


- Startpagina tijdschrift
- Volume 27 (2024)
- number 1-2
- The palaeoecological conditions of the tidal channel network of the Zwin region (Flanders, Belgium)


Weergave(s): 1175 (32 ULiège)
Download(s): 824 (26 ULiège)

The palaeoecological conditions of the tidal channel network of the Zwin region (Flanders, Belgium)

Abstract
The northeastern coastal plain of Belgium was a well-developed region during the late medieval period when Bruges and a series of smaller harbours became connected to the North Sea via the Zwin tidal channel. Yet, ecological data related to the evolution of the regional environment of the former Zwin region are still lacking. Diatoms, pollen, and macrobotanical remains have been studied for the first time from two cores drilled in the surroundings of the lost harbour of Hoeke. The sediments in both cores were deposited in four successive tidal channel systems spanning the pre-Roman to late medieval periods. Marine tychoplanktonic and planktonic diatoms were dominant and confirmed the presence of open marine tidal channels, while marine epipsammic diatoms were specific to channel deposits (systems 1 and 4) and marine epipelic diatoms characterised channel and accretion deposits (systems 2 and 3). The sediments of the late medieval Zwin main channel and tributary (system 4) are both characterised by marine tychoplanktonic and epipsammic species. The palynomorphs were typical of marine tidal deposits and (reworked) peat, while macroremains pointed to freshwater plants growing in the vicinity. The palynomorphs (system 4) suggest a more open landscape associated with the extensive reclamation and embankment of the Zwin region. Diatoms in the superjacent tidal deposits indicate that the late medieval Zwin silted up rapidly.
Inhoudstafel
1. Introduction
1The landscape of the eastern Belgian coastal plain was transformed when, according to historical sources, a large tidal channel developed following a storm surge in 1134 (Verhulst, 1959; Trachet et al., 2015; De Clercq et al., 2021). The tidal channel, called the Zwin, ensured for the entire region access to the sea. Despite its location further inland, the Zwin allowed the city of Bruges (Fig. 1) to grow into an important medieval metropolis as it became strategically located at the convergence of the newly established maritime gateway and the traditional continental trade routes. However, maritime commerce from Bruges via the North Sea was only viable through a network of harbour towns that emerged on the banks of the Zwin tidal channel, one of which was Hoeke (Dumolyn & Leloup, 2016; Trachet, 2016; Fig. 1).
2Traditionally, the overall development of the medieval Zwin landscape was reconstructed from historical sources (e.g. Verhulst, 1959; Pannier, 1970). In recent years however, the region has received new academic interest, notably from archaeological studies carried out at lost harbour sites, such as Monnikerede and Hoeke (Trachet, 2016). Despite this growing scholarly attention, geological and ecological data remained limited for the area. A recent research project provided a synthesis of both legacy and newly collected data specific to Holocene peatlands and archaeological findings in the eastern coastal plain of Belgium (Allemeersch et al., 2023). Furthermore, comprehensive geophysical, geological, and archaeological research in 2021 at Hoeke presented a unique opportunity to better understand the dynamics that shaped the late medieval Zwin tidal channel and its possible predecessors (De Clercq et al., 2022). In the shallow subsoil of Hoeke and its surroundings, the remains of a complex network of chronologically successive tidal channel systems appeared to be well preserved, covering a period from the pre-Roman period until the Late Middle Ages, the period in which the Zwin became the gateway of Bruges. The Quaternary evolution of these sedimentary environments has recently been described by sedimentological analyses of cores taken at Hoeke (Bogemans & Heyvaert, 2023; Bogemans et al., 2024).
3This paper aims to advance our understanding of the regional landscape evolution by investigating the ecological conditions in the different tidal channel systems of the Zwin region. Diatom and palynological assemblages were analysed throughout the sedimentary deposits from two undisturbed mechanically drilled cores (cores B3 and B5), while preserved macrobotanical remains were also investigated at specific levels in core B5. The primary objective was to assess whether diatom assemblages could effectively distinguish between the sedimentary subenvironments described by Bogemans & Heyvaert (2023) and thus follow the dynamics of the tidal channel systems and tidal flat over time. Additionally, we sought to investigate potential differences in environmental conditions during activity of the different types of channels with a focus on a main tidal channel and a tributary channel, to gain insights into the later phases of the Zwin’s evolution. Finally, palynological and macrobotanical analyses were carried out to determine whether distinct sources of material could be detected within the various sedimentary deposits, thereby contributing to our understanding of the overall development of this historical landscape.
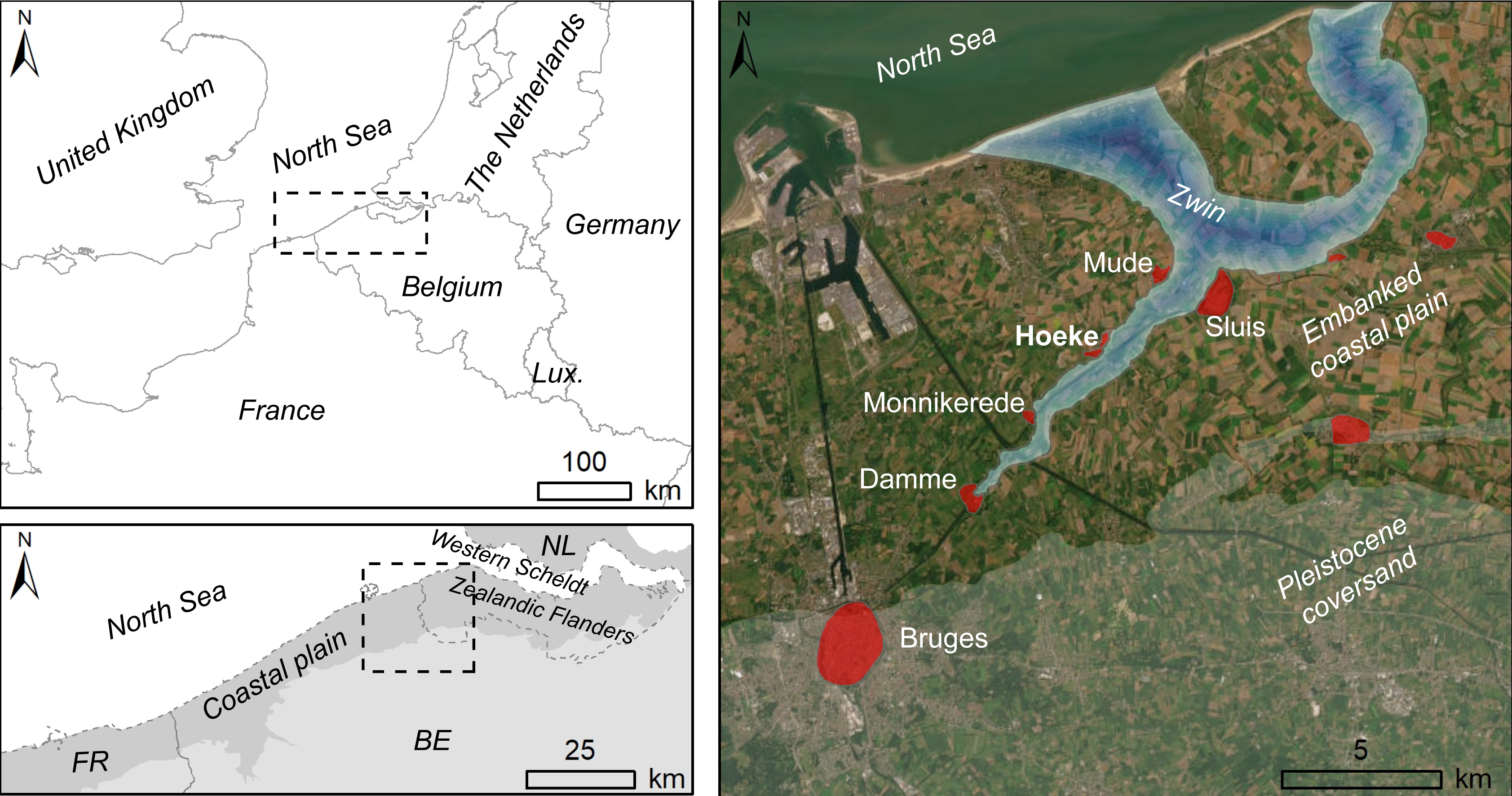
Figure 1. Location of the current Belgian coastal plain and the lost harbours (red areas) of the medieval Zwin region. The embankment of the coastal plain already started in the Middle Ages (modified from Bogemans et al., 2024).
2. Material
4Following electrical cone penetration test (CPT-e), seven cores were mechanically retrieved near Hoeke in 2021 using a hollow stem auger, yielding undisturbed 1 m liners (Bogemans & Heyvaert, 2023). The cores were sedimentologically described in detail for colour, texture, bedding types (primary sedimentary structures), deformation structures, biogenic structures, bedding plane characteristics and the presence of palaeontological and archaeological remains and pedological features. The grain size classes of the sand fraction were determined using a sand ruler. The sedimentary analysis of the cores allowed the identification of a series of architectural elements within a tidal environment, including channel elements sensu stricto, low energy channel elements, accretion forms, intertidal flats, salt marshes (supratidal), and abandoned channels (Bogemans & Heyvaert, 2023; Bogemans et al., 2024). The intertidal flat and salt marsh deposits are in this paper lumped under the term tidal flat deposits.
5Two cores (B3 and B5) were selected for palaeo-ecological analyses since they penetrated all four tidal channel systems detected in the cited sedimentary studies, dating from pre-Roman to late medieval times (Figs 2 and 3). Furthermore, the latest tidal channel system 4 also known as the Zwin is recorded in both cores. It was this channel that was active during the late medieval heyday of Bruges period (ca 1134–1500 AD).
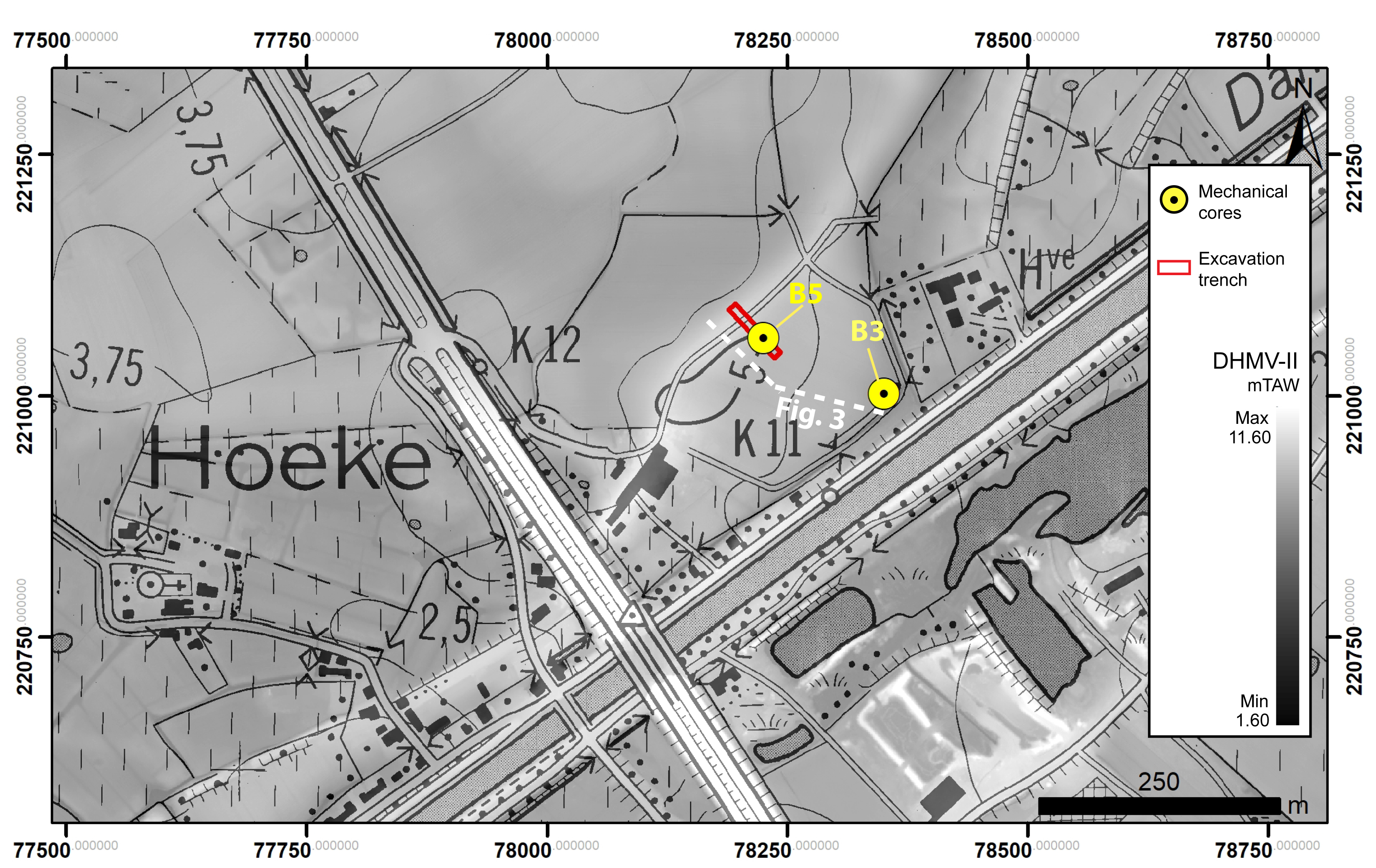
Figure 2. Study area of Hoeke with the location of the cores B3 and B5 (yellow circle) that were studied for ecological data in this paper. The white dashed line corresponds to the transect displayed in Figure 3. TAW = Tweede Algemene Waterpassing (Belgian Ordnance Datum).
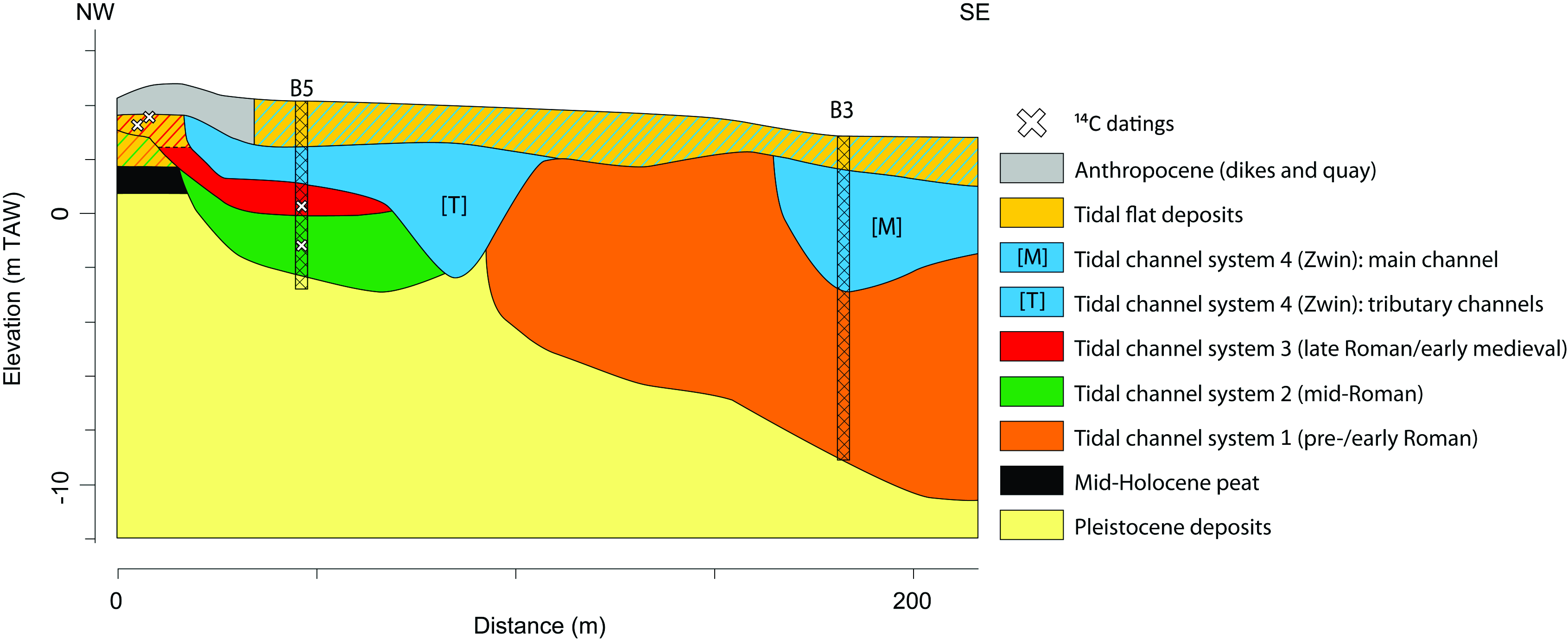
Figure 3. Schematic profile of the shallow subsurface near Hoeke with the location of cores B3 and B5. The sedimentary paleoenvironmental interpretation is based on the sedimentary analysis of data from undisturbed cores, observations made in the archaeological trenches, complimented with CPT-e data (the latter are calibrated on the basis of the core data). A more detailed description of the northwest section of the profile is given in figure 5 of Bogemans et al. (2024). The coloured stripes indicate what tidal channel system the tidal flat deposits belong to.
6Core B3 includes a more than 12 m thick Holocene sequence (Fig. 4), most of which consists of channel s.s. deposits (Bogemans & Heyvaert, 2023; Bogemans et al., 2024). However, these channel s.s. deposits are part of two different tidal channel systems; the oldest being active in the pre-/early Roman period (tidal channel system 1). The youngest system (tidal channel system 4) most probably developed after the storm surge of AD 1134 (according to historical sources), subsequently grew into an important commercial artery, but also silted up during the Late Middle Ages (Bogemans et al., 2024).

Figure 4. Location of the samples taken at 22 levels for diatom analysis (blue circles) in the depositional environments in core B3 (12 m deep) and their associated tidal channel systems. White circle: non-productive sample.
7The seven metre core B5 consists mostly of channel s.s. and accretion form deposits that belong to three different tidal channel systems (Fig. 5). The oldest tidal channel system 2 is dated to the mid-Roman period based on the articulated shell (Macoma) observed in living position at 5.37 m depth, which was radiocarbon dated to 195–489 AD (Laboratory code RICH-30446) (Bogemans et al., 2024). A second articulated shell (Cerastoderma) at 4.10 m depth radiocarbon dated to 548–795 AD (RICH-30447) allows to date tidal channel system 3 to a period in early medieval times. Archaeological excavations revealed the upper layers of channel system 3, which completely silted up and evolved into a salt marsh (De Clercq et al., 2022). A fragment of charcoal (Alnus) was dated to 682–880 AD (RICH-30881, youngest phase of the early medieval period), providing a terminus ante quem for the period of activity of system 3. Over more, a shallow cultural layer containing fragments of pottery and charcoal was found on top of it. The charcoal (Quercus) was radiocarbon dated to 895–1025 AD (RICH-30878, high medieval period). Finally, an abandoned/low energy channel is part of tidal channel system 4 (Bogemans et al., 2024). Along this channel the late medieval harbour of Hoeke emerged. It was demonstrated during archaeological excavations that this tributary channel was lined by a dike from which a wooden quay bordered the channel, indicating that the area was used as a harbour and location for ship repair during the late 13th and early 14th century (De Clercq et al., 2022). This channel 4 cut the aforementioned cultural layer for which the latter provides a terminus post quem dating, which is also well in line with the historical arguments for the emergence of the system in AD 1134 or shortly after.
8The final natural tidal channel system (4), active in the high and late medieval period and often referred to as the medieval Zwin, is thus documented in both cores B3 and B5 (tidal channel system 4, Fig. 3). Core B3 records the tidal channel s.s. deposits of the main navigable waterway during the heyday of Bruges, while the channel deposits in core B5 are thinner and correspond to a shallower tributary of the main tidal channel.
9Finally, intertidal and supratidal deposits (tidal flat deposits) cover the medieval Zwin channels deposits at both locations. The upper 30 to 40 centimetres are associated with plough layers from more recent periods.
10A total of 51 levels including sandy, clayey, and organic material were sampled for diatom analysis in core B3 (22 levels, Fig. 4) and core B5 (29 levels, Fig. 5). Organic deposits were only observed in core B5, especially in the accretion form deposits of system 2. A total of 17 levels, including these organic levels, together with silty clay levels intercalated in the different sediments of systems 3 and 4 in core B5, were also sampled for palynological analysis. The uppermost tidal flat deposits were not studied for palynological analysis, as spots of oxidation indicate poor pollen preservation. Macrobotanical analysis was only conducted on the very organic material accumulated in the accretion form deposits of system 2 between 4.56–4.92 m in core B5 (Fig. 5).

Figure 5. Location of the samples taken at 29 levels for diatom analysis (blue circles) and at 17 levels for palynological analysis (orange circle) in the depositional environments in core B5 (7 m deep) and their associated tidal channel systems. White circle: non-productive sample. The white dashed line between 4.56 and 4.92 m corresponds to the location of the macrobotanical remains that were analysed. The two shells analysed for radiocarbon dating are indicated at levels 4.10 and 5.37 m.
3. Methods
3.1. Diatom analysis
11Each sampled level (~0.5 g) was macerated for standard diatom analysis using H2O2 to remove organic material (Battarbee, 1986). At least 400 valves were counted using a ZEISS Axio Imager A1 transmitted light microscope at magnification 1000x (numerical aperture 1.3). The identification mainly follows Witkowski et al. (2000) and Lange-Bertalot et al. (2017) and the detailed taxonomy is given in Appendix 1 (see Supplementary material). The nomenclature was verified with AlgaeBase (Guiry & Guiry, 2024). The diatoms were classified according to life form and salinity preferences following the methodology developed by Vos & de Wolf (1993).
3.2. Palynological analysis
12About 1 mL of sediment was macerated following standard procedure for pollen analysis using HCl, KOH and HF treatments and acetolysis (Moore et al., 1991). The pollen concentration was calculated through the addition of Lycopodium tablets (batch number 100320201, X = 14 285, Appendix 2 in Supplementary material). The palynomorphs were analysed using a ZEISS Axio Imager A1 transmitted light microscope at magnification 400x (numerical aperture 0.75).
13The identification and nomenclature of the pollen and spores follow Moore et al. (1991) and Beug (2004), while non-pollen palynomorphs (NPPs) follow Miola (2012) and references therein, and Shumilovskikh et al. (2022). The pollen sum, used as a base for percentage calculations, includes only terrestrial pollen grains, for which at least 400 grains were counted. Other observed palynomorphs such as pollen grains from aquatic environments, spores and NPPs were also counted but not included in the pollen sum.
3.3. Macrobotanical analysis
14About 1800 mL of sediment was sieved over metal screens with a 500 µm and 250 µm mesh size. Material was picked under a binocular microscope Euromex ZE.1624 with magnification 10–45x. Macroremains of vascular plants have been identified using Cappers et al. (2006), Grosse-Brauckmann & Streitz (1992), Knörzer (2009) and a personal reference collection. Nomenclature of the vascular plants follows Lambinon et al. (2004). The classification of the ecological groups follows Runhaar et al. (2004).
3.4. Data analysis
15The diatom and palynological assemblages are plotted on percentage diagrams using Tilia (Grimm, 2020). Principal Component Analysis (PCA) was applied to both diatom and palynological records using Canoco (Ter Braak & Šmilauer, 2012) to interpret variation patterns in diatom succession through time. All levels were included in the PCA, except the diatom sample at 0.26 m in core B3, which represents an outlier with a very different species composition (see section 4.1.), related to distinctly different environmental conditions. All the species counted in both diatom and palynological analyses were included in the PCA, but for the readability of the figures, the number of species plotted was limited to the 50 best fitted species to the first two axes. The levels were classified according to lithology and the architectural elements defined by Bogemans & Heyvaert (2023).
4. Results
4.1. Diatom assemblages in core B3
16The channel s.s. deposits (systems 1 and 4, Fig. 4) and the overlying tidal flat deposits are dominated by marine (tycho)planktonic diatoms (~61–94%, Fig. 6). Delphineis minutissima (~26%) and Cymatosira belgica (~21%) are the dominant species. Delphineis surirella, Paralia sulcata and Rhaphoneis amphiceros are present in lower percentages (<8%). In addition, a large variety of marine/brackish epipsammic species (e.g. Navicula perminuta group and Plagiogrammopsis sigmoidea) are observed in lower percentages (~2–24%).
17The diatoms from the tidal flat sediments at 0.56 m were not counted because of the poor preservation of the valves, most likely due to dissolution. Although the uppermost disturbed deposits at 0.26 m hold fewer valves, they display an entirely different diatom assemblage than all lower levels, namely a reduction of the marine (tycho)planktonic species (~7%) and a dominance of brackish/freshwater species (~74%), mostly represented by Gedaniella cf. flavovirens (~30%) and species belonging to the genus Planothidium (~23%) and Gomphonema (~10%). The brackish epipelic Navicula cincta is also frequent (~6%). The diatom assemblage in this disturbed layer clearly points to eutrophic conditions.
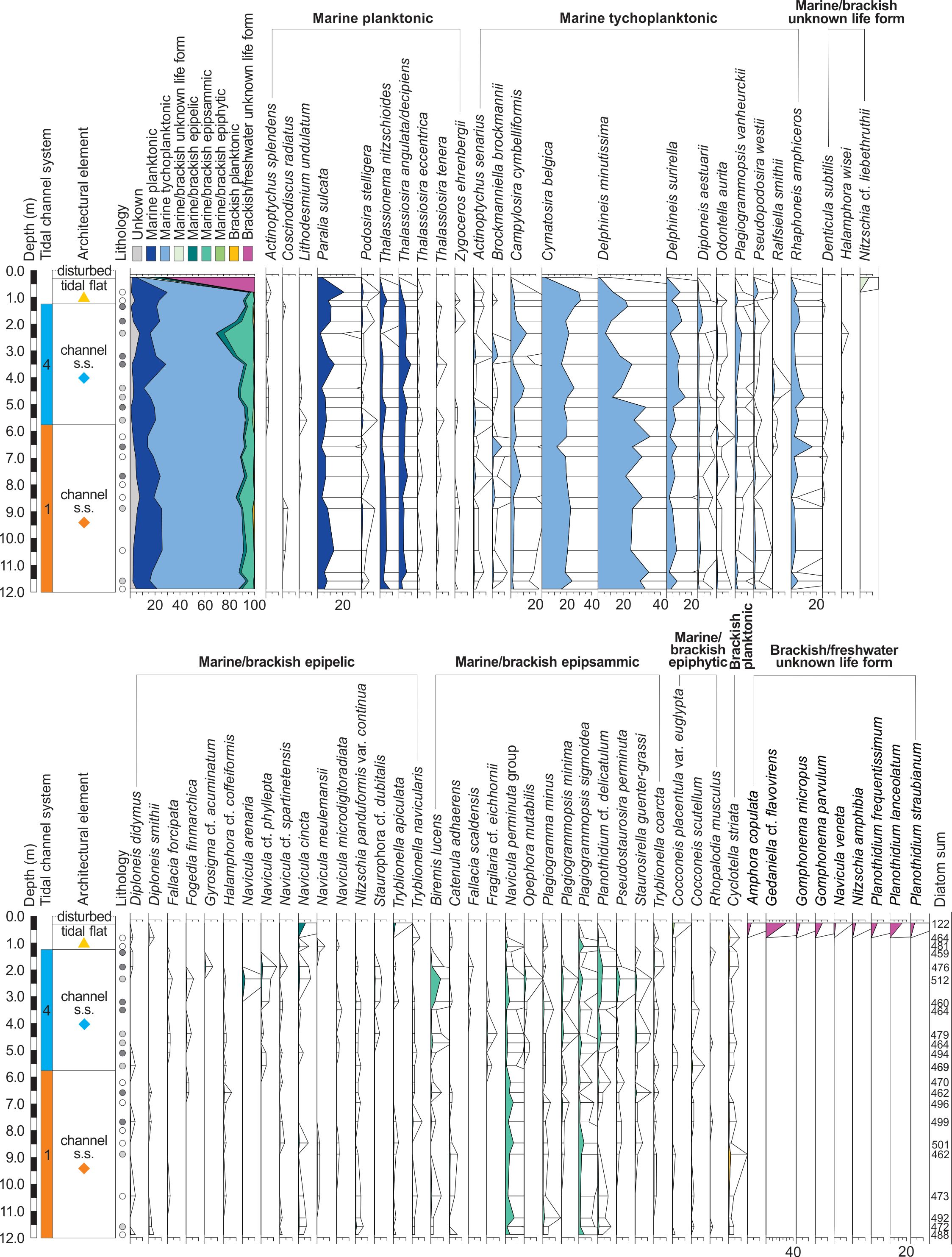
Figure 6. Percentage diatom diagram of core B3 (exaggeration x10, selected taxa) following the classification of Vos & de Wolf (1993). The legend of the lithology is given in Figure 8. The coloured geometrical symbols are specific to each architectural element within a tidal channel system and are used in the PCA analysis in Figure 8.
4.2. Diatom assemblages in core B5
18Marine (tycho)planktonic diatoms (~74–93%) dominate the sedimentary deposits in the three different tidal channel systems (2, 3, and 4, Fig. 5) and in the tidal flat deposits in core B5 (Fig. 7), while a multitude of marine/brackish epipelic and epipsammic diatoms are present in lower percentages (~4–20%). Five marine tychoplanktonic species (Cymatosira belgica, Delphineis surrirella, D. minutissima, Paralia sulcata and Rhaphoneis amphiceros) dominate the diatom assemblages in core B5, although their ratio varies within the different depositional entities.
19The accretion form deposits (system 2, Fig. 5) and the overlying channel s.s. deposits (system 3) display similar diatom assemblages dominated by Cymatosira belgica (~23%), Delphineis surirella (~16%) and D. minutissima (~13%). The accretion form deposits (system 3), the abandoned/low energy channel deposits (system 4) and the level at 1.54 m in the tidal flat deposits are dominated by Delphineis minutissima (~34%) and Cymatosira belgica (~20%). Finally, Paralia sulcata (~28%) and Cymatosira belgica (~23%) are dominant in the two uppermost sampled levels at 0.65 m and 1.06 m in the tidal flat deposits. The contribution of epipsammic and especially epipelic species is slightly higher in the accretion form deposits than in the channel deposits.
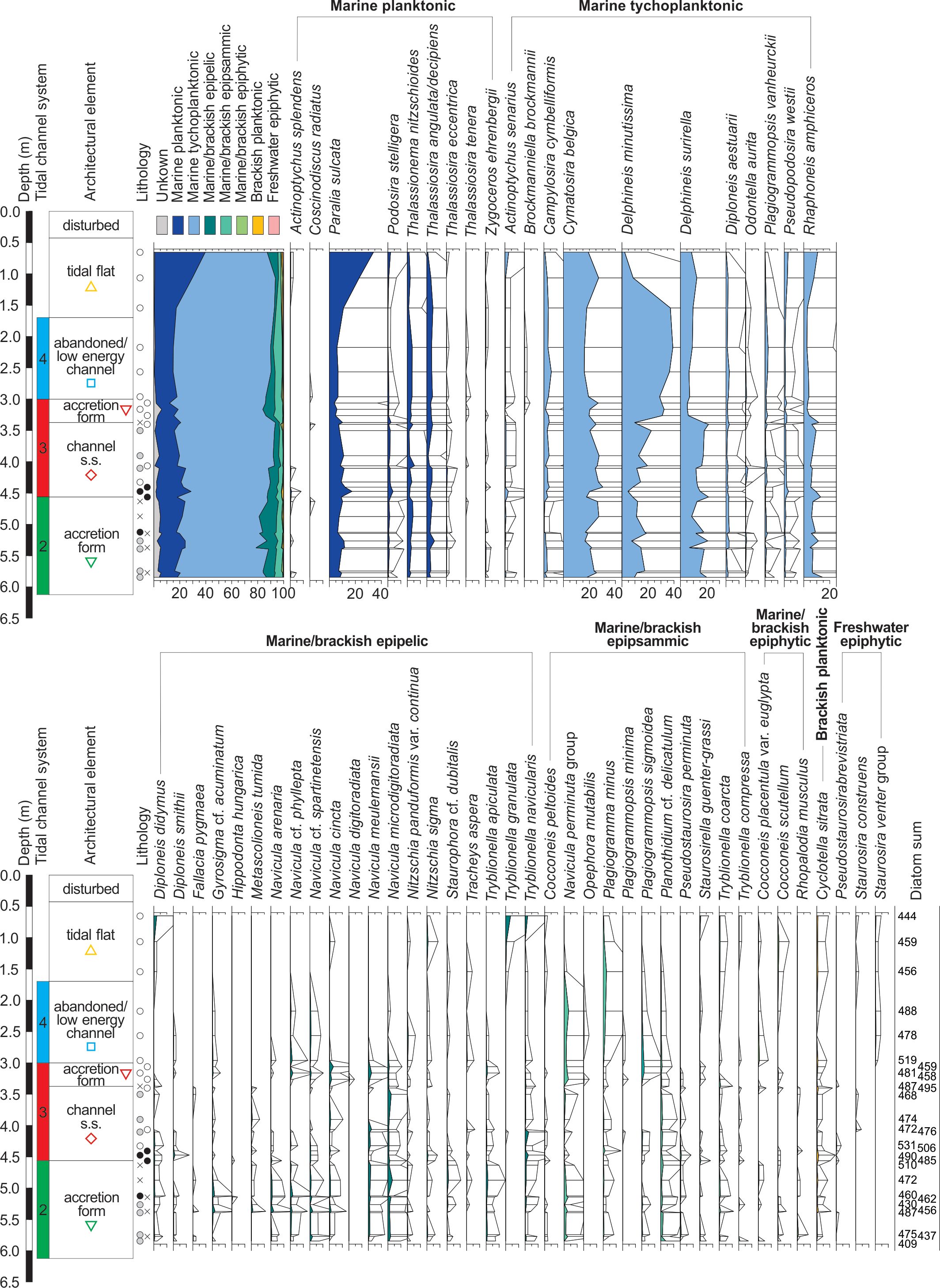
Figure 7. Percentage diatom diagram of core B5 (exaggeration x10, selected taxa) following the classification of Vos & de Wolf (1993). The legend of the lithology is given in Figure 8. The coloured geometrical symbols are specific to each architectural element within a tidal channel system and are used in the PCA analysis in Figure 8.
4.3. PCA of diatom assemblages in cores B3 and B5
20Axis 1 explains about 20% of the variation in the data and Axis 2 explains about 12%. The levels taken from the different architectural elements in cores B3 and B5 plot rather separately but do not show clusters with a specific lithology (Fig. 8). The channel s.s. deposits (systems 1 and 4) in core B3 plot close to each other and are characterised by a higher contribution of marine/brackish epipsammic species such as Plagiogrammopsis minima, Biremis lucens and Catenula adhaerens. The marine (tycho)planktonic Brockmanniella brockmannii, Plagiogrammopsis vanheurckii and Thalassiosira angulata/decipiens also characterise both channel s.s. deposits in core B3. The marine/brackish epipsammic Navicula perminuta group seems more typical of the oldest channel s.s. deposits. Also, the marine tychoplanktonic Odontella aurita and Delphineis minutissima seem more specific to the channels s.s. deposits of system 1, while Ralfsiella smithii and Campylosira cymbelliformis are more prevalent in the channel s.s. deposits of system 4.
21The sedimentary environments are well delimited in core B5. The levels from the channel s.s deposits (system 3) belong to one group mostly represented by heavily silicified diatoms such as the marine planktonic Paralia sulcata and Podosira stelligera and the tychoplanktonic Rhaphoneis amphiceros, Delphineis surirella and Pseudopodosira westii. Interestingly, Delphineis surirella and D. minutissima, which are dominant, display a negative correlation. The marine epipelic diatoms Diploneis didymus, Tryblionella navicularis, T. coarctata and Nitzschia sigma also characterise these channel s.s. deposits.
22The levels from both accretion form deposits (systems 2 and 3) plot close to each other in another group and are mostly represented by marine/brackish epipelic species that belong mostly to the genus Navicula (e.g. N. meulemansii, N. cf. phyllepta, and N. cincta), together with other epipelic species, such as Gyrosigma cf. acuminatum and Tryblionella apiculata. The lowest level at 2.96 m in the abandoned/low energy channel deposits (system 4) plots together with the underlying accretion form deposits (system 3), while the two upper levels at 2.56 m and 2.17 m in the abandoned/low energy channel deposits plot close to the channel s.s. deposits (systems 1 and 4) present in core B3.
23Finally, the levels taken from the tidal flat deposits in both cores seem more characterised by the marine tychoplanktonic Actinoptychus senarius and Paralia sulcata, as well as by the marine epipelic Tryblionella granulata, except for level 1.13 m in core B3 which plots more closely to the channel s.s. deposits (systems 1 and 4) of core B3.
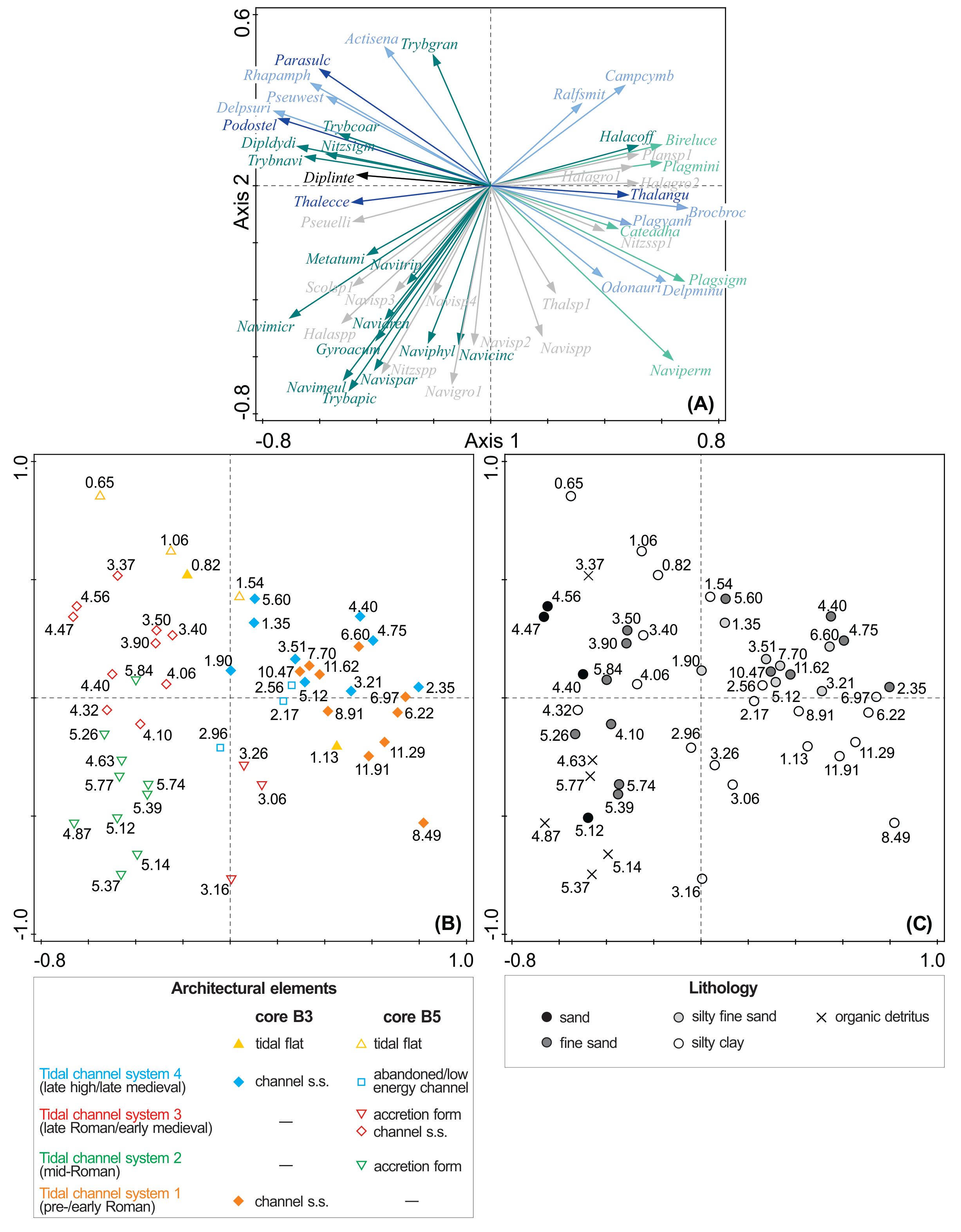
Figure 8. PCA plot of the 50 best-fit diatoms (A) in relation to the architectural elements (B) and lithology (C) in cores B3 and B5. The numbers plotted correspond to depth (m) and the colours of the arrows follow the ones used for the ecological groups in the percentage diatom diagram in Figure 7.
4.4. Palynological assemblages in core B5
24The pollen assemblages from the organic levels and the silty clay levels taken throughout the sedimentary record in core B5 (Fig. 9) are dominated by trees and shrubs (~55–81%). Corylus (~14–36%), Alnus (~9–22%), and Quercus (~8–14%) are the most abundant taxa. Other frequent trees include Betula (~7%) and Pinus sylvestris (~4%). Although present in low percentages on average (<2%), some trees display peaks in the organic level at 5.82 m in the accretion form deposits (system 2), such as Tilia (~5%), Ulmus (~3%), and Fraxinus excelsior (~3%). Heathland shrubs are frequent (~2–14%) and are mostly represented by Calluna vulgaris, which shows two peaks: the first (~10%) together with Vaccinium type (~4%) in the organic level at 5.82 m, and the second (~9%) in the silty clay level at 3.26 m in the accretion form deposits (system 3). Herbs reach on average ~19% in the organic layers in the accretion form deposits (system 2), with a minimum of ~5% at 5.82 m, and gradually increase to ~33% from 4.32 m onward in the silty clay intercalations present within the entities of systems 3 and 4. This is especially visible for Chenopodiaceae (~3% in system 2 vs ~8% in systems 3 and 4). Furthermore, herbs are dominated by Poaceae (~3–22%), while Cyperaceae are also frequent (~1–5%). From 3.26 m in the accretion form deposits (system 3) and in the overlying abandoned/low energy channel deposits (system 4), other herbs have become more common but still only in small percentages (<2%), such as Artemisia, Asteraceae-Liguliflorae, Cerealia type, Brassicaceae, and Senecio type.
25Aquatic taxa are mostly characterised by Sparganium type (~2%). Spores are mostly represented by Filicales and Sphagnum, and to a lesser extent by Pteridium aquilinum. Some Filicales were identified as Thelypteris palustris (~1%). Filicales and Sphagnum display opposite peaks in both accretion form deposits (systems 2 and 3). Sphagnum displays a peak of ~53% at 5.82 m (system 2), whereas Filicales reach only ~9%. Filicales is more abundant (~30%) in the upper accretion form deposits (system 3), together with Pteridium aquilinum (~3%), while Sphagnum is present in lower percentages (~3%). The fungus type HdV-13 (cf. Entophlyctis lobata, ~5%) and the protozoa types HdV-31 (Amphitrema flavum, ~5%) and HdV-32 (Assulina, ~2%) display peaks at 5.82 m. The protozoa type HdV-700 (linings of foraminifera, ~2%), acritarchs (~1%), and algae (e.g. Pediastrum spp., ~5%, and dinoflagellate cysts, ~3%) become more frequent from 4.32 m onward in the silty clay intercalations present in the entities of systems 3 and 4, except in the organic level at 3.37 m.
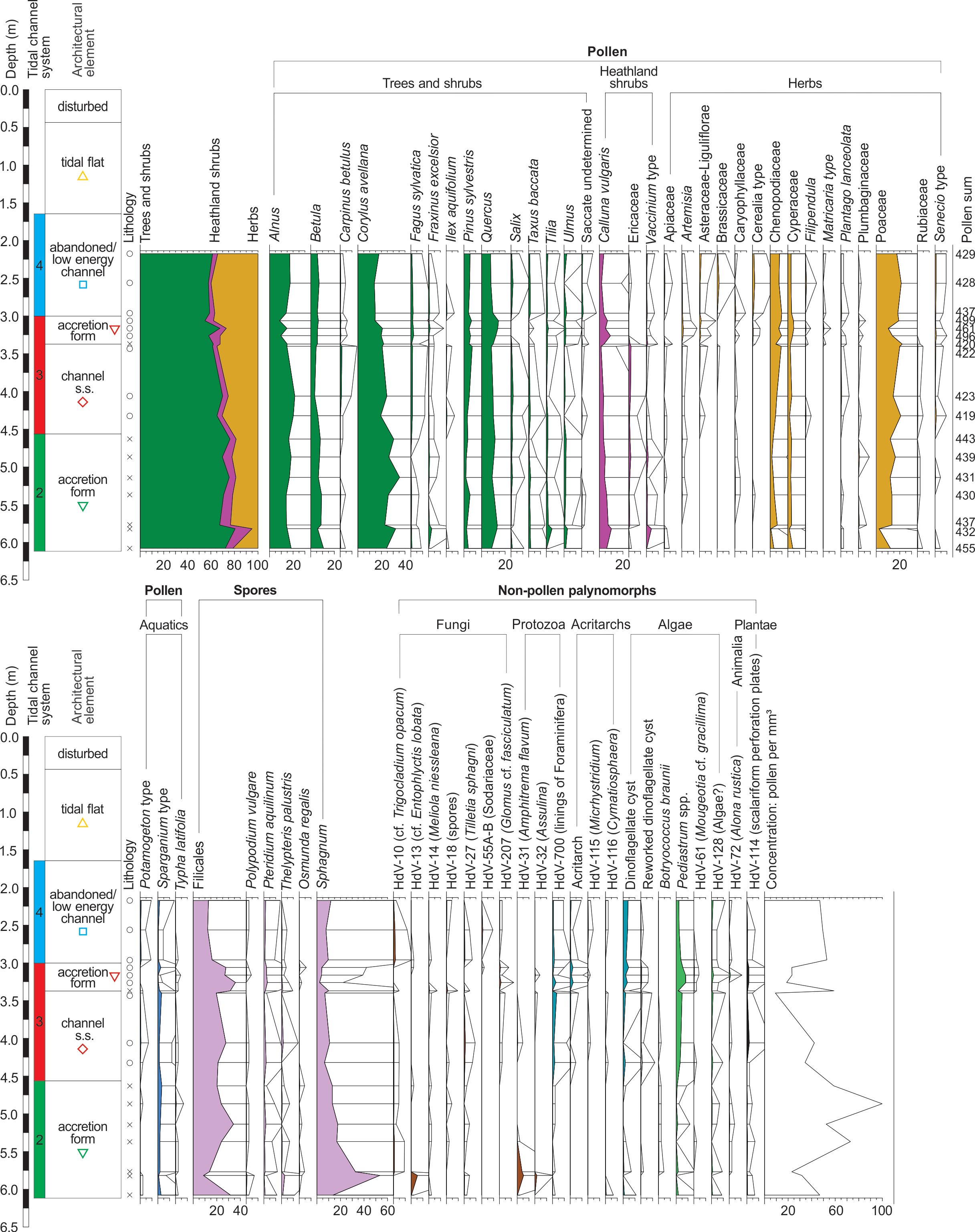
Figure 9. Percentage pollen diagram of core B5 (exaggeration x10, selected taxa). The legend of the lithology is given in Figure 10. The coloured geometrical symbols are specific to each architectural element within a tidal channel system and are used in the PCA analysis in Figure 10.
4.5. PCA of the palynological assemblages in core B5
26Axis 1 explains about 35% of the variation in the palynological data from core B5, and Axis 2 about 13%. Both architectural elements and lithology display well-delimited clusters (Fig. 10). The levels of silty clay plot separately from organic detritus, with distinct palynological assemblages, except for level 4.06 m where the silty clay intercalation is also rich in fine organic detritus; it plots together with the organic detritus.
27The organic samples display a cluster for the accretion form deposits (system 2), except level 5.82 m, which plots separately. Levels 3.37 m and 4.06 m from the channel s.s. deposits (system 3) plot close to the organic levels within the lower accretion form deposits (system 2). The organic deposits are globally characterised by Sphagnum, Botryoccocus braunii, and Corylus. The organic level at 5.82 m in the accretion form deposits (system 2) is more characterised by Ericaceae (including Calluna vulgaris and Vaccinium type) and Ulmus and Tilia, as well as by the fungi types HdV-13 (cf. Entophlyctis lobata), HdV-31 (Amphitrema flavum), and HdV-32 (Assulina). Alnus, Taxus baccata, and Salix seem to be more typical of the other organic levels.
28The silty clay intercalations plot opposite to the organic detritus and are divided into two clusters: the first is specific for the accretion form deposits (system 3), and the second includes two levels from the channel s.s. deposits (system 3) and the abandoned/low energy channel deposits (system 4). Both clusters are mostly characterised by dinoflagellate cysts, type HdV–700 (linings of foraminifera), and Chenopodiaceae, together with Pediastrum spp., Asteraceae–Liguliflorae, and Senecio type. Pinus sylvestris, Fagus sylvatica, Carpinus betulus, and Cerealia type are also specific for these deposits, although to a lesser degree.
29Additionally, the first cluster differs by taxa from or nearby aquatic environments, such as Menyanthes trifoliata, algae (e.g. types HdV-61 Mougeotia cf. gracillima and HdV-315 Spirogyra sp.), as well as Cyperaceae. The second cluster is more represented by herbs such as Poaceae, Brassicaceae, and Polygonum aviculare. Finally, Quercus seems to be equally significant in both silty clay and organic detritus intercalations from the two accretion form deposits (systems 2 and 3).
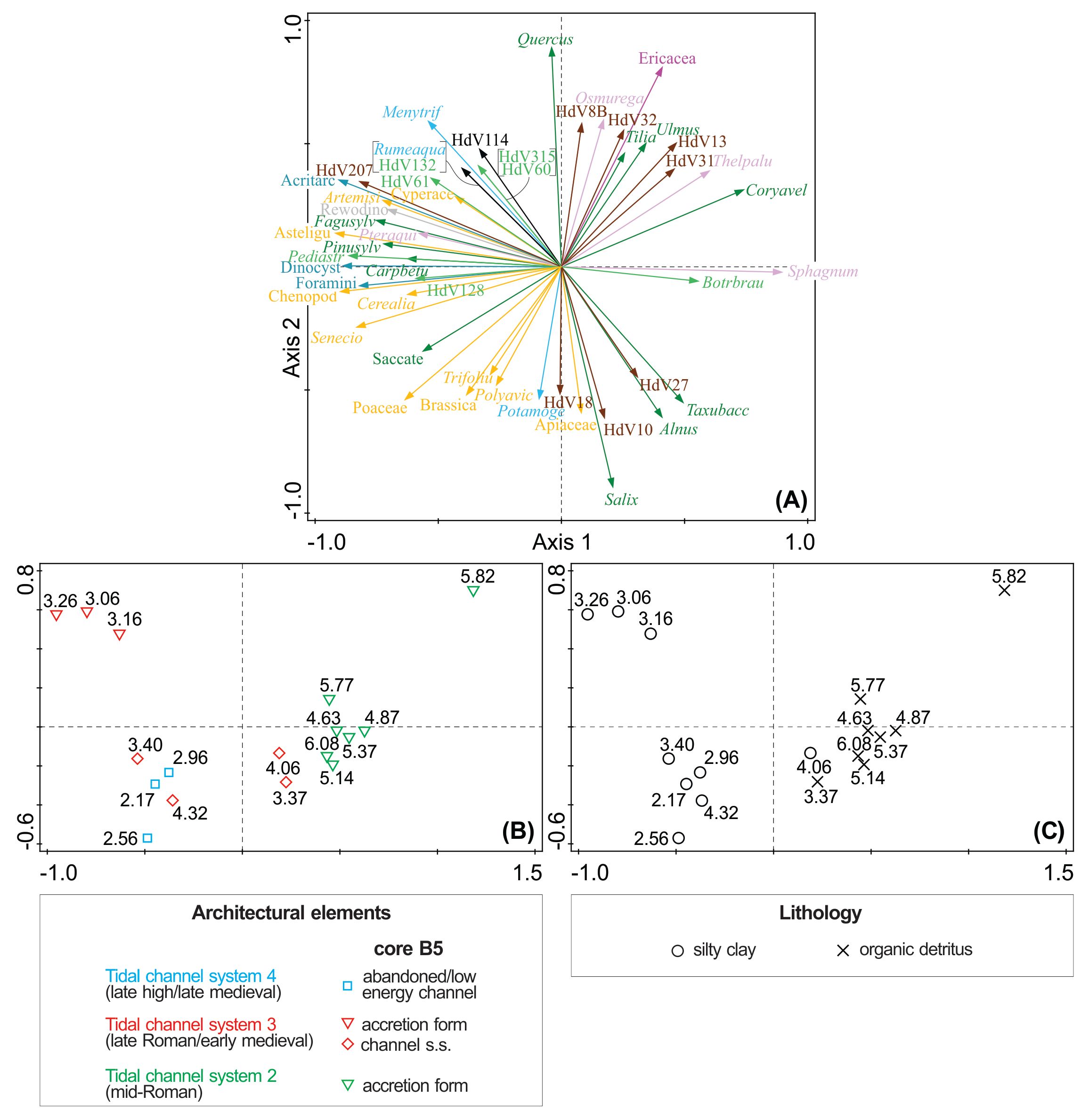
Figure 10. PCA plot of the 50 best-fit palynomorphs (A) in relation to the architectural elements (B) and lithology (C) in core B5. The numbers plotted correspond to depth (m) and the colours of the arrows follow the ones used for the ecological groups in the percentage pollen diagram in Figure 9.
4.6. Macrobotanical assemblages
30Two groups of macrobotanical remains were identified based on differences in preservation: one group had very good preservation, while the other had poor preservation (Table 1). Within the first group, two categories of taxa are observed. The first category holds seven types of mostly freshwater plants. Only Hippuris vulgaris and some species of the group Ranunculus subg. Batrachium, such as Ranunculus baudotii and Ranunculus trichophyllus, grow in brackish water (Runhaar et al., 2004). The second category includes three types, namely Atriplex sp., Glaux maritima or Anagallis arvensis, and Ranunculus sceleratus. More than 90% of the specimens belong to undifferentiated Atriplex sp. Seeds/fruits of species such as Urtica dioica, Chenopodium album, Stellaria media, Persicaria div sp., mostly very abundantly present in deposits of ruderal vegetations, are here totally absent. All the poorly preserved specimens in the second group belong to species that are present in peat, e.g. Alnus glutinosa and Menyanthes trifoliata (Allemeersch, 1991). The moss leaves of Sphagnum palustre and Drepanocladus sp. are also related to peat.

Table 1. Results of the macrobotanical analysis of a sample between 4.56 and 4.92 m in core B5. Sieved on 500 and 250 µm mesh size; sample volume 1800 mL; sp = sporadic; ../.. = whole specimens/fragments. All remains are seeds unless mentioned differently.
5. Discussion
5.1. The evolution of the diatom assemblages during the Late Holocene
31Diatom analysis has been widely used in palaeoenvironmental reconstruction of the Holocene coastal sequence in Belgium in the western coastal plain (Denys, 1985, 1989, 1994, 1999) and in east Flanders (Denys & Verbruggen, 1989), as well as in the Netherlands (Vos & de Wolf, 1993, 1994, 1997). No information however is available for the former Zwin region, where ecological data related to diatoms (and palynomorphs) are investigated for the first time in this study in the different tidal channel systems. Diatoms are usually studied in cores located adjacent to (and not directly in) tidal channels and inlets. It is important to stress that the infill of the tidal channel systems does not represent the environments where the diatoms actually lived but where the empty valves, behaving like mineral sediment particles, were transported into and deposited.
32The diatom assemblages from both cores are dominated by marine/brackish planktonic and tychoplanktonic species. In the Scheldt estuary, the dominance of these diatoms was always indicative of a usually less dynamic depositional environment with high silt content (Sabbe & Vyverman, 1991). Vos & de Wolf (1993) assume that marine planktonic and tychoplanktonic diatoms live in the offshore zone and that they are transported by tidal currents further inland, thus considering them to be allochthonous in the Dutch Holocene coastal deposits. Marine (tycho)planktonic diatoms outnumber local communities (benthic and epiphytic groups) in the very dynamic environments of coastal waters, tidal channels and inlets, but also sometimes in other sedimentary environments influenced by tides (e.g. intertidal and supratidal areas), which can hinder local palaeoecological reconstruction (Vos & de Wolf, 1993).
33Following the classification of ecological groups and sedimentary environments developed by Vos & de Wolf (1993), the dominance of marine tychoplanktonic and planktonic species in this study confirms the presence of open marine tidal channels that were strongly influenced by the sea. In such context, non-diatom criteria are often required to help the interpretation of the palaeoenvironmental conditions (Vos & de Wolf, 1993, 1997). Here, the results of the diatom analysis were combined with sedimentological characteristics. Furthermore, although marine tychoplanktonic and planktonic diatoms are dominant (Figs 6 & 7), it was still possible to differentiate the depositional entities in cores B3 and B5 based on diatom analysis, as they plot in distinct clusters (Fig. 8).
34The pre-/early Roman channel s.s. deposits (system 1) in core B3 are characterised by clay deposits with marine tychoplanktonic, as well as epipsammic species, which originally live on sandflats and in more exposed places (Sabbe & Vyverman, 1991; Vos & de Wolf, 1993).
35The mid-Roman to early medieval accretion form deposits in core B5 consist of an alternation of fine sand and organic detritus (system 2) and silty clay (system 3). Although the accretion form deposits hold different sediments, they are both more represented by epipelic species that are typical for mudflats and dominant in sheltered habitats. These epipelic diatoms must have been part of the local environment in the accretion form deposits during periods of lower energy, while a part might have been eroded and transported from nearby mudflats. In between the two accretion form deposits, the late Roman/early medieval channel s.s. deposits (system 3), composed of more sandy material, are characterised by strongly silicified marine (tycho)planktonic species, together with some epipelic species, reflective of more dynamic conditions, as well as mudflats in the vicinity.
36The medieval Zwin main channel deposits in core B3 (system 4) and the tributary channel deposits in core B5 (system 4) share similar diatom assemblages, and thus ecological conditions. Described as an abandoned/low energy channel by Bogemans & Heyvaert (2023), the tributary channel deposits are, however, characterised by marine (tycho)planktonic and epipsammic diatoms, which typically live in more exposed areas. The infill of the tributary channel consists of clay deposits and most probably received sediments and diatoms from a more dynamic environment such as sandflats located further away or from the main tidal channel recorded in core B3. The presence of a harbour along the tributary, where ship repairs occurred, suggests that the area must have been less dynamic compared to the main tidal channel.
37Aerophilous diatom assemblages that are adapted to intermittently wet conditions are usually abundant, together with marine planktonic and tychoplanktonic diatoms in supratidal environments (Vos & de Wolf, 1993). However, aerophilous diatoms were rare in the tidal deposits, i.e. Navicula cincta <1%. The tidal flat deposits in cores B3 and B5 consist of clay that is characterised by very similar diatom assemblages, with marine/brackish epipelic species of the high mudflat (Diploneis didymus and Tryblionella navicularis) together with marine (tycho)planktonic species (Paralia sulcata and Pseudopodosira westii). Such assemblages were also described in the western coastal plain of Belgium where intertidal sedimentation occurred (Denys, 1994, 1999). This suggests that marine conditions persisted in a more sheltered area. The reduction of the tidal influence in this area is most likely linked to the large-scale embankment that took place during the High and Late Middle Ages (Tys, 2013; Soens et al., 2014; Trachet et al., 2015). In addition, the much lower proportions of marine autochthonous diatoms (epipelic) compared to allochthonous diatoms (planktonic and tychoplanktonic) suggest that the sedimentation occurred at a fast rate (Denys, 1999).
38Level 0.26 m in core B3 is part of the plough layer, the top section of tidal flat deposits that has been disturbed by recent agricultural activity and hence reflects the local environmental conditions, namely eutrophic conditions and a decrease of the marine influence after embankment in more recent periods.
5.2. Terrestrial environments in tidal channel systems
39The pollen spectrum shows rather homogeneous terrestrial assemblages for the period from mid-Roman to late medieval times (systems 2, 3, and 4, Fig. 9). The sedimentary deposits are part of a dynamic environment, in which clastic sediment and organic material were transported in the tidal channel systems. Palynological data give us a general picture of the organisms and remains accumulated in the sedimentary deposits but do not provide a precise palaeoecological reconstruction of the surrounding environment. For example, the dominance of trees and shrubs does not necessarily reflect a woodland-dominated landscape during the entire period under study; instead, the sedimentary deposits include reworked material that can be much older than the landscape present at the time of deposition.
40The tidal channel systems received material from different sources. The organic layers in accretion form deposits of system 2 (mid-Roman period) are mainly associated with reworked peat, especially at 5.82 m, which has most probably been eroded and transported via tidal activity through channels. Indeed, the deceleration of the relative sea-level rise (to about 0.70 m/ka around 5500–5000 a cal BP) and the development of coastal barriers and dunes had resulted in an almost closed-off tidal landscape in the region, which gradually became covered by peatlands (Vos & Van Heeringen, 1997; Vos, 2015; Baeteman, 2018; Allemeersch et al., 2023). The peat accumulated over 2000–3000 years and eventually became an oligotrophic bog with taxa such as Sphagnum and Ericaeae. Such taxa are typical of the organic material, which most probably comes from mid-Holocene peat.
41The dryland trees and shrubs Ulmus, Tilia, and Corylus avellana also characterise the organic layers and were either part of the regional vegetation in the sandy inland at the time of peat growth or later at the time of tidal sediment deposition. This also applies to the wetland trees Salix and Alnus, which thrive in more humid environments. Taxus baccata was part of the carr vegetation in the coastal plain of Belgium during the deposition of the Subboreal biozone (the last half of the 5th and the 4th millenniums) and disappeared at the top of the Subboreal biozone (3rd millennium), when carr vegetation evolved into oligotrophic bogs (Deforce & Bastiaens, 2007). The peat deposits are rather thin at Hoeke (~1.20 m thick), and shallow tidal activity was sufficient to erode the stages of carr vegetation with Taxus.
42The badly preserved macrobotanical remains between 4.56 and 4.92 m in the accretion form deposits (system 2, Table 1) are also associated with older eutrophic to mesotrophic peat, which has most probably been reworked via tidal activity. Moreover, the well-preserved freshwater plants were not growing in situ at this location in the accretion form deposits, as a mix of organic and clastic material in these deposits indicates regular marine tidal influence, confirmed by the two levels at 4.63 and 4.87 m that were also studied for diatom analysis. Given their very good preservation, they most likely originate from the nearby vegetation, such as on salt marshes where freshwater pools could have developed. The seeds and fruits of the freshwater taxa may have been subsequently dispersed by floating upon the surface of the water, in the water column, or by aquatic birds such as ducks and geese (Oberdorfer, 1979).
43Despite some freshwater pools, a brackish environment remains characteristic of system 2. Atriplex sp. is dominant but could not be identified at the species level. The genus includes species such as Atriplex patula, which is a pioneer of very nutrient rich and rather wet soils, while the other Atriplex species are typical of brackish or even saltwater environments (Runhaar et al., 2004). In the absence of ruderal species, and with the type of material studied, the great number and nearly exclusive presence of Atriplex sp. within the group of pioneers indicates the presence of a brackish environment in the vicinity. The same problem occurs with Anagallis arvensis and Glaux maritima, which have very similar seeds that cannot be differentiated. Anagallis arvensis is more of a pioneer of nutrient-enriched wet soils, whereas Glaux maritima is present in brackish environments.
44The silty clay layers within the tidal channel systems 3 and 4 are mostly characterised by marine NPPs such as acritarchs, dinoflagellate cysts, and lining of foraminifera, which are typical of marine tidal deposits. Pollen types group several species thriving in different ecological conditions. In such marine tidal environment, the pollen types of many herbs are most probably associated with halophytes that grow in salt marshes, such as Chenopodiaceae (incl. Salicornia, Atriplex and Suaeda species), Senecio type (Aster tripolium), and Artemisia (Artemisia maritima) (Storme et al., 2020). Plumbaginaceae group only two species (Armeria maritima and Limonium vulgare), which are also typical of salt marshes. On the other hand, the freshwater environment identified in the macroremains suggest that some pollen types might also be associated with freshwater wetland taxa, such as Poaceae (Phragmites australis) Cyperaceae (e.g. Bolboschoenus), as well as freshwater aquatics (Sparganium type), that were also part of the surrounding vegetation.
45The dryland trees, Fagus sylvatica and Carpinus betulus, are more specific to the silty clay layers than the organic-rich layers. This might reflect the presence of beech-hornbeam woodland at the time systems 3 and 4 were active. Deforce et al. (2020) recently demonstrated the expansion of Fagus north of Belgium from about AD 600, associated with climatic conditions that became cooler and moister. Although in low percentages, Fagus does increase from 4.32 m onward, which corresponds to the phase in which the late Roman/early medieval tidal channel system 3 was active, a period that perfectly fits with the time of expansion of Fagus in the region, between AD 300 and 800 (Deforce et al., 2020).
46Finally, it seems that already in the accretion form of system 3 (late Roman/early medieval times), as well as in the medieval tributary channel deposits of system 4 (late high/late medieval times), the entities were characterised by more herbs from grassland and from agricultural activity, suggestive of an open landscape. This can be linked to the reclamation of the coastal plain, which became embanked on a large scale and was used for agriculture and pasturing (Tys, 2013; Soens et al., 2014).
6. Conclusion
47Ecological data were retrieved for the first time from sediment records of the former tidal channel systems in the Zwin region, spanning a period from pre-Roman to late medieval times. Although the tidal channel systems were dominated by allochthonous marine (tycho)planktonic diatoms that were transported from offshore, this research demonstrated that the sedimentary entities can be differentiated as they hold distinct diatom assemblages. These include marine/brackish epipsammic and epipelic diatoms, which indicate that the different tidal channel systems received material from mudflats and sandflats located further away. The pre-/early Roman channel s.s. deposits (system 1) consist of clay deposits that are mainly defined by epipsammic species originating from a dynamic environment located further away. The subsequent active tidal channel systems from the mid-Roman to the early medieval period (systems 2 and 3) include accretion form deposits that hold epipelic species, typical of more sheltered habitats. The channel s.s. deposits in between these accretion forms consist of more sandy material that is also characterised by epipelic diatoms, although different species, together with strongly silicified (tycho)planktonic diatoms, which are indicative of a more dynamic environment. The palynological assemblages are different in silty clay layers holding taxa typical of marine tidal deposits and organic layers mainly associated with reworked mid-Holocene peat, which has been eroded and transported via tidal activity. The macrobotanical remains included a poorly preserved group that is also associated with reworked peat, while the group with good preservation indicates the presence of brackish as well as freshwater plants, which were most probably growing in the vicinity. A beech-hornbeam woodland was probably part of the regional landscape from late Roman/early medieval times. During the later phases, the main and tributary tidal channels from the high and late medieval Zwin (system 4) share a similar aquatic ecology, with tychoplanktonic and epipsammic diatoms. Palynomorphs from grassland and possibly from agricultural activity in the tributary channel suggest an open landscape that can be linked to the reclamation and embankment of the coastal plain. The diatom assemblages in the tidal flat deposits overlying the deposits of the last tidal channel system of both cores suggest that the area rapidly silted up.
Acknowledgments
48This research was carried out as part of the UGent CRA project ‘High Tide, Low Tide’. Thomas Goovaerts is kindly thanked for assistance during the splitting of the cores and Sabine Van Cauwenberghe for macerating the pollen and diatom samples. We acknowledge the constructive comments of Luc Denys and Freek S. Busschers, which helped to improve the manuscript considerably.
Author contributions
49Coralie André, Stephen Louwye and Wim De Clercq carried out the conceptualization and the realisation of this study. Wim De Clercq, Jan Trachet and Maxime Poulain provided the studied material. Coralie André acquired the ecological data (pollen and diatom analysis) and performed the statistical analysis. Guidance in diatom analysis was performed by Bart Van de Vijver and Koen Sabbe. Guidance in palynological investigation was carried out by Annelies Storme. Frieda Bogemans conducted the sedimentological analysis and Luc Allemeersch realised the macrobotanical analysis. The draft of the manuscript was written by Coralie André and Stephen Louwye and all authors read and approved the final manuscript.
Data availability
50The diatom and pollen counts are available in Appendices 1 and 2 (see Supplementary material). The samples of this study are stored in the collection of the Geology Department at Ghent University.
References
51Allemeersch, L., 1991. Peat in the Belgian eastern coastal plain. In Gullentops, F. (ed.), Wetlands in Flanders: Contributions to Palaeohydrology of the temperate Zone in the last 15.000 years. Aardkundige Mededelingen, 6, 1–54.
52Allemeersch, L., Cruz, F., Germonpré, F., Laloo, P., Hinsch Mikkelsen, J., Storme, A., Vergauwe, R., Verhegge, J. & Verwerft, D., 2023. Veen als venster op het verleden. Onroerend Erfgoed, 16, 1–200. https://doi.org/10.55465/CTOM3365
53Baeteman, C., 2018. The coastal plain of Belgium, joint product of natural processes and human activities. In Demoulin, A. (ed.), Landscapes and Landforms of Belgium and Luxembourg. Springer, Cham, World Geomorphological Landscapes, 313–334. https://doi.org/10.1007/978-3-319-58239-9_19
54Battarbee, R.W., 1986. Diatom analysis. In Berglund, B.E. (ed.), Handbook of Holocene Palaeoecology and Palaeohydrology. John Wiley, Chichester, 527–570.
55Beug, H.J., 2004. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete. Friedrich Pfeil, München, 542 p.
56Bogemans, F. & Heyvaert, V.M.A., 2023. Sedimentary analysis of mechanically drilled cores nearby Hoeke, eastern coastal plain of Belgium. Professional Papers of the Geological Survey of Belgium, 321, 5–39.
57Bogemans, F., De Clercq, W., Poulain, M., Trachet, J., Verhegge, J. & Heyvaert, V., 2024. Sedimentary and stratigraphic architecture in the geoarchaeological context of Late-Holocene deposits in the eastern coastal plain of Belgium: Zwin region – Hoeke. The Holocene, 34, 519–530. https://doi.org/10.1177/09596836231225721
58Cappers, R.T.J., Bekker, R.M. & Jans, J.E.A., 2006. Digitale zadenatlas van Nederland = Digital Seed Atlas of the Netherlands. Barkhuis Publishing and Groningen University Library, Groningen, Groningen Archaeological Studies, 4, 502 p.
59De Clercq, W., Trachet, J. & Poulain, M., 2021. Verdwenen Zwinhavens. Provincie West-Vlaanderen, Brugge, 231 p.
60De Clercq, W., Demey, D., de Ruijsscher, D., Poulain, M., Trachet, J., Bogemans, F., André, C. & Vermeersch, J., 2022. De vorming van de Zwingeul en eerste bedijking in de laatmiddeleeumse haven te Hoeke : een eerste bilan (W.-Vl.). Archaeologia mediaevalis, 45, 45–46.
61Deforce, K. & Bastiaens, J., 2007. The Holocene history of Taxus baccata (yew) in Belgium and neighbouring regions. Belgian Journal of Botany, 140, 222–237.
62Deforce, K., Bastiaens, J., Crombé, P., Deschepper, E., Haneca, K., Laloo, P., Van Calster, H., Verbrugghe, G. & De Clercq, W., 2020. Dark Ages woodland recovery and the expansion of beech: a study of land use changes and related woodland dynamics during the Roman to Medieval transition period in northern Belgium. Netherlands Journal of Geosciences, 99, e12, https://doi.org/10.1017/njg.2020.11
63Denys, L., 1985. Diatom analysis of an Atlantic-Subboreal core from Slijpe (western Belgian coastal plain). Review of Palaeobotany and Palynology, 46, 33–53. https://doi.org/10.1016/0034-6667(85)90037-5
64Denys, L., 1989. Observations on the transition from Calais deposits to surface peat in the Western Belgian coastal plain: results of a paleoenvironmental diatom study. Geologische Dienst van België, Professional Papers, 241, 20–43.
65Denys, L., 1994. Diatom assemblages along a former intertidal gradient: a palaeoecological study of a subboreal clay layer (western coastal plain, Belgium). Netherlands Journal of Aquatic Ecology, 28, 85–96. https://doi.org/10.1007/BF02334249
66Denys, L., 1999. A diatom and radiocarbon perspective of the palaeoenvironmental history and stratigraphy of Holocene deposits between Oostende and Nieuwpoort (western coastal plain, Belgium). Geologica Belgica, 2, 111–140. https://doi.org/10.20341/gb.2014.014
67Denys, L., & Verbruggen, C., 1989. A case of drowning—the end of Subatlantic peat growth and related palaeoenvironmental changes in the lower Scheldt Basin (Belgium) based on diatom and pollen analysis. Review of Palaeobotany and Palynology, 59, 7–36. https://doi.org/10.1016/0034-6667(89)90003-1
68Dumolyn, J. & Leloup, W., 2016. The Zwin estuary: a medieval portuary network. In Solórzano Telechea, J.A., Arízaga Bolumburu, B. & Bochaca, M. (eds), Las sociedades portuartias de la Europa Atlántica en la Edad Media [Town port societies in Atlantic Europe in the Middle Ages]. Instituto de Estudios Riojanos, Logroño, Ciencias Históricas, 35, 197–212.
69Grimm, E.C., 2020. Tilia for windows: pollen spreadsheet and graphics program, version 3.0.1.
70Grosse-Brauckmann, G. & Streitz, B., 1992. Pflanzliche Makrofossilien mitteleuropäischer Torfe. III. Fruchte, Samen und einige Gewebe (Fotos von fossilen Pflanzenresten). Telma, 22, 53–102.
71Guiry, M.D. & Guiry, G.M., 2024. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. https://www.algaebase.org, accessed 11 January 2024.
72Knörzer, K.H., 2009. Geschichte der synanthropen Flora im Niederrheingebiet: Pflanzenfunde aus archäologischen Ausgrabungen. Verlag Zabern, Mainz am Rhein, Rheinische Ausgrabungen, 61, 484 p.
73Lambinon, J., Delvosalle, L. & Duvigneaud, J., 2004. Nouvelle flore de la Belgique, du Grand-Duché de Luxembourg, du nord de la France et des régions voisines (Ptéridophytes et Spermatophytes). 5e éd. Jardin botanique national de Belgique, Meise, 1168 p.
74Lange-Bertalot, H., Hofmann, G., Werum, M. & Cantonati, M., 2017. Freshwater Benthic Diatoms of Central Europe: over 800 Common Species used in Ecological Assessment. Koeltz Botanical Books, Schmitten-Oberreifenberg, 942 p.
75Miola, A., 2012. Tools for non-pollen palynomorphs (NPPs) analysis: A list of Quaternary NPP types and reference literature in English language (1972–2011). Review of Palaeobotany and Palynology, 186, 142–161. https://doi.org/10.1016/j.revpalbo.2012.06.010
76Moore, P.D., Webb, J.A. & Collison, M.E., 1991. Pollen Analysis. Blackwell scientific publications, Oxford, 216 p.
77Oberdorfer, E., 1979. Pflanzensoziologische Exkursionsflora. Ulmer Verlag, Stuttgart, 997 p.
78Pannier, N., 1970. De datering van de Duinkerke III-b transgressie en het dijksysteem ten noorden van Brugge. Handelingen der Maatschappij voor Geschiedenis en Oudheidkunde te Gent, 24, 113–126. https://doi.org/10.21825/hmgog.v0i0.1054
79Runhaar, J., van Landuyt W., Groen, C.L.G., Weeda E.J. & Verloove, F., 2004. Herziening van de indeling in ecologische soortengroepen voor Nederland en Vlaanderen. Gorteria, 30, 12–26.
80Sabbe, K. & Vyverman, W., 1991. Distribution of benthic diatom assemblages in the Westerschelde (Zeeland, The Netherlands). Belgian Journal of Botany, 124, 91–101.
81Shumilovskikh, L.S., Shumilovskikh, E.S., Schlütz, F. & van Geel B., 2022. NPP-ID: Non-Pollen Palynomorphs Image Database as a research and educational platform. Vegetation History and Archaeobotany, 31, 323–328. https://doi.org/10.1007/s00334-021-00849-8
82Soens, T., Tys, D. & Thoen, E., 2014. Landscape transformation and social change in the North Sea polders, the example of Flanders (1000–1800 AD). Siedlungsforschung. Archäologie–Geschichte–Geographie, 31, 133–160.
83Storme, A., Bastiaens, J., Crombé, P., Cruz, F., Louwye, S., Verhegge, J. & Deforce, K., 2020. The significance of palaeoecological indicators in reconstructing estuarine environments: A multi-proxy study of increased Middle Holocene tidal influence in the lower Scheldt river, N-Belgium. Quaternary Science Reviews, 230, 106113. https://doi.org/10.1016/j.quascirev.2019.106113
84Ter Braak C.J.F. & Šmilauer P., 2012. Canoco reference manual and user’s guide: software for ordination, version 5.0. Microcomputer Power, Ithaca, NY, 496 p.
85Trachet, J., 2016. Inland Outports. An interdisciplinary study of medieval harbour sites in the Zwin region. Unpublished Ph.D Thesis, Ghent University, Gent, 422 p.
86Trachet, J., Delefortrie, S., Dombrecht, K., Dumolyn, J., Leloup, W., Thoen, E., Van Meirvenne, M. & De Clercq, W., 2015. Turning Back the Tide: The Zwin debate in perspective. A historiographical review of the medieval port system northeast of Bruges. Revue du Nord, 413, 305–321. https://doi.org/10.3917/rdn.413.0305
87Tys, D., 2013. The medieval embankment of coastal Flanders in context. In Thoen, E., Borger, G.J., de Kraker, A.M.J., Soens, T., Tys, D., Vervaet, L. & Weerts, H.J.T. (eds), Landscapes or Seascapes? The History of the Coastal Environment in the North Sea Area Reconsidered. Brepols Publishers, Turnhout, Comparative Rural History of the North Sea Area, 13, 199–239.
88Verhulst, A.E., 1959. Middeleeuwse inpolderingen en bedijkingen van het Zwin. Bulletin de la Société belge d’Etudes Géographiques, 28, 21–54.
89Vos P., 2015. Origin of the Dutch coastal landscape: Long-term landscape evolution of the Netherlands during the Holocene, described and visualized in national, regional and local palaeographical map series. Barkhuis, Groningen, 372 p. https://doi.org/10.2307/j.ctt2204s8d
90Vos, P.C. & de Wolf, H., 1993. Diatoms as a tool for reconstructing sedimentary environments in coastal wetlands; methodological aspects. Hydrobiologia, 269, 285–296. https://doi.org/10.1007/BF00028027
91Vos, P.C. & de Wolf, H., 1994. Palaeoenvironmental research on diatoms in early and middle Holocene deposits in central North Holland (The Netherlands). Netherlands Journal of Aquatic Ecology, 28, 97–115. https://doi.org/10.1007/BF02334250
92Vos, P.C. & de Wolf, H., 1997. Palaeo-environmental diatom study of the Holocene deposits of the Province of Zeeland (SW Netherlands). In Fischer, M.M. (ed.), Holocene Evolution of Zeeland (SW Netherlands). Mededelingen Nederlands Instituut voor Toegepaste Geowetenschappen TNO, 59, 111–141.
93Vos, P.C. & Van Heeringen, R.M., 1997. Holocene geology and occupation history of the province of Zeeland. In Fischer, M.M. (ed.), Holocene Evolution of Zeeland (SW Netherlands). Mededelingen Nederlands Instituut voor Toegepaste Geowetenschappen TNO, 59, 1–109.
94Witkowski, A., Lange-Bertalot, H. & Metzeltin, D., 2000. Diatom Flora of Marine Coasts I. Koeltz Scientific Books, Königstein, Iconographia Diatomologica, 7, 925 p.
95Manuscript received 03.04.2024, accepted in revised form 24.09.2024, available online 17.12.2024.
96Supplementary material for this paper is available online at https://doi.org/10.20341/gb.2024.007.
