
New Rugose corals and refinements of the Tournaisian biostratigraphy of the Donets Basin (Ukraine)
Abstract
Specimens collected by Vassilyuk from the lower part of the Tournaisian strata of the southern Donets Basin and described as Campophyllum caninoides Sibly are re-examined. These specimens re-attributed to the Conilophyllum priscum (Münster, 1840), which is a well-known species in the lower part of the Lower Tournaisian of Western and Eastern Europe, Kazakhstan, North America and China (Junggar). Corphalia simplex (Perna, 1923) is for the first time identified at the top of Tournaisian in the Donets Basin. This species was described at similar stratigraphic levels on the eastern slope of the Southern Urals and in Belgium. The application of described species for interregional correlation of the Tournaisian subdivisions is discussed herein. Evolutionary relations between Corphalia and Dorlodotia near the Tournasian-Viséan boundary have been confirmed indirectly.
1. Introduction
1In the Donets Basin (Ukraine) the Tournaisian rocks are commonly fossiliferous carbonates. A detailed lithostratigraphic and biostratigraphic division has been achieved over the last decades (Aizenverg et al., 1963; Poletaev et al., 1990; Poletaev et al., 2011). Rugose and tabulate corals are widely distributed in some part of the sections. M.I. Lebedev and K.I. Lisitzin first identified Tournaisian corals of the Donbas early in the 20th century and used these fossils as guides to divide the sections and to correlate them with England and Belgium. I.I. Gorsky and V.D. Fomichev studied the Early Carboniferous corals of the Donets Basin during the 1920’s, but their studies have never been completed, and their collections were lost. Only brief and limited results of these studies have been published by Fomichev (1953) and Vassilyuk (1960). Vassilyuk studied the Early Carboniferous corals of the Donets Basin in detail and has proposed a coral zonation for the Lower Carboniferous strata (Poletaev et al., 1990). The stratigraphic distribution of corals taxa in the Tournaisian strata is highly heterogeneous. Most corals are found in the middle part of the Tournaisian (C1tc Zone) (Ogar, 2010). In the lower and upper parts of the Tournaisian, corals are much rarer. Unexpected new findings are reported in this paper, together with the redescription of specimens from Vassiljuk’s collection.
2. Settings
2The southern part of the Donets Basin is the only area in Ukraine where Tournaisian strata is easily accessible. They crop out in narrow bands through the towns of Novotroitske, Dokuchaevsk, Styla and Komsomolske (Donetsk region) (Fig. 1), where numerous natural sections along the Kalmius, Mokra Volnovakha and Sukha Volnovakha rivers have been studied since the 19th century. Most of these sections were unfortunately destroyed by anthropogenic activities.
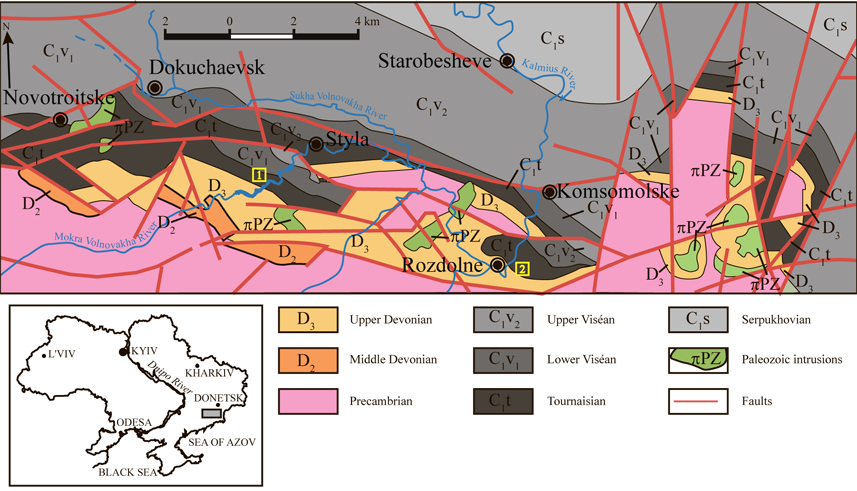
3Figure 1. Simplified geological map of the South Donbas (modified after Aizenverg et al., 1975) with location of coral localities: 1 - left bank of the Mokra Volnovakha River; 2 - left bank of the Kalmius River near Rozdolne town (destroyed section).
4The Tournaisian in the south of the Donets Basin is part of the regional Mokrovolnovakha Series (C11(A) lower part of the so-called "Continuous limestone strata" in older literature. This series is mainly composed of limestone, often dolomitized with rare interbedded mudstone and cherty inclusions. The Tournaisian part of the Mokrovolnovakha Series has a total thickness of 220 m (Poletaev et al., 2011).
5The Tournaisian deposits have been divided into complex biozones, mainly based on foraminiferal assemblages (Poletaev et al., 1990). In the regional stratigraphic chart of the Lower Carboniferous (Don-Dnieper trough) the Tournaisian corresponds to the Buzinov regional stage, including Bazaliev, Karakuba (lower substage), Volnovakha, Karpivka and Dokuchayevsk (upper substage) regional horizons.
6The basal part of the Basaliev horizon overlays disconformably the Devonian succession. The hiatus is approximately equivalent to the Gumerovo horizon of the East European platform and the Urals (Poletaev et al., 2011). The Dokuchayevsk horizon (C1va zone) that was historically considered to be lower Viséan is late Tournaisian in age.
7The rare corals of this succession are Conilophyllum collected in the light-grey massive dolomitized limestone of the middle part of the Karakuba horizon (40 m-thick) with the brachiopods Eudoxinia media; and Corphalia,collected from dark-grey and black argillaceous thin-bedded limestone of the upper part of the Karpivka horizon (9-10 m-thick).
3. Materials and methods
8The specimens described by Vassilyuk (1960) as Campophyllum caninoides Sibly were collected from two natural outcrops in the Kalmius River valley. Neither of them has been preserved in its original landscape. The Karst Valley located on the left bank of the Kalmius River near Rozdolne town (Karakuba town before 1946) has been completely destroyed by fluxing quarries. The second outcrop situated on the left bank of the Mokra Volnovakha River (right tributary of the Kalmius River) at the mouth of the Buzynova Valley, was only partly not flooded after the construction of the Styla reservoir (Fig. 2).
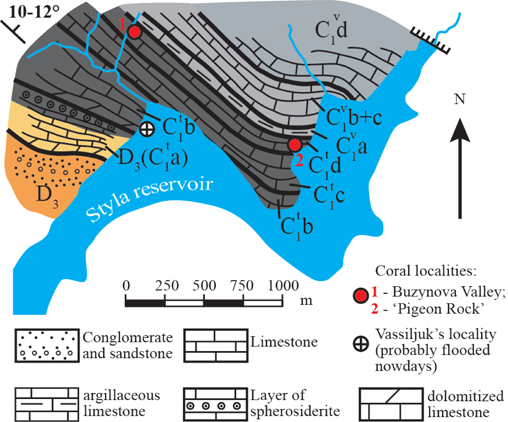
9Figure 2. Simplified geological scheme of the left bank of Mokra Volnovakha River (Styla Reservoir) with the coral localities. See figure 4 for zonal scheme.
10The Vassilyuk’s collection is housed in the National Museum of Natural History (NAS) of Ukraine (Kyiv, Ukraine) and available for additional studies. It contains several dozen specimens named as Campophyllum caninoides, including the specimen figured by Vassilyuk (1960, plate 14, figs 1-1b). The skeletons of all of the studied specimens are rock-free, partially eroded and altered by recrystallization.
11Cоrphalia specimens were collected by the author from two adjacent outcrops located on the left bank of the Mokra Volnovakha River (Fig. 2). One of them is situated on the left slope of the Buzynova Valley; the other is located in the Golub’yacha Skelya (‘Pigeon Rock’) section (Fig. 3), 10 m below the Tournaisian-Viséan boundary (Poletaev et al., 2011) (Fig. 4). The specimens are poorly preserved. Their external form is unknown. Fine microstructure of skeletal elements is not observed. The studied material is housed in Museum of Paleontology of Taras Shevchenko National University of Kyiv (Acronym TSNUK-2P264-A).
12Traditional methods to study corals (thin sections, polished surfaces and acetate peels) were applied.

13Figure 3. General view of the Golub’yacha Skelya (‘Pigeon Rock’) section, left bank of the Mokra Volnovakha River. The arrow indicates the sampling point of Corphalia simplex (Perna).
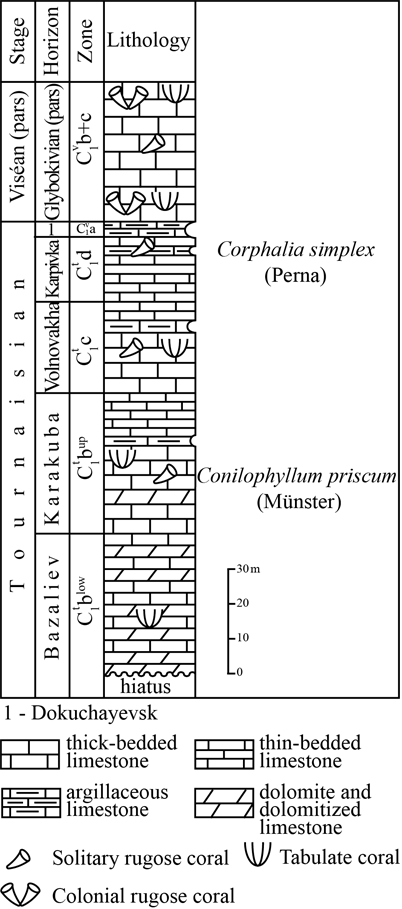
14Figure 4. General stratigraphic chart of the Tournaisian of the South Donbas (after Aizenverg et al., 1975; Poletaev et al., 2011) with position of the fossiliferous horizons.
4. Systematic palaeontology
15Suborder Caniniina Wang, 1950
16Family Cyathopsidae Dybowski, 1873
17Genus Conilophyllum Poty & Boland, 1994
18Type species. Conilophyllum streeli Poty & Boland, 1994, Hastière Formation, RC1β zone, Hastarian; Anseremme section, Belgium.
19Diagnosis.(After Denayer et al., 2011, p. 159). Cylindrical solitary coral. Major septa usually short, thick or not, limiting a free zone in the center of the tabularium. Minor septa of various length, not developed in some cases. Cardinal fossula inconspicuous. Dissepimentarium absent to wide, made of lonsdaleoid dissepiments and rare second order lonsdaleoid and simple interseptal dissepiments. Tabulae complete or slightly divided, horizontal or slightly convex, depressed laterally to form a gutter in some cases.
20Remarks. The presence of coral similar to Caninia cylindrica Scouler in McCoy, 1844 but with short septa (probably Conilophyllum) at layers with Spirifer medius (probably Karakuba horison) pointed by Fomichev (1953, pp. 525-526). The specimens described by Vassilyuk (1960) as Campophyllum showed that they actually belong to Conilophyllum Poty & Boland, 1994. Observations of the early growth stages of the Donetsk specimens demonstrate the thin septa that do not reach the axis, and an inconspicuous cardinal fossula that confirm the new taxonomic attribution. Conilophyllum from the Donets Basin is smaller than typical Siphonophyllia caninoides (Sibly, 1906) from the Arundian of North Wales (Somerville et al., 1986), and differs by its thin septa at all growth stages and its indistinct fossula. S. caninoides has usually a cardinal fossula marked by short cardinal septum. Only one specimen illustrated as Campophyllum caninoides Sibly by Vassilyuk (1960, plate 14, fig. 1c) from the uppermost Tournaisian (Dokuchaevsk horizon, C1va zone) probably really belongs to the Siphonophyllia caninoides (Sibly, 1906). It has large dimensions and more septa (n:d value 40:25 mm) than described below Conilophyllum priscum (Münster, 1840).
21Conilophyllum priscum (Münster, 1840)
22Fig. 5

23Figure 5. Conilophyllum priscum (Münster, 1840) from the left bank of the Kalmius River near Rozdolne town (destroyed section), lower Tournaisian, middle part of the Karakuba horizon, C1tbupzone. A-D: Specimen 1405/213-1. A: View of the specimen showing trochoid to cylindrical habitus, fragmentarily preserved calice and distinctly ribbed, covered by growth ridges. B: Transverse section. C: Detail of transverse section showing relationship between septa, dissepiments and tabulae. E-H: Specimen 1405/213. E: External view showing ceratoid habitus and growth ridges. F-G: Transverse sections. H: Longitudinal section. I-K: Specimen 1409/252 illustrated by Vassilyuk (1960, pl. 14, figs 1-1b). I-J: Transverse sections. K: Longitudinal section. L: Specimen 1405/219. View of preserved calice with signs of rejuvenation. Scale bar: 10 mm for all, except specially indicated D.
24*1840 Cyathophyllum priscum n. sp. Münster: p. 114, pl. 9, figs 12b-d (non fig. 12a).
251941 Campophyllum aff. caninoides Sibly; Volkova: p. 31, pl. 3, fig. 5.
26? 1959 Kassinella longiseptata Keller: p. 91, text-fig. 1, pl. 4, fig. 1.
27v 1960 Campophyllum caninoides Sibly; Vassilyuk: p. 62, pl. 15, figs 1-1b (non fig. 1c).
281967 Amplexus longiseptatus (Keller); Ivanovski: p. 38, pl. 1, figs 3, 4.
291982 Caninia tregaensis n. sp.; Poty: p. 54, figs 2-5.
301984 Caninia tregaensis Poty; Poty & Onoprienko: p. 204, pl. 31, figs 6-7
31? 1985 Guerichophyllum sp.; Sando & Bamber: p. 22, pl. 2, figs 6-7.
321987 Caninia tregaensis Poty; Wang & Zhao: p. 481, pl. 1, figs 1 a, b.
331993 Hebukophyllum priscum (Münster); Weyer: p. 187, pl. 1, figs 1-5; pl. 2, figs 1-6; pl. 3, figs 1-2; text-figs 4-6.
341994 Conilophyllum priscum (Münster); Poty & Boland: p. 205, pl. 1, figs 1-3; pl. 2, fig. 1.
351997 Conilophyllum priscum (Münster); Boland: p. 78, pl. 1, fig. 7; pl. 2, figs 2, 4; text-figs 4, 5.
362000 Hebukophyllum priscum (Münster); Weyer: p. 70, figs 1a, b.
372000 Conilophyllum tregaense (Poty); Weyer: p. 82, pl. 1, fig. 1.
382005 Conilophyllum priscum (Münster); Chwieduk: p. 430, pl. 16, figs 3, 4.
392011 Conilophyllum priscum (Münster); Denayer et al.: p. 159, pl. 1, fig. N.
40Diagnosis. (After Denayer et al., 2011, p. 159). Long Conilophyllum maximum 19 mm in diameter and having 35 major septa. Major septa usually short, amplexoid or caninoid. Minor septa short or not developed. Dissepimentarium absent to narrow, made of lonsdaleoid dissepiments. Outer wall undulating or arched.
41Lectotype. Chosen by Weyer (1993) from the syntypes housed in the Bavarian State Collection for Palaeontology and Historical Geology, Munich (Germany). Figured by Münster (1840, plate 9, fig. 12b) and by Weyer (1993, figs 4-1 and plate 1, figs 1a, b).
42Type locality and horizon. ‘Dark Clymenia limestone’ in former marble quarry Schübelhammer near Elbersreuth (Germany). Сonodonts extracted from rocks of the paralectotype no. 5 indicate the upper praesulcata conodont Zone (latest Famennian) or sulcata Zone (earliest Tournaisian) (Weyer, 1993).
43Material. Four specimens from Vassilyuk’s collection (1405/213, 1405/213-1, 1405/219 and 1405/243). All are eroded with fragments of the external surface and without proximal ends. Two specimens (1405/213-1 and 1405/219) have partly preserved calices. Five thin sections and 8 polished surfaces are available for study of these specimens. Lower Tournaisian, middle part of the Karakuba horizon, C1tbupzone.
44Description. Ceratoid, trochoid in the early stages of growth to cylindrical corals, up to 4.5 cm in height (Fig. 5A, E). Calice up to 2 cm in diameter. Specimen 1405/219 shows signs of rejuvenation within the 14 mm diameter calice (Fig. 5L). Central part of the calice flat 9 mm in wide, with steeply inclined edges up to 4 mm deep. External surface of corals distinctly ribbed with growth ridges. Major septa thin, variable in length (1.5-3 mm-long), occasionally continuous in inner row of the dissepiments and forming septal rugae on epitheca (Fig. 5D). Minor septa not developed. Cardinal fossula indistinct. Measured n:d values 22:6 mm; 25:8-9 mm; 28-29:10-11 mm; 29-34:16-17 mm; 30-39:18-22 mm. Dissepimentarium uneven in thickness in the same sections (Fig. 5B, K), up to 2.5 mm-wide in corallite 16-17 mm in diameter. Large elongated lonsdaleoid dissepiments. In longitudinal section dissepiments arranged in 1 to 4 indistinct rows, steeply declined (50-70º) towards the axis. Flattened dissepiments more steeply inclined appearing occasionally in the inner part of dissepimentarium (Fig. 5C, on the right). Tabularium biform, separated from dissepimentarium by thin inner wall, about 2/3 of the corallite diameter. Tabularium diameter/corallite diameter (mm) ratio: 8/10-11; 11.5-12.5/16-17; 13/18-22. Tabulae predominantly complete, horizontal in the central part and depressed near the dissepimentarium. Spaced at the distance of 0.5 to 1 mm.
45Discussion. The Donetsk specimens differ from those described in Belgium, Germany, Poland and the Omolon area by the underdevelopment of minor septa. In addition, some specimens from the Donets Basin have more major septa (39 in a diameter of 19-20 mm) than that specified in the diagnosis of the species. However considering the significant variability of Conilophyllum priscum these differences are not enough for the establishment of a new species in our opinion. Poty & Onoprienko (1984) noticed the similarity of some specimens described in Kazakhstan to Caninia tregaensis Poty, 1982 (= Conilophyllum priscum (Münster, 1840)). Kassinella longiseptata Keller (Keller, 1959) from the lowermost Tournaisian of Kazakhstan is very similar to Conilophyllum priscum. However, the illustrations of Kassinella longiseptata do not allow unambiguous conclusions about the presence of lonsdaleoid dissepiments. Therefore these specimens cannot be formally referred to the genus Conilophyllum. Similarly, Amplexus longiseptatus (Keller, 1959) from the Tournaisian of East Kazakhstan has thin septa, which are intercepted by ‘additional vertical plates’ (Ivanovski, 1967, p. 38). In our opinion, these ‘plates’ are lonsdaleoid dissepiments. Therefore, we include Amplexus longiseptatus into Conilophyllum priscum. Campophyllum aff. cannoides Sibly, 1906 (Volkova, 1941) from the Lower Tournaisian of Central Kazakhstan has shortened thin major septa, close n:d value (31:20 mm) and belongs to Conilophyllum priscum in our opinion. Specimens described as Guerichophyllum sp. (Sando & Bamber, 1985, p. 22, plate 2, figs 6-7) from middle Tournaisian (Coral Zones IB and C, IIA, B) of the Western Interior Province (United States) may be attributed to Conilophyllum priscum. They have typical features of the genus Conilophyllum but the scarce of the illustrated thin sections prevent a specific identification of the American specimens.
46Distribution. Germany, Rhenish Mountains, top of the presulcata to sanbergi Conodont zones; Rügen Island, upper Hastarian (‘Tn2’) (Weyer, 1993, 2000); Belgium, lowermost Tournaisian (Hastarian, RC1α-β zones) (Poty, 1982; Poty & Boland, 1994; Boland, 1997; Denayer et al., 2011); Omolon area (Russia), Strunian (Poty, 1999); China (Western Junggar), lower Tournaisian (Wang & Zhao, 1987); Pomerania (Poland), lower Tournaisian (Chwieduk, 2005); Kazakhstan, lowermost Tournaisian (Volkova, 1941; Ivanovski, 1967); Donets Basin, upper part of the lower Tournaisian, middle part of the Karakuba horizon (zone C1tbup)
47Family undetermined
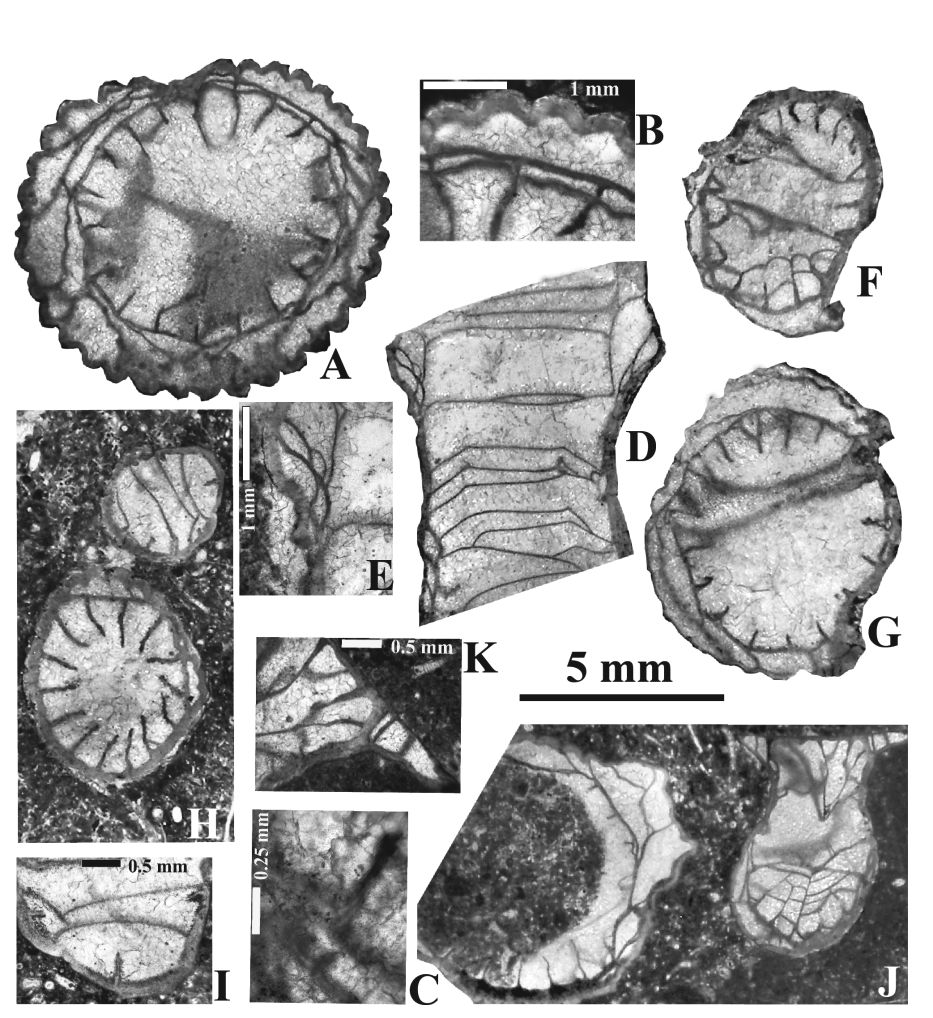
48Genus Corphalia Poty 1975
49Type species.Corphalia mosae Poty 1975, Lower Viséan of Corphalie section, Belgium.
50Diagnosis. (After Denayer et al., 2011, p. 173). Ceratoid then cylindrical solitary coral. Major septa short. Minor septa poorly developed or absent. No axial structure. Dissepimentarium narrow, made of small simple dissepiments and some lonsdaleoid dissepiments. Outer wall undulating, arched or festooned. Tabulae complete, subhorizontal.
51Remarks. Corphalia Poty includes small solitary corals with wavy outer wall commonly occurring in clusters. Its early morphology is amplexoid, but lonsdaleoid dissepiments appear in the late growth stage.
52Corphalia simplex (Perna, 1923)
53Fig. 6
54Figure 6. Corphalia simplex (Perna, 1923) from the uppermost Karpivka horizon (C1td2 subzone), left bank of the Mokra Volnovakha River. A-E: Specimen 2P264-A/d-3-1, Buzynova Valley section. A: Transverse section. B, C: Details of the transverse section. D: Longitudinal section. E: Detail of the longitudinal section. F-G: Specimen 2P264-A/d-6-1, Golub’yacha Skelya (‘Pigeon Rock’) section; transverse sections. H-K: Specimen 2P264-A/d-6-2, Buzynova Valley section. H-J: Transverse sections of the closely conivent corallites. I: Detail of transverse section showing development of the septa in early morphology. K: Fragment of flattened possible attachment wall area. Scale bar: 5 mm for all, except of specially indicated B, C, E, I and K.
55*1923 Thysanophyllum simplex n. sp.: Perna, p. 22, pl. 3, fig. 1.
562003 Dorlodotia sp.; Poty et al.: p. 104-105, pl. 3, fig. 6 (non fig. 12).
572011 Corphalia sp. nov.; Denayer et al.: p. 173, pl. 5, fig. U.
58Diagnosis (Modified from Perna, 1923). Corphalia with a mean diameter of 12-13 mm having 20 septa. Major septa of various length and thickness. Minor septa not developed. Dissepimentarium composed of one-two incomplete rows of lonsdaleoid dissepiments. External wall wavy or festooned, 0.2 mm-thick.
59Lectotype. Collection No. 927, St-Petersburg, CNIGR Museum. Figured by Perna (1923, plate 3, fig. 1).
60Type locality and horizon. Western slope of the South Urals near Ablyasovo village (Magnitogorsk Synclinorium); uppermost Tournaisian.
61Material. Eight specimens fully enclosed in rock fragments from the two contiguous localities situated on the left bank of the Mokra Volnovakha River: 1 – Buzynova Valley and 2 - ‘Pigeon Rock’ section. Five thin sections, 2 acetate peels and 2 polished surfaces. Upper Tournaisian, uppermost Karpivka horizon, 5 m below the Karpivka/Dokuchaevsk boundary, C1td2 subzone.
62Description. Small gregarious solitary corals commonly occurring in clusters. Attachment structures occasionally observed (Fig. 6J, K). Major septa thin, variable in length, very short (up to 0.3 mm) and widely spaced in early stages of development. Base of the septa included in the wall stereoplasm (Fig. 6I). Length of the septa half of the corallite radius (young stage of 5 mm in diameter). In later growth stages, septa shortened to 0.4 mm (Fig. 6J; left corallite) with development of dissepimentarium. Septa typically intercepted by lonsdaleoid dissepiments, but septal crests on the wall are occasional (Fig. 6C). Minor septa not developed but reflected by septal furrows on the wall (Fig. 6A, B). Cardinal septum and fossula inconspicuous. N:d values: 19:8-9 mm. Dissepimentarium appearing in corallites larger than 5 mm in diameter (Fig. 6D, H) and dominated by lonsdaleoid dissepiments of various size. Interseptal dissepiments rare or not developed. Dissepimentarium approximately 1/3 of corallite radius in width (in diameter of the corallite 9 mm). Tabularium consisting of mainly complete mesa-shaped tabulae, typically horizontal in the central part and depressed near the dissepimentarium. In longitudinal section (Fig. 6D) tabulae irregularly arranged and separated by 0.5 to 1.5 mm. External wall 0.2 mm-thick, wavy or festooned.
63Discussion. This species first was described as Thysanophyllum simplex Perna from the Southern Urals (Perna, 1923). Its assignment to the genus Thysanophyllum was incorrect because its colonial habitus inferred by Perna (1923) and subsequent studies (unpublished report from Degtyarev in 1962) has not been confirmed. The specimens from the Donets Basin differ from the Uralian ones by a slightly lesser maximum corallite diameter. C. mosae described by Poty (1975, 1981) from the early Viséan of Belgium is smaller than C. simplex n:d value (23:7.5 mm vs. 19:9 mm). Corphalia fourmarieri (Charles, 1933) (Denayer, 2015) from the latest Tournaisian (RC4β1 and MFZ8 biozones) is smaller in diameter and has less septa (16-17:5 mm). In both Turkish species Corphalia fourmarieri and Corphalia sp. (RC4β1 and MFZ8, Denayer, 2015) the development of concentric interseptal and lonsdaleoid dissepiments are very typical. Interseptal dissepiments are not developed in C. simplex nor in specimens figured under the name Dorlodotia sp. (Poty et al., 2003, plate 3, fig. 6, not fig. 12, a genuine Dorlodotia) and Corphalia sp. nov. (Denayer et al., 2011, p.173, plate 5, fig. U) from the latest Ivorian (RC4β1 biozone) of Belgium. These last examples display the same morphology and size than Ukrainian specimens and can be confidently attributed to C. simplex (Perna).
64Distribution. Uppermost Tournaisian of the eastern slope of the Southern Urals, (Beryozovka Suite, Kos’va horizon). The occurrence of Corphalia simplex in the marls among volcanic rocks of the Beryozovka Suite (3 km south of the Ablyazovo village) was noticed by Degtyarev (1964), Kachanov (1965) and Donakova et al. (1968). Uppermost Tournaisian of the Donets Basin (uppermost Karpivka horizon, C1td2 subzone); uppermost of Ivorian (Avins Member, RC4β1) of Southern Belgium.
5. Discussion
5.1. Phylogeny
65It should be noted that the two presently described species share similar features. Among them are a solitary habitus, the presence of lonsdaleoid dissepiments, the absence of an axial structure and a conspicuous cardinal fossula, and a wavy or festooned external wall. These similarities may only be the result of convergence because both species occur in distinct stratigraphic levels separated by large stratigraphic interval devoid of any fossil coral.
66The attribution of Corphalia at the family level is still unresolved. In our opinion it can be determined as a result of the establishment of related taxa. Significant progress in this direction has been made in regard of the recognition of the Corphalia-Dorlodotia lineage (Denayer & Poty, 2011), which suggests that these genera belong to the same family or subfamily (Denayer, 2013) and not to the Thysanophyllinae as suggested by Hill (1981).
67Considering the morphological similarity Corphalia simplex (Perna, 1923) and Dorlodotia pseudovermiculare (McCoy, 1849) as well their distribution in the adjacent horizons of the Tournaisian and Viséan on the eastern slope of the Southern Urals (Kachanov, 1965) and the Donets Basin (Ogar, 2010), the phylogenetic relationships of these species is very probable. In this case Corphalia simplex could be an ancestor of acolumellate Dorlodotia as suggested by Denayer & Poty (2011).
5.2. Correlation
68In Western Europe, Conilophyllum is a guide taxon for the lowermost Tournaisian (Poty et al., 2006). In Pomerania (Poland, Chwieduk, 2005) and the Rhenish Massif (Germany, Weyer, 1993), the genus appears slightly earlier, before the Devonian-Carboniferous Boundary, whereas in Omolon (E Russia, Poty, 1999), it is known in the late Famennian. Chwieduk (2005) interpreted this variable stratigraphic distribution as a migration from East to West. This hypothesis is compatible with palaeogeographic reconstructions but does not take into account local and regional sedimentation features. Inter-regional correlation of the Tournaisian units of the Donets Basin, the East European platform and other regions is controversial (Hecker, 2001, Menning et al., 2006, Poletaev et al., 2011). The Karakuba horizon of the Donets Basin is correlated with the lower part of the Cherepet or mostly Upa horizon (Aizenverg et al., 1963; Poletaev et al., 1990; Poletaev et al., 2011) of the East European platform. This correlation is confirmed by the presence in the Donets Basin (Poletaev et al., 2011) of the foraminifera Prochernyshinella disputabilis. In this case the Karakuba and Upa horizons correspond with Landelies Formation (former ‘Tn2a’) of Belgium (Hecker, 2001).
69The appearance of Conilophyllum priscum in the Donets Basin is close to the upper limit of its occurrence on the Rügen Island (Germany), where it was found in the basal upper Hastarian (‘Tn2a’, uppermost RC1β biozone) (Weyer, 2000).
70Unfortunately the Tournaisian conodonts of the Donets Basin are insufficiently studied. Indeed, Conilophyllum priscum was identified much higher than layers where Polygnathus parapetus and Siphonodella praesulcata were found (Lipnjagov, 1975). Hence, the stratigraphic distribution of Conilophyllum priscum is limited, but still rather long from the uppermost Famennian (Omolon area) to the lower Tournaisian (Conilophyllum interval zone RC1 of Belgium) (Poty et al., 2014).
71Corphalia simplex (Perna, 1923) may have a more significant correlation potential. The distribution of this species includes a narrow stratigraphic interval of the latest Tournaisian: the uppermost Karpivka horizon (C1td2 subzone) in the Donets Basin, the Kos’va (Lun’evo) horizon of the South Urals, and the uppermost Ivorian (RC4β1) Avins Member of the Longpré Formation of Belgium and in equivalent level in the Bristol area (England, Julien Denayer, written communication, December 2015). This corresponds approximately to the generally accepted correlation of the uppermost Tournaisian units (Hecker, 2001, Menning et al., 2006, Poty et al., 2006, Poletaev et al., 2011).
6. Conclusion
72The recent identification of Conilophyllum priscum (Münster, 1840) and Corphalia simplex (Perna, 1923) in the Donets Basin extends the data on their distribution and Early Carboniferous palaeogeography. Studied species are guide elements of the early Tournaisian RC1 biozone and the latest Tournaisian RC4 biozone of Western Europe that promotes interregional correlation with other regions of Eurasia.
7. Acknowledgements
73The author is very grateful to Julien Denayer (University of Liège, Belgium) for his critical remarks and great assistance to improve of the manuscript. Markus Aretz (Géosciences Environnement Toulouse, Université Paul Sabatier, France) is thanked for supporting and improving this paper.
8. References
74Aizenverg, D.E., Brazhnikova, N.E., Novik, E.A., Rotay, A.P. & Shul’ga, P.L., 1963. Stratigrafiya kamennougolnykh otlozheniy Donetskogo basseyna [Stratigraphy of the Carboniferous deposits of the Donets Basin]. Trudy Instituta geologicheskikh nauk, Akademia Nauk Ukrainskoy SSR, Seriya stratigrafiya i paleontologiya, Kiev, 37, 1-82. [In Russian].
75Aizenverg, D.E., Lagutina, V.V., Levenshtein, M.L. & Popov, V.S. (eds), 1975. Field excursion guidebook for the Donets Basin. VIII International Congress on Carboniferous Stratigraphy and Geology, Nauka, Moscow, 306 p. [In Russian and English].
76Boland, K., 1997. Caninoid rugose corals of the Lower Tournaisian (Hastarian) of Belgium: systematics and evolution. Boletín de La Real Sociedad Española de Historia Natural, Sección Geológica, 91, 73-84.
77Charles, F., 1933. Contribution à l’étude des terrains paléozoïques de l’Anatolie du Nord-Ouest (Asie mineure). Mémoires de la Société Géologique de Belgique, 7, 54-152.
78Chwieduk, E., 2005. Late Devonian and early Carboniferous Rugosa from Western Pomerania, northern Poland. Acta Geologica Polonica, 55/4, 393-443.
79Degtyarev, D.D., 1964. Korally Beryozovskoy svity (Nizhniy karbon Magnitigorskogo sinklinoriya) [Corals of the Beryozovka Suite (Lower Carboniferous of the Magnitogorsk Synclinorium)]. Sverdlovsk Mining Institute. Abstracts of the research works carried out in 1962. Sverdlovsk, 7-8. [In Russian].
80Denayer, J., 2013. Biostratigraphie et paléobiogéographie des coraux tétracoralliaires du Carbonifère inférieur de la Turquie. Unpublished PhD Thesis, Université de Liège, Liège, Belgique, 1, 346 p.; 2, 40 pls.
81Denayer, J., 2015. Taxonomy, biostratigraphy and palaeobiogeography of the Late Tournaisian rugose corals of north-western Turkey. Paläontologische Zeitschrift, 89, 313-333.
82Denayer, J. & Poty, E., 2011. Origin and evolution of Dorlodotia (Rugosa). In Aretz, M., Delculéee, S., Denayer, J. & Poty, E. (eds), 11th International Symposium on Fossil Cnidaria and Porifera, Liège, 19-29 August, 2011, Abstracts volume. Kölner Forum für Geologie und Paläontologie, 19, 37-39.
83Denayer, J., Poty, E. & Aretz, M., 2011. Uppermost Devonian and Dinantian rugose corals from Southern Belgium and surrounding areas. In Aretz, M. & Poty, E. (eds), 11th International Symposium on Fossil Cnidaria and Porifera, Liège, 19-29 August, 2011, Field-guides. Kölner Forum für Geologie und Paläontologie, 20, 151-201.
84Donakova, L.M., Katchanov, E.I. & Proskurina, L.K., 1968. Etapy razvitia rannekamennougolnoi fauny severnoi chasti Magnitiogorskogo sinclinoria. [Stages of development of the Early Carboniferous fauna from the northern part of the Magnitogorsk Synclinorium]. In Krimgoltz, G.Ya. (ed.), Laws of the organic world development according to paleontology, Proceedings of XII session of the VPO. Nauka, Leningrad, 51-63. [In Russian].
85Dybowski, W. N., 1873. Monographie der Zoantharia Sclerodermata Rugosa aus der Silurformation Estlands, Nord-Livlands und der Insel Gotland. Archiv für Livlands, Estlands, Kurlands, 1, 257–414.
86Fomichev, V.D., 1953. Korally Rugosa i stratigrafiya sredne- i verkhnekamennougolnykh i permskikh otlozheniy Donetzkogo basseyna. [Rugose corals and stratigraphy of Middle and Upper Carboniferous and Permian deposits of the Donets Basin]. Trudy VSEGEI, Gosgeolizdat, Moscow, 1-622. [In Russian].
87Hecker, M., 2001. Lower Carboniferous (Dinantian and Serpukhovian) rugose coral zonation of East European Platform and Urals, and correlation with Western Europe. Bulletin of the Tohoku University Museum, 1, 298–310.
88Hill, D., 1981. Coelenterata; Supplement 1, Rugosa and Tabulata. In: Teichert, C. (ed.), Treatise on invertebrate paleontology. The Geological Society of America (Boulder, Colorado) and the University of Kansas (Lawrence, Kansas), 762 p.
89Ivanovski, A.B., 1967. Etyudy o rannekamennougolinikh rougozakh [Study on Lower Carboniferous rugose corals]. Nauka, Moscow, 92 p. [In Russian].
90Kachanov, E.I., 1965. Rol’ corallov v razrabotke biostratigraficheskoy schemy nizhnekamennuugolnykh otlozheniy vostochnogo sklona Urala [The coral’s role in the development of the Lower Carboniferous biostratigraphic scheme of the Southern Urals Eastern slope]. In Sokolov, B.S. & Dubatolov, V.N. (eds), Devonian and Carboniferous Tabulatomorph corals of the U.S.S.R. Nauka, Moscow, 91-98. [In Russian].
91Keller, N.B., 1959. Novye nizhnekamennougolnye chetyriokhluchevye korally Dzhezkazganskogo rayona (Kazakhstan) [New Lower Carboniferous tetracorals of the Dzhezkazgan region (Kazakhstan)]. Paleontological Journal, 4, 90-99. [In Russian].
92Lipnjagov, O.M., 1975. The Conodonts of C1ta and C1tb of the Donetz Basin. Belgian Geological Survey Professional papers, Bruxelles, 161, 41-47. [In Russian and English].
93Mc Coy, F., 1844. A synopsis of the characters of the Carboniferous Limestone fossils of Ireland. University Press, Dublin, 5-207.
94Mc Coy, F., 1849. On some new genera and species of Palaeozoic corals and foraminifera. The Annals and Magazine of Natural History, ser. 2, 3, 1–20, 119–136.
95Menning, M., Alekseev, A. S., Chuvashov, B.I., Davydov, V.I., Devuyst, F.-X., Forke, H.C., Grunt, T.A., Hance, L., Heckel, P.N., Izokh, N.G., Jin, Y.-G., Jones, P.J., Kotlyar, G.V., Kozur, H.W., Nemyrovska, T.I., Schneider, J.W., Wang, X.-D., Weddige, K., Weyer, D. & Work, D.M., 2006. Global time scale and regional stratigraphic reference scales of Central and West Europe, East Europe, Tethys, South China, and North America as used in the Devonian-Carboniferous-Permian Correlation Chart (DCP 2003). Palaeogeography, Palaeoclimatology, Palaeoecology, 240, 318-372.
96Münster, G., 1840. Die Versteinerungen des Uebergangkalkes mit Clymenien und Orthoceratiten von Oberfranken. Beiträge zur Petrefacten-Kunde, Bayreuth, 3, 33-121.
97Ogar, V.V. 2010. New data on the Carboniferous corals of the Donets Basin. Paleoworld, 19/3-4, 284-293.
98Perna, A. Y., 1923. Korally iz nizhnekamennougolnykch otlogeniy vostochnogo sklona Yuzhnogo Urala [Rugose corals of the Lower Carboniferous on the southern slope of the South Urals]. Trudy Geologicheskjgo Komiteta, Novaya seria, 175, 3-34. [In Russian].
99Poletaev, V.I., Brazhnikova, N.E., Vassilyk, N.P. & Vdovenko, M.V., 1990. Local zones and major Lower Carboniferous biostratigraphic boundaries of the Donets Basin (Donbass), Ukraine, U.S.S.R. Courier Forschungsinstitut Senckenberg, 130, 47-59.
100Poletaev, V.I., Vdovenko, M.V., Shchogolev. O.K., Boyarina, N.I. & Makarov, I.A., 2011. Stratotypy regionalnych stratygrafichnych pidrozdiliv karbonu i nyznoi permi Dono-Dniprovskogo prohinu [The stratotypes of the regional stratigraphic subdivisions of the Carboniferous and Lower Permian of the Don-Dnieper Depression]. Logos, Kyiv, 236 p. [In Ukrainian].
101Poty, E., 1975. Un nouveau Tétracoralliaire du Viséen moyen de la Belgique: Corphalia mosae Gen. et sp. nov. Annales de la Société Géologique de Belgique, 98/1, 111-121.
102Poty, E., 1981. Recherches sur les tétracoralliaires et les hétérocoralliaires du Viséen de la Belgique. Mededelingen Rijks Geologische Dienst, 35/1, 1-161.
103Poty, E., 1982. Deux nouvelles espèces de Tétracoralliaire du sondage de Kastanjelaan -2 à Maastricht, Pays-Bas. Natuurhistorisch Maandblad (Orgaan van het Natuurhistorisch Genootschap in Limburg, Nederland), 71/3, 54-58.
104Poty, E., 1999. Famennian and Tournaisian recoveries of shallow water Rugosa following late Frasnian and late Strunian major crises, southern Belgium and surrounding areas, Hunan (South China) and the Omolon region (NE Siberia). Palaeogeography, Palaeoclimatology, Palaeoecology, 154, 11-26.
105Poty, E. & Boland, K., 1994. Révision des tétracoralliaires caninomorphes de l’Hastarien (Tournaisien) belge. Annales de la Société Géologique de Belgique, 117, 201-225.
106Poty, E. & Onoprienko, Y., 1984. Rugose corals. In Shilo, N.A., Bouckaert, J., Afanasjeva, M.J.M., Bless, M.J.M., Conil, R., Erlanger, O.A., Gagiev, M.H., Lazarev, S.S., Onoprienko, Y., Poty, E., Razina, T.P., Simakov, K.V., Smirnova, L.V., Streel, M. & Swennen, R. Sedimentological and palaeontological atlas of the Late Famennian and Tournaisian deposits in the Omolon Region (NE USSR). Annales de la Société Géologique de Belgique, 107, 200-213.
107Poty, E., Berkowski, B. Chevalier, E. & Hance, L., 2003. Biostratigraphic and sequence stratigraphic correlations between the Dinantian deposits of Belgium and Southern Poland (Krakow area). In Wong, T. E. (ed.), Proceedings of XVth International Congress on Carboniferous and Permian Stratigraphy. Royal Netherlands Academy of Arts and Sciences, Utrecht, 97-106.
108Poty, E., Devuyst, FX. & Hance, L., 2006. Upper Devonian and Mississippian foraminiferal and rugose coral zonations of Belgium and Northern France, a tool for Eurasian correlations. Geological Magazine, 143/6, 829-857.
109Poty, E., Aretz, M. & Hance, L., 2014. Belgian substages as a basis for an international chronostratigraphic division of the Tournaisian and Viséan. Geological Magazine, 151/2, 229-243.
110Sando, W.J. & Bamber, E.W., 1985. Coral zonation of the Mississippian System in the Western Interior Province of North America. U.S. Geological Survey Professional Paper, 1334, 61 p.
111Sibly, T.F. 1906. On the Carboniferous Limestone (Avonian) of the Mendip Area (Somerset) with special reference to the palaeontological sequence. Quarterly Journal of the Geological Society of London, 62, 324-380.
112Somerville, I.D., Mitchell, M. & Strank, A.R.E., 1986. An Arundian fauna from Dyserth area, North Wales and its correlation within the British Isles. Proceedings of the Yorkshire Geological Society, 46, 57-75.
113Vassilyuk, N.P., 1960. Nizhnekamennougol’nye korally Donetskogo basseyna [Lower Carboniferous corals of the Donets Basin]. Akademia Nauk Ukrainskoi SSR, Trudy Instituta Geologicheskikh Nauk, Seria Stratigrafia i Paleontologia, Kiev, 13, 1-179. [In Russian].
114Volkova, M. S., 1941. Nizhnekamennougol’nye korally Central’nogo Kazachstana [Lower Carboniferous corals from Central Kazakhstan]. Materials on the Geology and Useful Minerals of Kazakhstan, Gosgeolizdat, Moscow, Leningrad, 11, 1-120. [In Russian and English].
115Wang, H.C., 1950. A revision of Zoantharia Rugosa in the light of their minute skeletal structures. Philosophical Transactions of the Royal Society, London, 61/2-3-4, 175-246.
116Wang, Z. & Zhao, Z., 1987. Early Tournaisian tetracorals from the Hobok River Formation, Western Junggar, Xinjiang. Geological Review, 33/5, 479-483. [In Chinese with English abstract].
117Weyer, D., 1993. Korallen im Untertournai-Profil von Drewer (Rheinisches Schiefergebirge). Geologie und Paläontologie in Westfalen, 29, 177-221.
118Weyer, D., 2000. Korallen im Unterkarbon Deutschlands. Abhandlungen und Berichte für Naturkunde, 23, 57-91.
119Manuscript received 22.02.2016, accepted in revised form 04.03.2016, available on line 04.07.2016.
To cite this article
