from the wall to the axis of the corallum, but more often discontinuous or even disrupted at the periphery by presepiments. Major septa bilaterally arranged and reaching usually the centre of the tabularium where they are occasionally breaking up into isolated trabeculae. Minor septa discontinuous, represented by short segments and spines which may be lacking totally. Dissepimentarium composed of several rows of elongate dissepiments. Tabulae complete or incomplete, normally concave.
214Stringophyllum acanthicum (Frech, 1885)
215(Plate 5C–E)
216* 1885 Endophyllum acanthicum nov. sp.; Frech, p. 929, pl. 41, fig. 5.
2171925 Schizophyllum acanthicum Frech; Wedekind, p. 60, pl. 13, fig. 78.
218v 2011 Stringophyllum acanthicum (Frech, 1885); Coen-Aubert, p. 31, pl. 1, figs 1–6, pl. 2, figs 8–9, pl. 3, fig. 4.
2192017 Stringophyllum acanthicum (Frech, 1885); Nose et al., pl. 3, fig. 1.
2202020 Stringophyllum acanthicum (Frech, 1885); Jamart & Denayer, p. 296, figs 10A–B.
221pp. 2020 Stringophyllum buechelense (Schlüter, 1889); Jamart & Denayer, p. 296, figs 10C–D (non figs 10E–G).
222Remark. A more complete list of synonymy for the references before 2011 has been provided by Coen-Aubert (2011).
223Holotype. Specimen from the Middle Devonian of Blankenheim in the Eifel Hills, Germany which was probably stored in the Museum für Naturkunde from the Humboldt University of Berlin in Germany, according to Schröder (1995a, p. 398). Pl. 41, fig. 5 in Frech (1885) and pl. 13, fig. 78 in Wedekind (1925).
224Material. 2 specimens with 4 thin sections: Meerbüsch EIF425 and EIF426.
225Diagnosis. A species of Stringophyllum with 84 to 106 septa at a diameter of 18 mm to 33 mm. Septa rather discontinuous at the periphery. Minor septa variably developed in the inner part of the dissepimentarium.
226Description. The material consists of two fragmentary ceratoid and cylindrical coralla with a height of 3 cm. The outer wall is relatively well preserved and encrusted by a thin laminar stromatoporoid in one sample.
227The septa are non-carinate and dilated throughout their length. They are discontinuous at the periphery or locally separated from the outer wall by a few rows of presepiments. The major septa reach nearly the centre of the tabularium where there are rare coarse trabeculae. In one specimen, the minor septa are systematically developed in the inner part of the dissepimentarium; sometimes, they are discontinuous or they are projecting into the tabularium. In the other specimen, the minor septa are lacking or represented by rare spines and segments; the latter ones are restricted to the inner dissepimentarium and to the entry of the tabularium. Stereoplasma is often present on a row of inner dissepiments where it may form a more or less continuous ring.
228The dissepimentarium consists of 2 to 6 rows of large inclined dissepiments which are occasionally subhorizontal at the periphery. In one corallum, some spots of coarse and contiguous septal trabeculae are observed in the inner part of the dissepimentarium. The tabulae are closely spaced with a concave pattern; they are disrupted by thickened septa.
229There are 80 to 86 septa per corallum. The diameter of the corallum ranges from 16 mm to 19 mm whereas the width of the tabularium varies between 8 mm and 10 mm.
230Discussion. The two specimens described herein are relatively small representatives of Stringophyllum acanthicum with a variable development of the minor septa as is the case for the material of Coen-Aubert (2011) collected in southern Belgium, across the Eifelian–Givetian boundary. Jamart & Denayer (2020, figs 10C–D) assigned to S. buechelense (Schlüter, 1889) a corallum of S. acanthicum from the same area. As was explained by Coen-Aubert (2011), S. buechelense is not a synonym of S. acanthicum. Indeed, the former species from the Givetian of the Bergisches Land in Germany is easily separated from the latter by slightly smaller septal number and diameter of the corallum, by poorly developed minor septa and by septa more discontinuous at the periphery. The lectotype of S. buechelense has been refigured by Engel & Von Schouppé (1958, pl. 8, fig. 9) and by Schröder (2005, pl. 10, fig. 9a).
231Distribution. The two coralla of Meerbüsch quarry investigated herein come from the base of the Galgenberg Member whereas the specimen from the same outcrop figured by Nose et al. (2017) has been collected at the top of the Binz Member, also in the Lower Givetian Dreimühlen Formation. In Germany, Stringophyllum acanthicum is certainly known in the Upper Eifelian Freilingen Formation and in the Lower Givetian Rodert Formation from the Eifel Hills, in the Givetian Schwelm Formation from the Sauerland and in the Givetian from the Bergisches Land.
232Outside Germany, the species occurs on the south side of the Dinant Synclinorium in Belgium where it is present across the Eifelian–Givetian boundary, in the Jemelle and Hanonet Formations and at the base of the Trois-Fontaines Formation. Stringophyllum acanthicum has also been found in the Middle Devonian part of the Chalonnes Formation from the Southeastern Armorican Massif in France, in the Givetian from Moravia in the Czech Republic and in the Upper Eifelian from the Ma'der and the Moroccan Meseta in Morocco. It may also occur in the Givetian of the Urals in Russia.
233Family Cyathophyllidae Dana, 1846
234Genus Heliophyllum Hall in Dana, 1846
235Type species. By original designation, Heliophyllum halli Milne-Edwards & Haime, 1850 as replacement name for Cyathophyllum helianthoides Goldfuss, 1826 sensu Hall (1843, p. 209).
236Diagnosis. Solitary or weakly fasciculate rugose corals. Septa of two orders long and thin throughout their length in the adult stages with mainly yardarm carinae in the dissepimentarium. Septa possibly dilated and contiguous in the young stages. Minor septa traversing the entire dissepimentarium. Dissepimentarium composed of several rows of globose dissepiments arranged in horizontal layers in its outer part and inclined towards the axis of the corallum in its inner part. Tabulae usually incomplete.
237Heliophyllum cf. cribellum Oliver & Sorauf, 2002
238(Plate 5F–H)
239cf. 2002 Heliophyllum cribellum new species; Oliver & Sorauf, p. 29, pls 30–31.
240Holotype. Specimen USNM 518586 stored in the U.S. National Museum of Natural History, Smithsonian Institution at Washington DC, USA. Lower Givetian Centerfield Member of the Ludlowville Formation to the east of East Alexander, Genesee County, New York State, USA. Pl. 30, figs 1–8 in Oliver & Sorauf (2002).
241Material. 2 specimens with 4 thin sections: Meerbüsch EIF371 and EIF 435.
242Diagnosis. A solitary species of Heliophyllum with 44 to 68 septa at a diameter of 10 mm to 20 mm. Septa of both orders thin with abundant yardarm carinae and mostly restricted to the dissepimentarium.
243Description. The material consists of two fragmentary conical and cylindrical coralla with a height between 2.5 cm and 5 cm. They are affected by longitudinal ribs, growth rings and rejuvenescences. The thin outer wall is locally preserved.
244The septa are thin throughout their length with strong yardarm carinae in the dissepimentarium or at least in its outer part. In one specimen, they are very locally dilated in the inner part of the dissepimentarium with a little stereoplasma between the septa. Both orders of septa are mainly restricted to the dissepimentarium and they are sometimes discontinuous at their inner ends. However, some minor septa are slightly shorter whereas some major septa hardly enter into the tabularium.
245The dissepimentarium consists of 7 to 10 rows of small globose dissepiments arranged in horizontal layers in its outer part and inclined in its inner part. Coarse septal trabeculae which are frequently isolated and only locally contiguous, extend across the entire dissepimentarium. The tabulae are incomplete and intersecting laterally; they are occasionally horizontal or convex.
246There are 62 to 64 septa per corallum. The diameter of the corallum ranges from 23 mm to 25 mm whereas the width of the tabularium varies between 11 mm and 14 mm.
247Discussion. The two specimens of Meerbüsch quarry differ from Heliophyllum cribellum by slightly larger coralla with about the same number of septa. Additionally, the major septa of the American material are often longer in the tabularium. The same feature affects the paratype of H. rhopaliseptatum Stumm, 1968 from the Lower Givetian Tenmile Creek Formation of northwestern Ohio in USA; this taxon is also characterized by more septa and a greater diameter of the corallum. The sampling from the Givetian of the Pyrenees in France assigned to H. rhoplaliseptatum by Joseph & Tsien (1975, pl. 3, fig. 6) is particular due to some thickening of the septa in the dissepimentarium.
248Ma (1956, p. 51) introduced the species H. breviseptatum from the Middle Devonian of Gerolstein in the Eifel Hills which is only known by its holotype and is separated from the coralla of Meerbüsch quarry by its smaller size and less septa and by septa dilated and strongly carinate in a narrow dissepimentarium. On the contrary, H. dahlemense (Haller, 1936) and its synonym H. halleri Schröder, 1995b from the Eifelian of the Eifel Hills are very different from H. cf. cribellum. Indeed, both species are characterized by greater septal number and diameter of the corallum, longer major septa often reaching the centre of the tabularium and some local dilation of the septa in the dissepimentarium. The type material of H. dahlemense has been revised by Coen-Aubert (1996).
249Distribution. The questionable material of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation. For the rest, Heliophyllum cribellum is only known in the Lower Givetian Centerfield Member of the Ludlowville Formation from the New York State in USA.
250Family Disphyllidae Hill, 1939
251Genus Disphyllum De Fromentel, 1861
252Type species. By subsequent designation of Lang & Smith (1934, p. 80), Cyathophyllum caespitosum Goldfuss, 1826.
253Diagnosis. Fasciculate rugose corals. Septa of two orders, occasionally carinate, variably dilated in the dissepimentarium and thin in the tabularium. Major septa reaching the axis of the corallites or leaving an open space in the centre of the tabularium. Minor septa traversing the entire dissepimentarium. Dissepimentarium composed of several rows of globose dissepiments, often arranged in horizontal layers in its outer part and inclined towards the axis of the corallites in its inner part. Tabulae usually incomplete or compound.
254Disphyllum caespitosum (Goldfuss, 1826)
255(Plate 5I–M)
256v * pp. 1826 Cyathophyllum caespitosum nobis; Goldfuss, p. 60, pl. 19, fig. 2b (non figs 2a, 2c–d).
2571998 Disphyllum caespitosum (Goldfuss 1826); Schröder, p. 41, pl. 5, fig. 35.
258v 2008 Disphyllum caespitosum (Goldfuss 1826); Coen-Aubert, p. 38, pl. 1, figs 1–2, pl. 2, figs 7–9, pl. 3, figs 7–8.
259non 2008 Disphyllum caespitosum (Goldfuss, 1826); Brownlaw & Jell, p. 31, fig. 17.
260non 2010 Disphyllum caespitosum (Goldfuss, 1826); Blake, p. 106, fig. 71.
261Remark. A more complete list of synonymy for the references before 2008 has been provided by Coen-Aubert (2008).
262Lectotype. Specimen GMBo 205 of the Goldfuss collection chosen by Lang & Smith (1934, p. 80) and stored in the Paläontologisches Institut from the University of Bonn in Germany. Probably Givetian of "Bensberg", Bergisch Gladbach in the Bergisches Land, Germany. Pl. 19, fig. 2b in Goldfuss (1826), pl. 35, figs 4–6 in Lang & Smith (1935), pl. 2, fig. 7 and pl. 5, fig. 14 in Birenheide (1969) and figs 169, 2a–d in Hill (1981).
263Material. 6 specimens with 9 thin sections: Meerbüsch EIF350, EIF351, EIF352, EIF353, EIF387 and EIF392.
264Diagnosis. A species of Disphyllum with 46 to 52 septa at a diameter of 7 mm to 12 mm. Major and minor septa long. Tabulae incomplete, usually with axial flat-topped parts.
265Description. The material consists of cylindrical corallites with longitudinal ribs and a height of 2 cm to 3 cm. There are also some small fragments of fasciculate colonies which have an area of 4 x 2 cm and a height of 4.5 cm. The outer wall is partially preserved. A dark median line may be present when the corallites are locally in lateral contact. The wall is sometimes encrusted by thin laminar stromatoporoids and bryozoa.
266The septa are normally non-carinate though a few spinose and knobbly carinae occur in some specimens. They are weakly dilated in the dissepimentarium and become usually thin in the tabularium or slightly beyond their entry into it. The major septa leave a small open space in the centre of the tabularium. The minor septa traverse the entire dissepimentarium or even enter into the tabularium where they may be contratingent; they are rarely shorter.
267The dissepimentarium consists of 3 to 5 or even 1 or 2 rows of small globose dissepiments arranged in horizontal layers in its outer part and inclined in its inner part. The tabulae are incomplete and intersecting laterally; sometimes their axial part is flat-topped or they are horizontal or concave.
268There are 42 to 52 septa per corallite. The diameter of the corallites ranges from 8 mm to 11 mm. The width of the tabularium varies between 5.3 mm and 7.3 mm.
269Discussion. The material from Meerbüsch quarry is similar to the Belgian one and to the lectotype of the species though it differs somewhat by incomplete tabulae with rather rare flat-topped parts. The Frasnian Australian colonies investigated by Brownlaw & Jell (2008) and Blake (2010) coming respectively from the Canning Basin and Queensland are excluded from the synonymy list. Quantitatively, they are characterized by slightly greater size and septal number. Qualitatively, the specimens illustrated by Brownlaw & Jell (2008) have few dissepiments which are large and sometimes peneckielloid whereas the samples of Blake (2010) show slender septa leaving a wide open space in the centre of the tabularium.
270Distribution. The material of Meerbüsch quarry comes from the top of the Lower Givetian Binz Member, in the lower part of the Dreimühlen Formation. In Germany, Disphyllum caespitosum is also known in the Givetian Cürten Formation of the Eifel Hills and in the Givetian of the Bergisches Land.
271Outside Germany, the species occurs in the Givetian of the Pyrenees in France and in the upper part of the Lower Givetian Trois-Fontaines Formation, on the south side of the Dinant Synclinorium in Belgium.
272Genus Aristophyllum Bulvanker, Spassky & Kravstov in Besprozvannykh et al. (1975)
273Type species. By original designation, Aristophyllum terechovi Bulvanker, Spassky & Kravstov in Besprozvannykh et al. (1975).
274Diagnosis. Solitary rugose corals. Septa of two orders non-carinate or faintly carinate, slender to variably dilated in the dissepimentarium and thin in the tabularium. Major septa leaving a wide open space in the centre of the tabularium. Minor septa traversing the entire dissepimentarium. Relatively narrow dissepimentarium composed of a few rows of inclined dissepiments which may be also arranged in horizontal layers at the periphery. Incomplete tabulae often with a broad axial part.
275Discussion. The genus Aristophyllum Bulvanker, Spassky & Kravstov in Besprozvannykh et al. (1975), whose type species is A. terechovi Bulvanker, Spassky & Kravstov in Besprozvannykh et al. (1975) from the Frasnian of the Taymyr Kolyma Province in Russia, has been used by Coen-Aubert (1997) for A. luetti instead of Glossophyllum Wedekind, 1924. Indeed, G. dohmi Wedekind, 1924, type species of the latter genus from the Eifelian of the Eifel Hills, is only known by a few transverse sections figured among others by Wedekind (1924, figs 109–112) and Birenheide (1978, pl. 11, fig. 5) and showing septa systematically dilated in the tabularium, which suggests that they do not belong to mature stages. Several authors such as McLean (2014) consider that Aristophyllum is a synonym of Sinodisphyllum Sun, 1958. However, its type species S. variabile Sun, 1958 from the Frasnian of the Hunan Province in China is characterized by major septa leaving an open space in the centre of the tabularium or reaching occasionally the axis of the corallum whereas its minor septa may be variably developed.
276It must be mentioned that McLean (2014, p. 35) did not assign Aristophyllum luetti to Sinodisphyllum. But he referred to Sinodisphyllum different Givetian species such as Temnophyllum menyouense Hill & Jell, 1970 from Western Australia, Pseudozaphrentis sirius Schröder, 2004 from northern Pakistan, Campophyllum litvinovitshae Soshkina, 1949 from the Urals in Russia, Temnophyllum zamkowae Wrzolek, 1993 from the Holy Cross Mountains in Poland and Mictophyllum schlueteri Birenheide & Lütte, 1990 from the Kerpen Formation of the Eifel Hills. All these taxa have minor septa of variable length and this feature is not typical for Sinodisphyllum and Aristophyllum.
277Aristophyllum luetti Coen-Aubert, 1997
278(Plate 6A–G)
279v 1969 Ceratophyllum soetenicum (Schlüter), 1885; Tsien, p. 52, pl. 7, figs 8–10.
280v * 1997 Aristophyllum luetti n. sp.; Coen-Aubert, p. 16, pl. 1, figs 5–7, pl. 2, figs 1–4.
2811998 Aristophyllum luetti Coen-Aubert 1997; Schröder, p. 36, pl. 4, figs 27–29.
2822014a Aristophyllum luetti Coen-Aubert, 1997; Abbasi et al., p. 131, figs 5i–j, 6a–d.
2832014b Aristophyllum luetti Coen-Aubert, 1997; Abbasi et al., p. 11, pl. 2, fig. 4.
2842014c Aristophyllum luetti Coen-Aubert, 1997; Abbasi et al., p. 245, figs 4E–F.
2852020 Aristophyllum luetti Coen-Aubert, 1997; Jamart & Denayer, figs 7G–H.
286Holotype. Specimen IRScNB a10553 (= Houyet MC-1981-5-Z2831) stored in the Collection of Palaeontology of the Institut royal des Sciences naturelles de Belgique at Brussels in Belgium. Lower Givetian Hanonet Formation from the railway section situated to the north of Pondrôme, on the south side of the Dinant Synclinorium in Belgium. Pl. 2, figs 1–2 in Coen-Aubert (1997).
287Material. 14 specimens with 26 thin sections: Meerbüsch EIF348, EIF384, EIF388, EIF390, EIF393, EIF454, EIF455, EIF456, EIF457, EIF458, EIF462, EIF469, EIF471 and EIF472.
288Diagnosis. A species of Aristophyllum with 56 to 70 septa at a diameter of 13 mm to 26 mm. Septa slender to slightly dilated in the dissepimentarium. Several rows of globose dissepiments arranged in horizontal layers at the periphery and inclined in the inner part of the dissepimentarium.
289Description. The material consists of cylindrical, ceratoid, trochoid and conical coralla which are complete or fragmentary and whose height varies between 2.5 cm and 7.5 cm. Longitudinal ribs and occasionally growth lines or even growth rings are observed. Two specimens are affected by rejuvenescence; one of them is compressed laterally whereas another one has a lateral offset. The outer wall is thin and only preserved locally.
290The septa are non-carinate or bear a few small irregular, knobbly or even yardarm carinae in part of the material. They are slender or slightly dilated in the dissepimentarium. The septa may also be thin at the periphery where they are locally discontinuous in one sample. Sometimes, the septa show a triangular thickening against the outer wall which is occasionally more thickened at this place. In several specimens, a layer of dissepiments is partly affected by a weak deposit of stereoplasma. The septa become thin or more rarely less dilated in the tabularium; however, they are often thick in the tabularium of one specimen.
291The major septa leave a more or less wide open space in the centre of the tabularium; they are only longer in a few coralla. Their axial ends are sometimes rhopaloid, discontinuous or forming pseudofossulae. The minor septa traverse the entire dissepimentarium or even enter into the tabularium where they may be contratingent; they are rarely shorter.
292The dissepimentarium consists of 2 to 8 or even 0 to 10 rows of globose dissepiments arranged in horizontal layers in its outer part and inclined in its inner part. Only inclined dissepiments are present on one side of several coralla. Some spots of coarse and usually contiguous trabeculae occur in the dissepimentarium. The tabulae are incomplete and intersecting laterally; they are occasionally disrupted by septa or fragments of them. Sometimes, the tabulae are horizontal concave, horizontal or vesicular and they are more frequently characterized by a wide, flat-topped axial part.
293There are 62 to 72 or even 76 septa per corallum. The diameter of the corallum ranges from 17 mm to 28 mm or even to 30 mm. The width of the tabularium varies commonly between 11 mm and 15 mm and more generally between 10.5 mm and 17 mm.
294Discussion. The material of Meerbüsch quarry differs somewhat from the Belgian one investigated by Coen-Aubert (1997) by slightly greater septal number and diameter of the corallum, by sometimes longer major septa and by the variability of the septal dilation in the dissepimentarium. Indeed, the septa of the German specimens studied herein may be slender throughout their length (Plate 6F), locally weakly dilated in the dissepimentarium (Plate 6A) or sometimes more strongly thickened in part of the latter (Plate 6E). A. soetenicum (Schlüter, 1885) from the Lower Givetian Loogh and Cürten Formations of the Eifel Hills is very close to A. luetti. Its lectotype has been selected by Lütte (1987) whose sampling is often characterized by slender septa in the dissepimentarium. However, in the transverse section figured by Schlüter (1889, pl. 3, fig. 5), the septa are thin at the periphery and slightly dilated in the inner part of the dissepimentarium. Additionally, A. soetenicum is distinguished from A. luetti by more septa and larger coralla. A. trochiforme (Lütte, 1990) from the Loogh to Kerpen Formations from the Eifel Hills is also related to A. luetti though it has septa faintly dilated and carinate within the dissepimentarium as well as a small size with slightly less septa. The species has been assigned by Lütte (1990) to the genus Cyathophyllum Goldfuss, 1826 whose type species is C. dianthus Goldfuss, 1826 from the Lower Givetian of the Eifel Hills and which is represented by fasciculate and massive colonies (Coen-Aubert, 1990).
295Distribution. The material of Meerbüsch quarry comes from the top of the Lower Givetian Binz Member, in the lower part of the Dreimühlen Formation. In the Eifel Hills, Aristophyllum luetti is also known in the Lower Givetian Loogh and Cürten Formations or even in the Rodert Formation.
296Outside Germany, the species is well represented on the south side of the Dinant Synclinorium, mainly in the Lower Givetian Hanonet Formation and locally at the base of the overlying Trois-Fontaines Formation. Some coralla of A. luetti have been collected in the Givetian of the Alborz Mountains in Iran.
297Aristophyllum sp.
298(Plate 6H)
299Material. 1 specimen with 1 thin section: Meerbüsch EIF438.
300Description. The material is restricted to a fragmentary solitary corallum with a height of 1.5 cm and only one transverse section. The outer wall is thin and more or less preserved.
301The non-carinate septa are slender or slightly dilated throughout their length; sometimes they are thinner in the tabularium or at the periphery. A weak deposit of stereoplasma is locally present on a layer of dissepiments. The major septa leave a wide open space in the centre of the tabularium where their axial ends form a pseudofossula. The minor septa traverse the entire and broad dissepimentarium or even enter into the tabularium where they may be contratingent.
302There are 88 septa for a diameter of 32 mm to 34 mm whereas the width of the tabularium measures 18 mm.
303Discussion. This transverse section of Meerbüsch quarry without carinae is similar to those of Aristophyllum luetti, but it belongs to a larger corallum with much more septa. It resembles the lectotype of A. ceratites (Goldfuss, 1826) from the Givetian of the Eifel Hills figured among others by Birenheide (1969, pl. 3, fig. 9) and Lütte (1987, pl. 4, fig. 17). The latter author has sampled a good collection of this species in the Lower Givetian Kerpen Formation of the same area with usually rather slender septa. Nose et al. (2017, pl. 2, fig. 1) have illustrated one specimen of Meerbüsch quarry coming from the top of the Binz Member and assigned to Glossophyllum sp. aff. ceratites which is separated from my transverse section collected in the Galgenberg Member by septa more systematically dilated in the dissepimentarium, longer major septa and sometimes shorter minor septa.
304Distribution. The corallum of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation.
305Genus Spinophyllum Wedekind, 1922
306Type species. By monotypy, Campophyllum spongiosum Schlüter, 1889.
307Diagnosis. See Coen-Aubert (2019).
308Spinophyllum sp.
309(Plate 6K–L)
310Material. 1 specimen with 2 thin sections: EIF432.
311Description. The material is restricted to a fragmentary ceratoid corallum with a height of 5.5 cm. The outer wall is relatively well preserved and encrusted by a laminar stromatoporoid including a thin platy alveolitid.
312The septa are slender at the periphery and sometimes separated from the outer wall by a few rows of dissepiments. They are slightly dilated in it the inner dissepimentarium and become thin beyond their entry into the tabularium. A few yardarm and spinose carinae are present in the dissepimentarium.
313The major septa leave a small open space in the centre of the tabularium where their axial ends are united laterally and form pseudofossulae. The minor septa traverse the entire dissepimentarium or they are hardly projecting into the tabularium where they may be contratingent.
314The dissepimentarium consists of 7 to 14 rows of globose dissepiments arranged in horizontal layers in its outer part and inclined in its inner part. A few spots of coarse trabeculae which are isolated or contiguous, occur in the dissepimentarium. The tabulae are vesicular at the periphery of the tabularium; in its narrow inner part, they are horizontal or incomplete and bordered by more or less strongly convex tabellae.
315There are 44 septa for a diameter of 17 mm to 18 mm whereas the width of the tabularium measures 7 mm.
316Discussion. The corallum of Meerbüsch is related to Spinophyllum spongiosum (Schlüter, 1889), type species of the genus, well known in the Givetian of Germany and Belgium and investigated among others by Birenheide & Lütte (1990) and Coen-Aubert (2002, 2017). The specimen described herein differs from this taxon by much less septa for a relatively small diameter, by a particular tabularium and by septa thin and locally discontinuous at the periphery.
317Distribution. The corallum of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation.
318Genus Argutastrea Crickmay, 1960
319Type species. By original designation, Argutastrea arguta Crickmay, 1960.
320Diagnosis. See Coen-Aubert (2017).
321Argutastrea briceae (Rohart, 1988)
322(Plate 7A–C)
323* 1988 Cyathophyllum briceae sp. nov.; Rohart, p. 282, pl. 31, fig. 15, pl. 36, figs 1–2.
3242011 Cyathophyllum briceae Rohart, 1988; Mistiaen et al., p. 94, fig. 78.
325v 2017 Argutastrea briceae (Rohart, 1988); Coen-Aubert, p. 168, pl. 5A–C.
326Holotype. See Coen-Aubert (2017).
327Material. 5 colonies with 13 thin sections collected by the author and W. Wenning: Meerbüsch EIF341, EIF343, EIF344, EIF429 and EIF430.
328Diagnosis. See Coen-Aubert (2017).
329Description. The material consists of big massive colonies which are occasionally tabular. Their height varies between 2 cm and 18 cm whereas the largest fragment reaches an area of 15 cm x 25 cm. The polygonal corallites are large and show sometimes an excavated calice bordered by a more or less narrow flat peripheral platform. They are separated by a straight wall with frequently a dark median line. A few pericalicinal offsets have been observed.
330The septa are normally non-carinate with locally a few small irregular carinae which are spinose and rarely knobbly. They are usually slightly dilated in the dissepimentarium and they become thin in the tabularium; in a few corallites, they are sometimes less thick in part of the latter. Occasionally, the septa are slender at the periphery of the corallites or even throughout their length.
331The major septa reach more or less the axis of the corallites or leave in some cases a small open space in the centre of the tabularium where their axial ends may be rhopaloid or united laterally to form pseudofossulae or a plane of bilateral symmetry. The minor septa traverse the entire and wide dissepimentarium or even enter into the tabularium where they are sometimes contratingent.
332The dissepimentarium consists of 9 or only 6 to 18 rows of small globose dissepiments arranged in horizontal layers at the periphery and inclined in its inner part. Occasionally, the dissepiments are more and more inclined towards the axis of the corallites. The tabulae are incomplete and intersecting laterally with sometimes narrow and more or less flat-topped axial parts.
333There are 42 to 56 or even 62 septa per corallite. The diameter of the corallites ranges from 13 mm to 25.5 mm. The width of the tabularium varies commonly between 5 mm and 7.4 mm and more generally between 4.5 mm and 8.9 mm.
334Discussion. The material is similar to that investigated by Rohart (1988) and Coen-Aubert (2017). As discussed by the latter author, Argutastrea briceae occupies a marginal position in the genus Argutastrea. It is closely related to A. tenuiseptata Coen-Aubert & Lütte, 1990 which is known in the Rodert and Kerpen Formations from the Eifel Hills and mainly in the lower part of the Givetian Mont d’Haurs Formation from southern Belgium. A. tenuiseptata is separated from A. briceae by slightly smaller septal number and diameters of the corallites and by mostly inclined dissepiments. This species also occurs in the Givetian Blacourt Formation from the Boulonnais in France, but it has been identified by Rohart (1988) as A. periclada (Krämer, 1982) as mentioned by Coen-Aubert & Lütte (1990, pp. 22–23).
335Distribution. The material of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation
336Outside Germany, Argutastrea briceae is known in the Lower Givetian from the Boulonnais in France and in the lower part of the Givetian from the Zemmour in Mauritania.
337Family Phillipsastreidae Roemer, 1883
338Genus Macgeea Webster, 1889
339Type species. By subsequent designation of Fenton & Fenton (1924, p. 54), Pachyphyllum solitarium Hall & Whitfield, 1873.
340Diagnosis. See Coen-Aubert (2017).
341Macgeea mistiaeni n. sp.
342(Plate 6I–J; Plate 7F–I)
343Derivation of name. The species is dedicated to Bruno Mistiaen (1946–2019), a distinguished French specialist of Devonian stromatoporoids and tabulate corals mainly from France and Afghanistan.
344Holotype. IRScNB a 13563 (= Plate 6I–J). Specimen EIF410 collected by M. Coen-Aubert in 2000, in the Lower Givetian of the Eifel Hills, Germany.
345Type locality and horizon. Meerbüsch quarry in the Hillesheim Syncline, Eifel Hills in Germany. Base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation.
346Material. 12 specimens with 22 thin sections: Meerbüsch EIF373, EIF374, EIF375, EIF377, EIF378, EIF379, EIF380, EIF410, EIF419, EIF420, EIF421 and EIF422.
347Diagnosis. A species of Macgeea with 50 to 60 septa at a diameter of 8 mm to 13 mm. Septa slightly dilated in the dissepimentarium and leaving an open space in the centre of the tabularium. Several rows of inclined dissepiments on the inner side of the horseshoe dissepiments.
348Description. The material consists of often fragmentary coralla which are usually cylindrical, but may also be conical, trochoid or ceratoid; their length varies between 1.5 cm and 3 cm. Some of them have longitudinal ribs. Two specimens show lateral offsets whereas in one of them, there are four axial offsets at its top. The holotype shows laterally some small spines. The outer wall is thin and not very well preserved; it is rarely encrusted by a thin laminar stromatoporoid or by a favositid.
349The septa are commonly non-carinate though a few spinose and knobbly carinae occur in some coralla. The septa are weakly dilated in the dissepimentarium and become thin in the tabularium or slightly beyond their entry into it. A dark median line may affect the septa in the dissepimentarium. In rare specimens, the septa are locally slender in the outer zone of flat dissepiments or they are thickened throughout their length. In transverse section, the pipe of horseshoe dissepiments is often coated with stereome on one or both sides and it is bordered by a few inner dissepiments.
350The major septa leave a more or less wide open space in the centre of the tabularium; their axial ends are occasionally discontinuous, bifurcated, forked or forming pseudofossulae. The minor septa are restricted to the dissepimentarium or they are hardly projecting into the tabularium where they may be contratingent.
351The dissepimentarium consists of:
352- one peripheral row of flat dissepiments, sometimes concave or inclined;
353- one row of occasionally irregular horseshoe dissepiments with locally narrow symmetrical fans of rhipidacanths centred over them;
354- 1 to 3 or even 0 to 4 rows of inner inclined dissepiments.
355The tabulae are incomplete and intersecting laterally with frequently a flat-topped axial part; they are occasionally horizontal.
356There are 48 to 62 septa per corallite. The diameter of the corallites ranges from 7.2 mm to 13.5 mm. The width of the tabularium varies commonly between 5.3 mm and 7.9 mm and more generally between 4.5 mm and 8.5 mm.
357Discussion. The new species is a relatively small Macgeea with not so many septa. However, it is characterized by several rows of inner dissepiments as it is the case for this genus. By the weak dilation of the septa, M. mistiaeni can be compared to M. josephi Rozkowska, 1956 from the Upper Eifelian to the Lower Givetian of the Holy Cross Mountains in Poland. But this taxon has slightly more septa and larger coralla. Macgeea mistiaeni also resembles M. regularis Rozkowska, 1956 from the same age and area in Poland, which has thicker and typically carinate septa in the dissepimentarium. Additionally, there are some affinities between the new species and some corallites of Thamnophyllum germanicum Scrutton, 1968 whose lectotype revised by Coen-Aubert (1998) probably comes from the Givetian of the Eifel Hills; conspecific specimens from the Upper Eifelian Jemelle Formation in southern Belgium have also been investigated by this author. Thamnophyllum germanicum is distinguished from Macgeea mistiaeni by slightly smaller septal number and diameter of the corallites and by the weaker development of inner dissepiments.
358Distribution. The new species is only known in Meerbüsch quarry where it has been collected at the base of the Lower Givetian Galgenberg Member characterizing the upper part of the Dreimühlen Formation.
359Family Mucophyllidae Hill, 1940
360Genus Pselophyllum Pocta, 1902
361Type species. By subsequent designation of Hill (1940, p. 158), Pselophyllum bohemicum Pocta, 1902.
362Diagnosis. Solitary rugose corals. Septa of two orders short and slightly unequal, strongly dilated and contiguous in a peripheral stereozone. Axial ends of major septa sometimes a little thinner. Septa composed of coarse rhabdacanthine trabeculae. No dissepiments. Tabulae complete to incomplete, forming flat to concave surfaces.
363Discussion. Pselophyllum Pocta, 1902 is well defined as its type species P. bohemicum Pocta, 1902 from the Pragian of Bohemia in Czech Republic has been redescribed and reillustrated by Oliver & Galle (1971, p. 63) who have also selected a lectotype. The genus is considered as synonym of Pseudamplexus Weissermel, 1897 (Hill, 1981, p. F178) whose type species is P. ligeriense (Barrois, 1889) from the Emsian of the Southern Armorican Massif in France. According to Hill (1981), the type material of this taxon cannot be traced. It consists of two coralla figured by Barrois (1889) with their two longitudinal sections and only a small fragment of one transverse section. With no complete transverse section, Pseudamplexus is without doubt badly known. Another difficult genus to be used is Tryplasma Lonsdale, 1845 whose type species is T. aequabile Lonsdale, 1845 from the Lower Devonian of the Urals in Russia. According to Weyer (2008, p. 130), the designation of a neotype for this taxon made by Ivanovski & Shurigina (1975, p. 15) is not valid and not well documented. Indeed, the topotype of T. aequabile figured by Lonsdale (1845, pl. A, fig. 7) is characterized by a thin outer wall and not by a peripheral stereozone. Therefore, the neotype of Ivanovski & Shurigina (1975) does not belong to the same species.
364Pselophyllum sp.
365(Plate 7D–E)
366Material. 1 specimen with 3 thin sections: Meerbüsch EIF413.
367Description. The material is restricted to a fragmentary ceratoid corallum with a height of 6 cm. The outer wall is completely preserved, first encrusted by a laminar stromatoporoid and then by an alveolitid.
368The septa of both orders are short, thick and contiguous in a relatively narrow peripheral stereozone. The major septa are slightly projecting beyond this stereozone. In transverse section, some small trabeculae and circular traces of tabulae with small septal spines occur in the centre of the corallum.
369There are no dissepiments. The tabulae are widely spaced, complete or incomplete with a concave pattern. In longitudinal section, coarse rhabdacanth trabeculae are observed in the peripheral stereozone; they are straight and at a low angle to the horizontal.
370There are 86 septa at a diameter of 26.5 mm to 33 mm.
371Discussion. Pselophyllum sp. differs from P. bohemicum by a smaller diameter, a narrower stereozone and concave tabulae. The material collected in the Upper Eifelian Ahrdorf Formation from the Eifel Hills and identified by Glinski (2001) as Tryplasma rohrense Glinski, 2001 is closer to Pselophyllum sp., but it is characterized by smaller coralla. On the contrary, the specimens assigned to the same species by Jamart & Denayer (2020) and sampled close to the Eifelian–Givetian boundary in southern Belgium are different in having a thin outer wall and a very narrow peripheral stereozone. There are some affinities between Pselophyllum sp. and a fragmentary transverse section referred by Galle (1994) to cf. Pseudamplexus? obesus (Pocta, 1902). This specimen comes from the Lower Givetian part of the Acanthopyge Limestone in Bohemia (Czech Republic) as explained by Galle (1994, pp. 41–42). However, the holotype of Pseudamplexus? obesus from the Pragian of Bohemia, which has been revised by Oliver & Galle (1971, p. 66), is separated by a very thin outer wall without any stereozone.
372Distribution. The corallum of Meerbüsch quarry comes from the base of the Lower Givetian Galgenberg Member, in the upper part of the Dreimühlen Formation.
373Remark about the ptenophyllid genus Dohmophyllum Wedekind, 1923
374The collection of rugose corals gathered by Nose et al. (2017) in Meerbüsch quarry is certainly greater than mine, but it is restricted to the top of the Binz Member in the Dreimühlen Formation. Among their material, they figured two coralla of Dohmophyllum Wedekind, 1923: the first one (their pl. 2, fig. 2) assigned to D. helianthoides (Goldfuss, 1826) and the second one (their pl. 3, fig. 4) assigned to D. difficile (Wedekind, 1925). It is clear that these two specimens are very similar and that they probably belong to the same species, namely D. difficile. This taxon has been revised by Birenheide (1963, p. 418) whereas its lectotype from the Lower Givetian of the Sauerland in Germany has been chosen by Birenheide (1962). Dohmophyllum helianthoides has also been investigated by Birenheide (1963, p. 414) who selected its lectotype coming from the Upper Eifelian of the Eifel Hills. D. helianthoides is very different from D. difficile. Indeed, it is characterized by a much greater diameter of the corallum, by septa thin in the outer dissepimentarium and only dilated in its inner part and by numerous rows of dissepiments arranged in horizontal layers.
4. Stratigraphic and palaeobiogeographic conclusions
375My Lower Givetian collection of rugose corals from Meerbüsch quarry is highly diversified and comes mostly from two argillaceous and coralliferous levels of the Dreimühlen Formation: the lower one at the top of the Binz Member and the upper one at the base of the Galgenberg Member. Only Grypophyllum schroederi has been sampled close to the base of the Ley Member in a pure limestone with massive stromatoporoids. Additionally, my fauna consists mainly of solitary rugose corals associated with some fasciculate specimens of Disphyllum caespitosum at the top of the Binz Member whereas numerous and large massive colonies of Argutastrea briceae are present at the base of the Galgenberg Member. The latter occurrence has never been mentioned in the Givetian of the Eifel Hills. Argutastrea briceae is abundant close to the base of the Griset Member of the Blacourt Formation in the Boulonnais, France. These layers are still Lower Givetian in age, but younger than the Dreimühlen Formation (Mistiaen et al., 2011, p. 57).
376In Meerbüsch quarry, there are no massive rugose corals at the base of the Rodert Formation whereas many colonies of A. quadrigemina (Goldfuss, 1826) have been observed in the lower part of this lithostratigraphic unit, in the Sötenich Syncline to the north of the Eifel Hills (Fig. 1). This species has been investigated in the two neighbouring quarries of Sötenich and Keldenich, by Coen-Aubert & Lütte (1990) who have also mentioned a few specimens in the Cürten Formation. Argutastrea quadrigemina is very useful for the correlations between the Rodert Formation in the Eifel Hills and the Terres d’Haurs Formation on the south side of the Dinant Synclinorium in Belgium (Fig. 4). Indeed, A. quadrigemina is frequent in the lower part of the Terres d’Haurs Formation, but it is locally present close to the top of the underlying Trois-Fontaines Formation (Coen-Aubert, 2003, 2008).
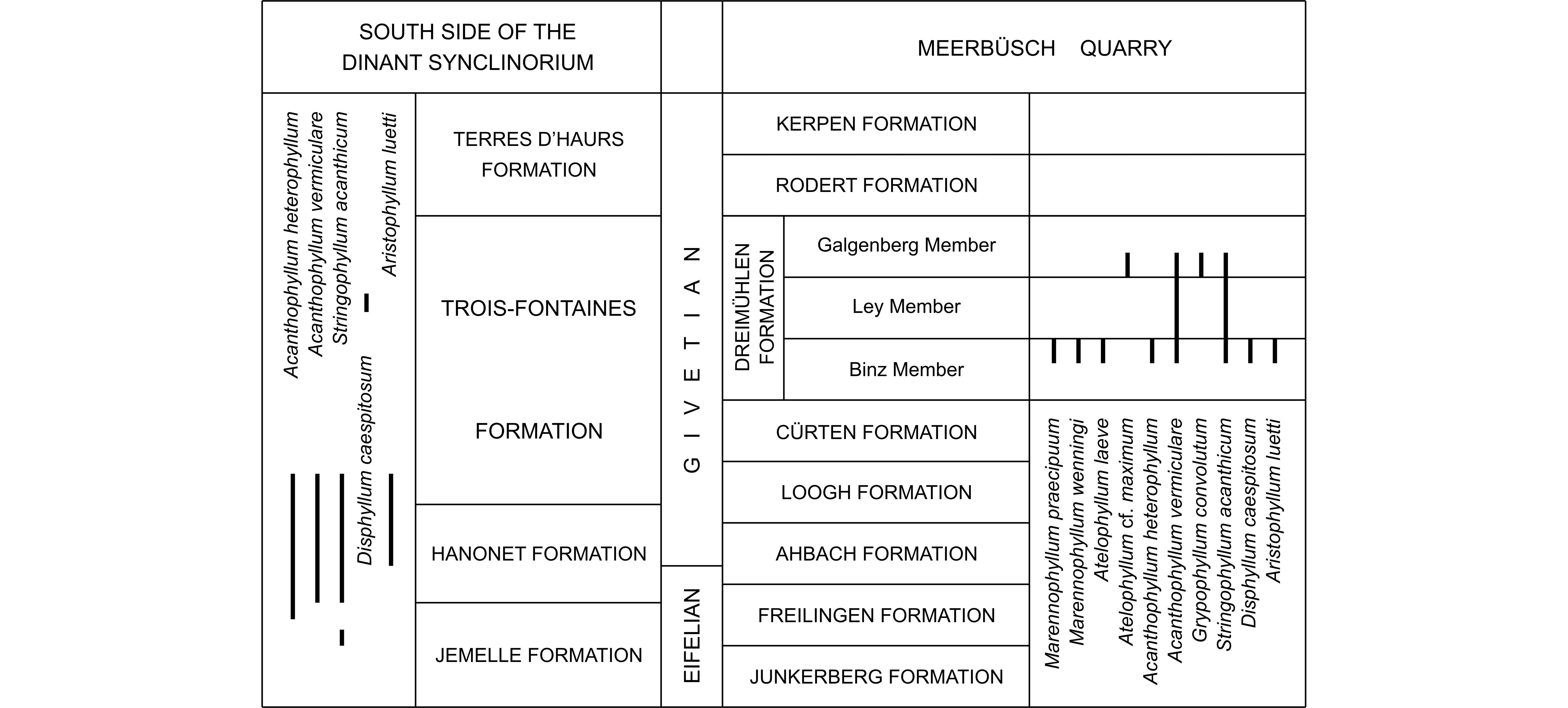
Figure 4. Correlations between the Lower Givetian lithostratigraphic units from the Meerbüsch quarry and the south side of the Dinant Synclinorium in Belgium with the distribution of the rugose coral species common to both areas and also known elsewhere in the Eifel Hills.
377For the rest, the latter lithostratigraphic unit starts with coarsely crinoidal and reefal limestones, very rich in diverse rugose corals, which are usually followed by a thick sequence of lagoonal limestones poor in fossils. However and more particularly in the area of Beauraing and Pondrôme (Coen-Aubert, 2003), the upper part of the Trois-Fontaines Formation is invaded by some layers of more or less crinoidal limestones with sometimes colonial rugose corals. Such a level with Disphyllum caespitosum has been described by Coen-Aubert (2008) at Pondrôme, in the Bois le Ban quarry, less than 25 m below the top of the lithostratigraphic unit. This occurrence of D. caespitosum can be compared with those of the Eifel Hills (Figs 4, 5).
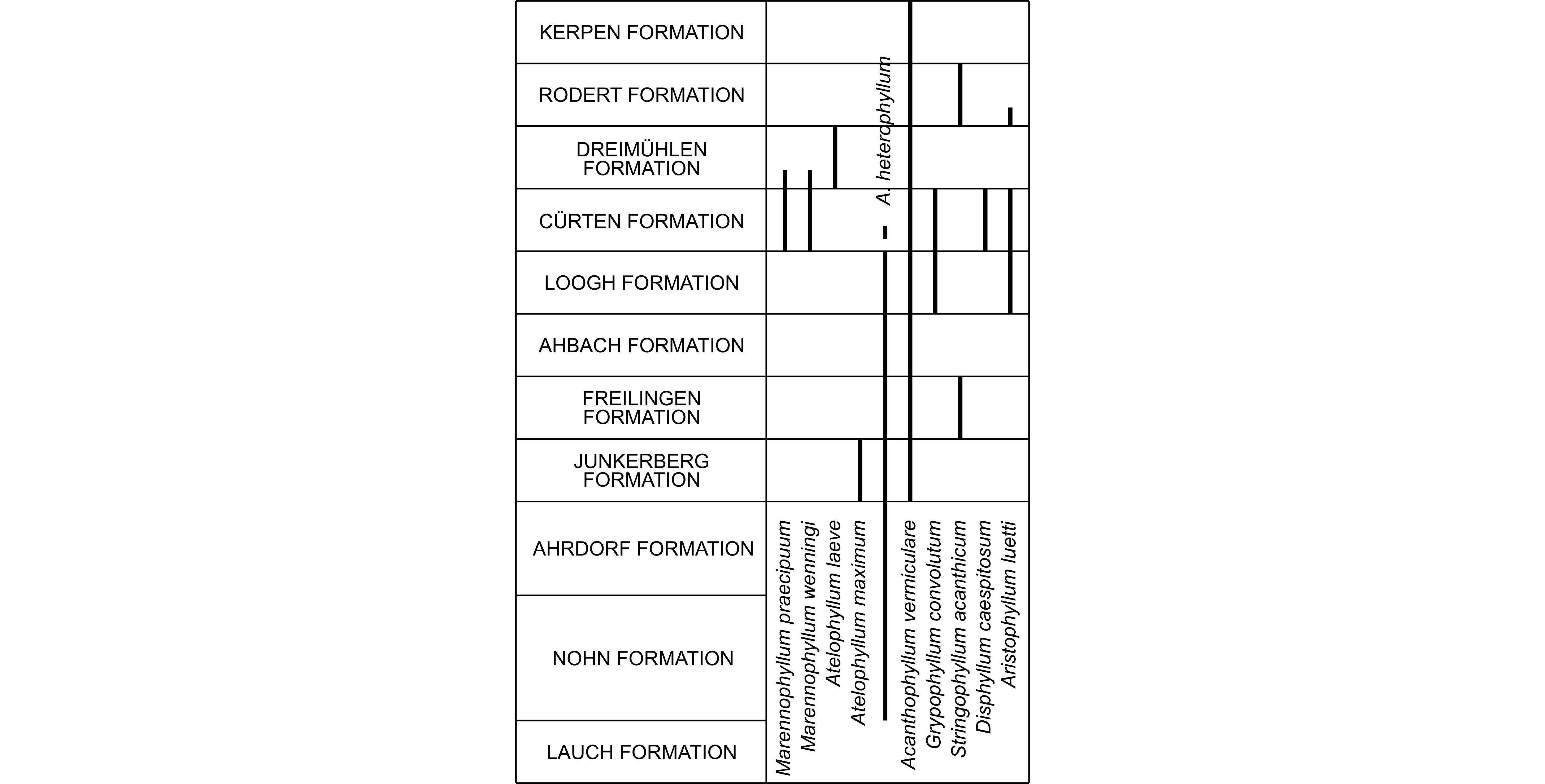
Figure 5. Stratigraphic distribution in the Middle Devonian of the Eifel Hills, for the species of rugose corals identified in the Meerbüsch quarry. For the Eifelian–Givetian boundary, see Fig. 4.
378On the contrary, species such as Acanthophyllum heterophyllum, A. vermiculare, Stringophyllum acanthicum and Aristophyllum luetti are restricted in southern Belgium, to the top of the Eifelian and to the base of the Givetian whereas they have a much greater range in the Eifel Hills (Figs 4, 5). This is probably due to the particular facies characterizing the main part of the Trois-Fontaines Formation. For the Meerbüsch quarry, these four species are present at the top of the Binz Member, but two of them, namely Stringophyllum acanthicum and Acanthophyllum vermiculare, occur also at the base of the Galgenberg Member. Additionally, A. vermiculare and A. luetti represented by several specimens in Meerbüsch quarry show the same variability, which affects mainly the dilation of the septa in the dissepimentarium, as in the Belgian material investigated by Coen-Aubert (1997).
379What about the other solitary rugose corals identified in Meerbüsch quarry? As mentioned in the systematic part, Marennophyllum praecipuum and M. wenningi are only known in the Hillesheim Syncline, from the Cürten Formation to the Binz Member of the Dreimühlen Formation. According to Birenheide (1978), Atelophyllum laeve is restricted to the Dreimühlen Formation in the Eifel Hills whereas Grypophyllum convolutum has also been found in the Loogh and Cürten Formations. Some specimens from the Cürten to the Dreimühlen Formations illustrated by Schröder & Salerno (2001, pl. 3, figs 23, 25–26) are related to the new species G. schroederi. On the contrary, Macgeea mistiaeni can only be compared with some species from the Middle Devonian of the Holy Cross Mountains in Poland. One specimen of Pselophyllum sp. has been collected in Meerbüsch quarry. With the exception of Tryplasma rohrense investigated by Glinski (2001), such coralla with a typical peripheral stereozone and no dissepiments seem to be rare in the Middle Devonian of the Eifel Hills.
380Besides Germany and Belgium, some species collected in Meerbüsch quarry are also present in other countries of Western Europe. However, Acanthophyllum heterophyllum, A. vermiculare, Grypophyllum convolutum, Stringophyllum acanthicum and Aristophyllum luetti have been additionally recognized in the Upper Eifelian and/or the Lower Givetian from the Ma’der in Morocco, in the lower part of the Givetian from the Zemmour in Mauritania and in the Givetian of the Alborz Mountains in Iran; these areas belong to the northern margin of Gondwana. Finally, two specimens of Heliophyllum cf. cribellum coming from the base of the Galgenberg Member suggest an eastern North American influence. The genus Heliophyllum Hall in Dana, 1846 was already known in the Eifel Hills and in Southern Belgium with the species H. halleri from the Upper Eifelian Freilingen Formation of the Dollendorf Syncline (Fig. 1) whereas Coen-Aubert (1996) has assigned conspecific specimens from the Upper Eifelian Hanonet Formation of Resteigne to H. dahlemense. For her identification, Coen-Aubert (1996) has illustrated and compared the type specimens of H. dahlemense with her material. However, Jamart & Denayer (2020, p. 288, figs 6A–B) have referred one corallum from Resteigne to H. halleri and to H. dahlemense sensu Birenheide (1963, pl. 62, figs 63–65) who has precisely figured the type specimens of the latter species. There is a problem with the type material of Haller (1936) which has been collected in fields from the Blankenheim Syncline (Fig. 1) so that its age and the collecting place are not sure.
Acknowledgements
381Walter Wenning introduced me to Meerbüsch quarry. Michel Coen made the geological survey of this outcrop. Stefan Schröder provided me stratigraphic information about Meerbüsch quarry. The thin sections have been made by René Cremers and by Joël Laval at the Laboratory of successively Eddy Poty and Valentin Fischer (Liège). Wilfried Miseur and Thierry Hubin helped me with photography of the corals. Ross McLean and Julien Denayer reviewed very carefully the manuscript. I am most grateful to all these people.
Author contribution
382Collecting of the rugose corals, detailed palaeontological identifications of the genera and species, writing of the article.
Data availability
383All the specimens and thin sections of the rugose corals are stored in the Collection of Palaeontology of the Institut royal des Sciences naturelles de Belgique at Brussels.
References
384Abbasi, M.A., Khaksar, K. & Ashouri, A., 2014a. Some rugose corals from the Devonian (Givetian and Frasnian) of Northeastern Iran. Neues Jahrbuch für Geologie and Paläontologie, 271, 123–139. https://doi.org/10.1127/0077-7749/2014/0379
385Abbasi, M.A., Ashouri, A. & Khaksar, K., 2014b. Solitary rugose corals from the Givetian of the Khoshyeilagh Formation (Eastern Alborz Mountains, NE Iran). Palaeodiversity, 7, 1–21.
386Abbasi, M.A. Khaksar, K & Ashouri, A., 2014c. Description of some rugose corals from the Givetian and Lower Frasnian of the Eastern Alborz Mountains, NE Iran. Geopersia, 4, 237–257.
387Barrois, C., 1889. Faune du Calcaire d'Erbray. Mémoires de la Société Géologique du Nord, 3/1, 1–348.
388Billings, E., 1859. On the fossil corals of the Devonian rocks of Canada West. The Canadian Journal of Industry, Science and Art, new series, 4, 97–140.
389Besprozvannykh, N.I., Dubatolov, V.N., Kravtsov, A.G., Latypov, Yu. Ya. & Spassky, N. Ya., 1975. Devonskie rugozy Taymyro-Kolymskoy provintsii. Trudy Instituta Geologii i Geofiziki, Akademiya Nauk SSSR, Sibirskoe Otdelenie, 228, 1–172. [In Russian].
390Birenheide, R., 1961. Die Acanthophyllum-Arten (Rugosa) aus dem Richtschnitt Schönecken-Dingdorf und aus anderen Vorkommen in der Eifel. Senckenbergiana lethaea, 42, 77–146.
391Birenheide, R., 1962. Die Typen der Sammlung Wedekind aus den Familien Cyathophyllidae und Stringophyllidae (Rugosa). Senckenbergiana lethaea, 43, 101–123.
392Birenheide, R., 1963. Cyathophyllum- und Dohmophyllum-Arten (Rugosa) aus dem Mitteldevon der Eifel. Senckenbergiana lethaea, 44, 363–458.
393Birenheide, R., 1964. Die « Cystimorpha » (Rugosa) aus dem Eifeler Devon. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 507, 1–120.
394Birenheide, R., 1968. Die Typen der Sammlung Wedekind aus der Gattung Plasmophyllum (Rugosa; Mitteldevon). Senckenbergiana lethaea, 49, 1–37.
395Birenheide, R., 1969. Typen mittel-und oberdevonischer Rugosa aus der Sammlung Goldfuss. Senckenbergiana lethaea, 50, 37–55.
396Birenheide, R., 1972. Ptenophyllidae (Rugosa) aus dem W-deutschen Mitteldevon. Senckenbergiana lethaea, 53, 405–437.
397Birenheide, R., 1978. Rugose Korallen des Devon. In Krömmelbein, K. (ed.), Leitfossilien begründet von G. Gürich. 2., völlig neu bearbeitete Auflage, n 2. Gebrüder Borntraeger, Berlin-Stuttgart, 265 p.
398Birenheide, R. & Lütte, B.P., 1990. Rugose Korallen aus dem Mittel-Givetium (Mittel-Devon) des Rheinischen Schiefergebirges. Senckenbergiana lethaea, 70, 1–28.
399Birenheide, R. & Soto, F.M., 1981. “Cystimorphe” rugose Korallen aus dem Devon des Kantabrischen Gebirges, N-Spanien. Senckenbergiana lethaea, 62, 251–275.
400Blake, P.R., 2010. Devonian Corals of the Yarrol Province, eastern-central Queensland. Memoir of the Association of Australasian Palaeontologists, 38, 1–191.
401Brice, D. (ed.), 2016. Stratotype Givétien. Muséum national d’Histoire naturelle. Paris; Biotope, Mèze, 272 p.
402Brownlaw, R.L.S. & Jell, J.S., 2008. Middle and Upper Devonian rugose corals from the Canning Basin, Western Australia. Memoir of the Association of Australasian Palaeontologists, 35, 1–126.
403Bulvanker, E.Z., 1958. Devonskie chetyrekhluchevye korally okrain Kuznetskogo basseyna. Vsesoyuznyi nauchno-issledovatelskiy Geologicheskiy Institut (VSEGEI), Leningrad, 212 p. [In Russian].
404Bulvanker, E.Z., Gorianov, V.B., Ivanovski, A.B., Spassky, N.Ya. & Shchukina, V.Ya., 1968. Novye predstaviteli chetyrekhluchevykh korallovykh polipov SSSR. In Markovskiy, B.P. (ed.), Novye vidy drevnikh rasteniy i bespozvonochnykh SSSR, 2, 2. Nedra, Moskva, 14–45. [In Russian].
405Cao, X.D., Ouyang, X. & Jin, T.A., 1983. Rugosa. In Xi'an Institute of Geology and Mineral Resources (ed.), Paleontological Atlas of Northwest China, Shaanxi, Gansu and Ningxia Volume. Part II Upper Palaeozoic. Geological Publishing House, Peking, 46–179.
406Chapman, E.J., 1893. On the corals and coralliform types of Palaeozoic strata. Proceedings and Transactions of the Royal Society of Canada, 10, section 4, 39–48. https://doi.org/10.5962/bhl.title.38717
407Crickmay, C.H., 1960. The older Devonian faunas of the Northwest Territories. Published by the author, Evelyn de Mille Books, Calgary, 21 p.
408Coen‑Aubert, M., 1990. Description de quelques Rugueux coloniaux du Couvinien supérieur de Wellin (bord sud du Bassin de Dinant, Belgique). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 59, 15–35.
409Coen-Aubert, M., 1996. Siphonophrentides et Cyathophyllides près de la limite Eifelien-Givetien à Resteigne (Ardenne, Belgique). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 66, 19–36.
410Coen-Aubert, M., 1997. Rugueux solitaires près de la limite Eifelien-Givetien à Pondrôme (Belgique). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 67, 5–24.
411Coen-Aubert, M., 1998. Thamnophyllides et Acanthophyllides près de la limite Eifelien-Givetien à Wellin et Pondrôme (Belgique). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 68, 5–24.
412Coen-Aubert, M., 2000. Stratigraphy and additional rugose corals from the Givetian Mont d’Haurs Formation in the Ardennes. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 70, 5–23.
413Coen-Aubert, M., 2002. Temnophyllids and Spinophyllids (Rugosa) from the Givetian Mont d’Haurs Formation in Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 72, 5–24.
414Coen-Aubert, M., 2003. Description of a few rugose corals from the Givetian Terres d’Haurs Formation in Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 73, 11–27.
415Coen-Aubert, M., 2008. Fasciculate Disphyllids (Rugosa) from the Early Givetian Trois-Fontaines Formation in Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 78, 31–50.
416Coen-Aubert, M., 2011. Reassignment to the Middle Devonian of some rugose corals investigated by Le Maître (1934) in the Chalonnes Formation from the Southeastern Armorican Massif (France). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 81, 27–53.
417Coen-Aubert, M., 2017. Givetian rugose corals from the Zemmour in Mauritania. Geologica Belgica, 20, 161–180. http://dx.doi.org/10.20341/gb.2017.009
418Coen-Aubert, M., 2019. Investigation of some Givetian rugose corals from the Mont d’Haurs Formation in southern Belgium. Geologica Belgica, 22, 121–138. https://doi.org/10.20341/gb.2019.008
419Coen-Aubert, M. & Lütte, B.P., 1990. Massive rugose corals from the Middle Devonian of the North Eifel Hills (Rheinisches Schiefergebirge, West Germany). Geologica et Palaeontologica, 24, 17–39.
420Dana, J.D., 1846. Genera of fossil corals of the family Cyathophyllidae. The American Journal of Science and Arts, series 2, 1, 178–189.
421De Fromentel, E., 1861. Introduction à l'étude des polypiers fossiles. Savy, Paris, 357 p.
422Dybowski, W.N., 1873. Monographie der Zoantharia Sclerodermata Rugosa aus der Silurformation Estlands, Nord-Livlands und der Insel Gothland. Archiv für die Naturkunde Liv-, Est- und Kurlands, series 1, 5, 257–414.
423Engel, G. & Von Schouppé, A., 1958. Morphogenetisch-taxionomische Studie zu der devonischen Korallengruppe Stringophyllum, Neospongophyllum und Grypophyllum. Paläontologische Zeitschrift, 32, 67–114. https://doi.org/10.1007/BF02987036
424Fenton, C.L. & Fenton, M.A., 1924. The stratigraphy and fauna of the Hackberry Stage of the Upper Devonian. Contributions from the Museum of Geology, University of Michigan, 1, 1–260. https://doi.org/10.3998/mpub.9690830
425Frech, F., 1885. Ueber das Kalkgerüst der Tetrakorallen. Zeitschrift der Deutschen Geologischen Gesellschaft, 37, 928–945.
426Frech, F., 1886. Die Cyathophylliden und Zaphrentiden des deutschen Mitteldevon. Palaeontologische Abhandlungen, 3/3, 1–120.
427Galle, A., 1994. Rugose corals of the Acanthopyge Limestone of Koneprusy (Middle Devonian, Barrandian, Czech Republic). Vestnik Ceskeho geologickeho ustavu, 69, 41–58.
428Glinski, A., 2001. Tryplasma (Anthozoa, Rugosa) im Mittel-Devon der Eifel (Rheinisches Schiefergebirge, Deutschland). Senckenbergiana lethaea, 81, 71–89. https://doi.org/10.1007/BF03043295
429Goldfuss, A., 1826. Petrefacta Germaniae 1. Arnz & Comp., Düsseldorf, 1–76.
430Gräf, W., 1958. Kritische Bemerkungen zu den als "Cyathophyllum" vermiculare aus dem Grazer Mitteldevon beschriebenen Korallen. Mitteilungen des Naturwissenschaftlichen Vereines für Steiermark, 88, 79–103.
431Hall, J., 1843. Geology of New York. Part IV comprising the survey of the fourth geological district. Albany, 683 p.
432Hall, J. & Whitfield, R.P., 1873. Description of new species of fossils from the Devonian rocks of Iowa. Annual Report of the New York State Cabinet of Natural History, 23, 223–239.
433Haller, W, 1936. Einige biostratigraphische Untersuchungen in der Rohrer Mulde unter besonderer Berücksichtigung der Keriophyllen. Jahrbuch der Preussischen Geologischen Landesanstalt zu Berlin, 56, 590–632.
434Hill, D., 1939. The Devonian rugose corals of Lilydale and Loyola, Victoria. Proceedings of the Royal Society of Victoria, new series, 51, 219–256.
435Hill, D., 1940. The Middle Devonian rugose corals of Queensland, II: The Silverwood-Lucky Valley area. Proceedings of the Royal Society of Queensland, 51, 150–168.
436Hill, D., 1981. Part F, Coelenterata, Supplement 1, Rugosa and Tabulata, 2 vols. In Teichert, C. (ed.), Treatise on Invertebrate Paleontology. The Geological Society of America and The University of Kansas, Boulder (CO) and Lawrence (KS), 762 p.
437Hill, D. & Jell, J.S., 1970. Devonian corals from the Canning Basin Western Australia. Geological Survey of Western Australia, Bulletin, 121, 1–158.
438Ivanovski, A.B. & Shurigina, M.V., 1975. Reviziya rugoz Urala. Trudy Instituta Geologii i Geofiziki Sibirskoe Otdelenie Akademiya Nauk SSSR, 218, 1–66. [In Russian].
439Jamart, V. & Denayer, J., 2020. The Kacak event (late Eifelian, Middle Devonian) on the Belgian shelf and its effects on rugose coral palaeobiodiversity. Bulletin of Geosciences, 95, 279–311. https://doi.org.10.3140/bull.geosci.1788
440Joseph, J. & Tsien, H.H., 1975. Calcaires mésodévoniens et leurs faunes de Tétracoralliaires en Haute Vallée d’Ossau (Pyrénées-Atlantiques). Bulletin de la Société d’Histoire naturelle de Toulouse, 111, 179–203.
441Königshof, P., Helling, S., Hartenfels, S., De Vleeschouwer, D., Schreiber, G., Becker, R.T. & Brett, C.E., 2016. Eifel Synclines – an overview and Emsian to Frasnian section descriptions. Münstersche Forschungen zur Geologie und Paläontologie, 108, 36–45.
442Krämer, T.J., 1982. Cerioide Rugosa aus dem Devon der Bergisch Gladbach-Paffrather Mulde (Rheinisches Schiefergebirge). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 1982/11, 648–666. https://doi.org/10.1127/njgpm/1982/1982/648
443Lang, W.D. & Smith, S., 1934. Ludwig’s “Corallen aus Paläolitischen Formationen” and the genotype of Disphyllum de Fromentel. The Annals and Magazine of Natural History, series 10, 13, 78–81. https://doi.org/10.1080/00222933408654793
444Lang, W.D. & Smith, S., 1935. Cyathophyllum caespitosum Goldfuss and other Devonian corals considered in a revision of that species. The Quarterly Journal of the Geological Society of London, 91, 538–589. https://doi.org.10.1144/GSL.JGS.1935.091.01-04.19
445Liao, W.H. & Birenheide, R., 1984. Rugose Korallen aus dem Givetium von Dushan, Provinz Guizhou, S-China. 1: “Cystimorpha”. Senckenbergiana lethaea, 65, 1–25.
446Lonsdale, W., 1845. Description of some characteristic Palaeozoic corals of Russia. In Murchison, R.I., De Verneuil, E. & Von Keyserling, A., The geology of Russia in Europe and the Ural Mountains, Volume 1 Geology, Appendix A. John Murray, London and P. Bertrand, Paris, 591–634.
447Lütte, B.P., 1984. Rugose Korallen aus dem Mitteldevon (Givetium) der Sötenicher Mulde (Rheinisches Schiefergebirge, Nord-Eifel). Münstersche Forschungen zur Geologie und Paläontologie, 61, 175–243.
448Lütte, B.P., 1987. Glossophyllum-Arten aus dem Mittel-Devon der Eifel (Rugosa; Rheinisches Schiefergebirge). Senckenbergiana lethaea, 67, 433–457.
449Lütte, B.P., 1990. Horn-und kegelförmige rugose Korallen aus dem Mittel-Devon der Eifel. Senckenbergiana lethaea, 70, 297–395.
450Ma, T.Y.H., 1956. A reinvestigation of climate and the relative positions of continents during the Devonian. Research on the Past Climate and Continental Drift, 9, 1–116.
451McLean, R.A., 1976. Middle Devonian cystiphyllid corals from the Hume Formation, Northwestern Canada. Bulletin Geological Survey of Canada, 274, 1–80. https://doi.org/10.4095/103067
452McLean, R.A., 2014. Solitary disphyllid corals from the Frasnian (Upper Devonian) of western Canada. Palaeontographica Canadiana, 34, 1–123.
453McLean, R.A., 2021. Devonian cystiphyllid rugose corals from western Canada and Eastern Australia. Palaeontographica Canadiana, 38, 1–159.
454Merriam, C.W., 1973. Middle Devonian rugose corals of the Central Great Basin. U.S. Geological Survey Professional Paper, 799, 1–53. https://doi.org/10.3133/pp799
455Middleton, G.V., 1959. Devonian tetracorals from South Devonshire, England. Journal of Paleontology, 33,138–160.
456Milne-Edwards, H. & Haime, J., 1850. A Monograph of the British Fossil Corals. Part 1, Introduction. The Palaeontographical Society, London, i–lxxxv. https://doi.org/10.5962/bhl.title.11691
457Milne-Edwards, H. & Haime, J., 1851. Monographie des Polypiers fossiles des terrains paléozoïques. Archives du Muséum d’Histoire Naturelle, 5, 1–502.
458Mistiaen, B., Brice, D., Hubert, B.L.M. & Loones, C., 2011. Field Trip 2: Classical Devonian and Carboniferous sites in the Ferques area, Boulonnais, Northern France. In Aretz, M. & Poty, E. (eds), Field Guides, 11th International Symposium on Fossil Cnidaria and Porifera, Liège, 2011. Kölner Forum für Geologie und Paläontologie, 20, 51–98.
459Nose, M., Schröder, S. & Fischer, U., 2017. Paläoökologie einer mergeligen Korallen-Assoziation aus dem Mitteldevon der Eifel (Hillesheimer Mulde), Rheinisches Schiefergebirge. Zitteliana, 89, 113–131.
460Oliver, W.A. & Galle, A., 1971. Rugose corals from the Upper Koneprusy Limestone (Lower Devonian) in Bohemia. Sbornik Geologickych Ved, Paleontologie, 14, 35–106.
461Oliver, W.A. & Sorauf, J.E., 2002. The genus Heliophyllum (Anthozoa, Rugosa) in the Upper Middle Devonian (Givetian) of New York. Bulletins of American Paleontology, 362, 1–72.
462Pedder, A.E.H. & McLean, R.A., 1982. Lower Devonian cystiphyllid corals from North America and eastern Australia with notes on the genus Utaratuia. Geologica et Palaeontologica, 16, 57–110.
463Pocta, P., 1902. Anthozoaires et Alcyonaires. In Barrande, J. (ed.), Système silurien du centre de la Bohême, 1ère Partie: Recherches paléontologiques, 8 (2). Musée Bohême, Prague, 347 p.
464Roemer, C.F., 1883. Lethaea geognostica. I. Theil, Lethaea palaeozoica. Zweite Lieferung. Stuttgart, 324–543 and Atlas.
465Rohart, J.C., 1988. Rugueux givetiens et frasniens de Ferques (Boulonnais-France). In Brice, D. (ed.), Le Dévonien de Ferques. Bas-Boulonnais (N. France). Biostratigraphie du Paléozoïque, 7, 231–297.
466Rozkowska, M., 1956. Pachyphyllinae from the Middle Devonian of the Holy Cross Mts. Acta Palaeontologica Polonica, 1, 271–322.
467Salerno, C., 2008. Stromatoporen-Fauna, Fazies und Paläoökologie von Plattformkarbonaten aus dem Unter-Givetium der Eifel (Devon, Rheinisches Schiefergebirge). Zitteliana, B27, 3–129.
468Schlüter, C., 1882. Neue Korallen des Mitteldevon der Eifel. Verhandlungen des naturhistorischen Vereines der preussischen Rheinlande und Westfalens, 39, 205–210.
469Schlüter, C., 1884. Über interessante neue Petrefakten. Verhandlungen des naturhistorischen Vereines der preussischen Rheinlande und Westfalens, 41, 79–84.
470Schlüter, C., 1885. Über neue Korallen aus dem Mittel-Devon der Eifel. Verhandlungen des naturhistorischen Vereines der preussischen Rheinlande und Westfalens, 42, 6–13.
471Schlüter, C., 1889. Anthozoen des rheinischen Mittel-Devon. Abhandlungen zur geologischen Specialkarte von Preussen und der Thüringischen Staaten, 8/4, 1–207.
472Schröder, S., 1995a. Die Korallenfauna des Kirchen-Berges (Freilingen-Formation) in der Blankenheimer Mulde (Rheinisches Schiefergebirge/Eifel). Münstersche Forschungen zur Geologie und Paläontologie, 77, 373–421.
473Schröder, S., 1995b. Rugose Korallen aus der Freilingen-Formation der Dollendorfer Mulde (Ober-Eifelium/Mitteldevon; Eifel/Rheinisches Schiefergebirge). Senckenbergiana lethaea, 75, 33–75.
474Schröder, S., 1998. Rugose Korallen und Stratigraphie des oberen Eifelium und unteren Givetium der Dollendorfer Mulde/Eifel (Mittel-Devon; Rheinisches Schiefergebirge). Courier Forschungsinstitut Senckenberg, 208, 1–135.
475Schröder, S., 2004. Devonian rugose corals from the Karakorum Mountains (Northern Pakistan). Rivista Italiana di Paleontologia e Stratigrafia, 110, 605–641.
476Schröder, S., 2005. Stratigraphie und Systematik rugoser Korallen aus dem Givetium und Unter-Frasnium des Rheinischen Schiefergebirges (Sauerland/Bergisches Land). Zitteliana, B25, 39–116.
477Schröder, S. & Kazmierczak, M., 1999. The Middle Devonian “coral reef” of Ouihlane (Morocco) - New data on the geology and rugose coral fauna. Geologica et Palaeontologica, 33, 93–115.
478Schröder, S. & Salerno, C., 2001. Korallenfauna und Fazies givetischer Kalksteinabfolgen (Cürten-/Dreimühlen-Formation) der Dollendorfer Mulde (Devon, Rheinisches Schiefergebirge/Eifel). Senckenbergiana lethaea, 81, 111–133. https://doi.org/10.1007/BF03043297
479Schulz, E., 1883. Die Eifelkalkmulde von Hillesheim. Nebst einem palaeontologischen Anhang. Jahrbuch der Königlich Preussischen geologischen Landesanstalt und Bergakademie, Abhandlungen, 1882/3, 158–250.
480Scrutton, C.T., 1968. Colonial Phillipsastraeidae from the Devonian of South-East Devon, England. Bulletin of the British Museum (Natural History), Geology, 15/5, 181–281.
481Scrutton, C.T., 1985. Subclass Rugosa. In Murray, J.W. (ed.), Atlas of Invertebrate Macrofossils. Longman and the Palaeontological Association, London, 13–31.
482Soshkina, E.D., 1949. Devonskie korally Rugosa Urala. Trudy Paleontologicheskogo Instituta, 15/4, 1–160. [In Russian].
483Soshkina, E.D., 1952. Opredelitel devonskikh chetyrekhluchevykh korallov. Trudy Paleontologicheskogo Instituta, 39, 1–127. [In Russian].
484Spassky, N. Ya., 1959. Korally Rugosa v nizhnem i srednem devone Urala. Zapiski Leningradskogo Gornogo Instituta, 36/2, 15–47. [In Russian].
485Stumm, E.C., 1968. The corals of the Middle Devonian Tenmile Creek Dolomite of Northwestern Ohio. Contributions from the Museum of Palaeontology, The University of Michigan, 22/3, 37–44.
486Sun, Y.C., 1958. The Upper Devonian coral faunas of Hunan. Palaeontologia Sinica, 144 new series B, 8, 1–28.
487Taylor, P.W., 1951. The Plymouth Limestone. Transactions of the Royal Geological Society of Cornwall, 18, 146–214.
488Tsien, H.H., 1969. Contribution à l'étude des Rugosa du Couvinien de la Région de Couvin. Mémoires de l’Institut Géologique de l’Université de Louvain, 25, 1–174.
489Vollbrecht, E., 1926. Die Digonophyllinae aus dem unteren Mitteldevon der Eifel. Eine morphologisch-chronologische Studie. 1. Teil. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 55, Beilage-Band, Abteilung B, 189–273.
490Walther, C., 1929. Untersuchungen über die Mitteldevon-Oberdevongrenze. Zeitschrift der Deutschen Geologischen Gesellschaft, 80, 97–152.
491Webster, C.L., 1889. Description of a new genus of corals from the Devonian rocks of Iowa. The American Naturalist, 23/272, 710–712. https://doi.org/10.1086/274991
492Weddige, K., 1996. Devon-Korrelationstabelle. Senckenbergiana lethaea, 76, 267–286. https://doi.org/10.1007/BF03042853
493Wedekind, R., 1922a. Beiträge zur Kenntnis der Mesophyllen. Palaeontologische Zeitschrift, 4, 48–63. https://doi.org/10.1007/BF03041558
494Wedekind, R., 1922b. Zur Kenntnis der Stringophyllen des oberen Mitteldevon. Sitzungsberichte der Gesellschaft zur Beförderung der gesamten Naturwissenschaften zu Marburg, 1921/1, 1–16.
495Wedekind, R., 1923. Die Gliederung des Mitteldevons auf Grund von Korallen. Sitzungsberichte der Gesellschaft zu Beförderung der gesamten Naturwissenschaften zu Marburg, 1922, 24–35.
496Wedekind, 1924. Das Mitteldevon der Eifel. Eine biostratigraphische Studie. I. Teil. Die Tetrakorallen des unteren Mitteldevon. Schriften der Gesellschaft zur Beförderung der gesamten Naturwissenschaften zu Marburg, 14/3, 1–93.
497Wedekind, R., 1925. Das Mitteldevon der Eifel. Eine biostratigraphische Studie. II. Teil. Materialien zur Kenntnis des mittleren Mitteldevon. Schriften der Gesellschaft zur Beförderung der gesamten Naturwissenschaften zu Marburg, 14/4, 1–85.
498Wedekind, R. & Vollbrecht, E., 1931. Die Lythophyllidae des mittleren Mitteldevon der Eifel. Palaeontographica, 75, 81–110.
499Weissermel, W., 1897. Die Gattung Columnaria und Beiträge zur Stammergeschichte der Cyathophylliden und Zaphrentiden. Zeitschrift der Deutschen Geologischen Gesellschaft, 49, 865–888.
500Weyer, D., 2008. Revision des Ludwig/Kunth-Gesetzes zur Septeninsertion der Supraordo Rugosa (Anthozoa, Ordoviz-Perm). Abhandlungen und Berichte für Naturkunde, 30, 85–145.
501Wrzolek, T., 1993. Rugose corals from the Devonian Kowala Formation of the Holy Cross Mountains. Acta Palaeontologica Polonica, 37, 217–254.
502Manuscript received 27.10.2021, accepted in revised form 03.06.2022, available online 22.08.2022.
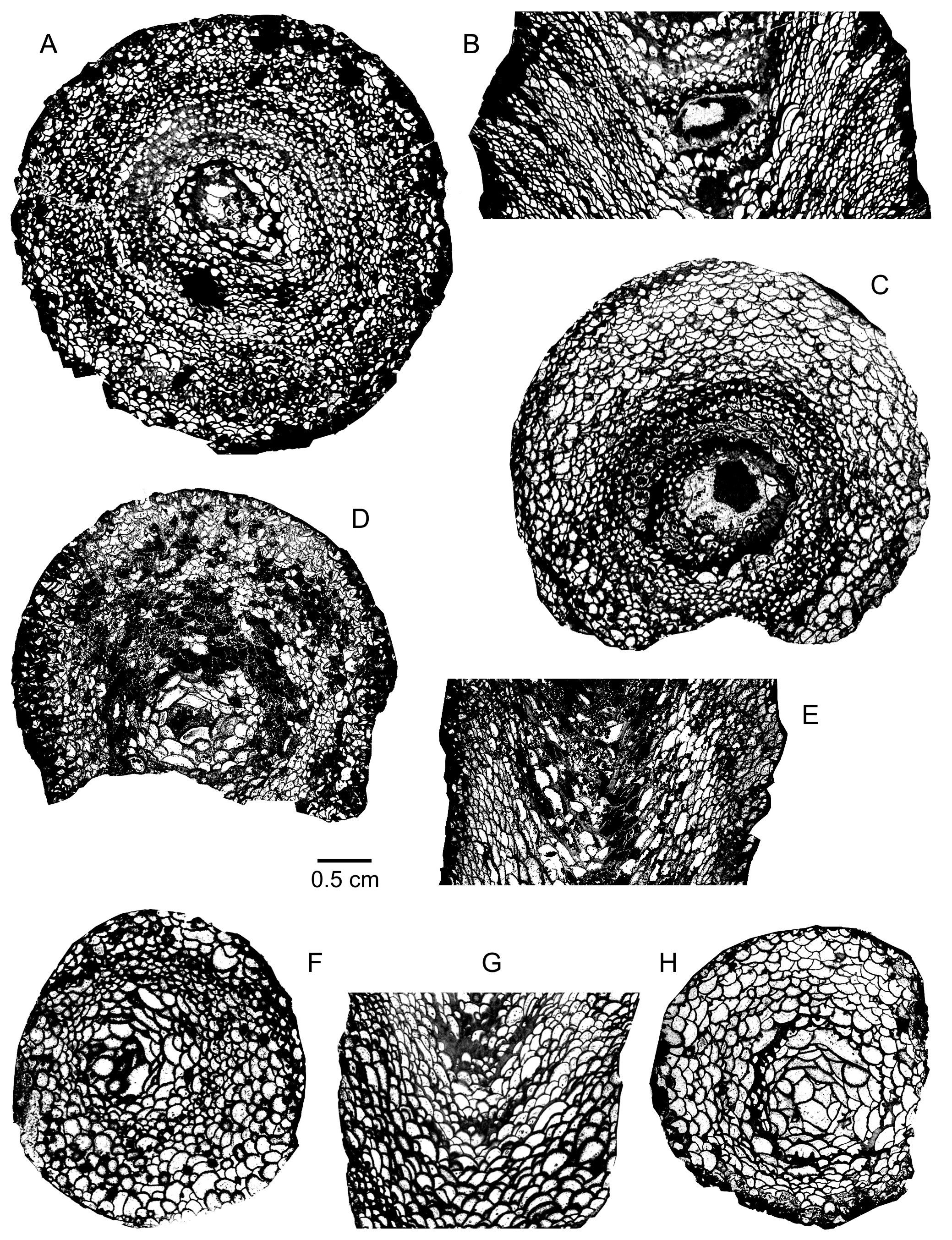
Plate 1. A–C: Marennophyllum praecipuum (Wedekind & Vollbrecht, 1931). A–B: IRScNB a 13527, Meerbüsch EIF395, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. C: IRScNB a 13528, Meerbüsch EIF346, Binz Member of the Dreimühlen Formation; transverse section. D–E: Marennophyllum sp. 2. IRScNB a13532, Meerbüsch EIF403, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. F–H: Marennophyllum wenningi n. sp. F–G: Holotype, IRScNB a13529, Meerbüsch EIF396, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. H: Paratype, IRScNB a13530, Meerbüsch EIF468, Binz Member of the Dreimühlen Formation; transverse section. Magnification x 2.

Plate 2. A–B: Marennophyllum sp. 1. IRScNB a13531, Meerbüsch EIF463, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. C: Acanthophyllum vermiculare (Goldfuss, 1826). IRScNB a13538, Houyet MC-1981-5-Z175, Hanonet Formation at Pondrôme, on the south side of the Dinant Synclinorium in Belgium; transverse section. D–G: Atelophyllum laeve (Schulz, 1883). D–E: IRScNB a13533, Meerbüsch EIF451, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. F–G: IRScNB a13534, Meerbüsch EIF394, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. Magnification x 2.
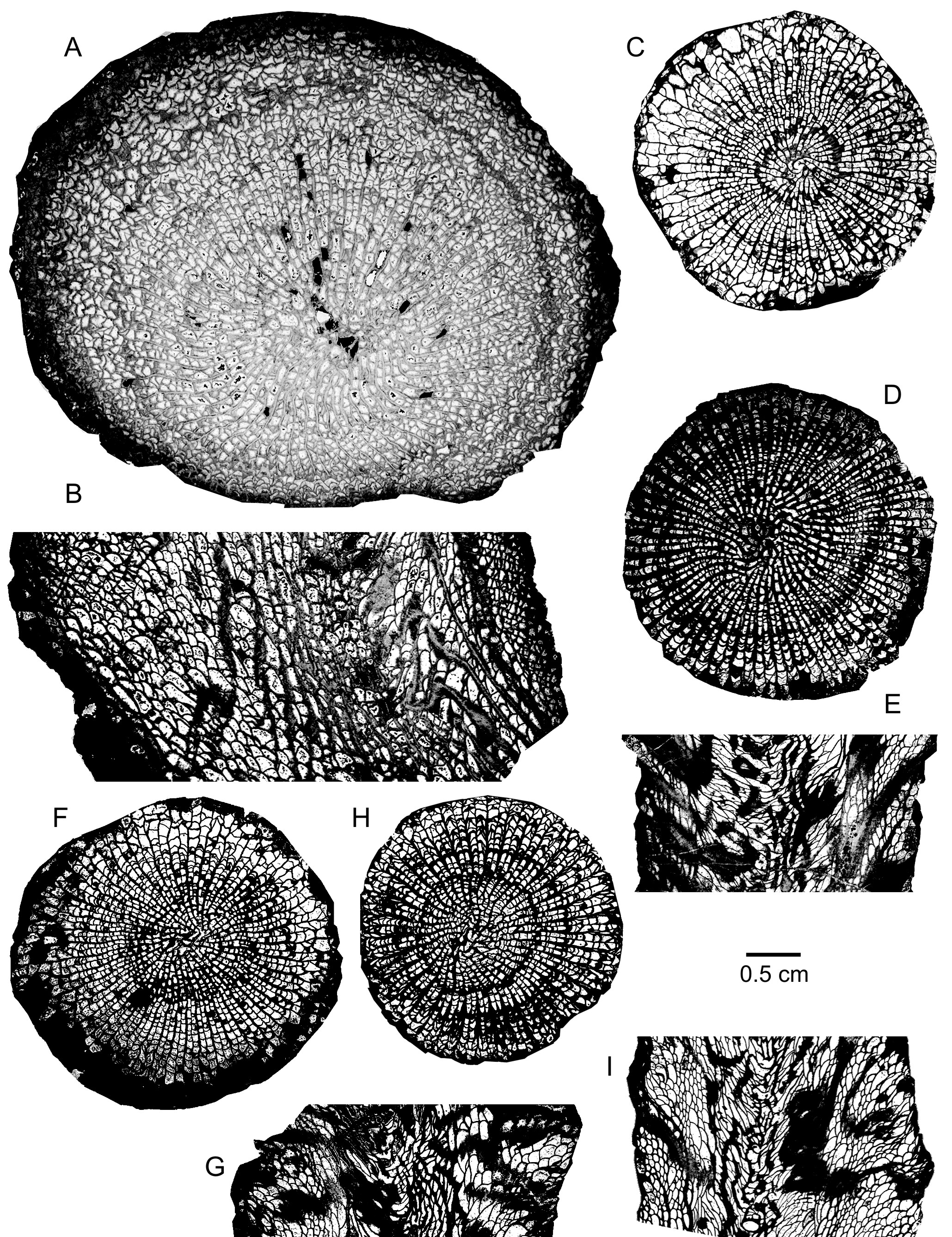
Plate 3. A–B: Atelophyllum cf. maximum (Schlüter, 1882). IRScNB a13535, Meerbüsch EIF423, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. C–I: Acanthophyllum vermiculare (Goldfuss, 1826). C: IRScNB a13539, Meerbüsch EIF453, Binz Member of the Dreimühlen Formation; transverse section. D–E: IRScNB a13540, Meerbüsch EIF464, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. F–G: IRScNB a 13541, Meerbüsch EIF467, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. H–I: IRScNB a13542, Meerbüsch EIF385 Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. Magnification x 2.
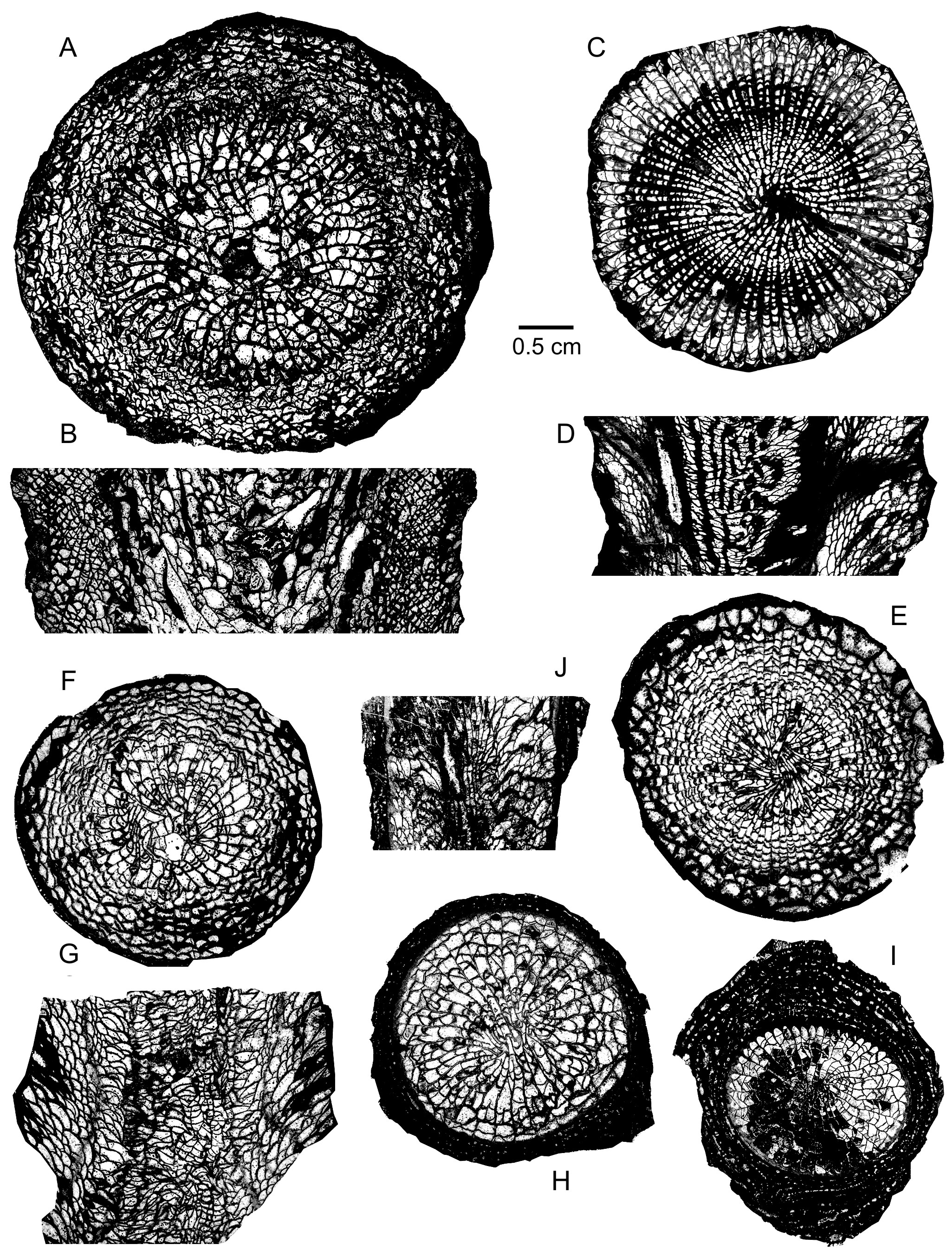
Plate 4. A–B: Atelophyllum cf. emsti (Wedekind, 1922). IRScNB a 13536, EIF431, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. C–D: Acanthophyllum heterophyllum (Milne-Edwards & Haime, 1851). IRScNB a13537, Meerbüsch EIF391, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. E–G: Grypophyllum convolutum (Wedekind, 1925). E: IRScNB a13543, Meerbüsch EIF433, Galgenberg Member of the Dreimühlen Formation; transverse section. F–G: IRScNB a13544, Meerbüsch EIF416, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. H–J: Grypophyllum schroederi n. sp. H: Paratype IRScNB a13546, Meerbüsch EIF448, Ley Member of the Dreimühlen Formation; transverse section. I–J: Holotype, IRScNB a13545, Meerbüsch EIF449, Ley Member of the Dreimühlen Formation; transverse and longitudinal sections. Magnification x 2.
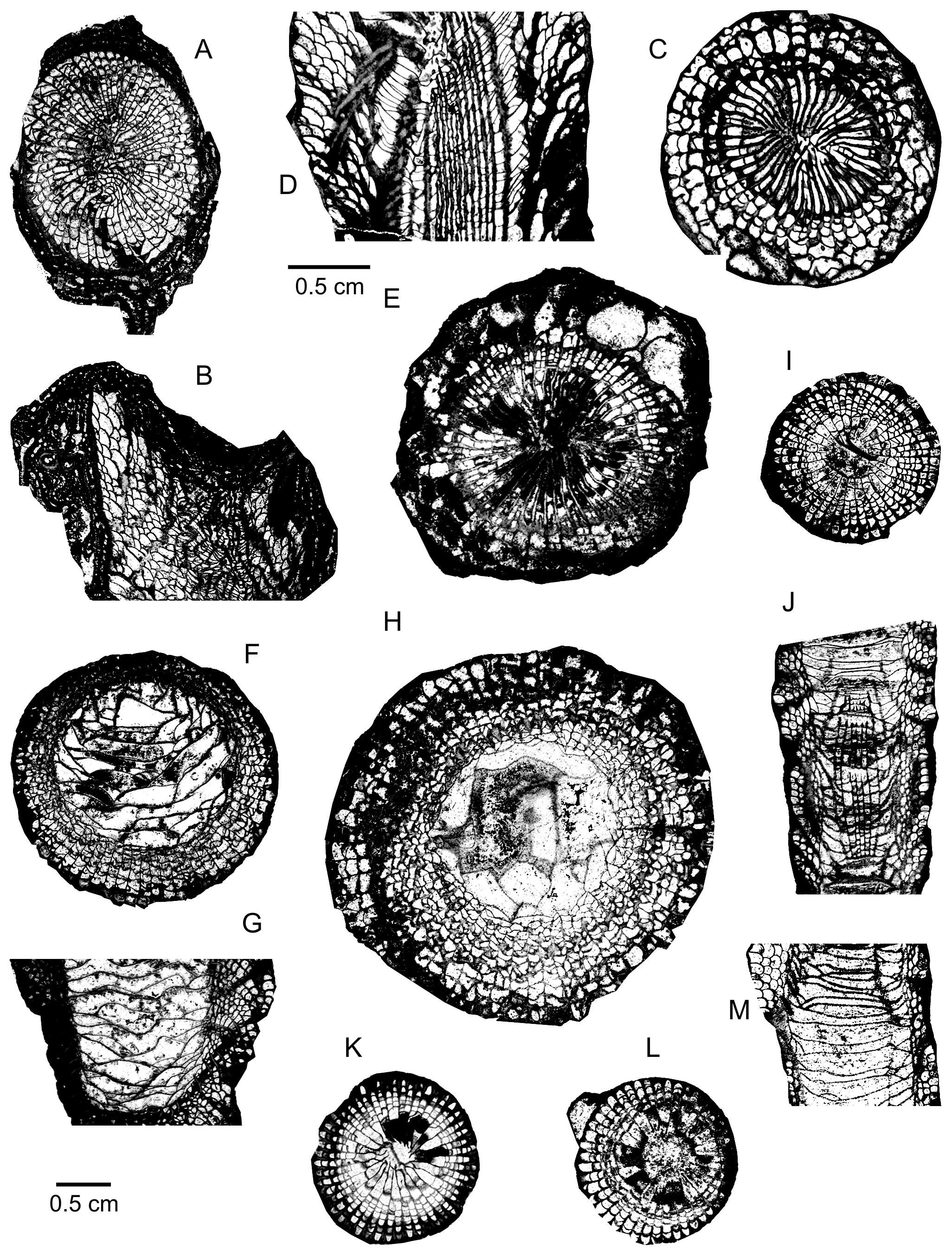
Plate 5. A–B: Grypophyllum schroederi n. sp. paratype, IRScNB a13547, Meerbüsch EIF446, Ley Member of the Dreimühlen Formation; transverse and longitudinal sections. C–E: Stringophyllum acanthicum (Frech, 1885). C–D: IRScNB a13548, Meerbüsch EIF425, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. E: IRScNB a13549, Meerbüsch EIF426, Galgenberg Member of the Dreimühlen Formation; transverse section. F–H: Heliophyllum cf. cribellum Oliver & Sorauf, 2002. F–G: IRScNB a13550, Meerbüsch EIF435, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. H: IRScNB a13551, Meerbüsch EIF371, Galgenberg Member of the Dreimühlen Formation; transverse section. I–M: Disphyllum caespitosum (Goldfuss, 1826). I–J: IRScNB a13552, Meerbüsch EIF351, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. K: IRScNB a13553, Meerbüsch EIF352, Binz Member of the Dreimühlen Formation; transverse section. L–M: IRScNB a13554, Meerbüsch EIF392, Binz Member of the Dreimühlen Formation: transverse and longitudinal sections. Magnification x 2 for figures A–B, F–H and x 3 for figures C–E, I–M.
Plate 6. A–G: Aristophyllum luetti Coen-Aubert, 1997. A–B: IRScNB a13555, Meerbüsch EIF455, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. C–D: IRScNB a13556, Meerbüsch EIF454, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. E: IRScNB a13557, Meerbüsch EIF458, Binz Member of the Dreimühlen Formation; transverse section. F–G: IRScNB a13558, Meerbüsch EIF462, Binz Member of the Dreimühlen Formation; transverse and longitudinal sections. H: Aristophyllum sp. IRScNB a13559, Meerbüsch EIF438, Galgenberg Member of the Dreimühlen Formation; transverse section. I–J: Macgeea mistiaeni n. sp. Holotype, IRScNB a13563, Meerbüsch EIF410, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. K–L: Spinophyllum sp. IRScNB a13560, Meerbüsch EIF432, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. Magnification x 2 for figures A–H and x 3 for figures I–L.
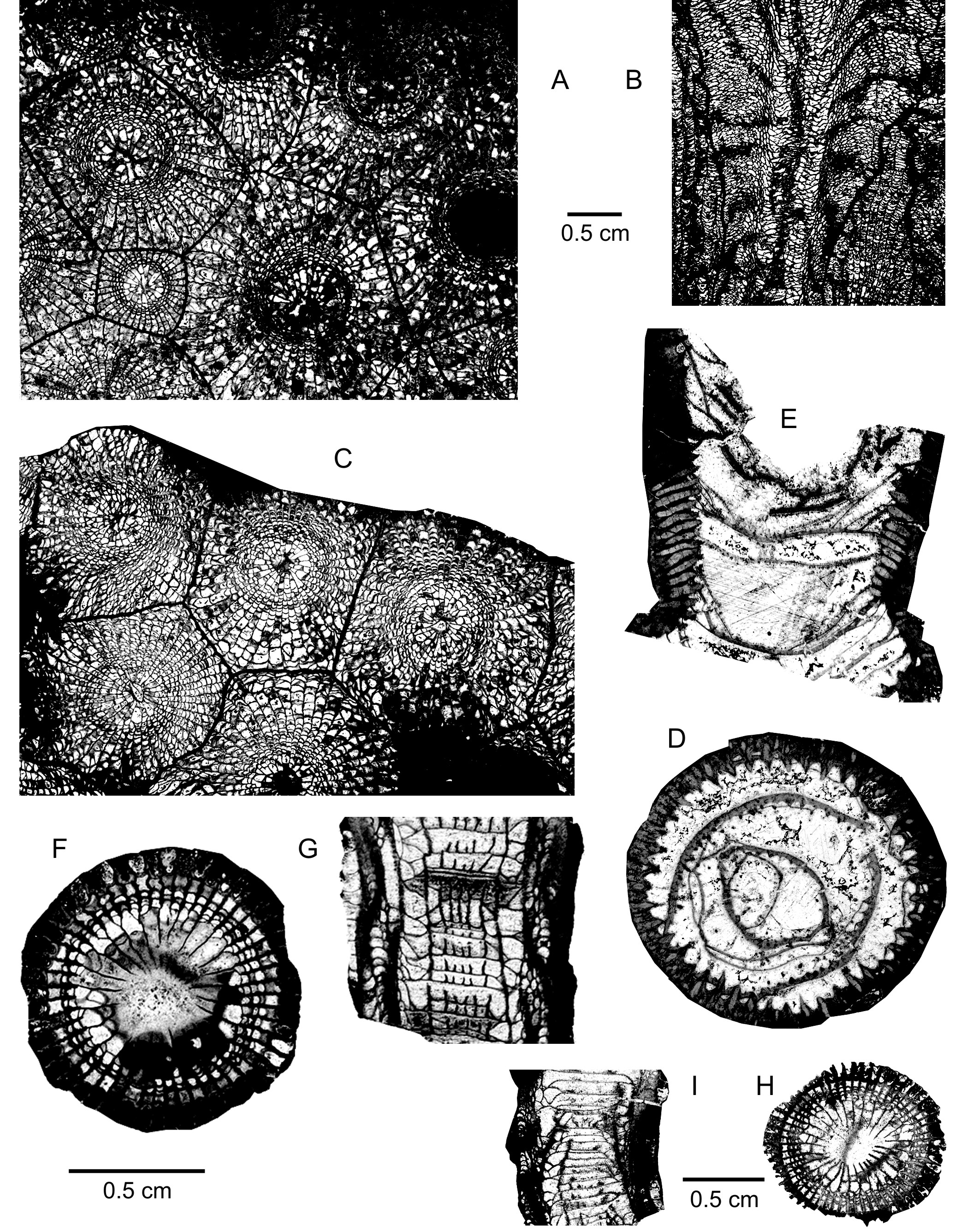
Plate 7. A–C: Argutastrea briceae (Rohart, 1988). A–B: IRScNB a13561, Meerbüsch EIF429, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. C: IRScNB a13562, Meerbüsch EIF341, Galgenberg Member of the Dreimühlen Formation; transverse section. D–E: Pselophyllum sp. IRScNB a13566, Meerbüsch EIF413, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. F–I: Macgeea mistiaeni n. sp. F–G: Paratype, IRScNB a13564, Meerbüsch EIF373, Galgenberg Member of the Dreimühlen Formation: transverse and longitudinal sections. H–I: Paratype, IRScNB a13565, Meerbüsch EIF379, Galgenberg Member of the Dreimühlen Formation; transverse and longitudinal sections. Magnification x 2 for figures A–E, x 3 for figures H–I and x 5 for figures F–G.
