


- Home
- Volume 26 (2023)
- number 1-2
- A Santonian record of the nautilid cephalopod Angulithes westphalicus (Schlüter, 1872) from the subsurface of the Campine, north-east Belgium, with comments on regional lithostratigraphic problems


View(s): 2766 (27 ULiège)
Download(s): 1234 (9 ULiège)

A Santonian record of the nautilid cephalopod Angulithes westphalicus (Schlüter, 1872) from the subsurface of the Campine, north-east Belgium, with comments on regional lithostratigraphic problems

Abstract
Newly recognised material of the Late Cretaceous nautilid Angulithes westphalicus is described from the subsurface of the eastern part of the Campine in north-east Belgium. This constitutes the first formal documentation of this genus and species from the Cretaceous of Belgium, having been identified amongst a large suite of fossils collected from the Voort Shafts I & II of the Zolder colliery during the first half of the twentieth century. The specimens originate from an interval of marine calcareous sand with a marly glauconiferous base, dated as late middle Santonian (Gonioteuthis westfalicagranulata belemnite Zone) and for which a deepening of the depositional environment is documented. Lithostratigraphically, the specimens occur within the Vaals Formation, within the upper part of the Asdonk Member or alternatively within the lower part of the Sonnisheide Member. The early Campanian age of the Asdonk Member suggested previously is refuted, the age of the Sonnisheide Member needs further study. The position of the siphuncle in A. westphalicus is illustrated for the first time; it is positioned closer to the venter than the dorsum, which confirms the close evolutionary relationship with Angulithes galea, which ranges from the upper Turonian to middle Coniacian in central Europe.
Table of content
1. Introduction
1At the time of its introduction, the nautilid cephalopod Angulithes westphalicus (Schlüter, 1872) (as Nautilus westphalicus) was known exclusively from lower Campanian beds near Dülmen (Westphalia, Germany). Recent data have shown this species to range from the early Santonian to late Campanian, with records from Germany, Austria, Poland and Spain, and possibly Hungary (Wilmsen, 2000; Frank, 2010; Summesberger, 2017a). Its supposed evolutionary ancestor, Angulithes galea (Fritsch in Fritsch & Schlönbach, 1872), ranges from the upper Turonian to the middle Coniacian, with records from the Czech Republic and Germany (Frank, 2010). While the number of documented occurrences has grown considerably for both species in the last two decades, our current knowledge is still largely biased and notably hampers a full understanding of the palaeoecological constraints defining the range of both taxa. This bias may in part be the result of the commonly poor state of preservation of nautilids in chalks, which is the predominant sedimentary facies of the European Upper Cretaceous. In this facies, nautilids are not only generally rare to extremely rare, these nearly always comprise composite moulds of their originally aragonitic shells, often also lacking in situ preserved buccal mass elements or other soft tissues, with the exception of isolated finds of calcified tips of their upper and lower jaws (rhyncholites and conchorhynchs; Mironenko et al., 2022). Only rarely (e.g., in siliceous porous limestones, or ‘opoka’, in Poland and Ukraine), may a thin nacreous lining be present in places on the mould (Janiszewska et al., 2018). In condensed intervals, the siphuncle may be partially phosphatised or calcified. These composite moulds are invariably distorted to varying degrees by sediment compaction. Moreover, their inner whorls are poorly to very poorly preserved, and the embryonic stage is generally unrecognisable. In addition, the once soupy nature of the chalky sea floor explains that the shells may occupy a wide array of orientations to the bedding surface. Therefore, compaction by dewatering and overburden had a marked impact on the (already fairly reduced) set of taxonomic characters, such as whorl proportions and suture details that are needed for species and genus identification in Late Cretaceous nautilids (Kummel, 1956).
2However, also in more marly sedimentary rocks, and even in sandier lithologies, imperfect preservation (which hampers assessment of taxonomically important features), is the rule rather than the exception. This means that taxonomic studies of Late Cretaceous nautilids are far from straightforward, explaining why their taxonomy and geographical distribution and stratigraphical ranges are largely understudied. This holds certainly true for nautilids from Belgium, irrespective of their stratigraphical age.
3Here we document A. westphalicus for the first time from the Upper Cretaceous of the Campine (Fig. 1). Specimens here assigned to this species were recognised within a large and mostly unstudied collection (>7,000 specimens, including ammonites, nautilids, belemnites, bivalves, gastropods, decapod crustaceans, brachiopods and echinoderms; see Goolaerts et al., 2013) recovered in the 1920–1930s by the staff of the Royal Belgian Institute of Natural Sciences (RBINS) from Upper Cretaceous levels of the ‘Smectite de Herve – Campanien’ (as noted on labels with specimens and collection drawers) penetrated in colliery shafts in the eastern part of the Campine, in particular of Shafts I and II at Zolder (Voort) and Houthalen (Fig. 1), from between 400 and 600 m below surface. The stratigraphy of these occurrences is here also discussed because there are inconsistencies in the most recent lithostratigraphical schemes for the Cretaceous of the Campine. In an attempt to obtain additional morphological data, some of these specimens have been imaged by X-ray micro-focus Computed Tomography (micro-CT).
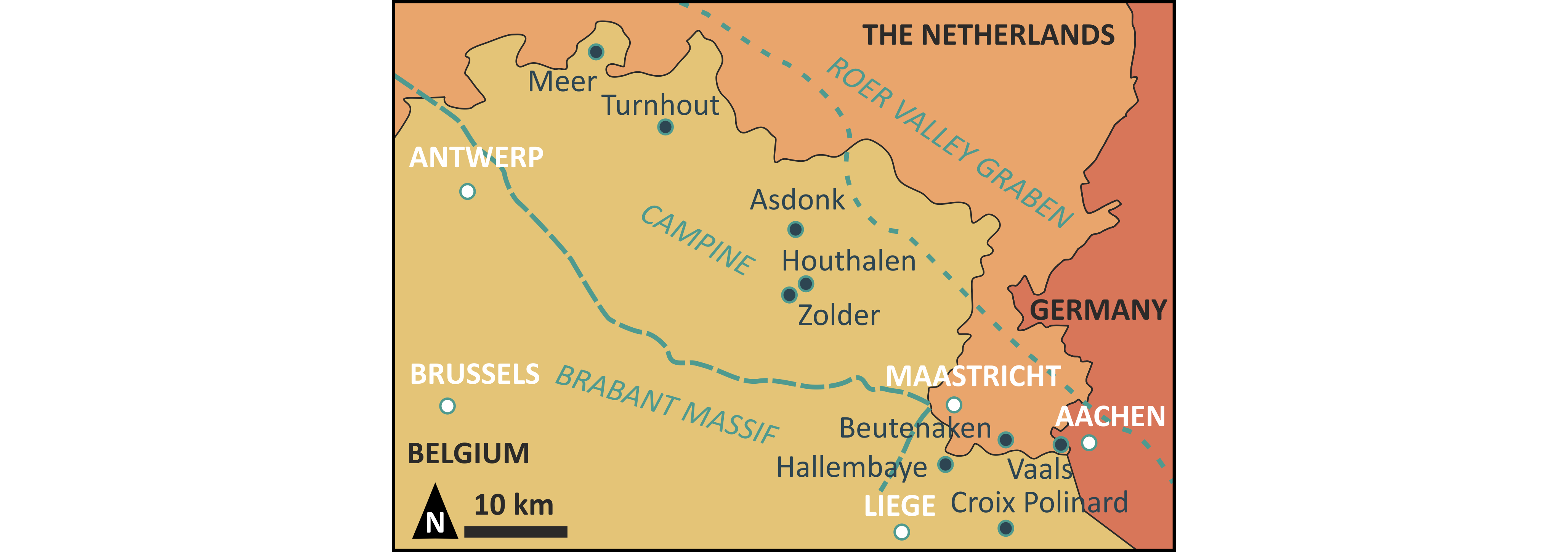
Figure 1. Map showing the location of the Zolder and Houthalen collieries and other nearby localities discussed in the text, as well as the position of the Campine, to the north-east of the Brabant Massif and to the west of the Roer Valley Graben, and most major cities for orientation.
2. Material and methods
4All specimens studied are housed in the palaeontology collections of the Royal Belgian Institute of Natural Sciences (Brussels) and are referred to here by catalogue numbers with the prefix IRSNB.
5Several specimens have been analysed using the RBINS micro-CT RX EasyTom 150 (from RX Solutions, France). Details on used voltages, spot size, beam filters and voxel sizes for each of these specimens may be found in Table 1. After scanning, extraction into 16-bit TIFFs has been performed with X-Act software, 3D-rendering and segmentation with Dragonfly ORS. Figures have been produced with the ‘export screenshot’ function, after optimising the 3D rendering by adjusting the histogram, contrast, shading, light source position, hard gradient and turning off all annotations, with the exception of scale bars. In the final step, image processing software has been used for scaling to 600 dpi. The primary scanning data and the set of Y slices of these specimens are stored on Belspo’s LTP platform and can be consulted upon request to the RBINS palaeontology collection manager. In addition to this, 3D-meshes of the outer surface of IRSNB 11506, IRSNB 11505, and IRSNB 11508 (based on micro-CT imaging) can be consulted via the RBINS Virtual Collections Platform (http://virtualcollections.naturalsciences.be/).
Table 1. Details of micro-CT imaging (acquisition mode, power of X-ray source [beam kV and µA] and obtained voxel sizes [in µm]) of scanning results for each specimen scanned with the RBINS RX EasyTom 150 studied in the present paper. For all scans, a 0.4 mm Cu filter was placed between the beam and the specimen.
|
Specimen N° |
Acquisition mode |
Beam kV |
Beam µA |
Voxel size (µm) |
Additional specimen number |
|
IRSNB 11504 |
large spot |
150 |
500 |
80.1567 |
Invert-8748-0145 |
|
IRSNB 11505 |
middle spot |
150 |
200 |
56.9206 |
Invert-8748-0146 |
|
IRSNB 11506 |
large spot |
150 |
500 |
96.1503 |
Invert-8748-0147 |
|
IRSNB 11508 |
large spot |
150 |
390 |
72.1817 |
Invert-8821-0093 |
6The terminology of the shell characters used in this paper is summarized in Figure 2. Measurements have been taken either traditionally (i.e., vernier calliper) or digitally from 2D- and 3D-renderings in DragonflyORS software. Specimens have also been photographed conventionally in white light, and some also after coating with NH4Cl.
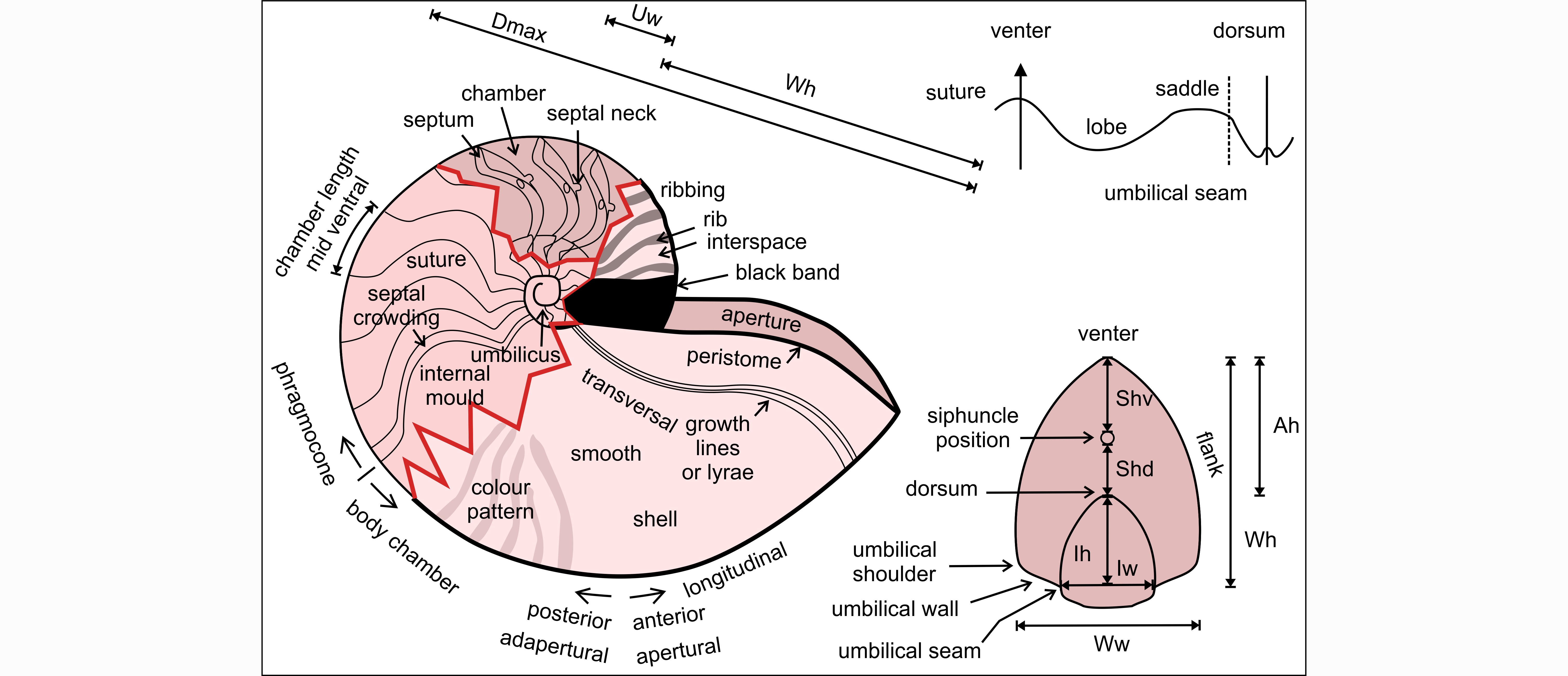
Figure 2. Terminology of nautilid shells. Abbreviations: A: aperture; D: diameter; d: dorsal; h: height; I: imprint zone; max: maximal; S: siphuncle; U: umbilicus; v: ventral; W: whorl; w: width.
7Boreholes (Table 2) are referred to with their corresponding DOV (‘Databank Ondergrond Vlaanderen’) and GSB (Geological Survey of Belgium) archive numbers. In some cases, they are referred to with their coal exploration borehole numbers (KS and/or KOEN) as well.
8Author citations are supplied only for taxa that are taxonomically discussed here. For those only mentioned discussing age dating, author citations can be found in the papers from which the data have been taken.
Table 2. Overview of all boreholes discussed in the text, with their corresponding DOV archive (‘Databank Ondergrond Vlaanderen’), GSB archive (Geological Survey of Belgium) and coal exploration (KS and KOEN) numbers.
|
DOV (with link) |
GSB |
KS |
KOEN |
cited in the paper as follows |
|
062W0205 |
|
B 79 |
borehole near future Voort Shaft II Zolder colliery, Schmitz & Stainier (1910) |
|
|
062W0226 |
|
|
borehole near Voort Shaft I Zolder colliery |
|
|
062W0227 |
|
|
borehole near Voort Shaft I Zolder colliery |
|
|
062E0250 |
|
|
borehole near Shafts I & II Houthalen colliery |
|
|
062E0251 |
|
|
borehole near Shafts I & II Houthalen colliery |
|
|
062E0248 |
|
B 95 |
borehole near Shafts I & II Houthalen colliery |
|
|
062E0270 |
KS 9 |
B 153 |
borehole KS 9 on figs 1, 5 of Lagrou et al. (2005) |
|
|
062E0269 |
KS 11 |
B 151 |
borehole KS 11 on figs 1, 5 of Lagrou et al. (2005) |
|
|
062W0304 |
KS 27 |
B 207 |
borehole KS 27 on figs 1, 5 of Lagrou et al. (2005) |
|
|
062E0280 |
KS 29 |
B 206 |
borehole KS 29 on figs 1, 5 of Lagrou et al. (2005) |
|
|
047W0265 |
KS 34 |
B 196 |
borehole KS 34 on figs 1, 5 of Lagrou et al. (2005) |
|
|
062E0281 |
KS 44 |
B 209 |
borehole KS 44 on figs 1, 5 of Lagrou et al. (2005) |
|
|
062E0282 |
KS 46 |
B 208 |
borehole KS 46 on figs 1, 5 of Lagrou et al. (2005) |
|
|
047W0267 |
KS 37 |
|
borehole KS 37, Asdonk (Leopoldsburg), Asdonk Member stratotype |
|
|
062E0280 |
KS 29 |
|
borehole KS 29, Houthalen-Helchteren, Sonnisheide Member stratotype |
|
|
007E0205 |
|
|
borehole Meer |
|
|
017E0225-120 |
|
|
borehole Turnhout (Turnhout Zwemdok), Gulinck (1954) |
3. Systematic palaeontology
9Subclass Nautiloidea Agassiz, 1847
10Order Nautilida Agassiz, 1847
11Genus Angulithes de Montfort, 1808
12Type species. Angulithes triangularis de Montfort, 1808, Cenomanian of Le Havre, Normandy (France), by original designation (de Montfort, 1808, p. 7).
13Angulithes westphalicus (Schlüter, 1872)
14(Figs 3A–J,4A–E, 5A, 6)
151872 Nautilus westphalicus Schlüter, p. 13.
161876 Nautilus westphalicus Schlüter; Schlüter, p. 175, pl. 47, figs 1, 2.
171906 Nautilus westphalicus Schlüter; Müller & Wollemann, p. 1, pl. 1, figs 1, 2.
181956 Angulithes westphalicus (Schlüter); Kummel, p. 475, text-fig. 33I.
191960 Angulithes (Angulithes) westphalicus (Schlüter); Wiedmann, p. 186, pl. 21, fig. O.
201975 Nautilus westphalicus (Schlüter); Shimansky, p. 136.
211991 Deltoidonautilus westphalicus (Schlüter, 1876); Riegraf & Scheer, p. 426.
221999 Deltoidnautilus westphalicus (Schlüter, 1872c); Wittler et al., p. 37, figs 51a, b, 52a, b.
232000 Angulithes westphalicus (Schlüter); Wilmsen, p. 37, pl. 3, fig. 1a, b; pl. 5, figs 6, 20.
24?2001 Angulithes cf. westphalicus (Schlüter, 1872); Fözy, p. 34, pl. 5.
252010 Angulithes westphalicus (Schlüter, 1872); Frank, p. 490, fig. 3A–N.
262013 Angulithes westphalicus (Schlüter, 1872); Frank et al., p. 797, fig. 7.
272017a Angulithes westphalicus (Schlüter, 1872); Summesberger et al., p. 10, pl. 1, figs 1–3; pl. 2, figs 1–3; pl. 3, figs 4, 5; pl. 4, figs 1–3, table 1.
282017b Angulithes westphalicus (Schlüter, 1872); Summesberger et al., p. 121.
29Type. Lectotype, designated by Frank (2010, p. 490), is specimen GMB 97 (Goldfuss Museum, Bonn, Germany), the original of Schlüter (1872, 1876) from the lower Campanian Dülmen Formation, Scaphites binodosus Zone, of Dülmen, Westphalia, Germany.
30Material. Five specimens from Zolder colliery: IRSNB 11504 from Voort Shaft I at a depth of 582–584.30 m (Fig. 3A–C), IRSNB 11505 (Fig. 5A) and IRSNB 11506 (Fig. 3E–J) from Voort Shaft I at a depth of 579.50–582 m, and IRSNB 11507 (Fig. 3D) and IRSNB 11508 (Fig. 4A–E) from Voort Shaft II at a depth of 582.50 m.
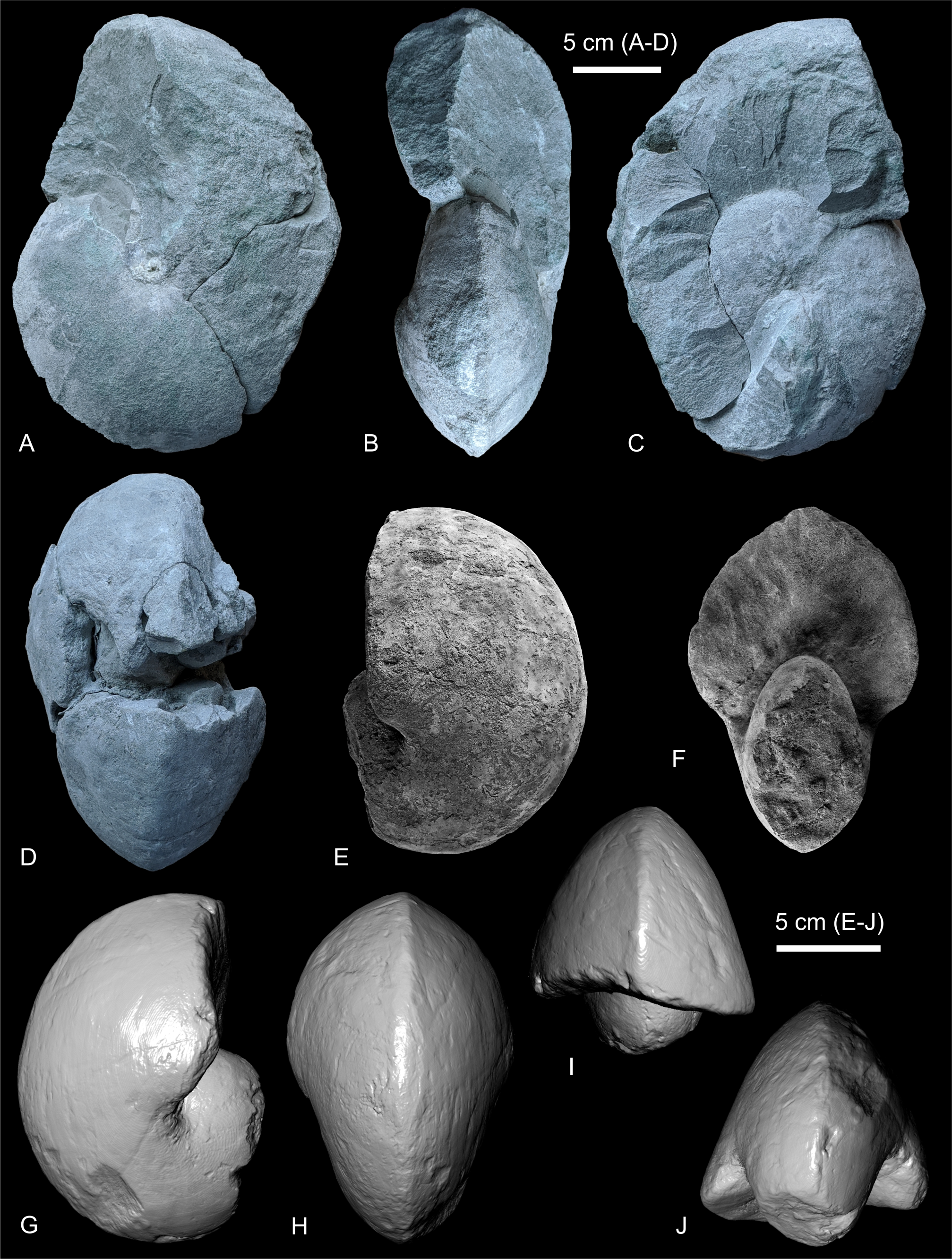
Figure 3. Angulithes westphalicus (Schlüter, 1872) from the Voort Shafts I and II, Zolder colliery. A–C. Specimen IRSNB 11504, photographed under natural light, from Shaft I, at a depth between 582–584.30 m. D. Keeled venter of Specimen IRSNB 11507, photographed under natural light, from Shaft II at a depth of 582.50 m. E–J. Specimen IRSNB 11506, from Shaft I at a depth of 579.50–582 m, photographed under white light after coating with NH4Cl (E, F) and screenshots of a 3D mesh obtained from segmenting a micro-CT imaging data set.
31Description. All specimens are incomplete internal moulds, in calcareous sandstone preservation, with fine-grained translucent and mostly angular quartz grains and low amounts of glauconite. Glauconite grains are mostly smaller than quartz, except on some of the surfaces of IRSNB 11504, where larger glauconite grains can be seen. The matrix of IRSNB 11505 and IRSNB 11504 also has a significant amount of clay. Specimens are light grey coloured with greenish hues (Fig. 3A–D; 5A), with IRSNB 11504 being the most greenish in colour (Fig. 3A–C). Specimen IRSNB 11506 is slightly darker grey in colour.
32Shell remains have been noted only in recrystallised or partially mineralised form, visible to the naked eye on the outer broken surface of septa of specimens IRSNB 11508 and IRSNB 11504 (only partially near the umbilicus) and discernable internally (Fig. 4E) as revealed by micro-CT imaging of IRSNB 11508. Overall mediocre preservation characterises these specimens, in particular where their inner whorls are concerned. Specimen IRSNB 11505 has a part of the inner whorls replaced by a cavity lined with calcite crystals, but also in the best-preserved specimen (IRSNB 11506) has incomplete preservation of the septa been documented by micro-CT imaging, precluding assessment of the ontogenetic trajectory or total chamber count per whorl. None of the micro-CT imaging data sets has identified the presence of (calcified) parts of the buccal mass, such as rhyncholites, conchorynchs or radulae. Lithological and taphonomic observations point to two (or three) nearby beds of origin of these five specimens.
33The external shape is that of an involute oval nautilicone in which the configuration of the venter transforms from rounded to angularly sharpened between conch diameters of 67 and 100 mm (IRSNB 11506; venter not preserved between both diameters). In IRSNB 11505, the onset of this change is slightly earlier ontogenetically, being seen, albeit faintly, from a Wh 55.0 mm onwards. The body chamber is large and elongated, measuring about half a whorl in IRSNB 11504 (D last observable septum ~165.6 mm) and about 274 degrees or two-thirds of the last whorl in IRSNB 11506 (based on the absence of preserved septa visible on the surface as well as internally by micro-CT imaging, with several 20 mm-large bivalves inside, septal crowding not present; D at last septum is 140.0 mm). Adult modifications, such as a re-rounding of the venter, as documented by Frank (2010, figs 3D–I) in specimens of diameters in excess of 250 mm are not observed, not in IRSNB 11504 at Dmax of 266.2 mm, nor in IRSNB 11506 at Dmax of 194.2 mm (Ww 128.1 mm and Wh 126.5 mm at this D). Ribbing or folding on the flanks and venter has not been seen either.
34Compaction-related deformation resulted in an angle of 13 degrees between the opposite umbilical shoulders for IRSNB 11506.
35The position of the siphuncle is seen only in IRSNB 11508 (Fig. 4B–E), a fragmentary body chamber with remains of five phragmocone chambers. Its position is slightly closer to the venter than to the dorsum. The septal neck extrudes about 5 mm in adapertural direction from the septum (at Ah 31.2 mm), and its opening in the septum measures 7.6 and 3.1 mm aperturally and adaperturally, respectively. In transverse section (Fig. 4D), it forms an obtuse angle with the septum ventrally, and a right angle dorsally. Micro-imaging has revealed the partial preservation of the connecting rings of the siphuncle (Fig. 4D).
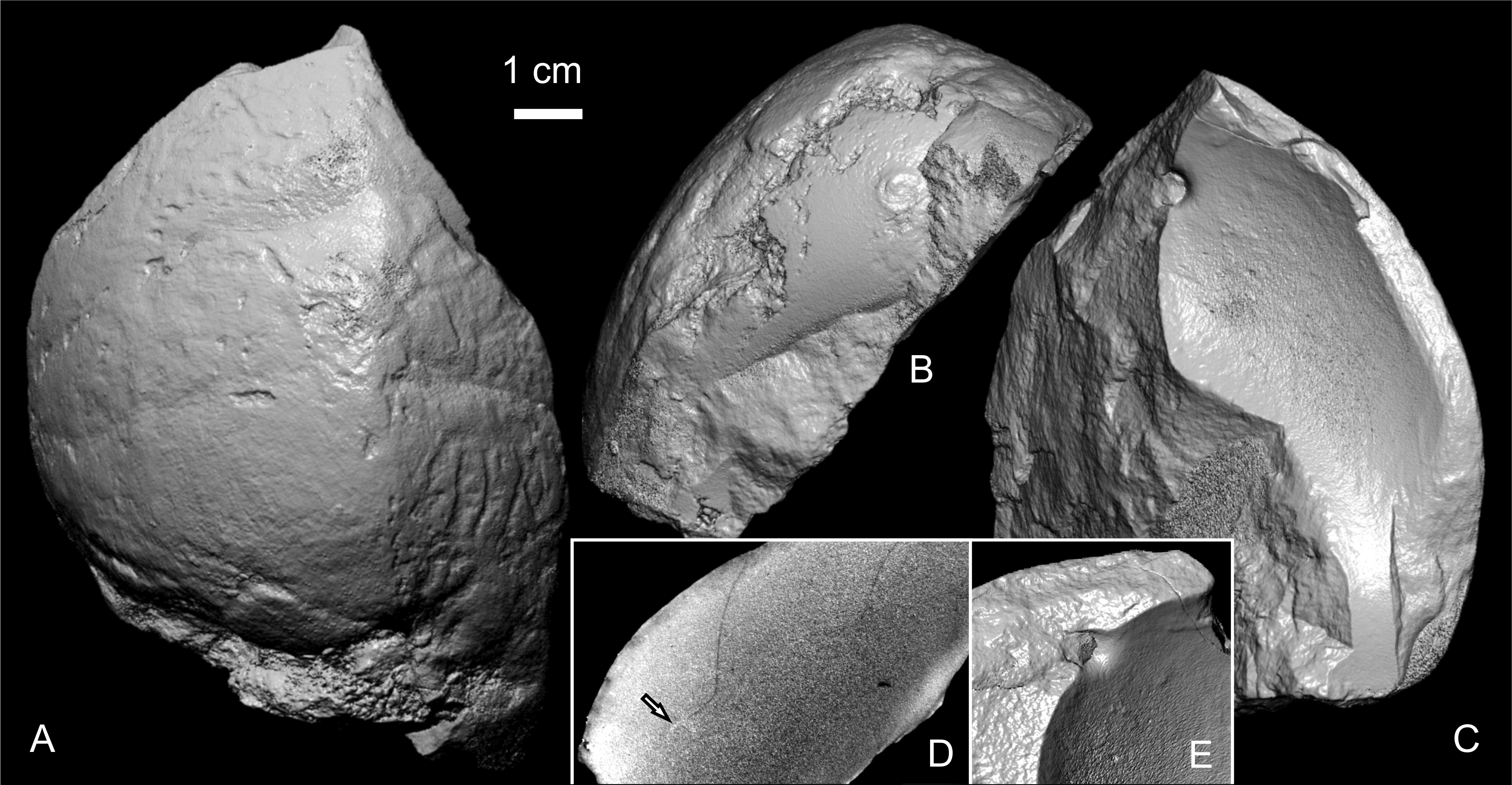
Figure 4. Angulithes westphalicus (Schlüter, 1872) from the Voort Shaft II, Zolder colliery, from a depth of 582.50 m, specimen IRSNB 11508. Screenshots from 3D rendering of micro-CT imaging data (A–C, E), and a virtual cross-section (D), showing the shape of the septal neck, next to some preserved remains (arrow) of the siphuncular tube (probably phosphatised originally organic material of the connecting rings).
36Remarks. Additional nautilid specimens, largely incomplete, from the same lithostratigraphical interval at Zolder colliery, as well as from corresponding depths in the nearby Houthalen Shafts I and II, are present in the RBINS collections. They all remain indeterminate, both generically and specifically, due to their very poor preservation. These specimens also have diameters that are smaller than the lower appearance limit of the angular venter that is typical of Angulithes westphalicus. Some of these have been studied by micro-CT imaging, so as to reveal their siphuncle position. However, these specimens have proved to be imperfectly preserved, to such an extent that the presence of a second taxon (such as Eutrephoceras sp.), co-occurring with A. westphalicus, in these deposits cannot be positively identified.
37The name Nautilus westphalicus is found in faunal lists for the now obsolete Belgian regional stage ‘Hervien’ (Hervian; see Moorkens & Herman, 2006), as provided by Maillieux (1922, p. 118; 1933, p. 153). To our knowledge, these are the only previous mentions of this species in the Belgian Cretaceous. One subadult specimen from the Croix Polinard section at Battice (Fig. 1) in the RBINS collection (former Rutot Collection, general inventory (IG) number 5425) might belong to Angulithes westphalicus, but this needs further study. The same holds true for the entire nautilid fauna from the ‘Hervien’. In this respect, it is interesting to note that Rutot (1875, p. 77) mentioned the presence of a large-sized ‘Nautilus’ at that locality, but failed to provide identifications and illustrations, nor explain what he exactly meant by ‘large-sized’. Such large-sized specimens could hint at the presence of the present species, but the largest specimen we have been able to trace in his collection from Croix Polinard is an adult Eutrephoceras.

Figure 5. A. Angulithes westphalicus (Schlüter, 1872) from the Voort Shaft I, Zolder colliery, from a depth between 579.50 and 582 m, specimen IRSNB 11505. B–D. Well-preserved major chelipeds of Mesostylus faujasi (Desmarest, 1822) from Voort Shaft II at a depth of 598.91 m (B: IRSNB 11509; C: IRSNB 11510) and Voort Shaft I 599 m (D: IRSNB 11511). Photographed under white light (B) after coating with NH4Cl (C–D).
4. Discussion
4.1. Lithostratigraphy and significant change of depositional environment
38According to their labels, the five specimens of Angulithes westphalicus described here originate from a very restricted stratigraphical interval, namely from depths between 579.50 and 584.30 m, part of the ‘Campanien – Smectite de Herve’, at Voort Shafts I and II of the Zolder colliery (Fig. 6).
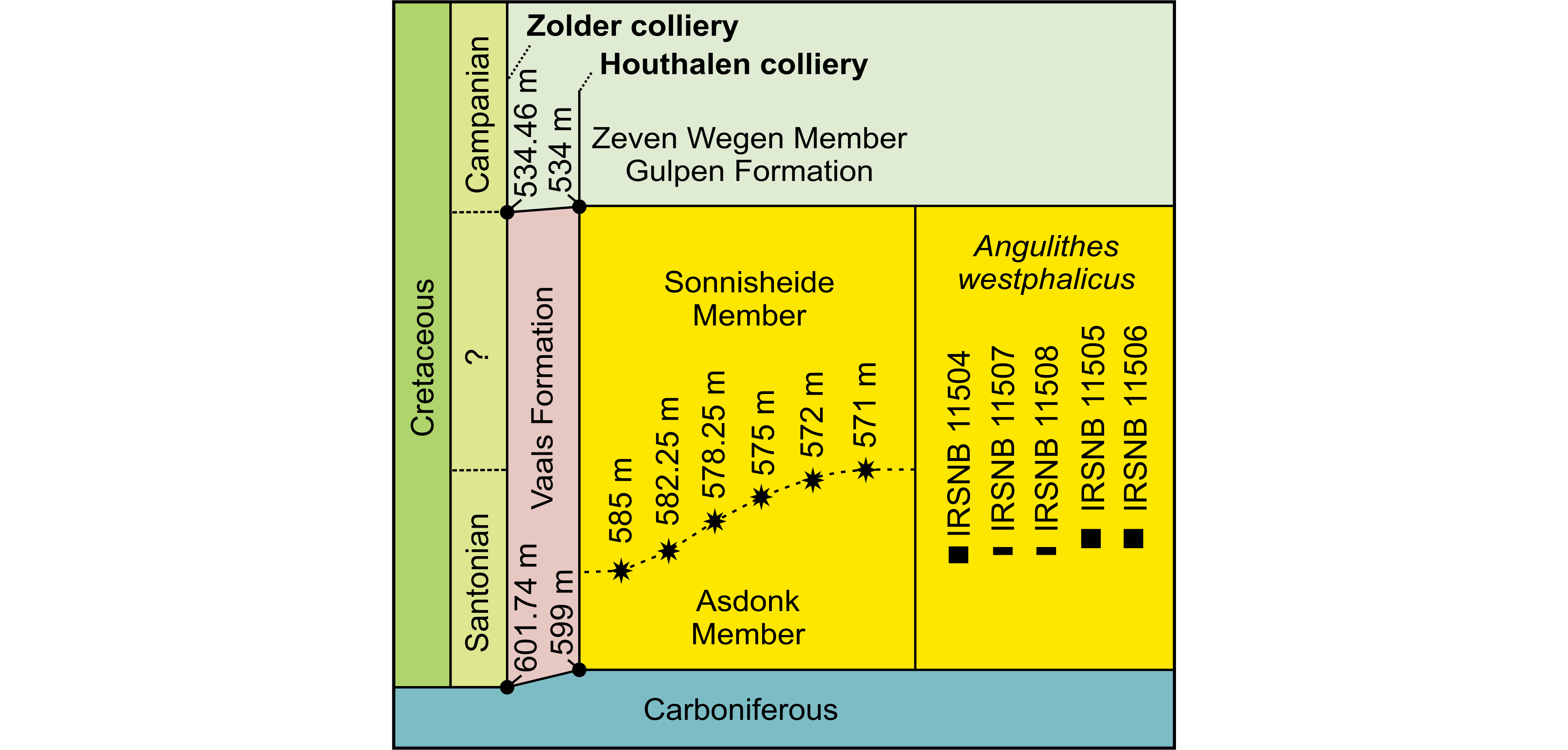
Figure 6. Lithostratigraphy of the Santonian and lower Campanian deposits found at Shafts I & II of the Zolder and Houthalen collieries. The depths of the base and top of the Vaals Formation interval at these shafts are nearly identical so that their fossil occurrences can be cross correlated on a 1:1 scale. The different possible depth levels of the boundary between the Asdonk and Sonnisheide members as discussed in this paper are displayed, as are the depths of the occurrences of our five Angulithes westphalicus (Schlüter, 1872) specimens. The interval downwards from 572 m clearly dates to the Santonian, the age dating of the part above 572 m but below the overlying Zeven Wegen Member (Gulpen Formation) needs further research.
39This ’Smectite de Herve’ is an old and disused term that translates in today’s lithostratigraphy to the Vaals Formation (Robaszynski et al., 2002). At least in its correct usage. This is, while all of the more than 7000 fossils collected from 600 to 400 m depth at Zolder and Houthalen by the RBINS staff in the 1920s and 1930s were labelled as originating from the ‘Campanien – Smectite de Herve’, only those coming from the lower part of this interval actually correlate to the Vaals Formation (see below).
40The most detailed lithological description of the Cretaceous strata encountered at Zolder, and by extension in the Campine coal mining area, was provided by Schmitz & Stainier (1910), when describing an exploratory borehole near the future Voort Shaft II of the Zolder colliery (DOV kb25d62w-B206, GSB 062W0205, KOEN B 79). Here, the top of the Vaals Formation, also the base of the overlying Zeven Wegen Member (Gulpen Formation), is at a depth of 534.46 m; its base, also the contact with the underlying Coal Measures of Westphalian age (Carboniferous), is at 601.74 m (Schmitz & Stainier, 1910). Additional but less detailed information can be found in the description of boreholes near Voort Shaft I of the Zolder colliery (DOV kb25d62w-B228, GSB 062W0226; DOV kb25d62w-B229, GSB 062W0227), and that of boreholes near Shafts I & II of the nearby Houthalen coal mine (DOV kb25d62e-B250, GSB 062E0250; DOV kb25d62e-B251, GSB 062E0251; DOV kb25d62e-B248, GSB 062E0248, KOEN B 95). Remarkably, while these pair of shafts are several kilometres apart, the base of the Cretaceous and the top of the Vaals Formation interval is at nearly identical depth (at 601.74 m and 599–601 m, and, 534.46 m and 534 m, respectively). This is also evidenced by the (other) fossils collected from the ‘Campanien – Smectite de Herve’ interval from these shafts, and their occurrence depths can be cross correlated on a 1:1 scale.
41A twofold subdivision of the 67.3 m-thick Vaals Formation interval can be made. In the lower part, the lithology is predominantly that of glauconite-bearing green sand, sandstone and calcareous sandstone. In the upper part, a glauconite-bearing but grey marl is found with levels in which the ichnofossil Gyrolithes davreuxi de Saporta, 1884 is common.
42A similar twofold division of the Vaals Formation was recognized in the subsurface deposits of the entire Campine area, both in its western and eastern parts (Laenen, 2002; Lagrou et al., 2005; Dusar & Lagrou, 2007). To accommodate this twofold character, two new lithostratigraphical units were introduced: the Asdonk and Sonnisheide members (Dusar & Lagrou, 2011a, b).
43The lower Asdonk Member, as defined by Dusar & Lagrou (2011a), is characterised by green, glauconite-rich calcareous fossiliferous clay and sand, occasionally more lithified by carbonate cement resulting in an alternation between indurated calcareous sandstone and loose sand layers. Its stratotype is at borehole KS 37 (DOV kb17d47w-B264, GSB 047W0267) at Asdonk (Leopoldsburg) (Fig. 1), between a depth of 754–771.5 m, in the eastern part of the Campine. It is said to show some lateral changes, with coarse-grained sand and indurated carbonate banks and a basal gravel in the eastern coal mining district. In the western coal mining district, it consists of green glauconitic clayey silt and fine sand, overlying a clay-rich base, thus with an upward increase of sand and carbonate content. In the Antwerp Campine, this sequence becomes a clayey marl that is reminiscent of the smectite facies in the Hallembaye section (Dusar & Lagrou, 2011, a) (Fig. 1).
44The lithology of the upper Sonnisheide Member is that of a greenish-grey, silty glauconite-bearing marl, often with Gyrolithes and other ichnofossils (Dusar & Lagrou, 2011b). It is said to be rather homogeneous, with an average carbonate content of 50%, remaining sandier in the east and siltier or marlier in the west. Its stratotype is borehole KS 29 (DOV kb25d62e-B284; GSB 062E0280), at Houthalen-Helchteren (Fig. 1), only a few kilometres northeast of the Zolder and Houthalen collieries, between a depth of 568–633 m. In this borehole, the 31 m-thick Cretaceous sediments below the Sonnisheide Member (base at 664 m) were subdivided into the Asdonk Member for the upper 16 m and the Aachen Formation for the lower 15 m, respectively (by Michiel Dusar, in DOV).
45The twofold nature of the Vaals Formation interval at Zolder and Houthalen was interpreted as representing the Asdonk (below) and Sonnisheide (above) members (see DOV for formal lithostratigraphic interpretations by David Lagrou for the boreholes listed above). In the Voort Shaft II borehole described by Schmitz & Stainier (1910), the intra-Vaals Formation boundary is placed at 582.25 m (in DOV, by David Lagrou), but the original description of Schmitz & Stainier (1910) also allows for an alternative at 578.25 m depth. In the other boreholes, the boundary is placed at 578, 575 or 571 m depth (in DOV, by David Lagrou).
46Looking solely at the fossils collected at both shafts by the RBINS staff, the boundary must be above 572 m, given the sandy nature of the matrix of the fossils found at (e.g., of the ammonite Placenticeras polyposis, see also below) and below this level.
47Interestingly in light of this discussion, within the lower sandy interval of the Vaals Formation at Zolder and Houthalen, a significant change in the depositional environment can be seen in a preliminary study of all collected fossils from this interval (Fig. 7). In the lowest levels, pectinids and ostreids largely dominate (about 80%) the bivalve fauna. This predominance stays roughly similarly high up to a depth of 585 m, to drop significantly to 20% (585–580 m) higher upsection, and to 6% (579–576 m) and 2% (575–571 m) even higher. This coincides with an increase in the number of collected belemnites, ammonites and nautilids. Since there are no data on how systematically this material was collected, and that the dual mode of shell preservation (originally calcitic shells preserved versus moulds and imprints for aragonitic ones) may have had some effect on the actual number of collected specimens, this observation must reflect a deepening of the depositional environment, under normal-salinity marine conditions. Additional confirmation for a shallower environment of the lowest part of the sandy interval comes from the abundance of decapod crustacean remains in the lowest 8 m, in which Mesostylus faujasi (Desmarest, 1822) with preserving both major and minor chelipeds in connection are not uncommon (Fig. 5B–D). This deepening is also an interesting observation with respect to the age-diagnostic belemnites. In combination with the observation that these belemnites are all well-preserved adult specimens (fide Jagt et al., 1995), it gives further evidence that these are an autochthonous part of the fauna, and not reworked from older strata (see e.g., Keutgen, 2011 on this topic).
48The depositional change insinuates to Asdonk–Sonnisheide boundary to be placed near 585 m depth. Remarkably, this level would fit best with the thickness of the Asdonk and Sonnisheide members in several boreholes located a few km north of the line connecting the shafts of Zolder and Houthalen. These boreholes (KS 9, KS 11, KS 27, KS 29, KS 34, KS 44 and KS 46) were interpreted by Lagrou et al. (2005, figs 1, 5), mostly on geophysical parameters (gamma ray). Here, the thickness of the members ranges from 13–18.5 m for the Asdonk Member and 44.5–65 m for the Sonnisheide Member.
49Thus, depending on which information is used, the five Angulithes westphalicus position in the top of the Asdonk Member, or in the basal part of the Sonnisheide Member. Lithologically, the matrix of these specimens seems to correspond better with the Asdonk Member than to the Sonnisheide Member, the reason for which we incline to put them in the Asdonk Member, awaiting further studies resolving the issue.
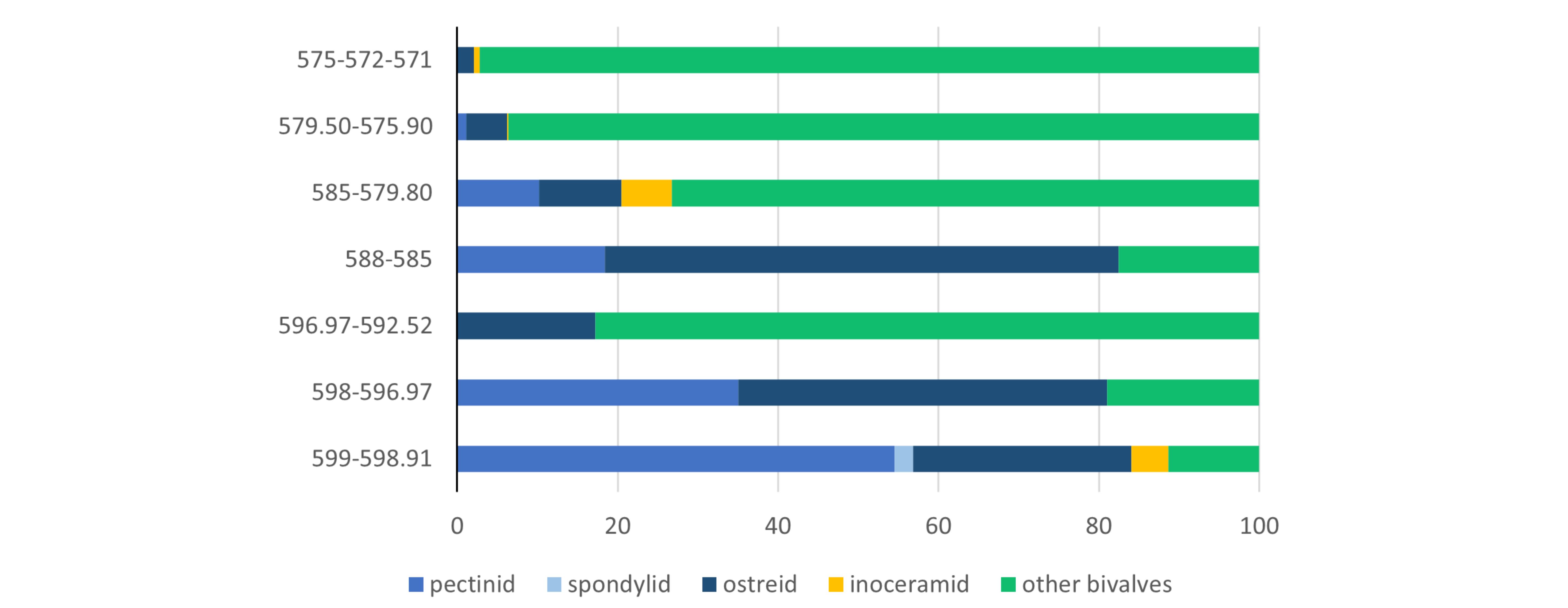
Figure 7. Vertical distribution of bivalve faunas in the lower sandy part of the ‘Smectite de Herve’ interval of Shafts I and II of the Zolder and Houthalen collieries (at depths between c. 600 and 571 m), as expressed in percentages for several ‘taxonomic’ groupings, plotted against depth (N: total number of specimens per level). Based on an inventory of c. 2000 invertebrate fossils in the RBINS palaeontology collection, depth levels taken from associated labels.
4.2. Biostratigraphy and age of the ‘Smectite de Herve’ interval at Zolder and Houthalen
50Traditionally, strata of the Vaals Formation have been dated as ranging from the early early Campanian to the early late Campanian (Robaszynski et al., 2002). However, Jagt et al. (1995), in a study of ammonites, belemnites, inoceramid bivalves and nannofossils collected from the shafts I and II of the Zolder and Houthalen collieries, revealed that the lower (sandy) part at Zolder and Houthalen was of late middle Santonian to possibly late Santonian age, making these beds correlative to the somewhat enigmatic Lonzée Member and (part) of the Aachen Formation (see also Jagt, 1999). Jagt et al.’s (1995) data are discussed below per fossil group.
51In more detail, amongst the ammonites of Jagt et al. (1995), it is especially Placenticeras polyposis that allows substantiating the Santonian age of this interval. Specimens of this species were collected at depths of 583–585, 582–584.30, 581.45 and 572 m (the latter being the seven specimens mistakenly indicated as originating from a depth of 527 m by Jagt et al., 1995). From nearby regions, such as England, France, Germany and Austria, this species is unknown from beds attributed to the lower Campanian (Summersberger et al., 2017b).
52Belemnites have permitted the most detailed age assignment. Fifteen complete specimens, from depths between 588 and 575 m, have been identified as Gonioteuthis westfalicagranulata, the index species of the westfalicagranulata belemnite Zone (Fig. 8), dated as late middle Santonian (Jagt et al., 1995; Keutgen, 2011). Although species and subspecies of the Gonioteuthis lineage are occasionally difficult to identify, it is the large distribution of the Riedel Quotient (see Jagt et al., 1995) that clearly positions these specimens well within the Santonian.
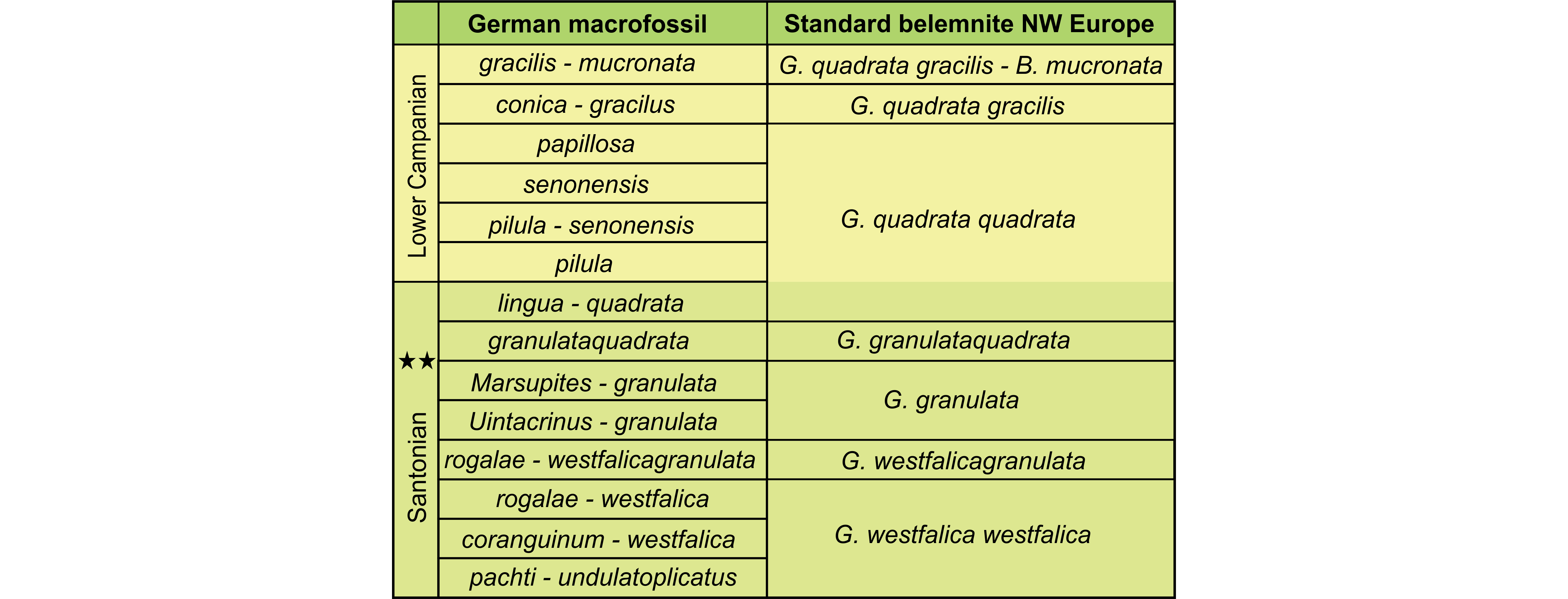
Figure 8. German macrofossil zonation (left) integrated with the standard belemnite zones of Northwest (NW) Europe (right) of the Santonian and Lower Campanian (after Christensen, 1991), with the Santonian–Campanian boundary placed two zones higher than the traditional definition (stars) as a result of the recent GSSP definition of the base of the Campanian in the Bottacione section at Gubbio, Italy, by Gale et al. (2023). B.: Belemnitella, G.: Gonioteuthis.
53Inoceramid bivalves found at a depth of 583–585 m are indicative of the middle to upper Santonian, with middle Santonian being the most probable age assessment (Jagt et al., 1995, with additional material from Walaszczyk & Dhondt, 2005). Another species, Endocostea baltica, of which a specimen was found at a depth of 579.5–582 m, has a known range from the upper Santonian to the Maastrichtian (Jagt et al., 1995). Higher upsection, already in the upper marly part, specimens found at depths of 559–563 and 563–571 m were referred to the Sphenoceramus patootensiformis–angustus group by Walaszczyk & Dhondt (2005), which has, according to the authors, a known range from the upper upper Santonian to the top of the lower lower Campanian.
54For the calcareous nannofossils studied by Jagt et al. (1995), zone CC16 was determined for six of the nine processed samples (from depths of 596.97–598, 585–590, 583–585, 581.45 and 572 m), CC16/C17 for two samples (from depths of 583–585 and 572 m) and CC15/CC16 for one sample (from a depth of 582–584.3 m), using the zonal scheme of Sissingh (1977) in a modified and summarised version of Perch-Nielsen (1985). Translating these biozonal dates into geological time is not straightforward, since diachroneity has been demonstrated for several of the first and last occurrences used to define the zonal boundaries, and basin to basin correlations, even within the Boreal Realm, for the Santonian have proved challenging. However, major advancements have been made in the last decade (see e.g., Thibault et al., 2016; Ovechkina et al., 2020; Miniati et al., 2020; Gale et al., 2021), leading, amongst other things, to the formal definition of a GSSP for the base of the Campanian (Gale et al., 2023), which is in the Bottacione section at Gubbio, Italy, at the 221.53 m level, corresponding to the magnetic polarity reversal from Chron C34n (top of Long Cretaceous Normal Polarity–Chron) to Chron 33r. The reversal, together with the late Santonian event (LSE, previously named Santonian–Campanian Boundary Event, or SCBE) in carbon isotopes and several bio-events (mostly foraminifera and calcareous nannofossils), should allow correlations with other Santonian–Campanian boundary sections worldwide (see Gale et al., 2023 for details). The choice for the polarity reversal implies for some—minor scale—adjustments to the boundary level in some well-studied sections. In the Boreal chalk domain, the lingua–quadrata and granulataquadrata macrofossil zones at Lägerdorf, northern Germany (Schönfeld et al., 1996), traditionally attributed to the lowermost Campanian, are now latest Santonian in age (Fig. 8). The same holds true for the lower part of the Offaster pilula Zone at Seaford Head, United Kingdom. According to the sediment accumulation rate estimates of Marron & Muttoni (2020) for Gubbio, taken together with the proposed detailed correlation of Gubbio, Lägerdorf and Seaford Head of Gale et al. (2023, fig. 4), the base of the Campanian has shifted upwards by ~250 to 300 kyr. While this may not seem much, it is over 10% of the total duration of the Santonian Stage: only 2.1 Myr (from 85.7 ± 0.2 Ma to 83.7 ± 0.5 Ma; see Gale et al. (2020), 2.3 Myr (Thibault et al., 2016, but with the new position of the base of the Campanian of Gale et al. (2023), by astronomical calibration) or 3.3 Myr (from 86.3 ± 0.5 Ma to 83.6 ± 0.2 Ma according to the ICS International Chronostratigraphic Chart version 2023.06, Cohen et al., 2023). The correlations presented by Gale et al. (2023) reveal that, albeit with some minor level diachronism, the observed first occurrence datum of the calcareous nannofossil taxon Aspidolithus parcus parcus (previously Broinsonia parca parca, used to define the base of CC18) falls, in most sections, within the topmost Santonian or lower(most) Campanian. Its absence in all nine samples studied herein is thus in line with the Santonian age of the sandy lower part of the Cretaceous succession at Zolder and Houthalen.
55In summary, all available age estimates for the lower sandy interval (at depths between c. 600 to c. 571 m) at Zolder and Houthalen, and thus also our five specimens of Angulithes westphalicus, point to a Santonian age, with belemnites further precising this to upper middle Santonian (Gonioteuthis westfalicagranulata belemnite zone, sensu germanico, Fig. 8).
4.3. Age of the Asdonk and Sonnisheide members
56The Asdonk Member was said to be of early Campanian age (Dusar & Lagrou, 2011a), and the Sonnisheide of early to late Campanian (Dusar & Lagrou, 2011b), based on dinoflagellate biostratigraphy of Slimani (2000), which is at odds with the Santonian age of the lower sandy interval evidenced above (see also Keutgen, 2011).
57Slimani et al. (2011, p. 130) noted that, “Moreover, particular correlation problems relate to different ages attributed to the same formation. A presumed Santonian age for the base of the Vaals Formation in colliery shafts (Jagt et al., 1995) was seen to be indicative of a Santonian marine transgression in the Campine area (Vandenberghe et al., 2004). However, other fossils generally favour an early Campanian date for the Vaals Formation in the Campine Basin, and geophysical well-log correlation suggests a great regularity in the depositional systems without diachronism.”. Slimani et al. (2011, p. 155) also repeated that, “The Cretaceous transgression over the Campine Basin, and hence the age of the Vaals Formation (interval between 1186–1146 m) [Meer borehole] is early Campanian, not Santonian”.
58Unfortunately, Slimani et al. (2011) did not have dinoflagellate data for sediments from Zolder and Houthalen to support their statement. Even more problematic is the total absence of any treated sample from the basal parts of the Cretaceous sequence anywhere in the Campine. To illustrate this further, we can refer to the lowermost sample documented by Slimani et al. (2011) of the Meer borehole (western part of the Campine; DOV kb8d7e-B224, GSB 007E0205) (Fig. 9) which is from (at 1152 m, said to be indicative of an early Campanian age, and with the lower/upper Campanian boundary tentatively placed between 1152 and 1109 m, the second lowest sample) the top of the Sonnisheide Member (1146–1177 m, see Slimani et al., 2011), and thus 25 m above the top of the Asdonk Member, and 36 m above the base of the Vaals Formation.
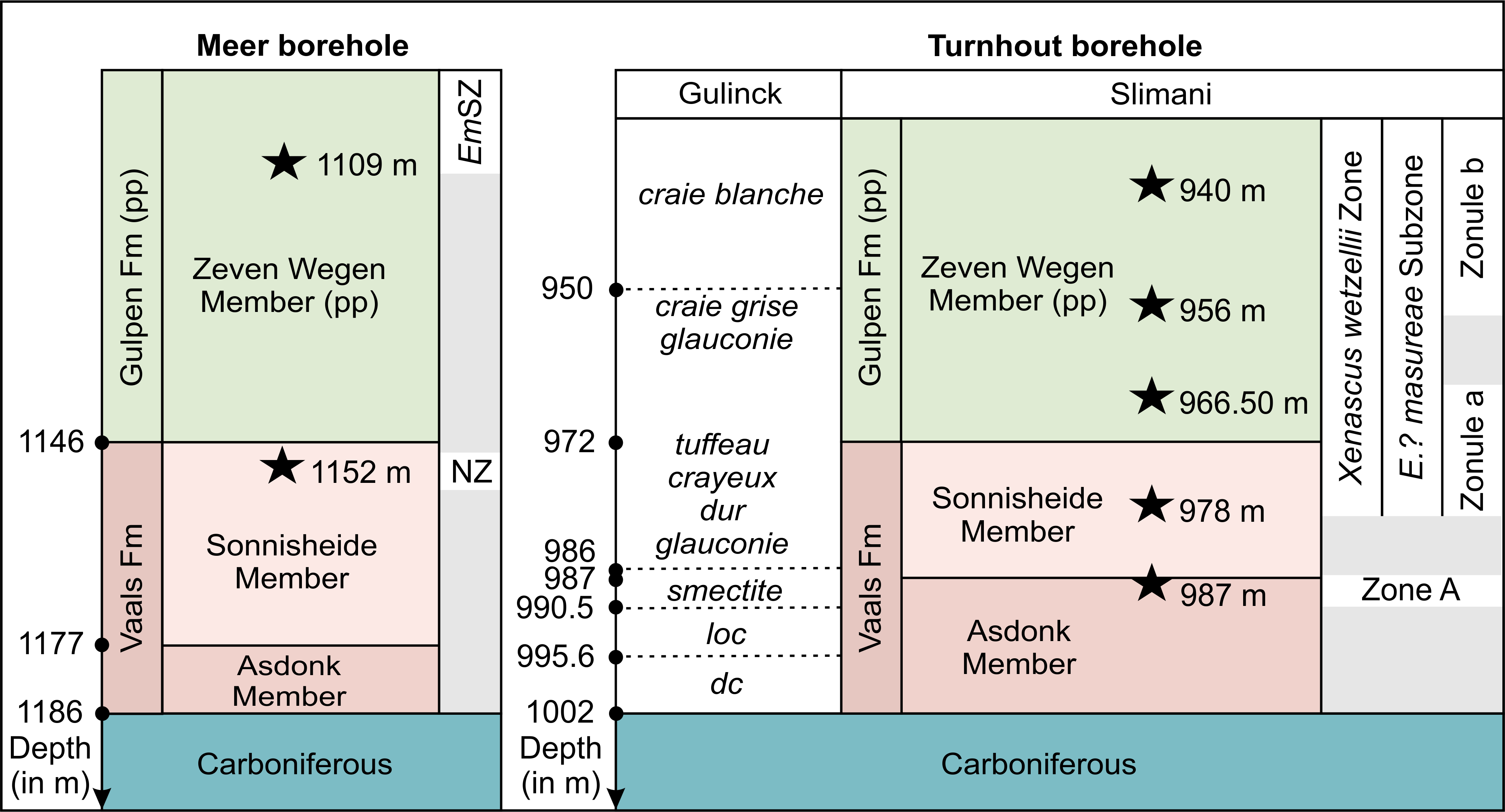
Figure 9. Lithostratigraphy, dinoflagellate samples (stars, with their position in m depth) and dinoflagellate zonation of the oldest Cretaceous deposits at the Meer (left, DOV kb8d7e-B224, GSB 007E0205) and Turnhout (right, DOV kb8d17e-B272, GSB 017E0225-120) boreholes. Abbreviations: dc: destructive coring; EmSZ: Exochosphaeridium? masureae Subzone; Fm: Formation; loc: loss of core; NZ: Not zoned; pp: pro parte. Grey-coloured bars: uncertainty interval. Compiled after Slimani (1994, 2000) (Meer), Slimani et al. (2011) (Meer, Turnhout), and Gulinck (1954).
59In the Turnhout borehole (Turnhout Zwemdok, DOV kb8d17e-B272, GSB 017E0225-120; Gulinck, 1954) (Figs 1, 9), frequently cited by Slimani et al. (2011), the lowest sample (at a depth of 987 m; Slimani, 1994, 2000) is from the top of the ‘smectite’ interval of Gulinck (1954), and dates to the early Campanian (Zone A of Slimani, 2000, the zone below the first appearance of markers of the Xenascus wetzellii dinoflagellate zone (early late Campanian following Slimani, 2000), samples indicative for the latter zone were identified between 978–933 m depth). It is also this sample, the only one, that was used to date the Asdonk Member at the time of its introduction by Dusar & Lagrou (2011a). However, the interval from 972–987 m depth of the Turnhout borehole was said to correspond to the Sonnisheide Member, the interval below to the Asdonk Member, with the top of the underlying Carboniferous at 1002 m depth (Lagrou et al., 2005; Slimani et al., 2011). The lithology of Slimani’s (1994, 2000) sample, as well as the lithological description of Gulinck (1954) from the depth of 986 m downwards (‘a homogenous grey marl with imprints of pyritized fossils, foraminifera and a possible Gonioteuthis quadrata belemnite’) are also more reminiscent to that of the Sonnisheide Member than to that of the Asdonk Member. Unfortunately, no lithological data are available for the interval below 990.5 m, due to the loss of cores (up to 995.6 m) and destructive coring (up to the top of the Coal Measures) (see Gulinck, 1954), complicating the stratigraphic interpretation of the Vaals Formation sediments in this borehole.
60In the outcrop area near Maastricht, the lowermost part of the Vaals Formation at Haccourt (Hallembaye) remains largely unstudied for age dating, but according to Jan P.M.Th. Meessen (cited by Schwarzhans & Jagt, 2022), benthic foraminifera suggested that only correlatives to the lower part of the Vaals Formation in its (Dutch) type area were preserved in the smectite facies. At Beutenaken (Fig. 1), only the top part of the section is well dated. At Croix Polinard (Fig. 1), age-diagnostic fossils (mostly coleoids and ammonoids) were only found in the topmost 4–6 m of the Vaals Formation, while the section still down dips for about 20 m (fide Felder, 1975, see e.g., Collins et al., 2000; Keutgen, 2011). Currently, it remains difficult to impossible to correlate deposits of the Belgian Vaals Formation with the detailed lithostratigraphy established in its Dutch type area (Vaals, near Aachen), where six members were defined, namely, from bottom to top, the Raren, Cottessen, Gemmenich, Vaalsbroek, Beusdal and Terstraten members. Belemnites and other fossils allow dating parts of the Cottessen and Gemmenich members as age correlative to the German lingua–quadrata and granulataquadrata zones (see Schwarzhans & Jagt, 2022 for a more elaborate overview). Considering the new GSSP for the base of the Campanian, these are now latest Santonian instead of early Campanian in age. Belemnites (see Christensen & Schmid, 1987; Schwarzhans & Jagt, 2022) allow to correlate the top part of the smectite facies at Hallembaye (former CPL SA quarry) to the German lingua–quadrata Zone; thus, this level should now also be included in the uppermost Santonian. The same must then also be true for (at least part of) Slimani’s (2000) Zone A, which was identified within the top of the Vaals Formation at Hallembaye.
61To conclude, there is no evidence to refute the Santonian age of the oldest deposits at the Zolder and Houthalen colliery shafts, as revealed by Jagt et al. (1995). Even more, there is no dating of the most basal part of the Vaals Formation anywhere in the Campine, and thus, there is currently no evidence either to refute a Santonian flooding of the Brabant Massif as envisioned by Slimani et al. (2011) and Dusar & Lagrou (2007). More research is needed, as several uncertainties remain, regarding the age of the Vaals Formation deposits in the Campine area. In this, a further study of the sediments and fossils from Shafts I & II of the Zolder and Houthalen collieries would seem very helpful.
4.4. Evolutionary picture
62It is the combination of the oval nautilicone shape, the transition from a rounded into an angular venter, the shape of the suture and the absence of ribbing that allow to assign the present five specimens from the Belgian Campine to Angulithes westphalicus, rather than to A. triangularis de Montfort, 1808 (upper lower to middle Cenomanian), A. galea (upper Turonian to the middle Coniacian), Deltocymatoceras leiotropis (Schlüter, 1876) (upper Turonian to middle Coniacian) or D. rugatum (Fritsch & Schlönbach, 1872) (lower upper Turonian). In addition, the placement in A. westphalicus fits best with the known stratigraphical range of that species.
63Interestingly, the position of the siphuncle was recorded as unknown for A. westphalicus (Frank, 2010), hampering a good understanding of the relationship between A. galea and A. westphalicus. This can now be resolved based on the rather ventral position of the siphuncle; it is confirmed that A. westphalicus is very closely related to A. galea, and most probably formed part of a single evolutionary lineage in which, early in the Santonian, the ribbing at the end of the body chamber of adult specimens became lost and the shell proportions modified slightly. In the light of this transition, the shell shape of the Belgian specimens still recalls somewhat A. galea in having slightly more convex flanks than the types of A. westphalicus.
64The possible relationships of Deltocymatoceras and its two constituents (D. rugatum and D. leiotropis) with Angulithes were discussed at length by Frank (2010) and Frank et al. (2013), who concluded that Deltacymatoceras (probably) descended from Cymatoceras rather than from Angulithes.
65Regardless of this discussion, it seems that successive attempts during the Late Cretaceous to combine a keeled venter with globular involute shells were not very productive, in view of their restricted geographical and temporal ranges, e.g., less than 3.5 Myr for Deltocymatoceras. Such morphologies are rare in the nautilid fossil record and appear unexplored since the Triassic. In addition, with doubts on factors responsible for this evolutionary test, this conspicuous morphology also disappears well before the end of the Cretaceous, with the youngest recorded A. westphalicus dated as late Campanian.
5. Conclusions
66Angulithes westphalicus, a Late Cretaceous nautilid with a globular involute smooth shell and remarkably keeled venter, is recorded on the basis of five specimens originating from a very restricted stratigraphical interval, from depths between 579.50 and 584.30 m in the Voort Shafts I and II of the Zolder colliery (Campine). This is the first formal description of the genus and species for the Cretaceous of the Campine area, and the Belgian Cretaceous as a whole, extending also the known geographical occurrence of this species westwards. The material comes from the upper part of a sequence of marine fossiliferous calcareous sand, with a marly glauconiferous base, for which we here also document a deepening of the environment based on an inventory of all collected specimens. The specimens are of late middle Santonian (Gonioteuthis westfalicagranulata belemnite Zone, sensu germanico) age. Lithostratigraphically, this level is within the Vaals Formation, within the upper part of the Asdonk Member or alternatively within the lower part of the Sonnisheide Member. Regardless of this uncertainty, the early Campanian age of the Asdonk Member that was (erroneously) suggested at the time of its introduction can be refuted. In addition, the age of the Sonnisheide Member needs further study, since the major parts of the Vaals Formation now date to the late Santonian.
67Siphuncle position is here identified for the first time in A. westphalicus; it confirms the close evolutionary relationship with A. galea, which is known from the upper Turonian to middle Coniacian of central Europe, and considered its immediate precursor. Both taxa can be differentiated on a few minor modifications of shell shape, ornament and septa. Presently, no other nautilid taxa can be positively identified flanking A. westphalicus.
Acknowledgements
68The present paper results from the Belgian Science Policy Office (Belspo)’s BRAIN2.0 project B2/191/P2/CT-CEPH; the authors gratefully acknowledge the Belgian Federal Government for funding. Micro-CT imaging of specimens was executed within the scope of DiSSCo-Fed, a Belspo-funded RBINS collection digitization project, led by Patrick Semal (2018–2022). Object Research Systems (ORS; Montreal, Canada, https://www.theobjects.com/dragonfly/index.html) and GOM (https://www.gom.com/) are thanked for allowing the use of their Dragonfly ORS (Version 4.1.0.647 for Windows) and GOM Inspect (Version 2019) software, respectively. Alain Drèze, Julien Lalanne and Annelise Folie (RBINS) are thanked for granting access to the RBINS palaeontology collections, database and virtual collections platform, Patrick De Saegher (RBINS) for aiding in the construction of a specimen-based inventory of fossils from the ‘Smectite de Herve’ of the Campine (2012–2014) and Wilfried Miseur (formerly at the RBINS) for conventional photography (in particular, Figures 3E–F and 5B–D). Reviewers Michiel Dusar (RBINS), John W.M. Jagt (Natuurhistorisch Museum Maastricht, The Netherlands) and Marcus Wilmsen (Senckenberg Naturhistorische Sammlungen Dresden, Germany) are thanked for their helpful comments allowing to improve the quality of the manuscript.
Author contributions
69The first author identified the Angulithes westphalicus specimens within the collection of >7000 fossils from the Campine collieries, inventoried this collection (with the aid of Patrick De Saegher, see acknowledgements), executed the micro-CT imaging, made the measurements, table and figures, and wrote most of the largest part of the text. Second author wrote remaining part of the text.
Data availability
70The figured specimens are housed in an official repository, the RBINS, guaranteeing their long-term safekeeping and availability to other researchers for future studies. The micro-CT imaging datasets can be obtained upon request from the RBINS palaeontology collections manager.
References
71Agassiz, L., 1842–1847. Nomenclator zoologicus, continens nomina systematica generum animalium tam viventium quam fossilium, secundum ordinem alphabeticum disposita, adjectis auctoribus, libris, in quibus reperiuntur, anno editionis, etymologia et familiis, ad quas pertinent, in singulis classibus. Jent et Gassmann, Soloduri, 393 p. https://doi.org/10.5962/bhl.title.49761
72Christensen, W.K., 1991. Belemnites from the Coniacian to Lower Campanian chalks of Norfolk and southern England. Palaeontology, 34/3, 695–749.
73Christensen, W.K. & Schmid, F., 1987. The belemnites of the Vaals Formation from the C.P.L. quarry at Hallembaye in Belgium: taxonomy, biometry and biostratigraphy. Geologisches Jahrbuch, A94, 3–37.
74Cohen, K.M., Finney, S.C., Gibbard, P.L. & Fan, J.-X., 2013 (updated, version 2023-06). The ICS International Chronostratigraphic Chart. Episodes, 36, 199–204. https://stratigraphy.org/ICSchart/ChronostratChart2023-06.pdf, accessed 20/08/2023.
75Collins, J.S.H., Fraaye, R.H.B. & Jagt, J.W.M., 2000. First record of the homolid crab Hoplitocarcinus gibbosus (Schlüter, 1879) from the Lower Campanian of Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 70, 121–127.
76de Montfort, D., 1808. Conchyliologie systématique et classification méthodique des coquilles ; offrant leurs figures, leur arrangement générique, leurs descriptions caractéristiques, leurs noms ; ainsi que leur synonymie en plusieurs langues. Coquilles univalves, cloisonnées. Tome premier. F. Schœll, Paris, 409 p. https://doi.org/10.5962/bhl.title.10571
77de Saporta, G., 1884. Les organismes problématiques des anciennes mers. G. Masson, Paris, 102 p.
78Desmarest, A.-G., 1822. Les Crustacés proprement dits. In Brongniart, A. & Desmarest, A.-G. (eds), Histoire naturelle des Crustacés fossiles sous les rapports zoologiques et géologiques. F.-G. Levrault, Paris, 67–154. https://doi.org/10.5962/bhl.title.66799
79Dusar, M. & Lagrou, D., 2007. Cretaceous flooding of the Brabant Massif and the lithostratigraphic characteristics of its chalk cover in northern Belgium. Geologica Belgica, 10/1-2, 27–38.
80Dusar, M. & Lagrou, D., 2011a. Asdonk Member. National Commission for Stratigraphy Belgium. https://ncs.naturalsciences.be/cretaceous/asdonk-member, accessed 09/01/2023.
81Dusar, M. & Lagrou, D., 2011b. Sonnisheide Member. National Commission for Stratigraphy Belgium. https://ncs.naturalsciences.be/cretaceous/sonnisheide-member, accessed 09/01/2023.
82Felder, W.M., 1975. Lithostratigraphische Gliederung der Oberen Kreide in Süd-Limburg (Niederlande) und den Nachbargebieten. Erster Teil: Der Raum westlich der Maas, Typusgebiet des “Maastricht”. Publicaties van het Natuurhistorisch Genootschap in Limburg, 24, 1–43.
83Fözy, I., 2001. Campanian (Late Cretaceous) cephalopods from Sümeg (Transdanubian Central Range, Hungary). Fragmenta Palaeontologica Hungarica, 19, 25–37.
84Frank, J., 2010. Taxonomy and palaeoecology of Cretaceous nautilids Angulithes galea (Fritsch in Fritsch & Schlönbach, 1872) and Angulithes westphalicus (Schlüter, 1872). Bulletin of Geosciences, 85/3, 487–496. https://doi.org/10.3140/bull.geosci.1194
85Frank, J., Wilmsen, M. & Košťák, M., 2013. The endemic and morphologically remarkable nautilid genus Deltocymatoceras Kummel, 1956 from the Late Cretaceous of Central Europe. Bulletin of Geosciences, 88/4, 793–812. https://doi.org/10.3140/bull.geosci.1402
86Fritsch, A. & Schlönbach, U., 1872. Cephalopoden der böhmischen Kreideformation. Prag, privately published, 52 p. https://doi.org/10.5962/bhl.title.46960
87Gale, A.S., Mutterlose, J., Batenburg, S., Gradstein, F.M., Agterberg, F.P., Ogg, J.G. & Petrizzo, M.R., 2020. The Cretaceous Period. In Gradstein, F.M., Ogg, J.G., Schmitz, M.D. & Ogg, G.M. (eds), Geologic Time Scale 2020, Volume 2. Elsevier, Amsterdam, 1023–1086. https://doi.org/10.1016/B978-0-12-824360-2.00027-9
88Gale, A., Kennedy, J. & Walaszczyk, I., 2021. Correlation of the late Santonian–early Campanian of Texas, USA with the Anglo-Paris Basin and other regions. Newsletters on Stratigraphy, 54/4, 433–460. https://doi.org/10.1127/nos/2020/0641
89Gale, A., Batenburg, S., Coccioni, R., Dubicka, Z., Erba, E., Falzoni, F., Haggart, J., Hasegawa, T., Ifrim, C., Jarvis, I., Jenkyns, H., Jurowska, A., Kennedy, J., Maron, M., Muttoni, G., Pearce, M., Petrizzo, M.R., Premoli-Silva, I., Thibault, N., Voigt, S., Wagreich, M. & Walaszczyk, I., 2023. The Global boundary Stratotype Section and Point (GSSP) of the Campanian Stage at Bottaccione (Gubbio, Italy) and its auxiliary sections: Seaford Head (UK), Bochieniec (Poland), Postalm (Austria), Smoky Hill, Kansas (U.S.A), Tepayac (Mexico). Episodes, 46, in press. https://doi.org/10.18814/epiiugs/2022/022048
90Goolaerts, S., De Saegher, P., Dusar, M., Fraaije, R.H.B., Jagt, J.W.M., Kennedy, W.J., Marquet, R., Simon, E., Van Bakel, B. & Van Knippenberg, P., 2013. The CRETACAM project: a new look at Santonian to Maastrichtian deposits of the Belgian Campine basin. 9th International Symposium on the Cretaceous System, Ankara, Turkey, 1–5/09/2013. Abstract book, 119.
91Gulinck, M., 1954. Coupe résumée des terrains post-paléozoïques au sondage de Turnhout. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 63, 147–154.
92Jagt, J.W.M., 1999. Late Cretaceous-Early Palaeogene echinoderms and the K/T boundary in the southeast Netherlands and northeast Belgium - Part 1: Introduction and stratigraphy. Scripta Geologica, 116, 1–57.
93Jagt, J.W.M., Kennedy, W.J., Burnett, J.A., Christensen, W.K. & Dhondt, A.V., 1995. Santonian macrofauna and nannofossils from northeast Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 65, 127–137.
94Janiszewska, K., Mazur, M., Machalski, M. & Stolarski, J., 2018. From pristine aragonite to blocky calcite: exceptional preservation and diagenesis of cephalopod nacre in porous Cretaceous limestones. PLoS ONE 13/12, e0208598. https://doi.org/10.1371/journal.pone.0208598
95Keutgen, N., 2011. The belemnite zonation of the uppermost Cretaceous in the Maastricht-Aachen-Liège, Brabant-Méhaigne and Mons areas (Belgium, southeast Netherlands). In Jagt, J.W.M., Jagt-Yazykova, E.A. & Schins, W.J.H. (eds), A tribute to the late Felder brothers – pioneers of Limburg geology and prehistoric archaeology. Netherlands Journal of Geosciences/Geologie en Mijnbouw, 90/2–3, 165–178. https://doi.org/10.1017/S0016774600001086
96Kummel, B., 1956. Post-Triassic nautiloid genera. Bulletin of the Museum of Comparative Zoology, 114/7, 324–494.
97Laenen, B., 2002. Lithostratigrafie van het pre-Tertiair in Vlaanderen. Studie uitgevoerd i.o.v. Vlaamse Overheid, LNE-ALBON. VITO-rapport 2002/ETE/R/063, 35 p.
98Lagrou, D., Dreesen R. & Dusar, M., 2005. Kartering en karakterisering (sedimentpetrografisch en petrofysisch) van de krijtgesteenten in Vlaanderen. Onderzoek i.o.v. Vlaamse Overheid LNE-ALBON, VITO-rapport 2005/MAT/R/0256, 104 p.
99Maillieux, E., 1922. Terrains, roches et fossiles de la Belgique. Les Naturalistes belges, Bruxelles, 164 p.
100Maillieux, E., 1933. Terrains, roches et fossiles de la Belgique. Deuxième édition revue et corrigée. Patrimoine du Musée royal d’Histoire naturelle de Belgique, Bruxelles, 217 p.
101Marron, M.& Muttoni, G., 2020. A detailed record of the C34n/C33r magnetozone boundary for the definition of the base of the Campanian Stage at the Bottaccione section (Gubbio, Italy). Newsletters on Stratigraphy, 54/1, 107–122. https://doi.org/10.1127/nos/2020/0607
102Miniati, F., Petrizzo, M.R., Falzoni, F. & Erba, E., 2020. Calcareous plankton biostratigraphy of the Santonian-Campanian boundary interval in the Bottaccione Section (Umbria-Marche Basin, Central Italy). Rivisita Italiana di Paleontologia e Stratigrafia, 126/3, 771–789. https://doi.org/10.13130/2039-4942/14399
103Mironenko, A.A., Jagt, J.W.M. & Jagt-Yazykova, E.A., 2022. An unusual conchorhynch from the upper Maastrichtian of the southeast Netherlands and the distinction between nautiloid and ammonoid conchorhynchs (Mollusca, Cephalopoda). Cretaceous Research, 130, 105037. https://doi.org/10.1016/j.cretres.2021.105037
104Moorkens, T. & Herman, J., 2006. Disused Cretaceous regional stage from Belgium: Hervian. Geologica Belgica, 9/1-2, 201–202.
105Müller, G. & Wollemann, A., 1906. Die Molluskenfauna des Untersenons von Braunschweig und Ilsede. II. Cephalopoden. Abhandlungen der königlich preussischen Geologischen Landesanstalt, Neue Folge, 47, 1–30.
106Ovechkina, M.N., Kopaevich, L.F., Aleksandrova, G.N., Proshina, P.A., Ryabov, I.A., Baraboshkin, E.Y., Guzhikov, A.Y. & Mostovski, M.B., 2020. Calcareous nannofossils and other proxies define the Santonian–Campanian boundary in the Central Crimean Mountains (Alan-Kyr section). Cretaceous Research, 119, 104706. https://doi.org/10.1016/j.cretres.2020.104706
107Perch-Nielsen, K., 1985. Mesozoic calcareous nannofossils. In Bolli, H.M., Saunders, J.B. & Perch-Nielsen, K. (eds), Plankton Stratigraphy. Cambridge University Press, Cambridge (UK), 329–426.
108Riegraf, W. & Scheer, U., 1991. Clemens August Schlüter: Cephalopoden der oberen deutschen Kreide. Reprint des in drie Teilen von 1867-1876 erschienenen Werkes, nomenklatorisch überarbeitet und ergänzt. Goldschneck-Verlag, Korb, 454 p.
109Robaszynski, F., Dhondt, A. & Jagt, J.W.M., 2002. Cretaceous lithostratigraphic units (Belgium). In Bultynck, P. & Dejonghe, L. (eds), Guide to a revised lithostratigraphic scale of Belgium. Geologica Belgica, 4/1-2 (for 2001), 121–134. https://doi.org/10.20341/gb.2014.049
110Rutot, A., 1875. Note sur le gisement de fossiles herviens de la Croix Polinard, près Battice. Annales de la Société géologique de Belgique, 2, Bulletin, 75–78.
111Schlüter, C., 1871–1876. Cephalopoden der oberen deutschen Kreide. Palaeontographica, 21, 1–24 (1871); 21, 25–120 (1872); 24, i–x, 123–204, 207–264 (1876).
112Schmitz, G. & Stainier, X., 1910. Sondage N°79 (Voort, n°2). Annales des Mines de Belgique, 15, 1838–1867.
113Schönfeld, J., Schulz, M.-G., McArthur, J.M., Burnett, J., Gale, A., Hambach, U., Hansen, H.J., Kennedy, W.J., Rasmussen, K.L., Thirlwall, M.F. & Wray, D.S., 1996. New results on biostratigraphy, palaeomagnetism, geochemistry and correlation from the standard section for the Upper Cretaceous white chalk of northern Germany (Lägerdorf-Kronsmoor-Hemmoor). In Spaeth, C. (ed.), New Developments in Cretaceous Research Topics. Proceedings of the Fourth International Cretaceous Symposium, Hamburg 1992. Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg, 77 (Jost Wiedmann Memorial Volume), 545–575.
114Schwarzhans, W. & Jagt, J.W.M., 2022. Silicified bony fish otoliths from the Vaals Formation (lower Campanian) of Vaals-Eschberg, the Netherlands. Cretaceous Research, 139, 105312. https://doi.org/10.1016/j.cretres.2022.105312
115Shimansky, V.N., 1975. Cretaceous nautiloids. Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR, 150, 1–208. [In Russian].
116Sissingh, W., 1977. Biostratigraphy of Cretaceous calcareous nannoplankton. Geologie en Mijnbouw, 56, 37–65.
117Slimani, H., 1994. Les dinokystes des craies du Campanien au Danien à Hallembaye, Turnhout (Belgique) et à Beutenaken (Pays-Bas). Mémoires pour servir à l’Explication des Cartes géologiques et minières de la Belgique, 37, 1–173.
118Slimani, H., 2000. Nouvelle zonation aux kystes de dinoflagellés du Campanien au Danien dans le nord et l’est de la Belgique et dans le sud-est des Pays Bas. Memoirs of the Geological Survey of Belgium, 46, 1–87.
119Slimani, H., Louwye, S., Dusar, M. & Lagrou, D., 2011. Connecting the Chalk Group of the Campine Basin to the dinoflagellate cyst biostratigraphy of the Campanian to Danian in borehole Meer (northern Belgium). In Jagt, J.W.M., Jagt-Yazykova, E.A. & Schins, W.J.H. (eds), A tribute to the late Felder brothers – pioneers of Limburg geology and prehistoric archaeology. Netherlands Journal of Geosciences/Geologie en Mijnbouw, 90/2–3, 129–164. https://doi.org/10.1017/S0016774600001074
120Summesberger, H., Kennedy, W.J. & Skoumal, P., 2017a. Early and middle Santonian cephalopods from the Gosau Group (Upper Cretaceous, Austria). 1. Nautiloidea and non-heteromorph Ammonoidea. Abhandlungen der Geologischen Bundesanstalt, 71, 5–99.
121Summesberger, H., Kennedy, W.J. & Skoumal, P., 2017b. Early and middle Santonian cephalopods from the Gosau Group (Upper Cretaceous, Austria). 2. Heteromorph Ammonoidea. Abhandlungen der Geologischen Bundesanstalt, 71, 101–149.
122Thibault, N., Jarvis, I., Voigt, S., Gale, A.S., Attree, K. & Jenkyns, H.C., 2016. Astronomical calibration and global correlation of the Santonian (Cretaceous) based on the marine carbon isotope record. Paleoceanography, 31, 847–865. https://doi.org/10.1002/2016PA002941
123Vandenberghe, N., Van Simaeys, S., Steurbaut, E., Jagt, J.W.M. & Felder, P.J., 2004. Stratigraphic architecture of the Upper Cretaceous and Cenozoic along the southern border of the North Sea Basin in Belgium. Netherlands Journal of Geoscience/Geologie en Mijnbouw, 83, 155–171. https://doi.org/10.1017/S0016774600020229
124Walaszczyk, I. & Dhondt, A.V., 2005. Santonian-Campanian (Upper Cretaceous) inoceramids from the Houthalen mineshaft, NE Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 75, 167–181.
125Wiedmann, J., 1960. Zur Systematik jungmesozoischer Nautiliden. Palaeontographica, Abteilung A, 115, 144–206.
126Wilmsen, M., 2000. Late Cretaceous nautilids from northern Cantabria, Spain. Acta Geologica Polonica, 50/1, 29–43.
127Wittler, F.A., Roth, R. & Legant, J., 1999. Die Nautiliden der oberen Kreide (Cenoman-Campan) vom Süd- und Westrand des Münsterländer Beckens. Arbeitskreis Paläontologie Hannover, 27, 1–52.
128Manuscript received 02.03.2023, accepted in revised form 22.08.2023, available online 07.09.2023.
