


- Startpagina tijdschrift
- Volume 25 (2022)
- number 3-4
- Functional morphology, coaptation and palaeoecology of Hollardops (Trilobita, Acastidae), with descriptions of new species and two new genera from the Devonian of Morocco


Weergave(s): 6067 (32 ULiège)
Download(s): 2881 (4 ULiège)

Functional morphology, coaptation and palaeoecology of Hollardops (Trilobita, Acastidae), with descriptions of new species and two new genera from the Devonian of Morocco

Documenten bij dit artikel
Bijlagen
Abstract
A group of asteropygine trilobites with 10 thoracic segments from the lower Emsian to lowermost Eifelian of western Europe and northwest Africa is deemed monophyletic. Available names for this clade are Hollardops Morzadec, 1997, Philipsmithiana Lieberman & Kloc, 1997, Modellops Lieberman & Kloc, 1997 and Pennarbedops Bignon & Crônier, 2013, the first of which has priority. Well-preserved Hollardops specimens from southern Morocco have revealed previously undescribed details of the mineralised exoskeleton that are interpreted here as coaptative and sensory devices. It is proposed that Hollardops, like many other asteropygines, practised a dual mode of coaptation: fully locked enrolment was alternated with a retracted pygidium, allowing temporary influx of oxygenated seawater and excretion through slits between the pygidial lappets. This putative ‘breathing device’ enabled the trilobite to remain enrolled for a longer period of time while maintaining vital body functions. A poor understanding of the problematic type species of Hollardops, H. mesocristatus (Le Maître, 1952), has clouded actual diversity in Algeria and Morocco. Hollardops klugi sp. nov. is the oldest known member of the genus and extends its confirmed stratigraphic range into the lower Emsian. Two species, H. kyriarchos sp. nov. and H. multatuli sp. nov., are recorded from just above the base of the upper Emsian. Additionally, H. angustifrons sp. nov., H. luscus sp. nov. and the first well-preserved specimens of H. boudibensis Morzadec, 2001 are recorded from the upper Emsian. The types of H. hyfinkeli (Lieberman & Kloc, 1997) and H. burtandmimiae (Lieberman & Kloc, 1997) are refigured and original species concepts reiterated. Feruminops Haas, 1968, including its junior subjective synonym Morzadecops Bignon & Crônier, 2013, from the lower Emsian of Morocco and Türkiye, may comprise the sister group of Hollardops. Additionally, the enrolment strategies and systematics of several other members of Asteropyginae are discussed. Platykardiapyge gen. nov. (type species: Metacanthina maderensis Morzadec, 2001) is erected for a group of Pragian–early Emsian asteropygines from Morocco, Spain and Türkiye with a widely heart-shaped pygidium and comparatively many pygidial pleurae, among other features. Bignonops gen. nov. (type species: Kayserops tamnrhertus Chatterton et al., 2006) is erected for some species previously included in Gandlops Bignon & Crônier, 2013. Minicryphaeus suavius sp. nov. is described from the lower Emsian of Morocco. The identity and generic affinity of the oldest known asteropygine, Ganetops gdoumontensis (Asselberghs, 1930) from the Pridolian of Belgium, are discussed.
Inhoudstafel
1. Introduction
1In 1997 Morzadec erected Hollardops for asteropygine trilobites with 10 thorax segments (as against the usual 11 for this subfamily) and selected Asteropyge mesocristata Le Maître, 1952 from the upper Emsian of Algeria as the type species. Hollardops species have hitherto been recognised in the Devonian of Algeria, Morocco and France (Morzadec, 1969, 1983; Lieberman & Kloc, 1997; Schraut, 1998, 2000; Lebrun, 2002, 2018; Jansen et al., 2004; Chatterton et al., 2006; Bonino & Kier, 2009; Chatterton & Gibb, 2010; Brett et al., 2012; Laurencin & Hoffman, 2012; Hoffman et al., 2014; Lawrance & Stammers, 2014; Levi-Setti, 2014; Khaldi et al., 2016; Schoenemann et al., 2017; Crônier et al., 2018). Subsequent to the papers of Morzadec (1997, 2001) there has been wide consensus that, besides the two endemic species H. lemaitreae Morzadec, 1997 and H. boudibensis Morzadec, 2001, diversity throughout the upper Emsian in Algeria and Morocco is accounted for by H. mesocristatus and H. burtandmimiae (Lieberman & Kloc, 1997).
2We have examined numerous well-preserved Hollardops specimens from various sections in Morocco, which led us to the viewpoint that current species concepts are in need of a critical reappraisal and that the diversity of the genus is higher than previously recognised.
2. Localities (Morocco)
3Hollardops is regularly found in upper Emsian strata in Morocco but only in certain limestone and marly levels. Commercially mined specimens have been offered for sale at least since the 1990s. Their profusion suggests that they were mined in localities frequented for their high commercial leverage, i.e., where comparatively rare trilobite species occur as well-preserved articulated exoskeletons relatively often. Unfortunately, the exact geographic and stratigraphic origins of Hollardops specimens offered for sale are almost never known and preparation is often substandard. Newly collected specimens are recorded from various sections in the Tindouf and Ma’der basins and the Tafilalt Platform (sensu Wendt, 2021), as well as others in extant collections of imprecise origin, in Morocco. Reference is made to Bultynck & Walliser (2000) for updated stratigraphic frameworks of the Ma’der Basin and the Tafilalt Platform subsequent to the principle work of Hollard (1974), and to Becker et al. (2004a) for the northern limb of the Tindouf Basin.
4Bou Tserfine locality of Becker et al. (2004c), east of Assa (Figs 1, 2); Khebchia Formation, upper Emsian. The base of the Khebchia Formation comprises a 5–6 m-thick interval of rhythmically bedded limestone-shale successions described by Becker et al. (2004a) as the Hollardops Limestone Member. This unit is a useful regional marker in the western Draa Valley, recognisable by the deposition of the first bioclastic solid limestone bed in dark grey marls. Abundant fossils occur in the limestones and marls of the Hollardops Limestone Member in various orientations to bedding; Brett et al. (2010, 2012) suggested that the animals were entrained in mudflows. Trilobites are often found articulated, uncompacted and well-preserved, as a result of rapid burial and early diagenetic cementation. Shaly intervals, barren in fossils and showing extensive bioturbation, indicate episodes of sediment starvation. The age of the Hollardops Limestone Member is not known exactly but an early upper Emsian age (probably laticostatus Zone of Bultynck & Hollard, 1980) is inferred from biostratigraphical evidence (Bultynck & Hollard, 1980; Becker et al., 2004a; Jansen et al., 2007; Dojen et al., 2011; van Viersen & Holland, 2016). A gradual change from neritic to pelagic sedimentation across the boundary between the Oui-n-Mesdoûr and Khebchia formations in the region has been suggested to reflect the increasing influence of the global transgressive Daleje Event (Becker et al., 2004b).
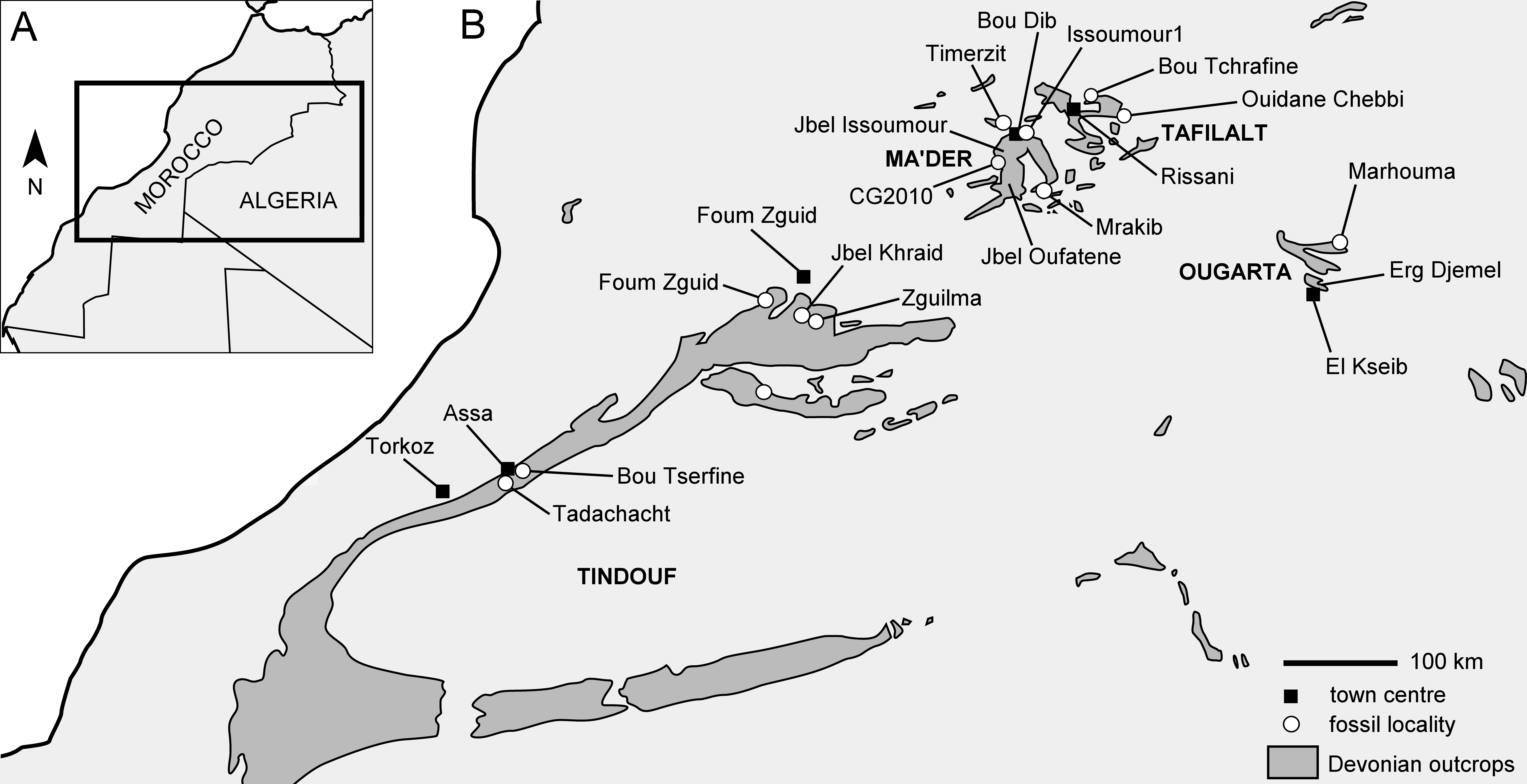
Figure 1. Simplified map of southern Morocco and western Algeria showing Devonian outcrops, fossil localities and town centres in the Tindouf, Ma’der and Ougarta basins, and the Tafilalt. Adopted from Hollard (1967, p. 204, fig. 1) and modified.
5Hollardops kyriarchos sp. nov. co-occurs with Morocops spinifer van Viersen et al., 2017 (Fig. 3C–E) in a marly layer just above the base of the Hollardops Limestone Member in Bou Tserfine (van Viersen et al., 2017, fig. 2a; Fig. 2). Van Viersen et al. (2017, fig. 2a) showed Hollardops mesocristatus to occur in the Hollardops Limestone Member starting just above Morocops spinifer, based on the accounts of Becker et al. (2004c) and Brett et al. (2012). One of us (AV) has examined high-resolution photographs of the original specimens from Bou Tserfine in the Westfälische Wilhelms-Universität Münster (courtesy of Dr Stephan Helling), concluding that those do not belong to H. mesocristatus and that they may represent one or more distinct species (“Hollardops” in our Fig. 2).
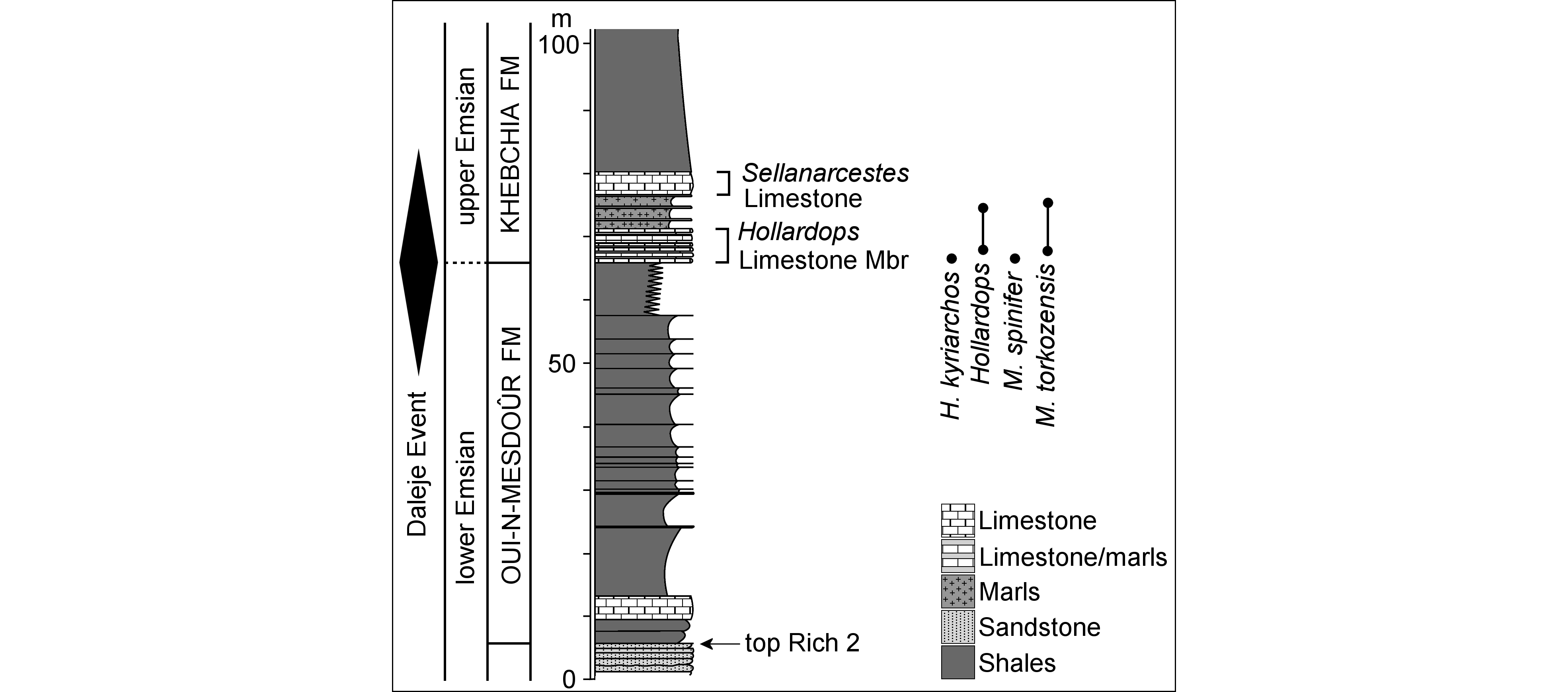
Figure 2. Lithostratigraphy and trilobite biostratigraphy in the Bou Tserfine section, modified and simplified from Becker et al. (2004c) and van Viersen et al. (2017).
6Tadachacht locality of van Viersen & Holland (2016), south of Assa (Fig. 1); Khebchia Formation, upper Emsian. A locality log is unavailable but the sequence is very similar to that in Bou Tserfine (van Viersen et al., 2017). Numerous complete, at times slightly disarticulated trilobites occur in a marly bed, informally called Cyphaspis couche, near the summit of the Hollardops Limestone Member. The Hollardops specimens have fine details of the mineralised cuticle preserved, some of which are described here for the first time. We describe Hollardops multatuli sp. nov. from this horizon, where it co-occurs with Cyphaspis lerougei van Viersen & Holland, 2016 (Fig. 3H–N), Cyphaspis tadachachtensis van Viersen & Holland, 2016 (Fig. 3O), Morocops torkozensis (Schraut, 2000) (Fig. 3A, B, Q) and Timsaloproetus gibbae van Viersen & Lerouge, 2019 (Fig. 3F, G).
7Jbel Khraid locality of van Viersen & Holland (2016), south of the village of Foum Zguid (Fig. 1); upper Emsian to Eifelian Timrhanrhart Formation. Two trilobite-bearing levels were identified during fieldwork carried out by the late H. Prescher (AV, pers. comm. with H. Prescher in 2014). A lower bed, which Prescher considered to be an equivalent of ZGEE2 (upper Emsian) in the nearby Zguilma section of Chatterton et al. (2006), has yielded Acastoides zguilmensis Chatterton et al., 2006, Koneprusia dahmani Chatterton et al., 2006, Walliserops trifurcatus Morzadec, 2001, Cyphaspis khraidensis van Viersen & Holland, 2016 and abundant Morocops granulops Chatterton et al., 2006. We record Hollardops angustifrons sp. nov. from this level. An upper bed, correlated by Prescher with ZGEE3 (lower Eifelian) in Zguilma, has yielded Acastoides zguilmensis, Austerops sp., Cyphaspis sp., Diademaproetus mohamedi Chatterton et al., 2006, Gerastos cf. marocensis Chatterton et al., 2006 and Thysanopeltis sp.
8Ouidane Chebbi locality of Klug et al. (2008), east of Rissani (Fig. 1); claystones and marls in the lower Emsian part of the Pragian to mid-Emsian Seheb El Rhassel Group (Bultynck & Walliser, 2000). Klug et al. (2008) studied the macrofauna of two lower Emsian intervals, yielding a lower Faunule 1 and a slightly younger upper Faunule 2. These correspond respectively to the “Marnes et calcaires” and “Calcaires et marnes” in the Seheb El Rhassel group (Tafilalt) of Hollard (1981, table 4). The lower unit consists of grey claystones and marlstones and belongs to the kitabicus Conodont Zone. Faunule 1 comprises an abundant macrofauna, including mainly relatively small invertebrates, abundant acanthodians, rare placoderms, and two fairly common species of asteropygine trilobites described here as Hollardops klugi sp. nov. and Minicryphaeus suavius sp. nov. Klug et al. (2008) noted similarities between faunules 1 and 2 but also an increase of nektonic taxa at the expense of the infauna in the latter. They tentatively referred this turnover to poorly oxygenated conditions on and in the sea floor while adding that it may also reflect other abiotic changes in the general structure of global marine ecosystems during this time. Faunule 2 does not include any asteropygine.
9Jbel Issoumour1 locality of van Viersen & Holland (2016), near Bou Dib (Issoumour1 in Fig. 1); lower part of the Tazoulaït Formation, upper Emsian. A single specimen of Hollardops boudibensis was obtained from a level just below the Psychopyge couche (see Chatterton & Gibb, 2010, p. 1189, fig. 1C), where it co-occurs with Cyphaspis kippingi van Viersen & Holland, 2016.
10Timerzit locality of van Viersen & Lerouge (2019) (Fig. 1); lower part of the El Otfal Formation, upper Emsian. A Walliserops couche has been mined for years in this locality. Van Viersen & Lerouge (2019) inadvertently placed the Walliserops couche in the Tazoulaït Formation. This horizon belongs to the basal part of the overlying El Otfal Formation instead. Trilobites include Acastoides sp., Adrisiops sp., Cyphaspis sp., Koneprusia sp., Leonaspis sp., Psychopyge sp., a cornuproetine, a scutelluid, a harpetid, several species of Walliserops (AV, pers. comm. with H. Prescher in 2007), Timsaloproetus pulchistriatus pulchistriatus van Viersen & Lerouge, 2019 and Hollardops luscus sp. nov.
3. Palaeoecology of trilobites from the basal Khebchia Formation in the western Draa Valley
11The interpretation of the trilobite-bearing beds in the Hollardops Limestone Member as obrution deposits implies that alpha diversity of the benthic macrofauna is largely original. Becker et al. (2004c) reported orthocones, nautiloids, corals, gastropods, styliolinids, ostracods, rare atrypids and other brachiopods. Trilobites (Fig. 3) include Hollardops, Morocops, Tropidocoryphe (two undetermined species), Timsaloproetus gibbae, Diademaproetus cf. praecursor (Alberti, 1969), Gerastos cf. marocensis, Rheiscutellum sp., Cyphaspis tadachachtensis, Cyphaspis lerougei, Leonaspis sp., Koneprusia sp. and Psychopyge sp. (Becker et al., 2004c; Brett et al., 2012; van Viersen & Holland, 2016; van Viersen et al., 2017; van Viersen & Lerouge, 2019; AV, pers. comm. with D. Holland and H. Prescher in 2014). At the genus level, this trilobite fauna is characteristic of the upper Emsian of the Rhenish-Bohemian Province (sensu Dowding & Ebach, 2019), where closely allied taxa in Acastidae, Phacopidae, Odontopleuridae, Proetidae, Scutelluidae and Aulacopleuridae, were widely distributed along the continental shelves of southern Laurussia and northern and peri-Gondwana, possibly owing their dispersion to the effects of the transgressive Daleje Event (Morzadec, 1992; Chlupáč, 1994; Chlupáč et al., 2000; van Viersen & Holland, 2016; van Viersen et al., 2017).
12Of note is the absence of trochurines, although this is generally the case in mid-Emsian strata (⁓ nothoperbonus–inversus Conodont zones) worldwide. In the Rhenish-Bohemian Province, the only potential exceptions known from this interval are referred to Huginarges from the Rhenish Mountains (Basse & Müller, 2004; Franke, 2010; van Viersen, 2021b). Huginarges is widely distributed in the Pragian of southern Morocco and reappears in the uppermost Emsian at Issoumour (Chatterton & Gibb, 2010; Corbacho & Kier, 2013; Lebrun, 2018). Jasperia from New South Wales is the only other confirmed mid-Emsian trochurine (Chatterton, 1971; Edgecombe & Wright, 2004) and Terranovia may occur in mid-Emsian strata in the Canadian Arctic (Yolkin & Ormiston, 1985); neither of these has invaded the Rhenish-Bohemian Province.

Figure 3. Trilobites from the Hollardops Limestone Member, Khebchia Formation (upper Emsian), in the western Draa Valley. A, B, Q. Morocops torkozensis (Schraut, 2000). A, B. NHMM 2016002, Tadachacht. Q. Paratype, SMF 53627, Torkoz. C–E. Morocops spinifer van Viersen, Holland & Koppka, 2017, holotype, NHMM 2016004, Bou Tserfine. F, G. Timsaloproetus gibbae van Viersen & Lerouge, 2019, holotype, IRSNB a13423, Tadachacht. H–N, Cyphaspis lerougei van Viersen & Holland, 2016, holotype, IRSNB a13014 (I–L and lower left specimen in H) and paratypes, IRSNB a13015 (M, N and upper articulated specimen in H) and IRSNB a13016 (cephalothorax in H), on a single rock slab, Tadachacht. O. Cyphaspis tadachachtensis van Viersen & Holland, 2016, holotype, IRSNB a13002, Tadachacht. P. Hollardops multatuli sp. nov., holotype, IRSNB a13567, Tadachacht.
Scale bars are 4 mm.
3.1. Modes of life
13The lifestyles of the trilobites of the Hollardops Limestone Member range from sluggish particle feeders to active scavengers or even predators. Van Viersen & Lerouge (2021) discussed the dorsal sculpture of Timsaloproetus which consists of asymmetric terrace ridges organised in a Bertillon pattern. Such sculptural elements in marine invertebrates are generally designed to prevent back slipping during burrowing. Van Viersen & Lerouge (2021) provided arguments to construe Timsaloproetus as a carnivorous scavenger or small predator with a semi-endobenthic life mode. If this notion is correct, the rareness of Timsaloproetus specimens in obrution beds was perhaps in part due to their frequent burrowing behaviour which may also have been a primary maneuver to evade danger.
14According to Fortey & Owens (1999), the impendent hypostome found in Phacopida was associated with forward expanding glabellas, reasonably explained within the context of predatory habits. Stürmer & Bergström (1973) studied the internal anatomy of “Phacops” and noted in particular the well-developed gnathobases of the coxopodites and the fringes of bristles distally on the endopodites. This led them to construe “Phacops” as a raptatory benthic animal, capable of walking on and slow swimming above the sea floor, but not well able to dig. Bruton & Haas (1997) studied the functional morphology of phacopids closely allied with Morocops, inferring a relatively slow gait with a promotor-remotor swing of the leg basis and circular motion of the legs. Bruton & Haas (1997) furthermore noted the long hypostome and the far anteriorly positioned mouth, which led them to suggest that the animal must have been stationary whilst feeding, with the legs being used to stir up the bottom. Bruton & Haas concluded that such phacopids fed on small food particles and that they were detritus feeders. Van Viersen et al. (2017) discussed the spiny tubercles of Morocops spinifer (Fig. 3C–E) which are dissimilar from other spiny phacopids with a more straightforward defensive strategy. They interpreted the spines of M. spinifer as modified tubercles found in congeners, and argued that these supported the displacement of the richly biodetrital substrate. Whatever the purpose of the spines was, they appear to be unique among Phacopinae and designed for a specific ecological niche. Morocops spinifer is confined to a 1 m-thick interval near the base of the Hollardops Limestone Member in Bou Tserfine and was succeeded here by Morocops torkozensis (Fig. 3A, B, Q) with conservative tubercular sculpture.
15Hollardops, with its relief-rich exoskeleton and towering schizochroal eyes, does not strike as a burrower. Bergström & Brassel (1984) noted that the legs of the grossly similar asteropygine Rhenops were not long enough for true digging. The subtriangular cephalon of Hollardops, with its broad, sloping borders, would appear to be ideal for ‘ploughing’ (see Birkenmajer & Bruton, 1971). We postulate that the ventrally deflected anterior border acted as a shovel, breaking the sediment surface and moving downward. The robust, abaxially concave (tr.), subexsagitally running genal spines, which were slightly dorsally higher than the lateral cephalic border, would have guided and disposed of sediment laterally. In this fashion the top layers of the substrate were excavated and the appendages exposed to fresh sediment. At the same time, Hollardops was probably streamlined enough to swim above the sea floor over short distances. We envisage that it was an active scavenger, perhaps even predator, capable of handling fairly large food particles. Bergström & Brassel (1984) previously proposed a possible predatorial lifestyle for Rhenops.
16It is noteworthy that, despite the fact that individuals of Hollardops grew to the largest sizes of trilobites in the Hollardops Limestone Member, they were well equipped for deterring predatorial attacks (see below). Hollardops probably lacked the tools for crushing the cuticle of similar-sized arthropods and this suggests that large holaspides were predated on by other, non-trilobite animals. Predators of trilobites included arthropods, cephalopods, worms, fishes, cnidarians, echinoderms, and enigmatic organisms (Babcock, 1993, 2003). The Devonian witnessed a drastic increase in durophagous predators including fish and cephalopods (Brett & Walker, 2002). Ammonoids have been reported in the basal Khebchia Formation and may have been culprits of attacks on trilobites. As a general statement, the characteristic triangular bite mark of the cephalopod parrot-like beak (assuming that this was universal during the Devonian) has not been identified by us on any of the hundreds of Devonian trilobites from Morocco that we have seen. This suggests that ammonoid attacks on trilobites were probably rare, or often lethal or unsuccessful. However, a sampling bias towards pristine trilobite specimens of comparatively high commercial value cannot be excluded.
17Fortey & Owens (1999) suggested a positive relationship between the numbers of particle-feeding trilobites and amounts of organic particle matter supply in the substrate. This may often be true for late Lower to early Middle Devonian trilobite faunas in western Europe and NW Africa that include abundant eurytopic, small proetide detritivores with a bulbous glabella and natant hypostome (e.g. Pillet, 1973; Šnajdr, 1980; Morzadec, 1983; Smeenk, 1983; Basse, 2002; Chatterton et al., 2006; Gibb & Chatterton, 2010; van Viersen & Holland, 2016; van Viersen, 2021a). The fauna of the basal Khebchia Formation is dominated by abundant individuals of Hollardops and Morocops whereas natant proetides are uncommon (Brett et al., 2012, p. 34, fig. 4; AV, pers. comm. with H. Prescher in 2014). The cosmopolitan Siluro-Devonian Cyphaspis is rare save for a single bed near the summit of the Hollardops Limestone Member, aptly named Cyphaspis couche, in which complete specimens of C. lerougei (Fig. 3H–N) are more regular occurrences (AV, pers. comm. with D. Holland in 2015). This species, with its comparatively large and strongly inflated glabella, was unlikely to be a successful forward burrower or swimmer. The ‘posterior tegimen’ was named by van Viersen & Holland (2016) for the steeply inclined, densely granulose area covering the posterior part of the pygidium and continuing obliquely onto the posterolateral extremities of the thorax of many Cyphaspis species. The anterior margin of the posterior tegimen is marked by a band of tubercles. Some species with a well-developed posterior tegimen, including C. lerougei, have the pygidium drastically reduced in terms of size and segmentation such that the legs were probably no longer fully functional. In the putative feeding position, the large genal spines of C. lerougei lay horizontal and the thorax was posteriorly weakly ventrally inclined to seabed (Fig. 4). The extremely vaulted (sag.) pygidium was aligned with the steep thoracic pleurae so that the entire posterior tegimen was dug in, its slopes standing perpendicular to the sea floor. The cephalon retained a horizontal position, stabilised on the sea floor by the long, robust genal spines. As the appendages stirred up the substrate and moved particles anteriorly towards the mouth, the pygidium was used for guiding and stabilisation. The dorsally convex posteromedian pygidial border seems to be a coaptative device. Enrolment was likely a principle defensive strategy for Cyphaspis against predators and other threats. Members of the Cyphaspis hamidi group (sensu van Viersen & Holland, 2016) which includes C. lerougei, were unable to align the cephalic doublure with the much smaller pygidium during enrolment (see, e.g., Basse & Müller, 2016, pl. 3, fig. 27, pl. 4, fig. 35). Instead, the area of the pygidium and the thorax covered by the thick-shelled posterior tegimen, with its steep, granulose surface and tubercular anterior band increasing friction, was embedded in the cephalic cavity (see, e.g., Chatterton et al., 2006, pl. 46, figs 1, 5–9). Many Cyphaspis species have a large dorsal spine on the sixth thoracic segment and some have additional smaller ones on the glabella and the occipital ring; their presence varies even in closely allied species (van Viersen & Holland, 2016). Although in some species the thoracic dorsal spine was long and weakly dorsally inclined so that it could be argued to have supported a horizontal position of the trilobite on the sea floor, in other species the spine would have been too short for this. The function of the dorsal spines may have been manifold although it was probably primarily a defensive one.
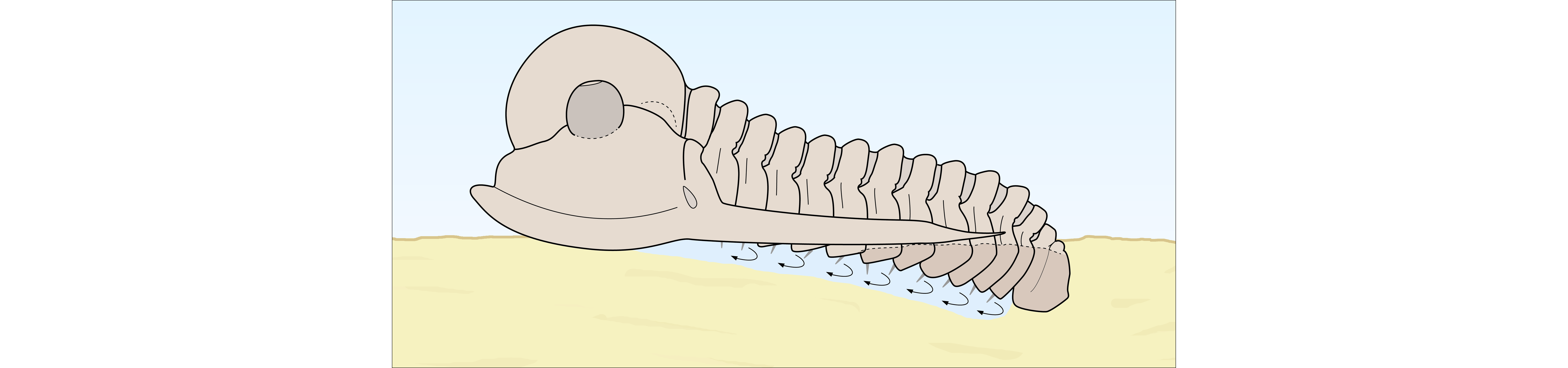
Figure 4. The otarionine trilobite Cyphaspis lerougei in putative feeding mode.
3.2. Pathologies
18Pathologies are shown by two of the trilobite specimens from the Hollardops Limestone Member studied. The exceedingly large paratype NHMM 2022001 of Hollardops multatuli shows two neoplasms on the left post-cephalic pleural region. The largest of these is located posterolaterally on the dorsal surface of the first pygidial pleural lappet, where it is recognisable as a distinct gall-like swelling of lighter colour (Figs 5A, D, G, 6D, G, I, 7M, 18K). A second, smaller neoplasm, similar in shape and location, is present on the ninth thoracic pleural lappet (Figs 6D, G, 7M, 18K). Such pathologies were likely caused by hyperplasia (Babcock, 1993). Their origins remain unknown and may be parasitic. If so, the culprit appears to have had a preference for the extremities of the hollow pleural lappets. Extant crustaceans can lodge a range of parasites, including other arthropods, dinoflagellates, fungi, yeasts, bacteria and viruses; some of these are known to affect the carapace (Trilles & Hipeau-Jacquotte, 2012).
19Vermiform, putative drilling holes are present on the same Hollardops multatuli individual, respectively on the left side of the anterior glabellar lobe (Figs 6A, B, 7J) and on the fourth left thoracic pleura (Figs 6H, 7K). Both holes are partially healed by lateral convergence of tissue, indicating that the attacks took place while the animal was alive and that they were not lethal (at least not instantly).
20A complete specimen of Timsaloproetus (Fig. 3F, G) shows a dorsally warped right anterolateral cephalic margin of aberrant outline. Just posterior of the affected margin, the lirae are effaced left and right of the preocular suture. This appears to be a partially healed, sublethal bitemark. Most examples of such healed injuries in trilobites are referred to as compensatory hypertrophy (Babcock, 1993).

Figure 5. Hollardops multatuli sp. nov., Cyphaspis couche, Hollardops Limestone Member of the Khebchia Formation, Tadachacht, Morocco. Paratype, NHMM 2022001, in dorsal view (A), close-up of left eye (B), close-up of librigena lateral of left eye (C), lateral view (D), close-ups of anterolateral cephalic border (E, F) as indicated in D, posterior view (G), close-up of left genal spine (H).
Scale bars are 10 mm.
3.3. Epizoic growth
21Epizoans on Devonian trilobites have rarely been reported in the literature; they include anthozoans, annelids and bryozoans (see Basse & Müller, 2016 for a review). Kloc (1997) reported encrusting organisms on Devonian odontopleurids and divided these into three types: (1) fossilised hard parts of bryozoans, brachiopods, corals, crinoids, foraminiferans and worm tubes, (2) endoliths of unknown origin that may represent algae and/or sponges, and (3) pyrite (limonitised) coating the spines, apparently formed from the decay of organic matter from an unknown encrusting organism. Some epizoans preferred trilobite sclerites (post-mortem or exuviae) as hosts. Zapalski & Klug (2018) reported tabulate corals on a phacopid cephalon and a harpetid cephalon (the latter appears to be a rotated proetine pygidium instead) from the Devonian of the Tafilalt. A study of Ordovician epibenthic bryozoans by Kácha & Šarič (2009) has revealed a host preference for cephalopod shells and conulariid exoskeletons over loose trilobite sclerites and brachiopods, and that when trilobites were encrusted, the central part of the cephalon was preferred.
22Epizoic growth may also have occurred on living trilobites but examples of this type are scarce (e.g. Alberti, 2014). Brandt (1996) listed as possible causes for the loss of epizoic growth in the fossil record, the opportunistic nature of the epizoan’s settlement and survival, preservation artifacts, and the life habit of the trilobite. Waugh et al. (2004) noticed that epizoans are commoner on living marine decapod crustaceans than on fossils, which they primarily attributed to a preservational bias. According to Basse & Müller (2016) most encounters on trilobites occur in pelitic facies and rarely to never in cephalopod limestones or carbonate facies, which seems to imply that the occurrence of epizoans was environmentally controlled. However, this is almost certainly also due to an observational bias since any traces of epibiosis can be expected to be more easily recognisable on the external moulds of decalcified fossils. For instance, the majority of Devonian trilobites from Morocco are found complete in limestones, recognised by miners by their distinctive cross sections. Only a fortunate strike of the hammer exposes areas of the dorsal cuticle, thus greatly limiting chances of detecting epizoic growth. In fact, such erratic structures of increased surface area should be expected to rupture often and the distinctive dorsal surface would be retained in the external mould, making it difficult to be recognised. Mechanical preparation of these trilobites with an air scribe is common practise and easily leads to the destruction of any fouling before its presence is noticed.
23The apparent rareness of epizoans on Devonian trilobites from Morocco does not imply that they did not occur regularly in life and it is tempting to reflect on the relationships with their hosts. Kloc (1997) reported epibionts on various spines of odontopleurids from the Devonian of North America which he postulated could have been beneficial by making the trilobite’s outline less conspicuous to predators. Similar putative examples of symbiosis, on the spines of the odontopleurid Radiaspis and the asteropygine Psychopyge have been reported from the Rhenish Mountains (e.g. Basse in Basse & Müller, 2004, p. 131, pl. 49, figs 624, 625; Basse & Müller, 2016, pl. 15, figs 140, 141). Brandt (1996) drew analogies with extant decorator crabs which actively attach algae, hydroids and bryozoans to their dorsal carapace, providing effective camouflage. Müller (2005) suggested that the blunted extremities of the dorsal spines of the homalonotid Spiniscabrella were ineffective against predators and that they may have been suitable spots for algae to grow on instead.
24Epizoans are far from always favourable to their hosts. Epizoic growth on contemporary crustaceans has been shown to decrease swimming or burrowing capabilities and can even lead to significantly increased casualties from predation (e.g. Bauer, 1981; Hidalgo et al., 2010). Grooming, i.e., preening and cleaning of various parts of the body involving secondary modification of locomotory or feeding appendages, is a frequent and important strategy adopted by a range of crustaceans (Bauer, 2013). Accepting current models of the biramous trilobite appendages known from certain taxa as ubiquitous within the class, grooming of the dorsal surface of the exoskeleton can be ruled out for the vast majority of trilobites because the appendages would not have been able to reach most dorsal areas. Other antifouling behaviours such as burrowing, frequent ecdysis and a nocturnal mode of life, have been shown to be effective in lowering occurrences of epibionts, e.g. in living brachyurans (e.g. Becker & Wahl, 1996; Bauer, 2013). However, in fossils, this is very difficult to prove and the absence or presence of epizoic growth on trilobite exoskeletons cannot be taken to evidence a certain lifestyle. The dorsal exoskeleton of Hollardops is covered with pits and pitted tubercles that were probably the exit points of setae. Different functions are construed for setae depending on their locations. On the majority of the surface area setae may have helped to reduce specific types of epibiosis as has been demonstrated, e.g., in extant marine arthropods (e.g. Key et al., 1997, 2010).
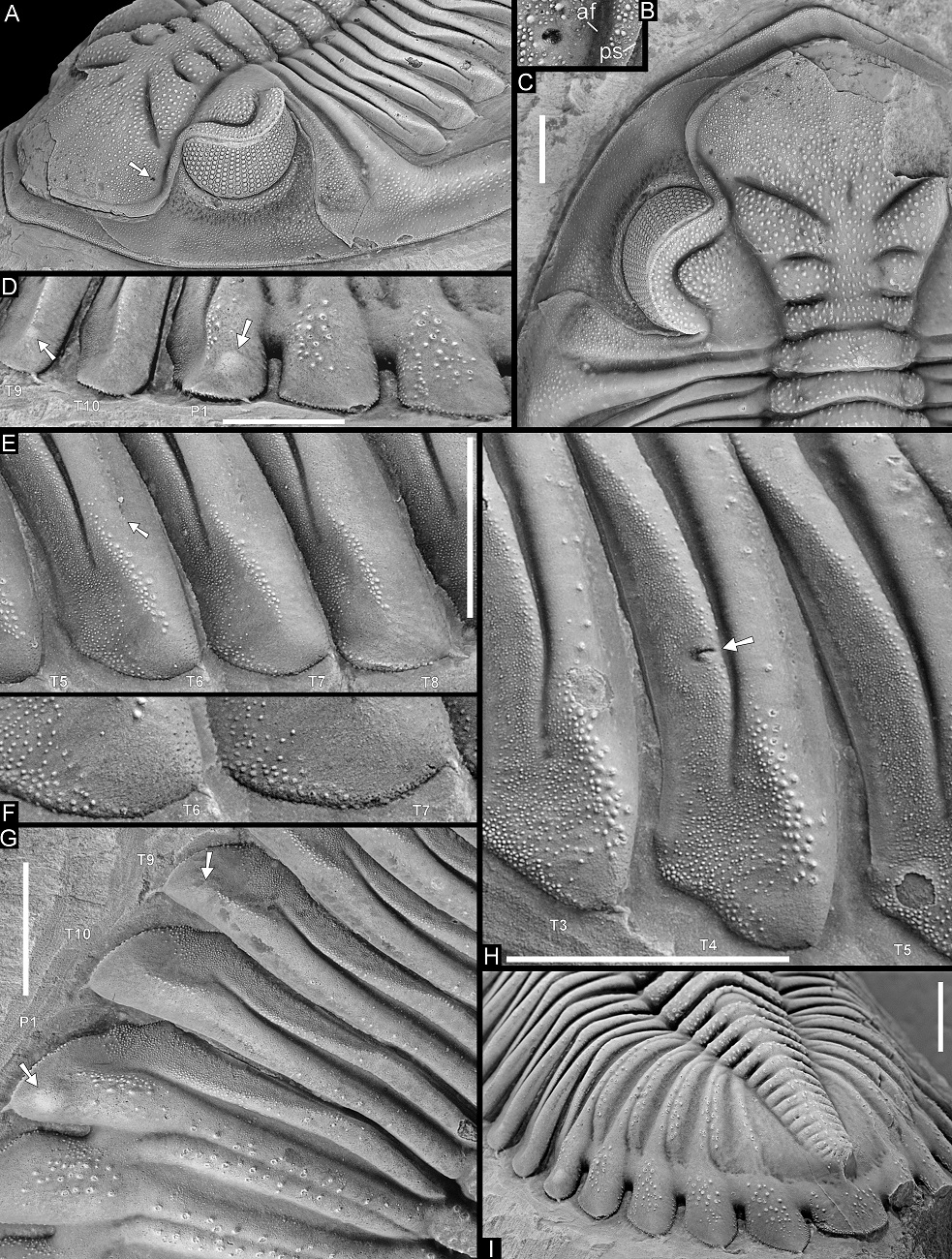
Figure 6. Hollardops multatuli sp. nov., Cyphaspis couche, Hollardops Limestone Member of the Khebchia Formation, Tadachacht, Morocco. Paratype, NHMM 2022001, in anterolateral view (A), close-up of vermiform hole, left on anterior glabellar lobe (B), dorsal view of cephalon (C), oblique lateral view of thorax and pygidium (D), oblique lateral view of left side of thorax (E), close-up of thoracic pleurae (F), dorsal view of left part of thoracopygidium (G), oblique lateral view of left side of thorax (H), oblique posterolateral view of pygidium (I).
Arrows in A and H indicate healed injuries. Arrows in D and G indicate neoplasms. Arrow in E indicates abaxial extremity of furrow dividing convex anterior and weakly vaulted posterior parts of posterior pleural band. Abbreviations in B: af – axial furrow, ps – preocular suture. Scale bars are 5 mm.
4. Functional morphology of Hollardops
4.1. Marginal pit and tubercle bands
25The dorsal sculpture of Hollardops consists of ubiquitous, small to moderately small tubercles and granules of varying size and spacing. Additionally, some specimens allow the observation of two conspicuous bands of close-set tubercles, respectively running submarginally and supramarginally along the cephalic border and onto the base of the genal spine (Figs 5D–F, 9M, 18D). The large paratype of H. multatuli shows that on the distal part of the genal spine, the tubercles are more sawtooth-like (Fig. 5H). Arrays of homologous, minuscule thorns or pointed tubercles, in places abraded as a side effect of preparation (compare, e.g., Fig. 7M and 7N), occur on the anterolateral to lateral margins of the thoracic pleurae and on the entire margins of the pygidial median and pleural lappets of H. multatuli (Figs 5D, G, 6D–I, 7M, N, 19M). Thorny margins of the pygidial lappets are also shown by H. angustifrons (Fig. 20H) and H. luscus (Fig. 21G). In between the tubercle bands of the large paratype of H. multatuli there is a dorsoventrally narrow surface which is devoid of sculpture and carries small pits that verge to forming a single row. The pits are only well discernible on the cephalic border (Fig. 5C, E, F) but the presence of dual tubercle bands on the thorax and the pygidium suggests that pits are also present there. A specimen of H. boudibensis clearly shows dual tubercle bands on cephalon, thorax and pygidium alike (Fig. 9M) and these probably enclosed the same rows of pits. Such structures in living and fossil arthropods are generally regarded as the exit points of cuticular canals that housed setae (Miller, 1976) (see, e.g., Eldredge, 1973, fig. 20F). The distribution of the pits horizontally along the fringe of the exoskeleton strongly suggests that they fulfilled a sensory function. We postulate that the setae formed a system that may have allowed the trilobite to detect and localise movement in its surroundings, shifting directions of currents, and perhaps even chemical changes in the seawater.
26Pit rows around the cephalic and pygidial margins are known to occur in many trilobite groups. In some phacopid and proetid clades, for instance, pits are commonly associated with marginal terrace ridges, although in other species their distribution is universal on the dorsal exoskeleton (e.g. Adrain, 1997; van Viersen et al., 2017; van Viersen, 2021a). Pits are easily lost to diagenetic destruction through calcite filling in crystallographic optical continuity with the cuticle (Teigler & Towe, 1975). Their marginal occurrence in Hollardops makes them particularly prone to damage from abrasion or preparation. This may explain why they have never been noticed before. The tubercle bands marking the inner and outer boundaries of the pitted surface are the only features that are occasionally recognisable in illustrations of published Hollardops specimens (e.g. Chatterton et al., 2006, pl. 14, fig. 1, pl. 18, figs 1, 3), but only if the peripheric matrix was sufficiently removed and photos were taken from low enough angles. From their marginal distribution alone, dual tubercle bands are possibly shown by specimens belonging to Coltraneia (Chatterton et al., 2006, pl. 12, fig. 5), Greenops (Lieberman & Kloc, 1997, p. 92, fig. 24.2; Fig. 14N–P), Walliserops (Chatterton et al., 2006, pl. 27, fig. 2, pl. 28, fig. 2, pl. 30, fig. 2, pl. 31, fig. 4) and the homalonotid Wenndorfia (Chatterton et al., 2006, pl. 32, fig. 7). The original specimens would have to be examined because the photos do not always allow distinctions to be made between tubercle rows and terrace lines. The importance of this distinction might be questioned when considering that terrace lines transitional with granules have been suggested to indicate similar underlying pattern formation mechanisms (Hammer, 2000). However, their combination with a single, smooth but punctured marginal strip appears to be a rare feature. The stratigraphically oldest potential cases that we could find date from the earliest upper Emsian. If these examples are all correct then considering their distribution among phacopide ranks, this type of dual tubercle bands is likely to have evolved much earlier.
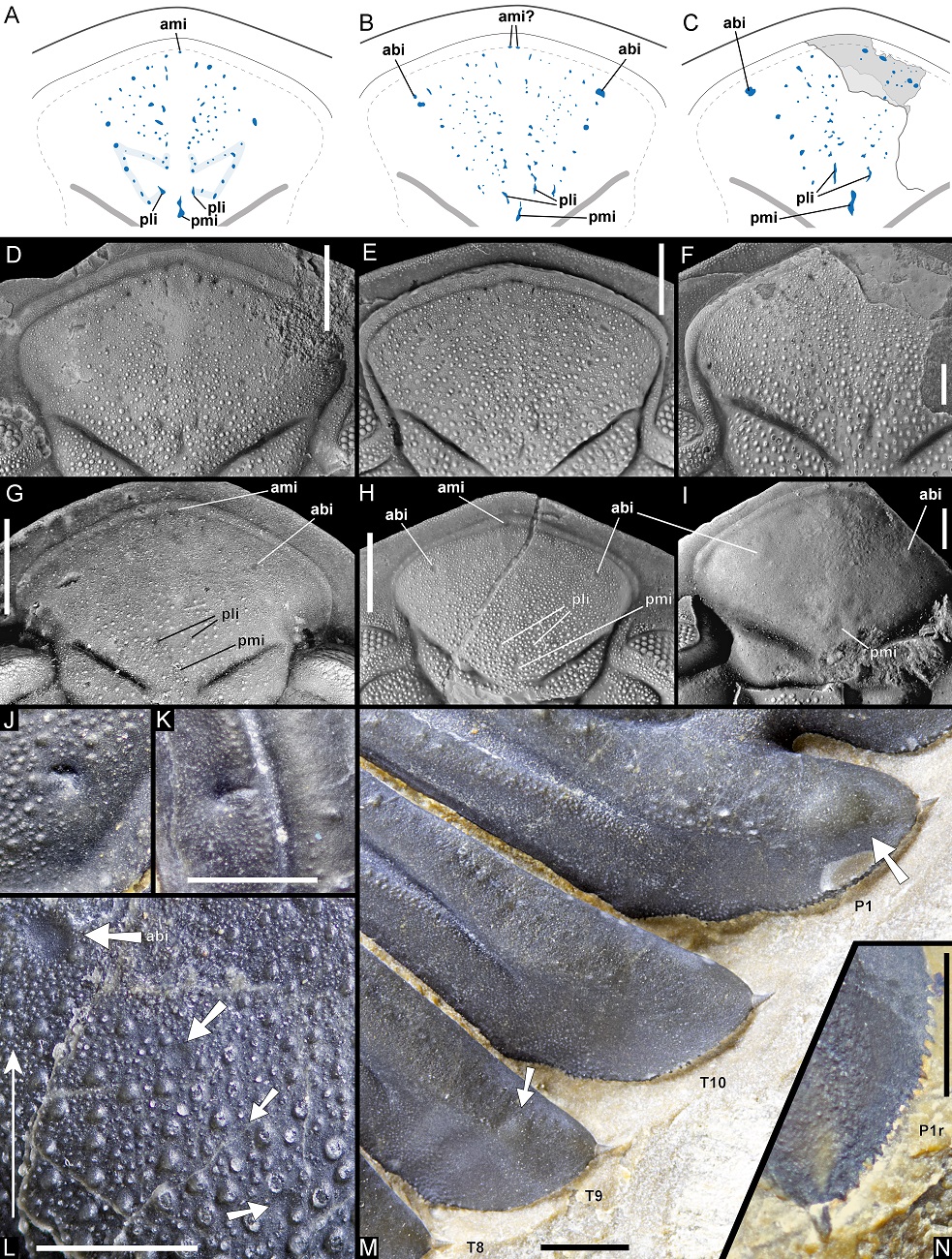
Figure 7. A–I. Frontal auxiliary impressions (FAIs) in Hollardops. A, D. Hollardops sp. A, NHMM 2022014. B, E. Hollardops multatuli sp. nov., holotype, IRSNB a13567. C, F. Hollardops multatuli sp. nov., paratype, NHMM 2022001. G. Hollardops boudibensis Morzadec, 2001, IRSNB a13571. H. Hollardops angustifrons sp. nov., holotype, IRSNB a13570. I. Hollardops sp. B, NHMM 2022015 (both pli’s are recognisable as dark depressions just anterolateral to pmi).
J–N. Selected features of Hollardops multatuli sp. nov., paratype, NHMM 2022001. J. Partially healed, vermiform hole on left side of glabella. K. Partially healed, vermiform hole on fourth left thoracic pleura. L. FAIs on left side of glabella. M. Neoplasms on the thorax and pygidium. N. Thorns adorning the right first pygidial pleural lappet.
Abbreviations for FAIs: ami – anterior median impression; abi – abaxial impression; pli – posterior lateral impression; pmi – posterior median impression. P1r is right first pygidial pleura. Elongated arrow in L points anteriorly (exsag.). Broad arrows in L indicate FAIs. Arrows in M indicate neoplasms. Scale bars are 2 mm. J is 2 mm wide.
4.2. Coaptation
27Stabilisation of thoracic segment numbers is common in phylogenetically derived trilobite taxa, which has been attributed by some workers to intrinsic developmental constraints (e.g. McKinney & McNamara, 1991). However, Hughes et al. (1999) have argued that stable thoracic segment counts in post-Cambrian trilobite groups reflect the evolution of derived morphotypes with more efficient enrolment mechanisms, i.e., as a behavioural response to deteriorating environmental conditions and as a defence against predators. If selective pressure favoured better enrolment then it is worthwhile to evaluate whether Hollardops had any distinctive coaptative devices. Many Hollardops species are subisopygous in that the pygidium is slightly narrower, but rarely much shorter, than the cephalon. This is an uncommon condition among Early Devonian, often micropygous asteropygines and likely a derived feature in balance with the 10-segmented thorax. Indeed, when comparing the relative lengths of the tagmata, e.g. of Hollardops multatuli and the phylogenetically close, 11-segmented Feruminops zguidensis comb. nov. (see 7.3. for revised classification), the pygidium of the former is longer at the cost of the thoracic length whereas the cephala of both species show similar values. As will be argued below, the position of the pygidium relative to the cephalon during full enrolment is more anterior in Hollardops than in some other asteropygines. A comprehensive study of the mechanics of enrolment of Hollardops is beyond the scope of the present work but published specimens (e.g. Morzadec, 1997, pl. 6, figs 1, 6; Chatterton et al., 2006, pl. 20, figs 8, 12, 15; Klug et al., 2008, pl. 11, figs 4–7, 13–16; Crônier et al., 2018, p. 247, fig. 3w–z) offer convincing evidence that its species were well capable of complete coaptation.
28Some of the individuals recorded are of comparatively large size. The large paratype of H. multatuli is also concavely outstretched, thus, fully exposing the lateral extremities of the thoracic pleurae and revealing important details of the dorsal cuticle. Other specimens reveal aspects of the ventral morphology of Hollardops. Here we discuss features considered by us to be pertinent to enrolment.
4.2.1. Thoracic facets and facetal conditions
29The abaxial parts of the thoracic pleurae form a bevelled (exsag.) gliding plane or facet. The anterior and posterior margins of the facets are respectively demarcated by the articulating flange and a change of inclination toward a dorsoventrally high ridge that runs obliquely laterally on the pleura. The highest point of this ridge is marked by comparatively large tubercles. The surface of each facet is covered with close-set tubercles (Fig. 6E–H). This ‘roughening’ is presumed to have increased friction between the overlapping thoracic segments upon full enrolment (e.g. Edgecombe, 1991; Chatterton & Campbell, 1993).
30Jaanusson (1975) noticed that many members of Phacopina have the thoracic pleural furrows continued onto the facets, which he termed the epifacetal condition. Speyer (1988) studied the phacopid Eldredgeops and concluded that it has postfacetal furrows which led him to infer a nearly perfect spherical mode of enrolment. Bruton & Haas (1997) identified a different hinge line for the thoracic segments than that of Speyer (1988) and rejected his claim that the epifacetal furrows could have played any part in ensuring enrolment or have functioned as a locking device. Whittington (1997) associated epifacetal and postfacetal conditions with certain trilobite groups, e.g., using it to contrast dalmanitids with phacopids. Such categorisations based on facetal modes appear ineffective. Firstly, it is often difficult to assess the abaxial course of the pleural furrow in articulated trilobite specimens because the anterior pleurae are usually obscured from sight by one another or by the genal spine. Secondly, the large paratype of H. multatuli, which has all the thoracic pleurae fully exposed, reveals a combination of both facetal conditions. On the posterior thoracic segments, the deep pleural furrow is abaxially continued as a posterolaterally flexed, shallow postfacetal furrow running along the posterior margin of the facet (= postfacetal) (Figs 5D, 6G). On the middle and anterior thoracic segments, the pleural furrow continues laterally onto the facet where it is somewhat less firmly impressed (= epifacetal) (Figs 5D, 6E, H).
4.2.2. Thoracic posterolateral and posterior pits
31Some of the thoracic segments of the large paratype of H. multatuli bear small pits, laterally and posterolaterally on the dorsal surface, that appear to be posterolaterally directed canal openings (Fig. 6E, F, H). These pits are more numerous and more densely spaced on the middle segments of the thorax. They appear to be lacking on the posterior segments and on the pygidial lappets. The pits are taken to have housed setae that may have functioned as brushes to clean the facets as segments slid over one another. Such brushes would have prevented residual sediment grains that otherwise might have (1) hampered coaptation, (2) weakened the gripping force of the tubercular surface of the facets upon full enrolment, or (3) caused abrasion of the articulating surfaces. Miller (1976) suggested similar functions of the setae that he argued were situated on the articulating half-rings and pleural tips of the phacopid Eldredgeops, while adding that these probably also provided some proprioceptive sense. Additionally, (4) these setae may have had an anti-fouling function (see 3.3.).
32A shallow transverse furrow is discernible distally on the thoracic posterior pleural bands of the large paratype of H. multatuli (Figs 6E, G–I, 18K). This furrow divides the convex (exsag.) anterior part from the weakly vaulted (exsag.) posterior part of the abaxial half of the pleural band. Large, moderately widely spaced pits occur in some of these furrows (see, e.g., on segments T6 and T7 in Fig. 6E, H). The holotype of Asteropyge boeckae van Viersen et al., 2019 shows the same row of pits on the abaxial part of each of the thoracic posterior pleural bands (Fig. 14K). It is conservative to assume that these openings, too, housed setae. The locations of the pits, far from where the segments interacted with one another during enrolment, indicate that these setae did not act as brushes and only fulfilled a sensory, and possibly also anti-fouling function.
33The adaxial parts of the thoracic anterior and posterior pleural bands of H. multatuli are homogeneous except for the tapering anterior band near the axial ring. These areas carry granules near their anterior and posterior margins and fairly widely spaced pits centrally (Fig. 17M). The pits also occur on the axial ring where they are alternated by dome-shaped (pitted) tubercles.
4.2.3. Aspects of the ventral morphology
34Heretofore, the cephalic doublure and pygidial posterior rim of Hollardops were only known from photos of ventrally prepared specimens on the Internet. Figures 17H and 9H–O, Q show these features in specimens of H. burtandmimiae and H. boudibensis. Another, partially exfoliated Hollardops sp. C specimen (Fig. 9C–E) reveals the lateral cephalic doublure to coincide with the lateral border.
35The posterior two to three thoracic segments of Hollardops are consecutively decreased in width. This gradual tapering affects both the axial and pleural regions and continues onto the pygidium. Upon complete enrolment, the ventrally flattened tips of the posterior thoracic pleurae partially overlapped one another, forming a broad platform in horizontal continuation with the posterior rim for the cephalic doublure to rest on. This is eloquently shown by the damaged but fully enrolled Hollardops individual of Chatterton et al. (2006, pl. 20, figs 8, 12, 15). A partially coiled H. boudibensis specimen (Fig. 9M–O) shows how the posterior thoracic pleural tips approach the cephalic doublure whereas the other thoracic pleural tips are laterally concealed by the base of the genal spine.
36The inner margins of the doublure converge anterior to the eyes where they reach the attachment point of the large hypostome (hence the impendent condition). All Hollardops species have the anterior cephalic border medially ventrally deflected to a certain extent, with H. burtandmimiae (Figs 16F, 17K) as the most extreme example and H. boudibensis (Figs 9I, 17E) having the weakest deflection. Hollardops species typically also have the anterolateral cephalic margin in front of the glabella withdrawn from the cephalic outline, often rendering the anterior border slightly longer sagittally than exsagittally, and causing a generally somewhat angular instead of smoothly rounded cephalic outline.
37Upon complete enrolment, the median parts of the cephalic doublure and the pygidial posterior rim of Hollardops did not coapt through flat surface contact (cf., e.g., sphaeroidal enrolment of Bergström, 1973). Instead, the pygidium was protruded relative to the cephalon, which implies a different mode of coaptation. This can be seen in the enrolled specimen of Chatterton et al. (2006, pl. 20, figs 8, 15) and in digital reconstructions of H. kyriarchos, showing how that looked had it been close to fully enrolled (Fig. 18A) and fully enrolled (Figs 8E2, 18B): the cephalic doublure lateral and anterior to the eyes rests on the posterior thoracic pleural lappets, the posterior rim of the pygidium, and the anterior three pygidial lappet pairs. Anterior from this point, the cephalic margin progresses inward, crossing the pygidial rim, until the ventrally deflected border abuts the inner margin of the medially slightly raised pygidial rim. Overlap drawings of the ventrally prepared H. burtandmimiae specimen (Fig. 8M, O) and the inferred ventral morphology of H. cf. circumapodemus (Smeenk, 1983) (comb. nov.; see 7.1.) (Fig. 8U) show a nearly identical configuration. From a lateral perspective, the ventrally deflected cephalic border of H. kyriarchos depicts a smooth downslope in sagittal continuation with the pygidial posterior rim (Fig. 18H). A nearly completely enrolled specimen of Breizhops lanceolatus Morzadec, 1983 figured by Morzadec (1983, pl. 16, fig. 1a–c) shows that it probably enrolled in a similar way to Hollardops. This mode of coaptation is equivalent to the pseudomegalaspid type of Harrington (1959) and was probably already present in Asteropyginae with 11 thorax segments during the Pragian (e.g. Fig. 8C2).
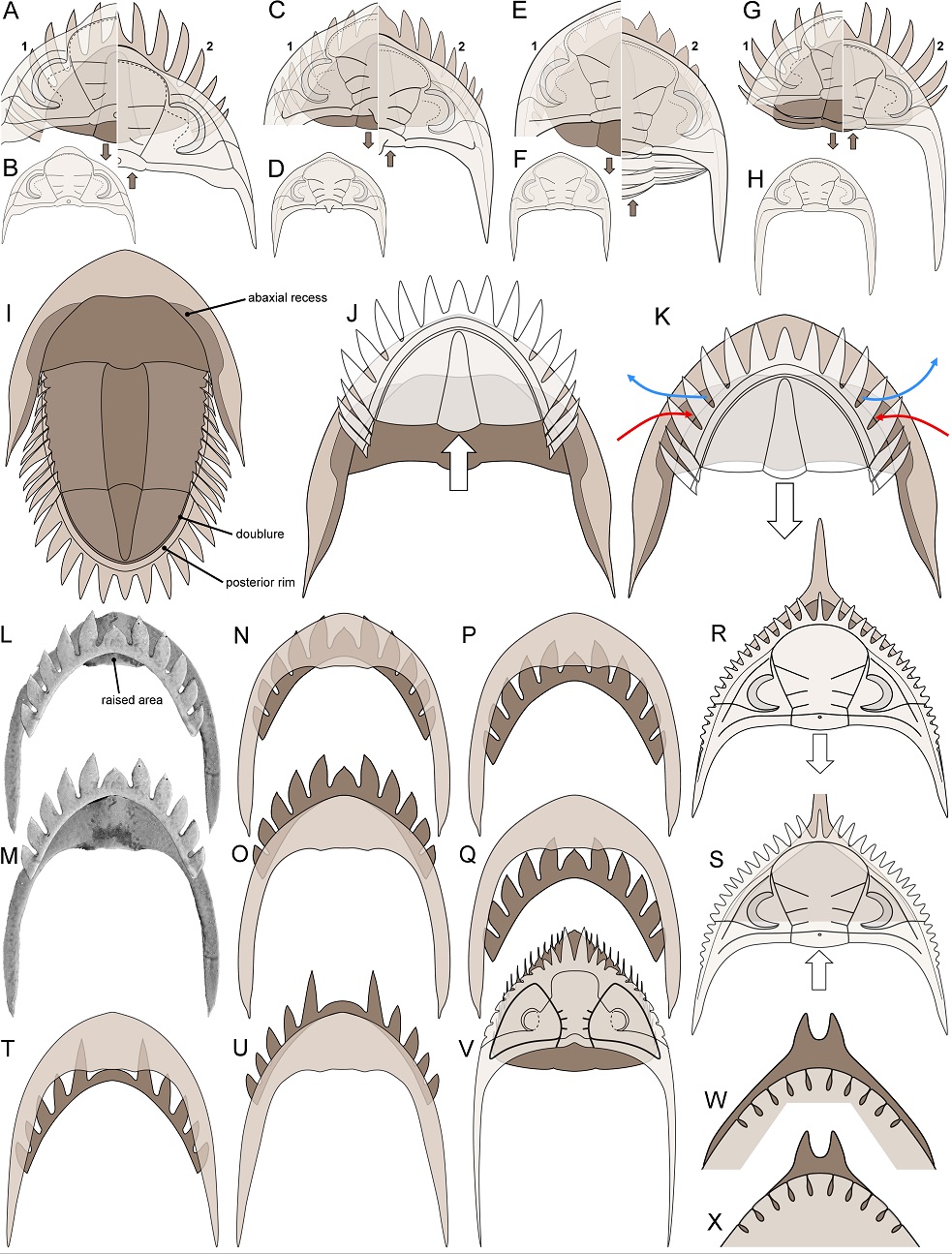
Figure 8. Ventral morphology and coaptation in selected trilobites. A, B. Delocare bensaidi (Morzadec, 2001), upper Emsian, Morocco. C, D. Platykardiapyge maderensis (Morzadec, 2001), lower Emsian, Morocco. E, F. Hollardops kyriarchos sp. nov., upper Emsian, Morocco. G, H. Asteropyge filoxenia van Viersen, Taghon & Magrean, 2019, lower Eifelian, Belgium. I–K. Bellacartwrightia sp. of Whiteley et al. (2002), Givetian, North America. J and K demonstrate the dual mode of coaptation, differentiating the locked state (J) and retracted pygidium (K). L–Q. Hollardops burtandmimiae (Lieberman & Kloc, 1997), Devonian, Morocco. NHMM 2021041, in retracted (L, N, P, Q) and locked (M, O) states. R, S. Huntoniatonia lingulifera (Ulrich & Delo, 1940), Lochkovian, North America, retracted state redrawn from Campbell (1977, p. 77, fig. 23a) (R) and locked state (S). T, U. Hollardops cf. circumapodemus (Smeenk, 1983), upper Emsian, Spain, based on specimen in Fig. 12, and ventral aspects inferred from Fig. 17H, in retracted (T) and locked (U) states. V. Coronocephalus gaoluoensis Wu, 1979, Silurian, China, redrawn from Chen et al. (2011, fig. 2). W, X. Odontocephalus bifidus (Hall, 1861), Middle Devonian, North America, in putative locked state (W) and with the pygidium slightly retracted (X).
Red and blue arrows indicate oxygenated seawater and excretion, respectively. Numbers in A, C, E, G indicate retracted (1) and locked (2) pygidial states.
38The enrolled Hollardops specimen of Chatterton et al. (2006, pl. 20, fig. 15) has the pygidial border broken off. This enables the observation that the cephalic border runs abaxial to the right pygidial pleural field and inward, towards the short (sag.) postaxial area, just anterior to where the broken posterior rim would have been. The anterior view of this same specimen (ibid., fig. 12) reveals that it is perfectly enrolled in showing how the cephalon and pygidium are tightly locked. Thus, during full enrolment, the ventrally deflected part of the cephalic border was positioned within the boundaries of the pygidial posterior rim, helping to prevent lateral shearing. The posterior border of the pygidium of Hollardops projected beyond the cephalic margin, with the pygidial median lappet and posterior pleural lappet pairs providing protection against predators attempting to force the trilobite. The protracted area should be expected to be prone to damage (from erosion, collecting and preparation alike) in fully exposed, enrolled individuals, and it is indeed broken in such specimens figured by Morzadec (1997, pl. 6, figs 1, 4), Chatterton et al. (2006, pl. 20, figs 8, 12, 15), Klug et al. (2008, pl. 11, figs 4–7, 13–16; Fig. 21H, M) and Crônier et al. (2018, p. 247, fig. 3w–z).
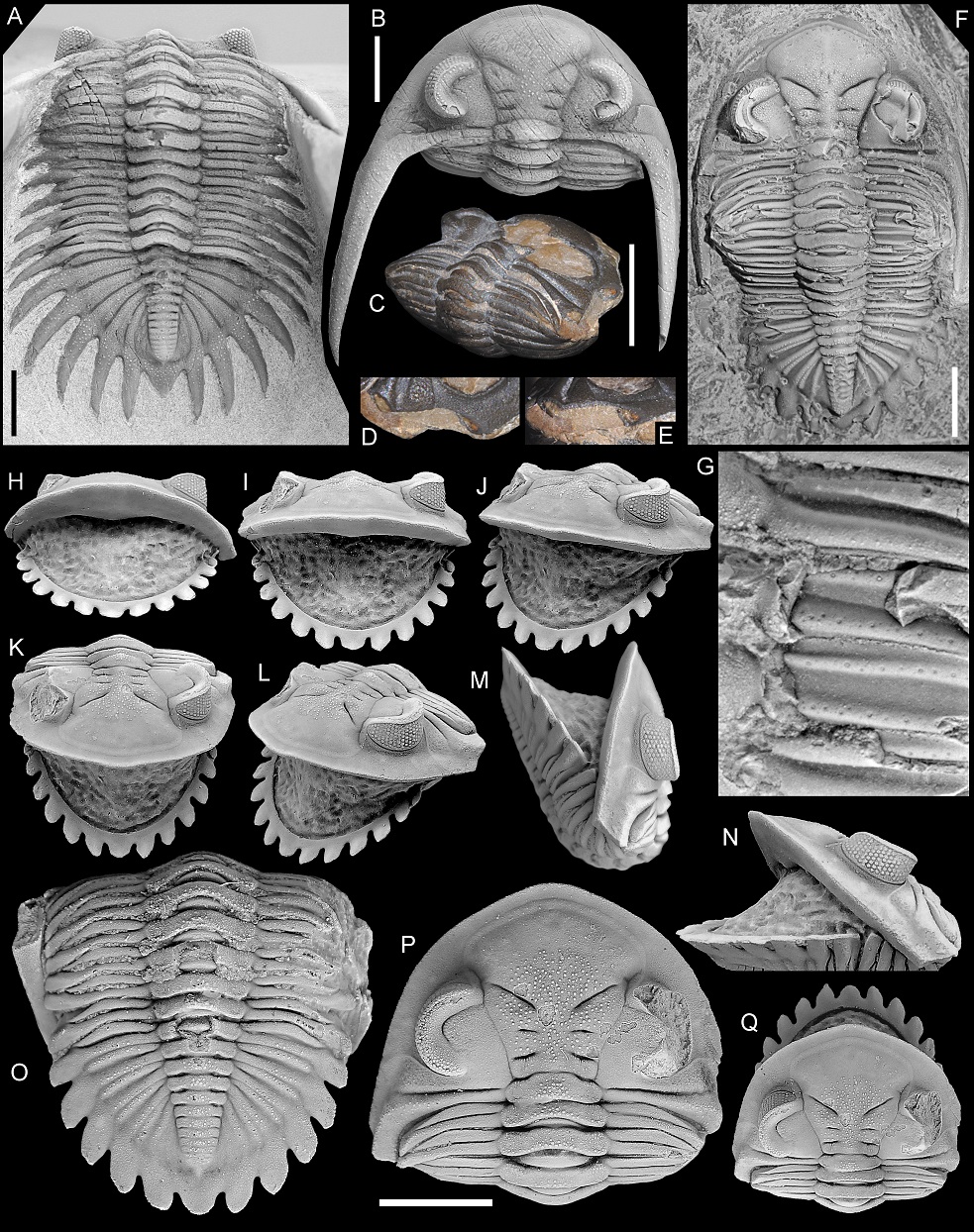
Figure 9. A, B. Asteropyge filoxenia van Viersen, Taghon & Magrean, 2019, Vieux Moulin Member of the Jemelle Formation (lower Eifelian), Nismes, Belgium. Specimen in the collection of P. Taghon, in dorsal views of pygidium (A) and cephalon (B).
C–E. Hollardops sp. C, “Devonian of Morocco”. NHMM 2022016, posterolateral view (C) and close-ups of right cephalic border (D, E).
F, G. Hollardops sp. A, “Devonian of Morocco”. NHMM 2022014, close-up of exfoliated area on the right thoracic pleural region (F) and dorsal view (G).
H–Q. Hollardops boudibensis Morzadec, 2001, “Devonian of Morocco”. NHMM 2021042, in various views.
Scale bars are 5 mm.
4.2.4. Robust genal spine bases
39Genal spines in trilobites are generally believed to have served a variety of functions or combinations thereof (Fortey & Owens, 1997). It is conservative to assume that if an individual was enrolled, the genal spines, with their sharp tips, functioned as a primary defence against predator attacks. But the robust bases of the genal spines of Hollardops also inhibited lateral shearing to a certain extent. From the back to the front of the animal each thoracic pleura slid over the previous. The anteriormost facet was topped by the large base of the genal spine which blocked transverse movement of the interlocked thoracic pleurae (Fig. 18H).
4.3. Spiny lappets
40The thoracic pleurae of Devonian members of Asteropyginae and Dalmanitinae are often extended into tapered projections. These may rise from the anterior and posterior pleural bands or from the posterior band alone. The pleural projections range from broad, short, rounded protuberances to elegant, long spines of subcircular section, with a full spectrum of intermediary shapes and sizes. In the first Trilobite Treatise, Harrington et al. (1959) defined the term ‘pleural spine’ as the pointed extension of distal end of a pleura. Struve (1959a), also Trilobite Treatise, described Greenops as having broad, flat lappets, but the lappets were nowhere defined. The term ‘pleural lappet’ has been in use at least since the work of Delo (1940). Whittington & Kelly (1997) described the pleural spine as a hollow, pointed extension of distal end of a pleura or of either band of the pleura, of thorax or pygidium. Again, the term lappet was not defined. In our descriptions we have used the term ‘pleural lappet’ for the comparatively short, flattened or deflated, somewhat truncated (non-spinous) projections of the thoracic and pygidial pleurae of Hollardops species. Although we do not doubt that the pleural lappets are homologous with the pleural spines of most Asteropyginae, this does not necessarily imply that they fulfilled the same functions. We contemplated on a universal definition for the term ‘pleural lappet’ but concluded that the progressive change spanning a range of forms up to ‘true’ pleural spines, impedes the recognition of morphologically demarcated groupings. Thus, both terms are gradations of a homologous feature and their use remains a matter of personal preference.
41Hollardops has relatively short, blunted, ventrally flattened pleural lappets. Each lappet carries a single minuscule spine posterolaterally; the spines are smallest on the anterior pleurae (Figs 5D, 6A, 18K) and of increased size on the posterior ones (Figs 5D, 6D, E, G, I, 7M, N, 18K). Because of their small size, marked protrusion from the contour of the lappet, and locations far posteriorly on the pleurae, we do not consider these spines to be homologous with the pleural spines of other asteropygines. Instead, they formed the apex of a saw-like array together with the marginal thorns (see 4.1.) that would have deterred predators trying to infringe the enrolled animal. As far as we are aware, such spines have never been described in a member of Acastoidea, probably because specimens are usually too small to have them preserved. With this knowledge, their remains can be recognised in several species of Hollardops (e.g. Chatterton et al., 2006, pl. 17, fig. 3; Figs 15A, 16B, 20B, H). Whiteley et al. (2002, pl. 44) figured a specimen of Bellacartwrightia phyllocaudata Lieberman & Kloc, 1997 which shows similar spines on the thoracic and pygidial spines. Outside of Asteropyginae, we could only find one case of conceivably homologous short spines, which is in the homalonotid Burmeisterella (Müller & Alberti, 2010, fig. 1c). Somewhat similar, yet larger spines are shown by the homalonotid Spiniscabrella (Lebrun, 2018, p. 243, fig. G). Thus, at present, the feature is only known to occur in members of Phacopida.
5. Enrolment strategies in Asteropyginae
42Little attention has been given in the literature to coaptation in asteropygines. Varying cephalic and pygidial outlines and size ratios, lengths of thoracic and pygidial pleural lappets, and genal spine sizes and shapes, among other features, which are shown by various members of the subfamily, may point to divergent enrolment strategies.
5.1. Taxa with short to moderately long pleural spines
43Asteropygines with short to moderately long pleural spines are often associated with exsagittally running to slightly divergent, proximally robust genal spines. Such taxa typically have a fan-like configuration of the posterior thoracic and pygidial pleural spines in that these are longer than the anterior thoracic pleural spines adjacent to (and to accommodate) the genal spines. In many genera, the posterior two to four thoracic segments including their spines or lappets, become progressively narrower (tr.): e.g. Hollardops (see above), Feruminops (Morzadec, 2001, pl. 4, fig. 1a, b), Neometacanthus (Basse, 2003, pl. 24, figs 395, 406), Greenops (Whiteley et al., 2002, pls 50, 52, 55, 56) and Bellacartwrightia (Whiteley et al., 2002, pls 45, 47, 48). This is not the case in other genera like Mrakibina (Morzadec, 2001, pl. 14, fig. 3) and Asteropyge (Fig. 9A) which have the posterior thoracic pleural spines becoming consecutively longer and projecting further abaxially. Yet the posterior thoracic axial rings and pleurae excluding the spines, show the same declining width like Hollardops and others. This means that Asteropyge and Mrakibina probably coapted in ways similar to other fan-bearing taxa and that their longer spines projected further beyond the cephalic margin. Like Hollardops, all these genera have non-matching cephalic and pygidial outlines. Their modes of coaptation, including the pertinent question of whether or not the anterior cephalic border abutted the pygidial rim in the same way as Hollardops, are ideally inferred from the ventral morphology of complete exoskeletons yet often remain inconclusive because published photos or preservation of the specimens do not allow such observations.
44No published records are known to us of well-preserved, uncompacted, fully enrolled specimens of asteropygines with moderately long or long pygidial pleural spines. The most widely acknowledged reconstruction is that of Kayserops kochi (Kayser, 1883) by Richter & Richter (1943, pl. 8, fig. 5), repeated by Struve (1959b, p. O481, fig. 381.2) in the first Trilobite Treatise and Kowalski (1990, pl. 5, fig. 4), and elaborated on by Bergström (1973). The drawing shows an enrolled individual with matching cephalic and pygidial margins such that a defensive array of pygidial spines projects anteriorly. This reconstruction is based on an incompletely prepared, deformed specimen that was later selected as the type of Rheingoldium feldbergense by Basse (2003, pp. 63, 64, pl. 11, fig. 171). From Basse’s photo, it becomes clear that only the cephalon is exposed besides parts of the pygidial margin and several spines. Basse (2003, pl. 11, fig. 172) illustrated a conspecific pygidium the outline shape of which cannot possibly be matched with that of the cephalon of the holotype. We underline the speculative nature of the drawing by Richter & Richter (1943) and reject Bergström’s (1973) conclusion that the cephalic and pygidial borders of this species coincided. The extent to which the posterior part of the pygidium was overlapped during enrolment depends on its position relative to the cephalic margin along the sagittal line. The enrolment strategy of R. feldbergense is difficult if not impossible to determine based on the currently available material.
5.2. Dual mode of coaptation
45Comprehensive photographic evidence of the ventral morphology of asteropygines is rare. Whiteley et al. (2002, pl. 47) illustrated a fully ventrally prepared, outstretched and articulated exoskeleton of a Bellacartwrightia species, repeated as a line drawing here (Fig. 8I). This specimen reveals valuable clues about how it would have enrolled. We take note of several observations here: (1) pygidial width (tr.) is 0.6 of cephalic width across occipital ring, (2) pygidial length (sag.) is 0.75 of cephalic length, (3) the cephalic doublure has developed a large abaxial recess (i.e., the adaxial part of the lateral doublure is steeply inclined, into the cephalic cavity), and (4) pygidial width (tr.) is 0.7 of maximum width across abaxial recess. Thus, all measurements of the pygidium are smaller than those of the cephalon, indicating that their margins could not possibly have fully coincided during enrolment. There is slight room for interpretation in matching the cephalon and the pygidium along the sagittal line, ranging from a position of the anterior cephalic border abutting the pygidial rim, to a position in which the pygidial doublure makes flat surface contact with the cephalic doublure (the option we have chosen). The large concave abaxial recess of the otherwise broad cephalic doublure is a striking feature. We postulate that it allowed effortless passage of the comparatively long thoracic pleural spines as the thorax coiled in a swing motion during coaptation. However, this does not explain the far anterior process of the recessed doublure, well beyond the area of the cephalon that is covered by the thoracic segments during enrolment.
5.2.1. Functional mismatches between tagmata
46The functional morphology of ‘intentional’ mismatches between the cephalon and the pygidium during coaptation has been recognised in a variety of trilobites from the Silurian, Devonian and Permian periods. Clarkson & Henry (1973) noted a small triangular open space in the Silurian encrinurid Encrinurus tuberculatus (Buckland, 1836), where the pygidium projects beyond the cephalon during full enrolment (however, see remarks by Clarkson & Whittington, 1997). Clarkson & Henry (1973) interpreted this opening as a pygidial ‘breathing-hole’ in suggesting that it allowed the trilobite to circulate water freely and thus, spend longer periods of time in an enrolled state. Ormiston (1976) noticed that the pygidial margin of the Devonian proetid Camsellia is narrower than the cephalon and that it protrudes beyond the anterior cephalic margin during enrolment. This mode of coaptation left anterior and lateral gaps which Ormiston believed would have permitted feeding and excretion. In analogous examples, Lerosey-Aubril & Angiolini (2009) suggested that the anteriorly projected pygidium of the Permian proetid Ameura allowed water to flow internally and Feist et al. (2010) made a case for the Devonian scutelluid Paralejurus. Morzadec (1983) described the asteropygine Coltraneia celtica and noticed that in a fully enrolled state, the dorsally convex anterior cephalic border would leave a space between the cephalon and the pygidium. Morzadec also pointed out a similar structure in the asteropygine Deloops haasi (Morzadec, 1983) and suggested that both species were able to let water pass through the open space during enrolment. Klug et al. (2008, pl. 11, figs 8–11; repeated here as Fig. 21O–R) figured a fully enrolled specimen of Minicryphaeus which has a broadly rounded opening anteriorly between the cephalon and the pygidium that may have had the same purpose. Campbell (1977) showed the anterior cephalic margins of the Devonian dalmanitids Huntoniatonia, Prosocephalus and Phalangocephalus to be adapted in such ways, that they were overlapped by the anteriorly protruding pygidial cavity during enrolment. Anterior cephalic crenulations bridged the gap that remained but Campbell (1977) concluded that numerous slits between the spines would have been left open and suggested that they allowed entry of water and the protrusion of the antennae during enrolment. Holloway (1981) accepted Campbell’s (1977) explanation of the gaps and suggested the same for the crescentic cephalic margin of the Silurian dalmanitid Glyptambon. Campbell (1977) also proposed a ‘breathing’ function for small openings in the cephalic border of the dalmanitid Odontocephalus. When examining photographs of the cephala and pygidia of O. aegeria, O. bifidus and O. selenurus (see Whiteley et al., 2002, pls 123–125; Fig. 8W, X) their outlines seem to match well. Provided that the pygidium protruded far enough, its ventral cavity may have coincided with the cephalic border openings (Fig. 8W) which would be consistent with Campbell’s interpretation. However, an assessment of their ventral morphologies is required to corroborate this. Other trilobites, such as the dalmanitids Neoprobolium and Synphoroides (see Delo, 1940), the phacopid Echinophacops (see Zhou & Campbell, 1990), the homalonotid Wenndorfia (see Chatterton et al., 2006, pl. 33, figs 4, 5) and the cheirurid Coronocephalus (see Chen et al., 2011; Fig. 8V), also had openings during enrolment that are reasonably explained as breathing devices. With the exceptions of the proetids Camsellia and Ameura and the scutelluid Paralejurus, all these examples occur among taxa in the three suborders of Phacopida during the Silurian and the Devonian (we have not performed an exhaustive literature search but suspect that functionally analogous features may be scattered among enrolling members of the class). Clearly, the functional mismatch between the cephalon and the pygidium was an advantageous feature that evolved, independently, in multiple lineages. Such a perquisite, however, would have come at a price. Although the openings allowed the animal to remain enrolled for a longer period of time while maintaining important body functions, there would have been reduced protection against smaller hostile beings and temporary adverse changes in seawater quality.
47Lespérance (1975) studied phacopides from the Devonian of North America. He attributed major phylogenetic value to the crenulated anterior cephalic processes within his concept of Synphoriidae, in recognising two distinct evolutionary trends classified as Synphoriinae and Trypaulitinae. Eldredge (1977) accepted Campbell’s (1977) explanation of a water circulation system and suggested that Lespérance’s trends in part at least, seem to imply progressively more sophisticated control and monitoring of the passage of water to the ventral surface of the enrolled trilobite. Lespérance’s (1975) Synphoriidae was later rejected, and Synphoriinae (= Trypaulitinae) is regarded as a subfamily of Dalmanitidae (see Holloway & De Carvalho, 2009 for discussion). However, Eldredge (1977) had made a crucial point in stating that the anterior cephalic border of Lespérance’s (1975) Trypaulitinae remained unmodified except for one genus, Coronura, which, he reasoned, independently developed a water-monitoring system by the addition of spines along the periphery of the pygidium. Holloway & De Carvalho (2010) studied the closely allied genus Chacomurus from the Devonian of Bolivia and concluded that the pygidium would not have overlapped the front of the cephalon during enrolment, and that there would not have been horizontal openings between the cephalic spines and the inner edge of the pygidial doublure. Both Coronura and Chacomurus have a pair of robust spines near the posterior end of the pygidium, the functional morphology of which has remained enigmatic.
5.2.2. Locked versus retracted states
48It might seem that, with few exceptions, members of Asteropyginae lacked a breathing device for enrolment. Yet the advantages should be expected to have outweighed the risks, as the device afforded significantly increased resilience against prolonged harmful conditions. Of interest would be an assessment of whether the grossly similar asteropygine and dalmanitine Bauplans could be taken as having had comparable energy demands, potentially rendering a breathing device during enrolment an important function.
49The fully enrolled state of most asteropygines, as far as known from published specimens, is a tightly locked spheroid, regardless of whether species possess 10 or 11 thorax segments (e.g. Figs 8A2, C2, E2, G2, 21H–R) and the same can be assumed for Bellacartwrightia (Fig. 8J). However, if the pygidium of Bellacartwrightia is slightly retracted (Fig. 8K), with the posterior rim resting on the cephalic doublure and bordering the hypostome, large slits appear between the pygidial spines, the tips of which rest on the cephalic doublure even where the latter is recessed. Retracting the pygidium would be a controlled and relatively safe movement for the animal and there is no reason to assume that it was not possible from a functional perspective. Perhaps the posterior margin of the abaxial recess functioned as a doorstop for the posterior thoracic segments when the pygidium was retracted. We propose that this dual (‘locked’ versus ‘retracted’) mode of coaptation is an analogue, if not improved version, of a device designed to allow seawater to circulate internally and antennae to protrude. The retracted state of Bellacartwrightia is basically a mirrored version of the fully enrolled state of the above examples in dalmanitids and cheirurids which, in the absence of a spiny pygidium, evolved various adaptations of the cephalon (crenulations, spiny protuberances, border openings) to create breathing holes. Basse (2017, p. 33, fig. 1) figured a ventrally exposed Rhenops specimen with soft parts preserved. The emergence of the antennae lateral to the hypostome and their anterior course could match the size and position of the slits between the third and fourth pygidial pleural spine pairs in the retracted pygidial state. Importantly, dual coaptation had few prerequisites in order to function. Considering its marked advantages, it may be expected that effective dual coaptation was under selective pressure and that it was widely distributed within Asteropyginae. This will be difficult to prove, especially without knowledge of the ventral morphology of most species, but the basic rule is that, in the putative retracted state, posterior to lateral outline of pygidium + pleural projections ≈ cephalic anterior to lateral outline. This rule can be tested by comparing the dorsally exposed cephala and pygidia of complete specimens or the associated cephalon and pygidium of the same individual. It appears that throughout the Devonian, many genera in Asteropyginae adhere to this principle (e.g. Fig. 8A1, C1, E1, G1). It is furthermore arguable that the retracted state was the ‘normal resting’ mode and that full coaptation (locked state) only occurred in case of imminent danger. Upon revisiting Campbell’s (1977, pp. 76, 77, figs 21–23) dalmanitid examples, one species, Huntoniatonia lingulifera (Ulrich & Delo, 1940), has partially coinciding cephalic and pygidial outlines (excluding the cephalic crenulations) that could have supported a locked state (Fig. 8S), in addition to Campbell’s reconstruction (redrawn here as Fig. 8R) that would then represent the retracted state.
50Turning to the evidence in Hollardops, most species have comparatively large pygidia with proportionally short pygidial lappets. This condition complies with the basic rule regarding their dimensions relative to the cephalon, as alluded to above. With the knowledge that the dorsal lateral cephalic border coincides with the lateral part of the cephalic doublure (see 4.2.3.) it becomes possible to reconstruct the retracted state of H. multatuli, which reveals that slits between the anterior pygidial lappets provided access to the ventral side (Fig. 8E1). One specimen of H. burtandmimiae (Fig. 17H) allows the corroboration that putative slits occurred in the retracted state, based on its ventral morphology (Fig. 8L). Additionally, this specimen offers valuable insights into the various possible stages of dual coaptation of a Hollardops species: (1) The locked state (Fig. 8M, O) of pseudomegalaspid enrolment type was already described above. (2) Of note is a raised area posteromedially on the cephalic doublure, adjacent to the attachment point of the hypostome, which may have afforded occlusion to the pygidial rim when the pygidium was slightly retracted (Fig. 8L, N). In this stage, small slits are created between the anterior four pleural lappet pairs. (3) More and larger slits could be created if the pygidium was retracted up to the point where the ventrally flattened median lappet overlaps the raised area of the cephalic doublure (Fig. 8P). The different dorsoventral profiles of these features meant that flat surface contact would not have been possible; hence this stage was unstable and probably not preferred. (4) The long fifth lappet pair of the pygidium might have provided a suitable platform for the cephalic doublure to rest on in the maximum retracted state (Fig. 8Q). Species formerly assigned to Pennarbedops (= Hollardops, see 7.1.), which have a cephalon of high parabolic outline like H. burtandmimiae and an even longer fifth pygidial pleural lappet pair, may have regularly assumed this maximum retracted state considering that their rudimentary median lappet would have adjoined the cephalic doublure instead of overlapping it (Fig. 8T). The pygidial spines of H. klugi are all broken off in the available specimens. Since this species has a fairly small pygidium compared to congeners it may be expected to have proportionally longer pygidial spines.
51Another piece of argumentation in support of dual coaptation comes from a partially coiled H. boudibensis specimen (Fig. 9H–Q) which shows that the cephalic doublure is not a horizontal plane, and that instead it is weakly sloped, forming a somewhat cushion-like inflated, ventral side of the cephalon that would be pressed against the slightly ventrally directed pygidial lappets (e.g. Fig. 5G, 18J, 19E) during the retracted state of enrolment. The ventral surface of Hollardops is generally ‘roughened’ by granules which increased friction. It follows that these features enabled the trilobite to remain in a firmly fixed retracted state and may be taken as evidence to suggest that this was indeed a frequently assumed or ‘normal resting’ mode.
52The pair of robust spines posteriorly on the pygidia of Coronura and Chacomurus may be a convergence on Asteropyginae and could have supported the cephalon in the same way as former Pennarbedops species. In that case the marginal embayment of the posterior pygidial border (e.g. Holloway & De Carvalho, 2010) allowed a larger volume of water to pass simultaneously. Another example of a putative convergence on Asteropyginae is provided by the Devonian dalmanitine Erbenochile (see Chatterton & Gibb, 2010 and Bignon & Crônier, 2013 for revised subfamilial assignment). Erbenochile is unusual among Dalmanitinae in having a comparatively small, somewhat Asteropyginae-like pygidium that carries an array of pleural spines and that would be overlapped by the cephalon during enrolment. These adaptations are perfectly accounted for from a functional perspective in that the spines provided the same support for the cephalon when the pygidium was slightly retracted during enrolment.
5.3. Anterior vincular and lateral ventral cephalic furrows
53The asteropygine Bignonops tamnrhertus comb. nov. (see systematic chapter for revised classification) does not show a distinctly fan-like spine configuration like some other asteropygines (see 5.1.). Instead, it has a posteriorly strongly tapered thorax and pygidium. Furthermore, the cephalon has a firmly impressed, V-shaped anterior vincular furrow in which the raised posterior rim of the pygidium fitted (Chatterton et al., 2006, pl. 20, figs 9, 11). The lateral cephalic doublure is divided along its length by a broad, concave (tr.) furrow (Chatterton et al., 2006, pl. 20, figs 5, 9) that would seem to be capable of accommodating the lateral pygidial rim when fully enrolled. If indeed correctly considered a coaptative device, these lateral furrows would qualify as a vincular structure according to the definition of Whittington & Kelly (1997). However, this appears not to be the case, and instead, the pygidial margin retains the more inward direction of the V-shaped vincular furrow, crossing the cephalic margin to adjoin the inner margin of the lateral cephalic doublure. In this way the anterior pygidial spine pairs slightly protruded from the cephalic outline during full enrolment. The posterior spines projected fully beyond the cephalon outline as did the long, upward-curved median spine.
54The tapered pygidium and short, thin pleural spines imply that Bignonops would have struggled with sustaining a retracted state of the pygidium. Perhaps it was only able to create openings during enrolment by protracting the pygidium instead. If this assumption is correct, the furrows dividing the lateral cephalic doublure may have been true vincular furrows; i.e., they fitted the posterior thoracic pleurae and the lateral pygidial rim during a protracted state (for example, by slightly extending the thoracopygidium of the enrolled specimen of Chatterton et al. (2006, pl. 20, fig 14) anteriorly, its lateral margin could match these furrows). Whether or not this created a breathing hole sagittally and subexsagittally, between the pygidial posterior rim and the cephalic doublure would need to be tested. Such a hypothesis requires thorough knowledge of the ventral morphology of Bignonops and this is currently unavailable.
55We are aware of Greenops species that have a trench-like furrow in the lateral cephalic doublure, including G. widderensis Lieberman & Kloc, 1997 (Fig. 14O), G. barberi Lieberman & Kloc, 1997 and G. grabaui Lieberman & Kloc, 1997. The well-developed pygidial lappets of these species suggest that they had no need for a protracted state of enrolment. Greenops grabaui at least, has a flattened surface of the anteriomedian portion of the cephalic doublure and it lacks the V-shaped vincular furrow of Bignonops tamnrhertus. Unpublished specimens of G. barberi show the distal extremities of the thoracic pleurae to be ventrally reflected, forming flanges that might have fit the trench-like furrow in the cephalic doublure but despite the deformation that these specimens have undergone, the thoracic width seems to be less than the width across the furrows. Coaptation in Greenops species will be dealt with by us in a future paper.
5.4. Exceedingly spiny forms
56Exceedingly spiny taxa such as Comura, Psychopyge, Quadrops and Walliserops tend to have slender, widely curved genal spines and long pleural spines, and they often also have arrays of dorsal spines all over the exoskeleton. It is questionable whether enrolment was an important defensive strategy for such forms. This idea is consistent with the remarks of Chatterton et al. (2006, p. 12), who studied numerous trilobites from the Zguilma section and reported that extremely spinose specimens were almost never found enrolled as opposed to more ‘conservative’ forms. We consider such spiny taxa to have been poor diggers or swimmers, with an exclusively epibenthic lifestyle. Epizoans are thus more likely to occur on individuals of these trilobites. Their spines may have had a primary defensive purpose against predator attacks or they provided suitable foundations for epizoans to grow on as a means of camouflage.
5.5. Submarginal flange
57Speyer (1988) described a steeply inclined submarginal flange on the ventral surface of the pygidium of Greenops which is concentric with the posterior rim. Speyer postulated that the anterior cephalic margin fitted inside this flange and showed how the cephalon would be tucked away between the pygidial spines during enrolment (Speyer, 1988, p. 129, fig. 6, p. 136, fig. 12.4; Speyer, 1990, p. 453, fig. 370B). However, the cephalon of Greenops, like that of most asteropygines, is too broad to fit within the pygidial cavity. This leads us to conclude that Speyer’s submarginal flange had no function in facilitating coaptation. Instead, the submarginal flange appears to be homologous with the pygidial doublure.
6. Other Hollardops features
58Several observations are listed here that we cannot estimate the potential significance of at this time, mainly because they require large amounts of specimens to evaluate.
6.1. Frontal auxiliary impressions
59Cephalic muscle impressions in trilobites have been known since the pioneering works of Barrande (1852, 1872). Lerosey-Aubril et al. (2011) proposed the term FAIs (frontal auxiliary impressions) for metamerically paired markings on the anterior part of the glabella which they regarded as attachment sites for muscles connecting the anterior portion of the digestive tract. FAIs occur on the dorsal cuticle of all Hollardops species. They are often preserved as small depressions of varying size, depth and shape – circular or scar-like, in their simplest forms. Ventrally, FAIs may be recognisable as depressions or as pads.
60Fatka & Budil (2021) studied FAIs on internal moulds of the dalmanitid Dalmanitina from the Ordovician. They distinguished the posterior median impression (pmi)—a deep elongate pit on the posterior part of the frontal glabellar lobe; the anterior median impression (ami)—a conspicuous, transversally elongate pit located sagitally in the preglabellar furrow; anterior lateral impressions (ali)—represented by four pits arranged in two pairs flanking the ami; and abaxial impressions (abi)—a pair of depressions placed posterior to the anterolateral margin of the glabella, abaxially opposite the mid-distance between ami and pmi.
61Overlay drawings (Fig. 7A–C) were made of H. sp. A, and the holotype and large paratype of H. multatuli (Fig. 7D–F). The distribution of FAIs generally conforms to the Greenops pattern of Eldredge (1971, p. 55, fig. 2c) (revised as the Bellacartwrightia pattern herein since Eldredge’s specimen is a Bellacartwrightia sp.) except that note is made of the asymmetrical outline (tr.) of the pmi. Anterolateral to the pmi is a pair of scars of irregular, elongated outline, which we term here posterior lateral impressions (pli). The ami is located sagittally in the preglabellar furrow of H. sp. A (Fig. 7A, D). Two abaxially disposed impressions occur near the sagittal line, in the preglabellar furrow of the holotype of H. multatuli (Fig. 7B, E). The outer contour of the FAI system of H. sp. A might be more diamond-shaped and H. multatuli more kite-shaped, although H. multatuli has largely erratic patterns with few fixed landmarks. The pattern of H. sp. A shows certain consistencies left and right from the sagittal line although landmarks are not particularly well aligned (tr., exsag.). The cephala of H. boudibensis (Fig. 7G) and H. angustifrons (Fig. 7H) have weak ami and pmi, accentuated in the latter species by the lack of tubercles there. A mostly exfoliated cephalon of H. sp. B shows the ventral distribution of FAIs (Fig. 7I). These are preserved on the internal mould as pads abaxially and as impressions adaxially (i.e., respectively representing impressions and pads on the ventral cuticle).
62The distributions of FAIs on the anterior glabellar lobe have been used by workers to define clades within the Phacopina (e.g. Eldredge, 1971) but recent studies have questioned the phylogenetic significance of FAIs (Lerosey-Aubril & Peel, 2018; Fatka & Budil, 2021). The limited amounts of Hollardops specimens studied by us offer little insight into intra- and interspecific variation of this feature. Nonetheless, FAIs can be detected in published photos of various species in different asteropygine genera from the Lower to early Middle Devonian, generating a vast amount of data (e.g. Arbizu, 1977, pl. 4, fig 1a; Smeenk, 1983, pl. 1, figs 5a, 12a, pl. 2, figs 1b, 2, pl. 3, fig. 4c, pl. 4, figs 2, 10, 14, pl. 5, fig. 11, pl. 6, fig. 1a, pl. 8, fig. 5, pl. 13, fig. 1a; Morzadec, 2001, pl. 1, fig. 2a, pl. 3, fig. 4, pl. 4, fig. 2, pl. 5, figs 1a, 5, pl. 6, figs 1a, 2a, 3, pl. 7, figs 1, 2, pl. 10, figs 1a, 2a, pl. 11, figs 1a, 2b, pl. 12, fig. 2a, pl. 13, fig. 5a, pl. 14, figs 1d, 5a, 7, pl. 15, fig. 3d; Basse, 2003, pl. 3, fig. 59, pl. 4, fig. 69, pl. 9, figs 141, 152, 153, pl. 13, fig. 214, pl. 14, fig. 226, pl. 19, fig. 323; Bignon et al., 2014, p. 286, fig. 3). Many of these have the pmi firmly incised and some also have well-developed pli’s. FAIs seem to be less common in late Middle to Late Devonian asteropygines where sometimes only a weakly incised pmi can be recognised clearly (e.g. Haas & Mensink, 1970, pl. 3, fig. 1; Morzadec, 2002, figs 5.3b, 6.1; Whiteley et al., 2002, pls 45, 48; Basse, 2003, pl. 27, figs 446, 448), or the traces of an impression system (e.g. Haas & Mensink, 1970, pl. 5, fig. 4; Lieberman & Kloc, 1997, fig. 19.6; Whiteley et al., 2002, pl. 46). However, cephala of Neocalmonia orientalis (Feist et al., 2001, fig. 4A, C, D, F), Treveropyge cf. wiltziana (Basse, 2003, pl. 5, figs 83–88) and Bellacartwrightia sp. (Eldredge, 1971, pl. 14, fig. 6), for instance, show that FAIs can be well-developed ventrally even if they cannot be recognised dorsally on the cuticle.
6.2. Allometric growth
63The development of trilobites, like many extant arthropods, has been shown to be hemianamorphic (Minelli et al., 2003; Hughes et al., 2006). After the anamorphic release of trunk segments, stabilisation of thoracic and pygidial segment counts early in the holaspid period marked the beginning of the epimorphic stage involving considerable allometric growth. The number of holaspid instars is generally difficult to determine and unknown in the vast majority of trilobite species. It is also often unknown whether terminal anecdysis occurs. The abundant articulated and uncompacted Hollardops specimens reported by Brett et al. (2012) from the basal part of the Khebchia Formation could offer a valuable opportunity to identify instars. This would be pertinent to assessing the intraspecific variation of morphological traits observed in Hollardops.
64Of the three H. multatuli specimens collected from a single bank in Tadachacht, paratype NHMM 2022001 is of extraordinarily large size for a Hollardops (Fig. 10). There is a plethora of parameters that can affect body-size variations, such as temperature, oxygenation, nutrient availability and predation (e.g. Saleh et al., 2020) but many of these were probably homogeneous for these three H. multatuli specimens. The large paratype specimen shows morphological differences compared to smaller ones, including the courses and lengths of the lateral glabellar furrows, the distance between eye and posterior border, the width (exsag.) of the pygidial pleural furrows, the shape of the anterior pygidial axial rings, and the shape and size of the pygidial lappets. Such variations imply that these characters should be used with caution in taxonomic descriptions. The limited number of specimens available to us does not permit any further discussion of this topic. We have based our descriptions and comparisons on regular-sized specimens, generally for the genus, or as far as comparative sizes among individuals of the same species could be estimated.

Figure 10. Relative sizes of Hollardops specimens. A–C. Hollardops multatuli sp. nov. A. NHMM 2022001. B. IRSNB a13567. C. IRSNB a13568. D. Hollardops kyriarchos sp. nov., IRSNB a13569. E. Hollardops angustifrons sp. nov., IRSNB a13570. F. Hollardops boudibensis (Morzadec, 2001), IRSNB a13571. G. Hollardops luscus sp. nov., NHMM 2021043.
Scale bar is 10 mm.
6.3. Tubercle distribution
65A striking aspect of Hollardops is the distinctive distribution and size of tubercles. Tubercles are often larger or more densely spaced in certain areas of the cephalon and the thorax. We suspect that this is a potential feature to discriminate species. Tubercle distribution patterns of selected Hollardops species are shown in Figure 11. These highlight the areas that are most commonly covered with comparatively large tubercles. Obviously, a comprehensive analysis requires predefined criteria and landmarks as well as larger amounts of specimens from a single population. This is beyond the scope of the present work.
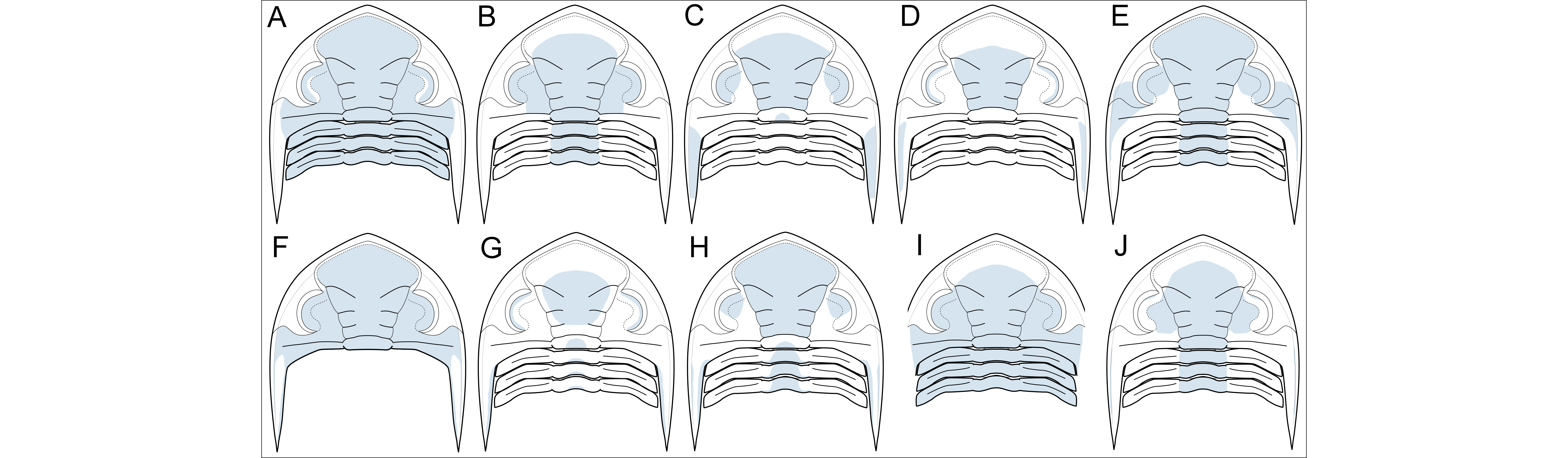
Figure 11. Tubercle distribution (highlighted in blue) in selected Hollardops species from Morocco and Algeria. A. Hollardops angustifrons sp. nov. B. Hollardops mesocristatus of Chatterton et al. (2006). C. Hollardops hyfinkeli (Lieberman & Kloc, 1997). D. Hollardops burtandmimiae (Lieberman & Kloc, 1997). E. Hollardops multatuli sp. nov. F. Hollardops lemaitreae Morzadec, 1997. G. Hollardops boudibensis Morzadec, 2001. H. Hollardops sp. A. I. Hollardops klugi sp. nov. J. Hollardops luscus sp. nov.
6.4. Eye lens numbers
66The numbers of lenses in the visual surface are known to vary within phacopide species and the same intraspecific variation may be expected to occur in Hollardops. Lens counts are expressed in the maximum of dorsoventral files and the maximum lenses per file. Specimens that we would consider including in H. hyfinkeli can have 29/9, 29/10 or 30/10 lenses, and H. burtandmimiae may have 27/7 (AMNH 45266) or 29/7 (AMNH 45267, 45272), rarely with a small 8th lens developed (AMNH 45271). Hollardops multatuli is known from three specimens, the smaller two of which have 30/9 whereas the exceptionally large paratype has 31/11 lenses. The other species recorded here are known from few specimens. Hollardops boudibensis has 27/7 lenses, H. kyriarchos has 29/9, H. luscus has 28/8 and H. angustifrons 28/7.
7. Systematics and the phylogenetic affinities of Hollardops
67Subsequent to their rise, presumably in the Silurian, members of Asteropyginae became regular components of Early to early Late Devonian trilobite faunas in the Rhenish-Bohemian Province. According to Morzadec (1992) the group slowly evolved during the Lochkovian and the Pragian, followed by accelerated diversification in the Emsian. However, we believe that Pragian to Emsian diversity, at least, is not particularly well resolved and that equivocal low-rank taxonomy is largely to blame for this. The main issue is that a number of “classic” (i.e., regularly reported, widely geographically and stratigraphically distributed) and other species from the Lower Devonian of Europe are known exclusively from inadequately preserved, poorly illustrated specimens. Type series are not rarely comprised of sclerites from different localities and (vaguely defined) stratigraphic intervals, and united into single polymorphic species. These defects become problematic when the type species of genera are affected that have often been used to accommodate taxa (e.g. Metacanthina Pillet, 1954, Pseudocryphaeus Pillet, 1954, Pilletina Haas, 1970, Paracryphaeus Gandl, 1972, Cryphina Oehlert & Oehlert, 1889) (see Pillet, 1973 for photos of all of these). Other genera (e.g. Echinopyge Haas, 1968 and Feruminops Haas, 1968 from the Asian part of Türkiye, and Delocare Struve, 1958 from Germany) have been largely ignored and asteropygine systematics insufficiently integrated with palaeogeography. This has led to divergent standpoints of workers and unstable generic concepts. Solving all these issues is beyond the scope of the present work and for obvious reasons we have been conservative with the classifications of taxa.
7.1. Proposed, accepted and rejected synonymies
68Morzadec (1997) diagnosed Hollardops as having an anteriorly strongly expanded glabella, long genal spines, 10 thorax segments, and a pygidium with 10–14 axial rings, boothi-type pleural segmentation, wide pleural furrows, and five pairs of short, tongue-shaped lateral spines with flat ventral sides. Above all, Morzadec (1997) attributed great significance to the number of thoracic segments of Hollardops which he regarded as a unique condition within the Asteropyginae. Besides the type species, Asteropyge mesocristata, he included in Hollardops: Greenops? struvei Morzadec, 1969 from Brittany and Hollardops lemaitreae Morzadec, 1997 (thorax unknown) from Algeria.
69In the same year, Lieberman & Kloc (1997) erected Philipsmithiana (type species: Philipsmithiana hyfinkeli Lieberman & Kloc, 1997 from southern Morocco). Morzadec (2001) regarded P. hyfinkeli as a synonym of H. mesocristatus. This led to the junior synonymy of Philipsmithiana (the publication of Hollardops preceded that of Philipsmithiana in the same year and thus the former has priority). In our opinion, P. hyfinkeli is rightly included in Hollardops; its synonymy with H. mesocristatus, however, is rejected and dealt with below.
70Lieberman & Kloc (1997) also erected the monotypic Modellops and selected Greenops? struvei as the type species. At the outset, this species was described by Morzadec (1969) as having 11 thoracic segments. Although Morzadec (1969, 1983) listed complete specimens among his examined material, he did not illustrate these. Later, Morzadec (2001) stated that G.? struvei has 10 segments which led him to regard Modellops as a junior subjective synonym of Hollardops. Bignon & Crônier (2013), in performing a cladistic analysis of Asteropyginae, could neither corroborate nor reject the placement of G.? struvei in Hollardops but concluded that they were inclined to leave it there (we are in agreement with this decision). A comprehensive revision of Hollardops struvei would be timely. Bignon & Crônier (2013) already made note of considerable intraspecific variability among specimens re-examined by them. According to Morzadec (1983) this species occurs in the Le Fret through Bolast formations (serotinus to patulus Conodont zones), and thus, a large part of the upper Emsian of Brittany. Such a long stratigraphic range is unusual for an asteropygine species. It is furthermore uncertain whether all specimens assigned to H. struvei at present actually have 10 thoracic segments. Morzadec (2001) did not explain his adjusted segment count (10, instead of 11 as stated in his original description from 1969); hence, in the absence of photographic evidence of a topotypical complete H. struvei specimen, the condition of 10 thorax segments remains uncertain. The only published photos of articulated, potential representatives of this species were provided by Hoffman et al. (2014, p. 54, figs 3, 5), who recorded two internal moulds with 10 thoracic segments, from the uppermost Emsian Kerdréolet and Bolast formations. The poor state of preservation of those specimens precludes comparisons with the cephala and pygidia recorded by Morzadec (1969, 1983). That is to say, collectively, the material from Brittany may represent more than one species.
71Schraut (2000) doubted the independent generic rank of Hollardops, claiming that it is exceedingly similar to Greenops and that only H. mesocristatus is certain to have 10 thoracic segments. Schraut did not mention Philipsmithiana or Modellops, apparently not having noticed the paper by Lieberman & Kloc (1997). Schraut furthermore suggested that Greenops, like Hollardops, may have 10 thoracic segments although he did not provide a reason for this. We do not subscribe to any of Schraut’s ideas. As to our knowledge, no other worker has ever questioned the validity of Hollardops and not a single Greenops species is known from large holaspides with less than 11 thorax segments. The phylogenetic analysis of Bignon & Crônier (2013) did not support a close relationship between Greenops and Hollardops. Because Schraut (2000) was not specific about the shared characters between these genera we regard the similarities as superficial.
72Bignon & Crônier (2013) erected Pennarbedops (type species: Rhenops babini Morzadec, 1983 from the upper Emsian of Brittany) and also included Rhenops circumapodemus Smeenk, 1983 from the upper Emsian of the Cantabrian Mountains in their new genus. Bignon & Crônier (2013) coded P. babini for analysis as having 11 thorax segments. However, no worker seems to have noticed that the single articulated complete specimen of P. babini figured by Morzadec (1983, pl. 7, fig. 3) has only 10 thoracic segments. The same is true for the holotype of P. circumapodemus figured by Smeenk (1983, pl. 9, fig. 8) and for putatively congeneric specimens from the upper Emsian of the Cantabrian Mountains figured by Guy (2020, fig. 5) and herein (Fig. 12). Both Pennarbedops species share a comparatively long fifth pygidial pleural lappet pair, or true pleural spines, but that feature is known to vary significantly between species in other asteropygine genera (e.g. van Viersen & Vanherle, 2018). Additionally, the fifth pleural lappet pair of H. burtandmimiae is also rather long (Fig. 16B, H). It has been argued above that the exceedingly spiny condition of former Pennarbedops species and H. burtandmimiae is a coaptative device designed to support the retracted pygidium, allowing the long pleural spines to rest on the cephalic doublure. None of the diagnostic characters mentioned by Bignon & Crônier (2013) render their genus distinct from Hollardops or its putative synonym Modellops. We prefer not to recognise Pennarbedops.

Figure 12. Hollardops cf. circumapodemus (Smeenk, 1983), Lezna Member of the Abadía Formation (upper Emsian), Polentinos, Spain. External mould of an articulated specimen in the collection of F. Collantes.
Scale bar is 10 mm.
7.2. The 10-segmented state
73The phylogenetic importance of thoracic segment counts in Asteropyginae is somewhat equivocal even if only for the principle argument that it is unknown in the majority of species. Struve (1959a) erected the Ordovician–Devonian Phacopina which he typified as having 11 thorax segments. Silurian dalmanitids and Early Devonian asteropygines that are known from complete specimens all have 11 segments (e.g. Bignon & Crônier, 2013; Bignon et al., 2014) and it stands to reason that this count is primitive for the subfamily. The 10-segmented state of species included in Hollardops at present is likely to imply recent common ancestry in view of their similar morphologies and overlapping stratigraphic and palaeogeographic distributions.
74Whiteley et al. (2002, pl. 40) illustrated a member of an undescribed asteropygine genus with 10 thoracic segments from the Givetian of Eastern North America and one of us (GK) has both 10- and 9-segmented specimens in his possession. Those are morphologically rather dissimilar from Hollardops species and no intermediary forms from the Eifelian are known.
75Chatterton et al. (2006) reported 10 thorax segments for their new species Bignonops tamnrhertus but they did not discuss the possible implications of this lower amount. It appears that Chatterton et al. (2006) had inadvertently based their count on the disarticulated paratype (ibid., pl. 19, figs 1–4, 6) which almost certainly has the 11th segment missing or concealed under the 10th segment. As far as can be observed in their photos, all the other specimens, including the holotype (ibid., pl. 21, figs 4, 7, 9–11), have 11 thorax segments.
7.3. Origins of Hollardops
76There is a group of asteropygines from the lower Emsian of North and peri-Gondwana that may be ancestral to Hollardops. Haas (1968) erected Feruminops (type species: Feruminops crepidus Haas, 1968 from the lower Emsian of Türkiye; see Haas, 1982 for revised stratigraphy) based on cephala alone. Haas (1968, p. 133) mentioned that cephala of F. crepidus and pygidia of Metacanthina acinacifera Haas, 1968 are by far the commonest trilobites in the type locality. He considered the possibility that the two are conspecific but nonetheless favoured assignment of his pygidia to Metacanthina because a single cephalon of Metacanthina had also been found. The generic concept of Metacanthina sensu Haas (1968) was revised subsequent to the analysis of Bignon & Crônier (2013), who no longer listed M. acinacifera as one of its species (they regarded Metacanthina as monotypic). This leaves the most parsimonious option that Haas’ pygidia of M. acinacifera belong to F. crepidus. Of note is that the cephala and pygidia of Feruminops crepidus (Haas, 1968, p. 132, fig. 34, p. 141, fig. 39, pl. 34, figs 1–3, 15, 16), in a revised sense, are exceedingly similar to Pilletina zguidensis Morzadec, 2001 (Morzadec, 2001, pl. 4, figs 1–4) from the lower Emsian of southern Morocco, with only minor differences in the eye lens count, the more prominent median occipital node, and the abaxial depth of S2 (all three characters are variable in other asteropygine genera). Thus, the similarities to P. zguidensis, which is known from articulated material, appear to be consistent with the proposed association of the cephala and pygidia of F. crepidus here. Bignon & Crônier (2013) rejected Morzadec’s (2001) generic concept of Pilletina and selected P. zguidensis as the type species of their new genus Morzadecops which, in their cladistic analysis, was resolved as the sister group of Pennarbedops (a genus considered by us to be synonymous with Hollardops). Bignon & Crônier (2013) diagnosed Morzadecops as having frontal lobe diamond shaped; axial furrows curved between S2 and S3; L3 more elevated than L1 and L2; 8 lenses per dorsoventral file of visual surface; genal spine shorter than glabella, without development of lateral border; 10 to 11 pygidial axial rings; 5 pleural segments; rounded pleural bands; pleural furrows rather large; 5 pairs of pygidial spines developed from posterior and anterior pleural bands with a similar length, shorter or as long as pleural width (tr.); terminal spine as wide as axial extremity and as long as other pygidial spines. Bignon & Crônier (2013) did not compare Morzadecops with Feruminops and the listed diagnostic features are insufficient to discriminate the two. Because of the minor differences between their type species we regard Morzadecops as a junior synonym of Feruminops.
77Heretofore, all Hollardops species were recorded from the upper Emsian, with the exception of the earliest Eifelian H. aithassainorum Chatterton et al., 2006. We propose Hollardops klugi sp. nov. for asteropygines with 10 thorax segments from the lower Emsian of Ouidane Chebbi that were identified by Klug et al. (2008, pl. 11, figs 4–7, 13–16) as Pilletina zguidensis. Hollardops klugi is the oldest known member of the genus and so it might be expected to possess a less derived morphology than stratigraphically younger species. At the same time, H. klugi shows putative juvenile features (e.g. the 10 thorax segments). This underlines once more the potential paedomorphic origin of Hollardops and complicates comparisons with other Early Devonian asteropygines because their ontogenies are as good as unknown. Accepting the 10-segmented condition as an apomorphy of Hollardops, this state must have been achieved by the release of one less segment into the thorax of an ancestor with 11 segments in Asteropyginae. Additionally, H. klugi carries median nodes on the occipital and thoracic and pygidial axial rings. Median nodes are shown by a degree 10 meraspid of the Middle Devonian Greenops grabaui (Whiteley et al., 2002, pl. 53). A conspecific, large holaspis illustrated by Whiteley et al. (2002, pl. 54) reveals that only the occipital node is retained into adulthood. A large holaspis of another, undescribed Greenops species (Whiteley et al., 2002, pl. 56) carries nodes posteromedially on most axial rings. This specimen shows other juvenile characters shared with the meraspid of G. grabaui, including a comparatively small pygidium and reduced genal spines. However, drawing analogues between the character states of Greenops and Hollardops may be precarious since the two clades are phylogenetically distantly related (see Bignon & Crônier, 2013). The slightly shorter pygidium of H. klugi compared to congeners should probably be regarded as a plesiomorphy. Other Hollardops species have large pygidia and it would seem likely that this feature was under selective pressure if it facilitated improved coaptation in harmony with 10 thoracic segments. Nonetheless, a pseudomegalaspid type of coaptation, with the anterior cephalic border pushed against the ventral pygidial rim, was already practised by H. klugi (Fig. 21H, M). An increase of pygidial size was achieved in stratigraphically younger Hollardops species through the accumulation of pygidial axial rings and changes in ontogenetic allometry (e.g. increased width (exsag.) of the pleural furrows). The early Emsian Feruminops possesses median nodes throughout the axis and comparatively short genal spines. Thus, Feruminops itself shows potentially paedomorphic features. It is conceivable that Feruminops and Hollardops belong to a paedomorphocline and that the latter evolved from an arrested degree 10 meraspid ancestor in Feruminops during the lower Emsian. This would have required only few character state changes (see codings of Bignon & Crônier, 2013).
8. The oldest Asteropyginae
78Asselberghs (1930) described cranidial and pygidial remains of an acastid from the Gdoumont sandstone (Waimes [German: Weismes] Member of the Marteau Formation) in the Hohes Venn, East Belgium, as Asteropyge gdoumontensis. The age of the Gdoumont sandstone has been much debated on with opinions ranging from the middle Ludlow to upper Lochkovian. Richter & Richter (1954) and Richter (1964) reviewed papers dealing with this topic and concluded that the Gdoumont sandstone belongs to the upper Lochkovian. Morzadec (1990, 1992) and Bignon et al. (2014) assumed a lower Lochkovian age but did not explain their standpoints. Godefroid & Cravatte (1999), Schemm-Gregory & Jansen (2009) and Jansen (2016), however, provided compelling biostratigraphic evidence that is consistent with a Pridolian age of the Gdoumont sandstone and this is accepted herein.
79Asselberghs (1930, pl. 6, figs 6–9) figured a cranidium and three pygidia from the type locality whilst only briefly describing these. Richter & Richter (1942) united cranidia from two nearby localities with the material of Asselberghs (1930) and expanded the original description notably with pygidial characters, including 9–10 (+ ? 1–2) axial rings, six pairs of pleurae with enough space for a seventh pair, seven pairs of pleural spines and a large median spine that is broader and longer than the adjacent lappet pair. Richter & Richter (1942) selected pygidium IRSNB a3269 (Asselberghs, 1930, pl. 6, fig. 7; see Fig. 13A, B for new photos) as the lectotype since Asselberghs had not designated a holotype. Richter & Richter (1942, p. 159, fig. 5; repeated here as Fig. 13C) figured the lectotype and apparently retouched the area of the pygidial margin in the photograph in order for it to show seven distinct lateral spine pairs and a rather long median spine. One of us (AV) has examined Asselberghs’ types in the Maillieux collections of the Institut royal des Sciences naturelles de Belgique (IRSNB) (Fig. 13) and was unable to confirm these features. The internal and external moulds of the lectotype (Fig. 13B) are poorly preserved in a coarsely grained siliciclastic stone and reveal little to no information on the morphology of the spines. The contour of the external mould (Fig. 13A) appears to have been accentuated by red paint, probably with the aim of increasing contrast for the photos used by Asselbergs (1930). The posterior region of the pygidium is damaged and parts are missing, including the posteriormost spine pairs and the median spine. The seventh right pleural spine indicated by Richter & Richter (1942) is located postaxially and thus, more likely represents a misapprehended remnant of the base of a large median spine (cf., e.g., the spiny structure just adaxial to the right sixth pleural lappet of the paralectotype internal mould in Fig. 13E: we suspect that this information was used by the Richters to enhance their photo of the incompletely preserved lectotype). We advise against the use of the photo of the lectotype by Richter & Richter (1942, p. 159, fig. 5) because it is misleading.
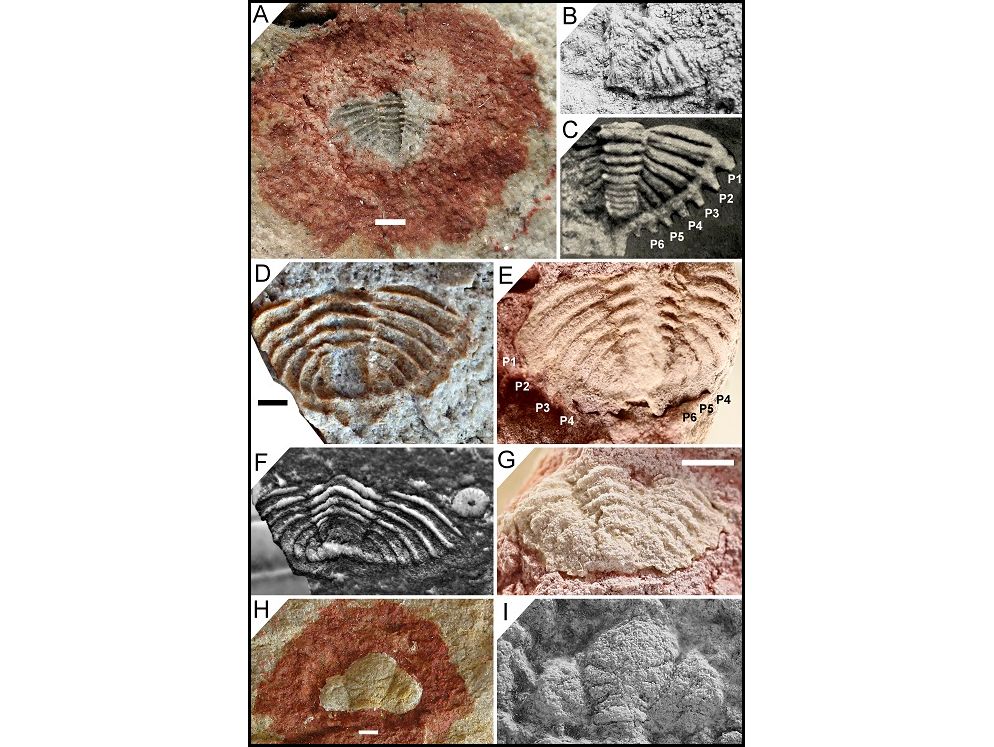
Figure 13. Ganetops gdoumontensis (Asselberghs, 1930), Gdoumont sandstone (Pridolian), Gdoumont, Belgium. A–C. Lectotype, IRSNB a3269, external mould in dorsal view (A), internal mould in oblique lateral view (B), retouched image, mirrored along sagittal line, from Richter & Richter (1942, p. 159, fig. 5) (C). D–F. Paralectotype, IRSNB a3271, external mould (D) and internal mould (E) in dorsal view, and external mould in oblique posterior view (digitally inverted image) (F). G. Paralectotype, IRSNB a3270, internal mould of a pygidium in oblique posterior view. H, I. Paralectotype, IRSNB a3268, internal mould of a cranidium in dorsal view.
Numbering in C and E indicates pleural spine numbers. Scale bars are 2 mm.
80Richter & Richter (1942) provided reasons for the tentative inclusion of A. gdoumontensis in Cryphina. Haas (1968) underlined the provisional character of this assignment, noting that A. gdoumontensis has ‘true’ asteropygine pygidial spine pairs and a long median spine. Morzadec (1990) retained A. gdoumontensis as a questionable Cryphina species although he considered that only a complete revision of this species will enable a definite generic assignment. Nevertheless, Morzadec (1990) typified A. gdoumontensis as having 10 axial rings, six pairs of pleurae and seven pairs of lateral spines, while adding that the holotype is a juvenile specimen. We are assuming that Morzadec meant to refer to the lectotype pygidium, yet in the absence of argumentation for his statement, we fail to see why its morphology should be regarded as juvenile. The lectotype is neither smaller than the paralectotype pygidia nor is it principally different.
81Bignon et al. (2014) repeated the characters listed by Morzadec (1990) and concluded that the segment count is rather high compared to the earliest Asteropyginae. According to Bignon et al. (2014), Asselberghs’ material is poorly preserved making the pygidial segmentation and the connection with the pygidial spines difficult to observe, but they noted a very faint connection with the spines or no connection at all, rendering A. gdoumontensis close to Protacanthina (an acastavine). Bignon et al. (2014) did not mention the additional cranidia described by Richter & Richter (1942); those specimens lack characters considered by Bignon & Crônier (2013) to be diagnostic of Protacanthina, including axial furrows straight between S1 and S3, and eyes, not in contact with posterior border furrow or S3. None of the pygidia of A. gdoumontensis allow the confirmation of the long median spine that was claimed by Richter & Richter (1942) and which, according to Bignon & Crônier (2013), is also diagnostic of Protacanthina.
82Straw (1933, pp. 127, 128, pl. 10, fig. 7) recorded a pygidium of Calmonia sp. along with other trilobite remains from the Little Missenden borehole in Buckinghamshire, SE England. Richter & Richter (1954) included Straw’s pygidium in A. gdoumontensis but failed to mention that Straw (1933, p. 128) reported only 6 pleural spine pairs as well as a short median spine. Richter & Richter (1954) suspected that Straw’s (1933, pl. 10, fig. 9) putative dalmanitid cranidium also belongs to A. gdoumontensis but Straw’s description is rather superficial and his figure shows an unidentifiable fossil. Thomas et al. (1984) and Morris (1988) accepted the conspecific relationship between the British and Belgian specimens and the tentative placement in Cryphina as proposed by the Richters, while underlining the uppermost Silurian age of this species. Finally, Storey (2012, pl. 19, fig. 7) provided a contemporary photo of a pygidium from the Little Missenden boring that allows five, possibly six pairs of pleurae and five spine pairs to be discerned, leaving room for a potential sixth pair of spines (this area of the pygidium is not well preserved).
83Problems with the generic concept of Cryphina have been briefly pointed out above and we do not advocate its usage. Pillet (1973) redescribed the type species, C. andegavensis Oehlert, 1889 from Brittany. Pillet united, somewhat arbitrarily, poorly preserved cephalic and pygidial remains from various localities in the Massif armoricain with the neotype from the upper Emsian. The similarities between C. andegavensis and A. gdoumontensis that were claimed by Richter & Richter (1954) are either incorrect or superficial. There is a large stratigraphic gap between both species and no intermediate forms are known. We therefore remove A. gdoumontensis from Cryphina.
84The types in the collections of the IRSNB reveal that A. gdoumontensis has no more than nine identifiable pygidial axial rings, six pairs of pleurae, and six pairs of lateral spines. There is a broad, distally somewhat upturned, pointy median spine which is about twice as long as the pleural spines. Of note are similarities of one of the paralectotype pygidia (Fig. 13D) and especially its median spine, to pygidia of Treveropyge djemelensis Morzadec, 1997, from the Lochkovian of Algeria (cf., e.g., Morzadec, 1997, pl. 1, fig. 6a). Morzadec (1997) reported nine pygidial axial rings and five pleural and spine pairs for his species. One of the Algerian pygidia (Morzadec, 1997, pl. 1, fig. 6) has the cuticle partially preserved and shows six pairs of pleurae, but a sixth pleural lappet pair is not discernible. However, one of the internal moulds (ibid., pl. 1, fig. 4) shows a poorly developed sixth pair of pleurae and, connected to each of these, a very weak ridge on the posterior border, indicating the onset of a rudimentary sixth lappet. Bignon et al. (2014) erected Ganetops (type species: Asteropyge (Rhenops?) ebbae Richter & Richter, 1954 from the Lochkovian of the Sauerland, Rhenish Mountains) and transferred Treveropyge djemelensis to it. As far as the cranidial and pygidial remains of A. gdoumontensis can be compared, they adhere to the generic diagnosis of Bignon et al. (2014) except for three features: (1) axial furrows straight between S1 and S3—these appear to be convex in the cranidia of G. ebbae figured by Richter & Richter (1954, pl. 6, figs 83–88) except in places where they are tectonically deformed; (2) no contact between S2 and axial furrows—this character occurs on the internal moulds of G. ebbae but not on the single external mould illustrated (Richter & Richter, 1954, pl. 6, fig. 85); (3) five pairs of pleural segments and spines—as concluded above, G. djemelensis has rudimentary sixth pleural and spine pairs. If our reassignment of Ganetops gdoumontensis comb. nov. to this genus is correct then it is, once more, the stratigraphically oldest known asteropygine and the five pleural and spine pairs of younger Ganetops species a putatively derived feature. The origins of Ganetops and Asteropyginae remain difficult to determine without complete specimens or the prospect of a potential ancestor.
9. Openings of the exoskeleton
9.1. Fenestrae
85Some trilobites had fenestrae, i.e., slit-like to circular openings in the mineralised exoskeleton of the post-cephalic pleural regions. Holloway & De Carvalho (2009) reviewed the occurrences and possible functions of this feature in the dalmanitid Fenestraspis and members of Acastidae and Odontopleuridae. They considered that fenestrae may have been actual openings or covered with a membrane in life, and that possible functions were (1) respiration by circulation of oxygenated water (or diffusion of oxygen through a permeable membrane) during enrolment, (2) the accommodation of some sensory organ, or, less likely, the reduction of (3) weight or (4) volume of the mineralised exoskeleton. Holloway & De Carvalho (2009) concluded that apart from their problematic function, it is difficult to reconcile the presence of fenestrae in some trilobites with their absence in closely related taxa.
86There appears to be no elegant universal solution for the problem. A respiratory function was superfluous to trilobites that were unable to tightly enrol or that may not have employed coaptation as an important defensive strategy (e.g. Comura, Laethoprusia, Heliopyge). If the above examples of respiratory devices in dalmanitids, cheirurids, encrinurids, scutelluids, homalonotids and proetids are all correct, then a large total surface area of openings in the thoracic and pygidial pleurae, e.g. in Fenestraspis, was not necessary for them to be effective unless they were covered with a membrane. In species with very small fenestrae, on the other hand, true openings probably would have just sufficed to let enough water pass through.
87An interesting case is presented by members of Asteropyginae from the Belgian middle Frasnian carbonate platform where pygidial fenestrae evolved, independently, in several lineages and environments (van Viersen & Bignon, 2011; van Viersen & Vanherle, 2018). Their patterns, however, are erratic in that sizes and shapes are axially asymmetric and even their presence is variable intraspecifically. Species of Heliopyge occur in Belgium both in shales and limestones. The holotype pygidium of Heliopyge helios (Richter & Richter, 1926) (Fig. 14E, F, H) has sizable interpleural embayments extending across the pleural fields and nearly continuous with large lateral inter-ring openings. Towards the posterior end of the pygidium, axial asymmetry of the fenestrae is increasingly manifested. Topotypical pygidia of another asteropygine, Quadratispina excelsa van Viersen & Vanherle, 2018, which is restricted to limestones, variably exhibit large, axially asymmetric (especially posteriorly on the pygidium) openings (Fig. 14A), or they are small (Fig. 14C, D) or absent altogether (Fig. 14B). Such variations more or less rule out a sensory or respiratory function that would be expected to have been intimately linked with the animal’s behaviour and to have required more consistent fenestral patterns, both in terms of their axial symmetry and presence within populations or species. The low numbers of specimens under study here may render these conclusions premature but published specimens of other asteropygines, all from the uppermost Givetian to Frasnian, also show intraspecific variations of the fenestrae, including Radiopyge sardarensis Morzadec, 2002 (Morzadec, 2002, p. 425, fig. 9.5, 9.6, 9.8–9.15) and “Heliopyge” sohensis Ghobadi Pour et al., 2013 (Ghobadi Pour et al., 2013, p. 155, fig. 4B–H) from Iran. Bradocryphaeus neptuni van Viersen & Bignon, 2011 (Fig. 14G) occurs both in shales and limestones in Belgium; it has exceedingly small fenestrae that are only developed adaxially as narrow (exsag.) openings. Pygidia of B. neptuni and Q. excelsa can show openings abaxially in the inter-ring furrows; these are evident in two specimens of the latter species, by the convex posterior recesses abaxially on the axial rings (Fig. 14A, C).
88Biomineralisation of the arthropod exoskeleton generally comes at high energy costs. Prevalent gaps like the ones of H. helios and Q. excelsa would have compromised sturdiness of the exoskeleton and reduced its effectiveness against predator attacks. If structural integrity became of lesser importance as a result of behavioural changes (e.g. the abandonment of coaptation as a defensive strategy) then energy could be diverted to more important functions. The openings in the pleural and axial regions probably enabled average holaspid growth increments and it is evident that normal tagma size was achieved in adulthood, but with a smaller energy demand. We envisage that as the new exo-cuticle was stretched and expanded along the openings directly after shedding of the moult, the size of the axis and pleural regions was increased. It is unlikely, however, that fewer moults were required to reach adulthood because that would probably have induced a more juvenile cephalic morphology. This seems not to be the case.
89Another noteworthy aspect of the examples in Asteropyginae from the Belgian Frasnian is that not only the openings are axially asymmetric, but parts of the pygidium are sometimes skewed beyond extents that are normally referable to tectonic deformation (cf. Fig. 14E and digitally enhanced alternative 14H). The median spine of the holotype pygidium of H. helios stands left of the sagittal line. The cuticle around the fenestrae of this same specimen projects dorsally higher on the left pleural region than on the right pleural region of the pygidium (Fig. 14F). The asymmetric pleurae are also shown by Q. excelsa (Fig. 14A–D).
90It is possible that the pygidial openings of H. helios and Q. excelsa were entirely involuntary. Holloway & De Carvalho (2009) did not favour the notion of reduced volume of the mineralised exoskeleton in response to carbonate deficient conditions because trilobites with fenestrae occur both in environments rich and poor in carbonate. The examples of asteropygines from the Belgian Frasnian are not inconsistent with this argumentation. But perhaps access to carbonate material did not always imply that it could be used readily. According to Whiteley (2011) the acidification of contemporary oceans by the absorption of anthropogenic carbon dioxide is known to impact calcification and growth rates of many exclusively marine crustaceans, but especially species that are poor iono- and osmoregulators with limited abilities to compensate for acid-base disturbances. Whiteley concluded that the ability to tolerate oceanic acidification is species specific and varies within phyla and even between closely related species. Admittedly, the parallel with trilobites is notional, yet the distribution, size and asymmetry of the openings, at least in the Belgian asteropygines under consideration here, are so arbitrary that the fenestrae cannot be assumed to have been under strict genetic control. An extrinsic cause (e.g. distress from changing environmental conditions), thus, remains a very viable explanation.
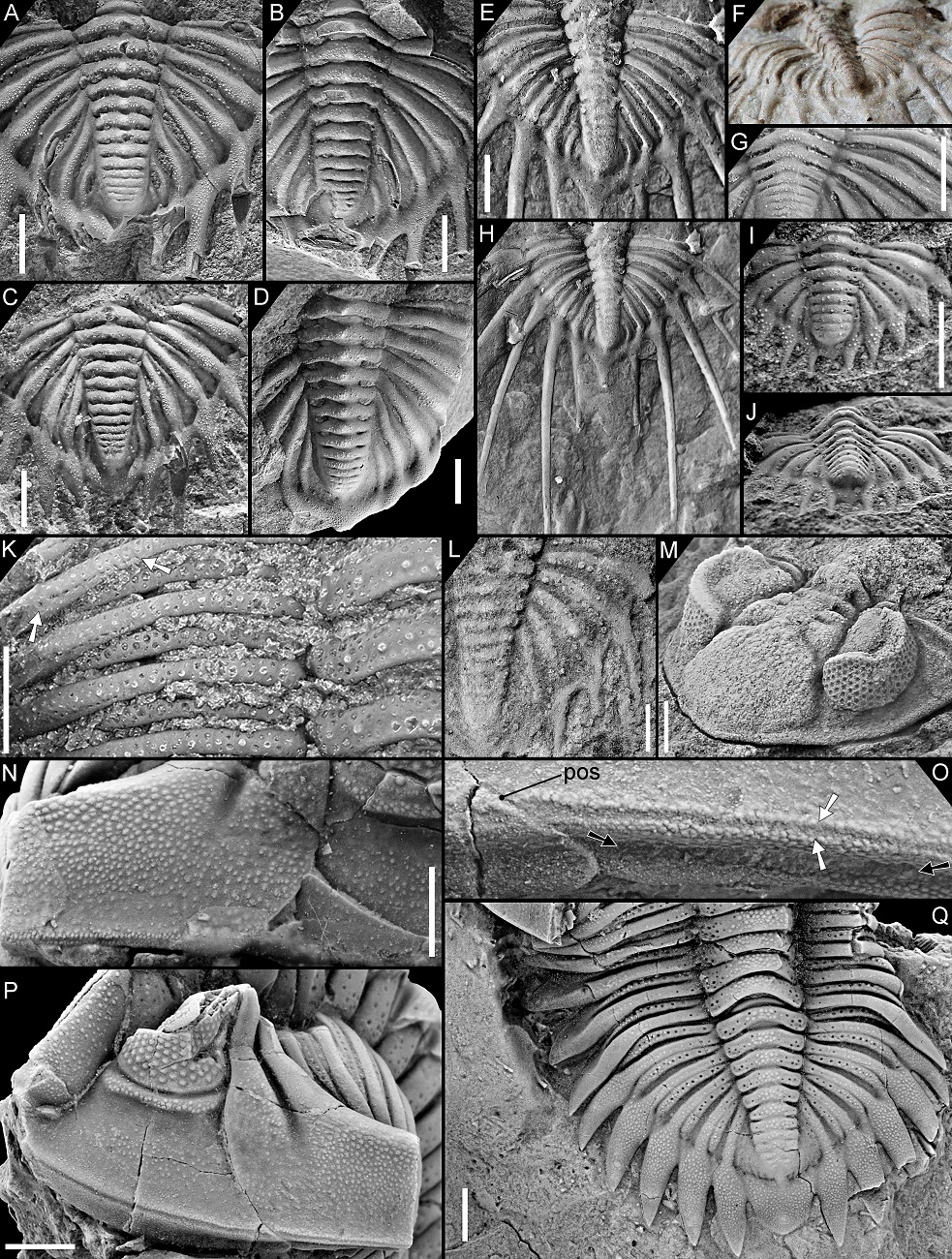
Figure 14. A–D. Quadratispina excelsa van Viersen & Vanherle, 2018, Bovesse Formation (middle Frasnian), Emines, Belgium. A. Holotype, IRSNB a13196. B. Unnumbered specimen, Coll. Piret IRSNB I.G. 9694. C. Paratype, IRSNB a13200. D. Unnumbered specimen, Coll. Piret IRSNB I.G. 9694.
E, F, H. Heliopyge helios (Richter & Richter, 1926), Ermitage Member, Moulin Liénaux Formation (middle Frasnian), Nismes, Belgium. Holotype (external mould), IRSNB a7808, in dorsal (E) and oblique posterolateral (F) views (both digitally inverted), and same specimen after digital correction of tectonic skewing (H).
G. Bradocryphaeus neptuni van Viersen & Bignon, 2011, Boussu-en-Fagne Member, Grands Breux Formation (middle Frasnian), Lompret, Belgium, IRSNB a13170.
I, J. Gudralisium rossumi van Viersen, 2015, Hanonet Formation (Eifelian–Givetian transition), Boutonville, Belgium. Holotype, NHMM 2015009, in dorsal (I) and oblique posterior (J) views.
K. Asteropyge boeckae van Viersen, Taghon & Magrean, 2019, Vieux Moulin Member of the Jemelle Formation (lower Eifelian), Vierves, Belgium. Holotype, IRSNB a13272, close-up of thoracic axial rings 7 to 9 and associated left pleurae. Arrows indicate row of posterior pits.
L, M. Jaylopygiella debruni van Viersen, 2013, Rivière Formation (Eifelian), Kin, Belgium. L. SMF 79526a, internal mould of pygidium on same rock slab as holotype. M. Paratype, SMF 79525, internal mould of cephalon.
N–Q. Greenops widderensis Lieberman & Kloc, 1997, presumably from the Widder Formation, Hamilton Group (Givetian), Hungry Hollow, Arkona, Canada. Complete specimen in the collection of the first author, in lateral view of right fixigena (N), close-up of base of left genal spine and doublure (O), oblique left view of cephalon (P) and dorsal view of pygidium (Q). Postocular suture indicated by “pos”. White arrows indicate supramarginal and submarginal tubercle rows; black arrows indicate broad ventral furrow dividing the lateral doublure into an adaxial and an abaxial band (subexsag.).
Scale bars are 2 mm.
9.2. Perforations
91Holloway & De Carvalho (2009) reviewed the exoskeletal perforations (i.e., small, circular openings or holes extending through the mineralised cuticle) of Asteropyge, Bellacartwrightia, Greenops, among other asteropygines, and concluded that these are principally different from fenestrae in being smaller and not restricted to intersegmental boundaries. Published specimens from several asteropygine genera offer clues to suggest that fenestrae and perforations have unique origins. The perforations of Greenops widderensis (McNamara & Tuura, 2011, p. 107, fig. 1; Fig. 14P, Q) are predominant on the anterior bands of the thoracic and pygidial pleurae, the abaxial parts of the axial rings, and on the lateral glabellar lobes. Perforations are nearly ubiquitous on a pygidium of “Comura” praecursor Pillet, 1961 (Pillet, 1961, pl. 2, fig. 9) where they occur even on the hollow(?) bases of the pleural spines. The holotype pygidium of Gudralisium rossumi van Viersen, 2015 (Fig. 14I, J) shows similar perforations, abaxially on some of the axial rings, in the pleural furrows, on the second, third and fourth anterior pleural band pairs, on the third left and fourth right posterior pleural bands, and on the fifth pleural pair; their asymmetries, both in presence and size left and right of the sagittal line, may appear erratic like those of fenestrae but the affected areas, especially on the left side of the pygidium, are less well preserved. Perforations are pervasive on the pygidial axial and pleural regions of Jaylopygiella debruni van Viersen, 2013 (Monseur, 1959, pp. 92, 97, figs 5, 6; van Viersen, 2013, p. 8, fig. 6E) and Asteropyge filoxenia van Viersen et al., 2019 (Fig. 9A). Asteropyge boeckae (Fig. 14K) bears a mixture of perforations and tubercles: there are comparatively large holes on the anterior pleural bands and intercalating tubercles and holes on the posterior pleural bands, on thorax and pygidium alike. This pattern continues anteriorly and posteriorly on the thoracic and anterior pygidial axial rings. Perforations are ubiquitous on the cranidium of A. filoxenia (Fig. 9B) except on the anterior glabellar lobe where it borders the preglabellar area, and on the librigenal border and genal spines.
92We agree with Holloway & De Carvalho (2009) that such perforations are principally different from true fenestrae in that they are not restricted to the interpleural furrows, which are weakly impressed in the species under review here. Although perforations occur intersegmentally in the pygidium of Jaylopygiella jungheimi (Struve, 1992) (Thormann & Weddige, 2001, pl. 10, fig. 77), their distribution is generally characterised as dense and ubiquitous, and so we do not consider any of them to be fenestrae. The smallest fenestrae (e.g. Quadratispina, Bradocryphaeus) are hardly smaller than the largest perforations (e.g. Gudralisium), which disqualifies size as being distinctive.
93When comparing the tubercles of A. boeckae to the perforations of the closely related A. filoxenia, their distributions and sizes appear to match. Whether a tubercle or a perforation occurs in A. boeckae may seem arbitrarily determined in places, yet patterns are more or less symmetric left and right of the sagittal line. The dome-like tubercles of A. boeckae, where these are damaged, show that they are hollow. These observations suggest that the perforations of Asteropyge are modified tubercles. Further evidence in support of this view comes from a specimen of Hollardops sp. A (Fig. 9F) which bears small tubercles that verge to forming a subtransverse row, centrally (exsag.) on the thoracic pleural bands, similarly positioned to the perforations of Asteropyge and Greenops. Exfoliated patches on the same individual (Fig. 9G) reveal large, circular impressions of canal openings on the internal mould of corresponding distribution to the dorsal tubercles. Such sculpture was shown by Størmer (1980, pl. 26, fig. 1) in the pterygometopid Chasmops and classified as smooth tubercles of low convexity that housed sensillae. Other examples in Asteropyginae include a partially exfoliated Neocalmonia pygidium figured by Morzadec (2002, p. 419, fig. 6.12), and even the comparatively large, tubercle-like structures on the internal mould of a pygidium of Gandlops brevispinosus (Gandl, 1972) (Gandl, 1972, pl. 13, fig. 6) which are preservational artifacts of secondary infill subsequent to the decortication of the outer cuticle layer that carried smaller tubercles (Gandl, 1972, pl. 13, fig. 3). Internal moulds of Jaylopygiella debruni show the same tubercle-like artifacts (Fig. 14L, M). Perhaps the perforations are analogous to the comparatively large pits with underlying canals through the cuticle of some homalonotid trilobites (Dalingwater et al., 1999; Rustán et al., 2020). Clearly, more research will be needed to evaluate these structures in asteropygines. Even when hypothesising that the tubercles of Hollardops and the tubercles and perforations of the other genera are all homologous, which we do not advocate (for example, the perforations in the pleural furrows of Gudralisium are unlikely to be modified tubercles because the latter normally do not occur there), it would be precarious to assume that they fulfilled the same function.
10. Systematic palaeontology
94Remarks. The material is housed by the American Museum of Natural History (AMNH), Institut royal des Sciences naturelles de Belgique (IRSNB), Paläontologisches Institut und Museum der Universität Zürich (PIMUZ), Senckenberg Naturmuseum (SMF), and the Natuurhistorisch Museum Maastricht (NHMM). Morphological terms used in the descriptions of the trilobite specimens follow the Revised Trilobite Treatise (Whittington & Kelly, 1997). Thoracic axial rings or pleural projections and pygidial pleural projections are numbered in the figures as T1 to T10 and P1 to P5, respectively.
95The trilobite specimens were coated with ammonium chloride sublimate prior to photography except for Figures 7J–N, 9C–E, 12, 13A, D, H, 14F, 15E (unknown for Figure 13C). We refer to Levi-Setti’s (2014) excellent discussion on the merits of colour photography and the varying details of the mineralised exoskeleton revealed by it, which incited us to deviate in several cases from whitening as the standard. The colour photo used for Figure 12 was provided by the owner of the specimen.
96Order Phacopida Salter, 1864
97Suborder Phacopina Struve, 1959
98Superfamily Acastoidea Delo, 1935
99Family Acastidae Delo, 1935
100Subfamily Asteropyginae Delo, 1935
101Genus Hollardops Morzadec, 1997
102Type species. Asteropyge mesocristata Le Maître, 1952 from the upper Emsian part of the Emsian to Eifelian Chefar el Ahmar Formation, Erg Djemel northeast of El Kseib, Ougarta Basin, Algeria, by original designation.
103Subjective synonyms. Philipsmithiana Lieberman & Kloc, 1997, Modellops Lieberman & Kloc, 1997, Pennarbedops Bignon & Crônier, 2013.
104Diagnosis (provisionally emended, herewith). Asteropygines with weakly anteriorly and ventrally deflected anterior border (sag.); 10 thoracic segments; subisopygous pygidium with short to moderately short lappets (except, probably, H. klugi).
105Species assigned. Hollardops aithassainorum Chatterton, Fortey, Brett, Gibb & McKellar, 2006 (lower Eifelian, Morocco), Hollardops angustifrons sp. nov. (upper Emsian, Morocco), Rhenops babini Morzadec, 1983 (upper Emsian, France), Hollardops boudibensis Morzadec, 1997 (upper Emsian, Morocco), Philipsmithiana burtandmimiae Lieberman & Kloc, 1997 (upper Emsian?, Morocco), Rhenops circumapodemus Smeenk, 1983 (upper Emsian, Spain), Philipsmithiana hyfinkeli Lieberman & Kloc, 1997 (upper Emsian?, Morocco), Hollardops klugi sp. nov. (lower Emsian, Morocco), Hollardops kyriarchos sp. nov. (upper Emsian, Morocco), Hollardops lemaitreae Morzadec, 1997, (upper Emsian, Algeria), Hollardops luscus sp. nov. (upper Emsian, Morocco), Hollardops multatuli sp. nov. (upper Emsian, Morocco), Greenops? struvei Morzadec, 1969 (upper Emsian, France).
106Discussion. As explained above, Hollardops is considered by us to include the junior synonyms Philipsmithiana, Modellops and Pennarbedops. However, a revision of species previously assigned to those genera remains necessary. The choice of A. mesocristata as the type species of Hollardops is unfortunate. The ambiguous identity of this species will be discussed below.
107Bignon & Crônier (2013) diagnosed Hollardops as having frontal lobe rounded; axial furrows curved between S2 and S3; L2 and L3 more elevated than L1; 6 to 11 lenses per dorsoventral file of visual surface; genal spine as long as glabella; anterior tip of palpebral lobe posterior to intersection of axial furrows and S3; thorax with 10 segments; 10 to 16 pygidial axial rings; 5 pleural segments; rounded pleural bands; very wide pleural furrows; 5 pairs of pygidial spines developed from posterior and anterior pleural bands, shorter than pleural width (tr.); terminal medial spine shorter than adjacent pygidial spines, wider than axis. Despite being largely correct in a purely descriptive sense, most of these characters are phylogenetically not very informative, rendering them little useful to discussions of the apomorphies of Hollardops. For instance, the convex axial furrows between S2 and S3, 6 to 11 lenses per dorsoventral file, 10 to 16 pygidial axial rings, 5 pairs of pygidial pleurae, the far anterior junction of axial furrow and S3, and genal spine as long as glabella, are all scattered within the subfamily. This last feature is furthermore not shown by all Hollardops species (e.g. H. burtandmimiae). Pygidial size and lengths of pygidial median and pleural lappets were intimately linked as they furnished support to effective dual coaptation (see 5.2.2.). The width (exsag.) of the pygidial pleural furrows, which helped to control pygidial length, was operated by allometric changes (hyperallometric in the large paratype of H. multatuli) and should be treated with caution. The oldest species, H. klugi, has a comparatively small pygidium, narrow pygidial pleural furrows and probably slightly longer pygidial pleural lappets or spines. The larger pygidia, widened pygidial pleural furrows and comparatively short pleural lappets of stratigraphically younger species are synapomorphies governed in accordance with the condition of 10 thoracic segments. This last feature is a potentially robust apomorphy of Hollardops, under the caveat that this count is unknown for many asteropygine species (see 7.2.). The marginal pit rows enclosed within the sub- and supramarginal tubercle bands (see 4.1.) is a synapomorphy of all Hollardops species except the early Emsian H. klugi where their occurrence cannot be assessed because of extensive damage of the margin. The presence or absence of this feature in members of Asteropyginae is often difficult to corroborate based on published photos, but it is clearly not exclusive to Hollardops and might be synapomorphous of a larger phacopide clade (see 4.1.). Another rarely observed feature in asteropygines is the rostral plate (Basse, 2017) but this is not developed in Hollardops specimens that are known from the ventral morphology.
108Hollardops mesocristatus (Le Maître, 1952)
109* e.p. 1952 Asteropyge mesocristata Le Maître, p. 153, pl. 20, figs 24–26.
110e.p. 1997 Hollardops mesocristata; Morzadec, pl. 6, fig. 1 [only photos of the holotype].
111non 2000 Hollardops mesocristata; Schraut, pl. 13, figs 1–5 [= H. sp.].
112non 2000 Hollardops mesocristata; Schraut, pl. 13, figs 6–8 [= H. struvei].
113non 2001 Hollardops mesocristata; Morzadec, pl. 9, figs 1–4 [= H. aithassainorum].
114non 2004 Hollardops mesocristata; Jansen et al., pl. 2, figs 1, 4 [= H. sp.].
115non 2006 Hollardops mesocristata; Chatterton et al., pp. 28, 29, pl. 14, figs 1, 2, 4–6, pl. 15, figs 1–7, pl. 16, figs 1–6, pl. 20, figs 8, 12, 15 [?= H. luscus].
116non 2012 Hollardops mesocristata; Brett et al., p. 33, fig. 3B [?= H. luscus].
117non 2014 Hollardops mesocristata; Levi-Setti, pls 73, 74 [= H. spp.].
118e.p. 2016 Hollardops mesocristata; Khaldi et al., p. 369, fig. 6m–p [only photos of the holotype].
119non 2017 Hollardops mericristata (sic); Schoenemann et al., p. 6, fig. 4g–i, m [= H. sp.].
120non 2018 Hollardops cf. mesocristata; Crônier et al., p. 247, fig. 3v–z [= H. sp.].
121Discussion. Hollardops mesocristatus is a problematic species and in need of revision. According to Le Maître (1952), the type locality and horizon are the lower Eifelian strata on the northeastern flank of the Erg Djemel syncline. Morzadec (1997, pl. 6, fig. 1) revised the type horizon as the lower (upper Emsian) part of the Chefar el Ahmar Formation and showed the holotype enrolled specimen to be a juvenile. Only three other specimens, an incomplete cephalon (Morzadec, 1997, pl. 7, fig. 3), a small (juvenile?) pygidium (ibid., pl. 8, fig. 8) and the poorly preserved paratype internal mould of an enrolled specimen (ibid., pl. 6, fig. 4), were recorded; these are assumed to have come from the type locality and horizon. Still, the lower part of the Chefar el Ahmar Formation on the northeastern flank of the Erg Djemel syncline encompasses a considerable range of strata. It is not known, for example, whether the three mentioned specimens were collected from the same bank as the holotype or anywhere near it. The fact that the holotype is a small holaspis raises questions as to what allometric changes occurred in later holaspid instars. These uncertainties inhibit fundamental comparisons with other specimens from Erg Djemel and different sections in Algeria and Morocco. The remaining specimens assigned to this species by Morzadec (1997) come from various stratigraphic levels in nearby and distant localities in Algeria. The morphologies of some of these are so disparate that their union in a single species is conjectural. This is exemplified by the dissimilar specimens figured by Morzadec (1997, pl. 7, figs 1, 3, 6), and furthermore by an enrolled specimen (Morzadec, 1997, pl. 6, fig. 6) from Marhouma, some 50 km NNE from Erg Djemel, which bears hallmarks of H. angustifrons including a subtriangular cephalic outline, and the occipital ring as wide as glabella across L1 and narrower (tr.) than glabella across L2.
122A newly collected, enrolled, putative H. mesocristatus from the type locality was recorded by Khaldi et al. (2016, p. 369, fig. 6m, n). The cephalon of that specimen is contrasted with Morzadec’s (1997, pl. 7, fig. 3) topotypical cephalon in having an expressively rounded subtriangular cephalic outline (as against widely rounded with a marked anterior deflection), β slightly more abaxial than γ (as against β far more abaxial than γ because of the laterally expanded anterior glabellar lobe), ε lies opposite centre (sag.) of L1 (as against ε in the same transverse line as median part of S0), and a more prominent subocular ridge anteriorly (weakly developed in Morzadec’s cephalon). Khaldi et al. (2016) only provided a small photo in dorsal view which precludes further comparison at this point, but the differences with Morzadec’s (1997) cephalon demonstrate that at least two distinct Hollardops species occur in Erg Djemel, in stratigraphic proximity of one another, either of which could be conspecific with the type of H. mesocristatus. Hollardops specimens from Morocco have revealed that significant morphological differences can occur in brief stratigraphic intervals, some of which have led to the descriptions of distinct species (e.g. H. aithassainorum by Chatterton et al., 2006; H. kyriarchos and H. multatuli herein). Such observations are pertinent to the identity and morphological boundaries of H. mesocristatus.
123According to Morzadec (1997, 2001), H. mesocristatus is an abundant species in the upper Emsian of Morocco, in the Tafilalt (Hamar Laghdad West and Bou Tchrafine: Amerboh Formation), the Ma’der (Tazoulaït, Jbel Issoumour, Ofaten and Mrakib: Er Remlia and Tazoulaït formations) and the Draa Valley (El Anhsar (sometimes referred to as El Anhsour) SE of Foum Zguid, El Gara, South of Hassi Talah: Timrhanrhart Formation). The lumping of a wide variety of Hollardops morphs from different localities and strata in Algeria and Morocco (e.g. Morzadec, 1997, 2001; Schraut, 2000; Jansen et al., 2004; Chatterton et al., 2006; Brett et al., 2012; Khaldi et al., 2016; Crônier et al., 2018) to include H. hyfinkeli, has resulted in a confounded H. mesocristatus of unclear affinities with the type from Algeria. This has led to considerable problems with the identification of specimens, even if the material is plentiful and well preserved. Schraut (2000) went as far as to place Hollardops struvei in synonymy of Hollardops mesocristatus, because he could not distinguish cephala and pygidia of these species (despite the problems with these species discussed here and above, we prefer to retain both). Chatterton et al. (2006) stated that they were unable to divide their material from the upper Emsian part of the Timrhanrhart Formation in Zguilma into two morphs corresponding with H. mesocristatus and H. burtandmimiae and so they chose to include it in the former species. However, this is at least partly also due to a misapprehended definition of H. burtandmimiae (see below). Chatterton et al. (2006) included in H. mesocristatus, specimens recorded by Morzadec (2001, pl. 9, figs 1–4) from the upper Emsian part of the Timrhanrhart Formation in the nearby Foum Zguid section. Those are indistinguishable from H. aithassainorum described by Chatterton et al. (2006) from the lower Eifelian part of the Timrhanrhart Formation in Zguilma and almost certainly belong to that species. Possibly, Morzadec’s material was mislabelled and actually came from a lower Eifelian, instead of an upper Emsian, level in Foum Zguid.
124The replacement of the unidentifiable name-bearing type of H. mesocristatus with the poorly preserved paratype of evenly imprecise geographic and stratigraphic provenance would be pointless. Therefore, we prefer to restrict H. mesocristatus to the holotype. This is a rigorous measure but one that enables the objective recognition of distinct species, albeit with the knowledge that one of these might potentially belong to H. mesocristatus when future collections at the type locality are conducted. Still, that would require the near-impossible identification of the specific bank that the holotype was extracted from as well as sufficient specimens to span a range of holaspid instars, in order to validate an unambiguous relationship with the type.
125Pillet (1961) assigned Asteropyge mesocristata to Pseudocryphaeus with question and added a new subspecies, Pseudocryphaeus? mesocristata jacqueti, from the Givetian of Mauritania. Based on their resemblances to Asteropyge perforata Morzadec, 1983, Smeenk (1983) reasoned that Pillet’s cephalon of P. mesocristata jacqueti (in fact an incomplete, enrolled specimen) and a pygidium identified by Pillet as Greenops (Neometacanthus) stellifer (Burmeister, 1843) probably belong to the same species, whilst referring both specimens to G. (Neometacanthus) sp. Although the poorly preserved specimens from Mauritania do not allow corroboration of such observations, they are very different from Hollardops and clearly do not belong in that genus (e.g., both show numerous closely spaced perforations of the dorsal exoskeleton and the pygidium has long, weakly tapered spines). Because Smeenk (1983) provided characters to contrast Pillet’s material with other G. (Neometacanthus) species, we are assuming that his suppression of Pseudocryphaeus? mesocristata jacqueti was unintentional.
126Hollardops hyfinkeli (Lieberman & Kloc, 1997)
127(Figs 11C, 15)
128*v 1997 Philipsmithiana hyfinkeli Lieberman & Kloc, pp. 65–68, figs 6.1–6.5, 7.7, 8.9, 21.2, 21.5–21.9, 22.1, 22.2, 22.5, 22.6, 22.8, 22.9, 22.11.
129? 2001 Hollardops mesocristata; Morzadec, pl. 8, fig. 6.
130Material. AMNH 45265, paratype specimen from the Devonian of Morocco. AMNH 45259, holotype from the Devonian of Morocco (refigured here but specimen could not be traced).
131Discussion. Lieberman & Kloc (1997) provided an extensive diagnosis for H. hyfinkeli which includes unique characters that render it distinct from H. mesocristatus and other Hollardops species. Figure 11C shows the cephalic tuberculation of H. hyfinkeli to be restricted to the glabella between S0 and a smooth (sag., exsag.) band running along the front of the anterior glabellar lobe, the posteromedian part of the occipital ring, the palpebral lobe and abaxial part of the palpebral area, and on the genal spine adaxial to the longitudinal furrow.
132The types of H. hyfinkeli were recorded by Lieberman & Kloc (1997) from the “Middle Devonian of Mt. Issomour” (sic). As correctly noted by Morzadec (2001) and according to our present knowledge, no Hollardops species occurs in those strata and it is likely that they came from an upper Emsian level instead. Morzadec (2001) illustrated specimens that he identified as H. mesocristatus, from two different localities and formations; these belong to at least two distinct species, none of which is H. mesocristatus (see synonymies under H. mesocristatus and H. hyfinkeli). The specimen illustrated by Morzadec (2001, pl. 8, fig. 6) has the following characteristics like H. hyfinkeli: maximum of 9 lenses per dorsoventral file, curved S3, angle of the S3 furrows, and the distinctive tubercle pattern on the cephalon. It was recovered from Morzadec’s (2001) level 22 in the upper Emsian Tazoulaït Formation in Jbel Ofaten. Even though this specimen is damaged, it is the only published indicator where H. hyfinkeli may occur. Morzadec (2001) illustrated the ranges of H. mesocristatus in the Er Remlia to Tazoulaït formations in the Ma’der, from sections Mrakib, Ofaten and Issoumour. Because we have not observed H. mesocristatus in the Ma’der, we are uncertain what species this range may represent except for the one published specimen that looks like H. hyfinkeli. One of us (GK) has a specimen of H. hyfinkeli in his possession that is associated with a specimen of Morocconites malladoides Struve, 1989 on the same rock slab. Numerous photos of specimens with characters of H. hyfinkeli are available on the Internet. Some of these are associated with the phacopids Morocops ovatus (McKellar & Chatterton, 2009) and Boeckops stelcki McKellar & Chatterton, 2009. McKellar & Chatterton (2009) identified a Morocconites horizon in the Tazoulaït Formation in Jbel Ofaten, just south of Jbel Issoumour (Fig. 1), and indicated that this level also contained M. ovatus and B. stelcki. From these observations, a likely occurrence of H. hyfinkeli would be the Morocconites horizon in the Tazoulaït Formation at Jbel Ofaten. However, we have seen several specimens in a different limestone matrix of unknown origin; these are like H. hyfinkeli but they also show variations in the curvature of the S3 furrows and the length of the genal spines. It is unknown if this is a variation of H. hyfinkeli or a different species. Bed-by-bed sampling in the Tazoulaït Formation at Jbel Ofaten would be helpful in determining the occurrence of H. hyfinkeli and its morphological variation. McKellar & Chatterton (2009) and Chatterton & Gibb (2010) illustrated a Hollardops horizon above the Psychopyge horizon in the Tazoulaït Formation at Jbel Issoumour but there is no photo of a Hollardops from this level, so it is unknown what species might occur there. One of our H. boudibensis specimens was recovered from a level just below the Psychopyge horizon in the Tazoulaït Formation at Jbel Issoumour.
133In May of 2021, we requested the loan of the type specimens of H. hyfinkeli from the AMNH. We were advised by Dr Bushra Hussaini that neither she nor Dr Melanie Hopkins had been successful at tracing the types, except for paratype AMNH 45265. Three specimens, holotype AMNH 45259 and paratypes AMNH 45260 and 45264, are missing and were originally entered in the catalogue as unavailable (GK, pers. comm. with B. Hussaini in 2021). Photos of the holotype taken by one of us (GK) prior to the publication of this species are reproduced here as Figure 15A–C, E. New photos of paratype AMNH 45265 are also provided (Fig. 15D, F–H).
134Morzadec (2001) placed H. hyfinkeli in synonymy of H. mesocristatus but he did not provide an explanation for this choice. Even when considering the unrealistic hypothesis that Morzadec’s (1997) specimens from Erg Djemel all belong to H. mesocristatus, there are a range of features that render H. hyfinkeli distinct: S3 convex anteriorly, whereas it runs straight in “H. mesocristatus”; anterior glabellar lobe bears well-developed frontal auxiliary impressions (FAIs) according to a diamond-shaped pattern, whereas only pmi and pli are well discernible in “H. mesocristatus”; glabella has an angular instead of a smoothly rounded anterior outline; longer (sag., exsag.) preglabellar field; anterior margin of glabella and preglabellar field run subparallel to anterior border whereas the cephalic outline of “H. mesocristatus” markedly protrudes medially; broad, angular as against parabolic cephalic outline; prominent eye ridge visible both anteriorly and lateral in dorsal view; visual surface is adjacent to, or overhangs the lateral border furrow in dorsal view (this is never the case in “H. mesocristatus”). No photo of a large holaspid pygidium of “H. mesocristatus” from Erg Djemel is known to compare H. hyfinkeli with except for the coiled specimen figured by Khaldi et al. (2016, p. 369, fig. 6m, n). The pygidium of that specimen has sustained extensive damage but it is principally different from H. hyfinkeli in having the anterior two pairs of interpleural furrows run in a continuous line onto the border towards the base of the associated pleural lappet. The anterior two pairs of interpleural furrows of H. hyfinkeli are sigmoidally flexed near the lateral border before adjoining the rather broad bases of the pleural lappets, the pleural lappets are wider, and the median and pleural lappets are shorter relative to the border width.
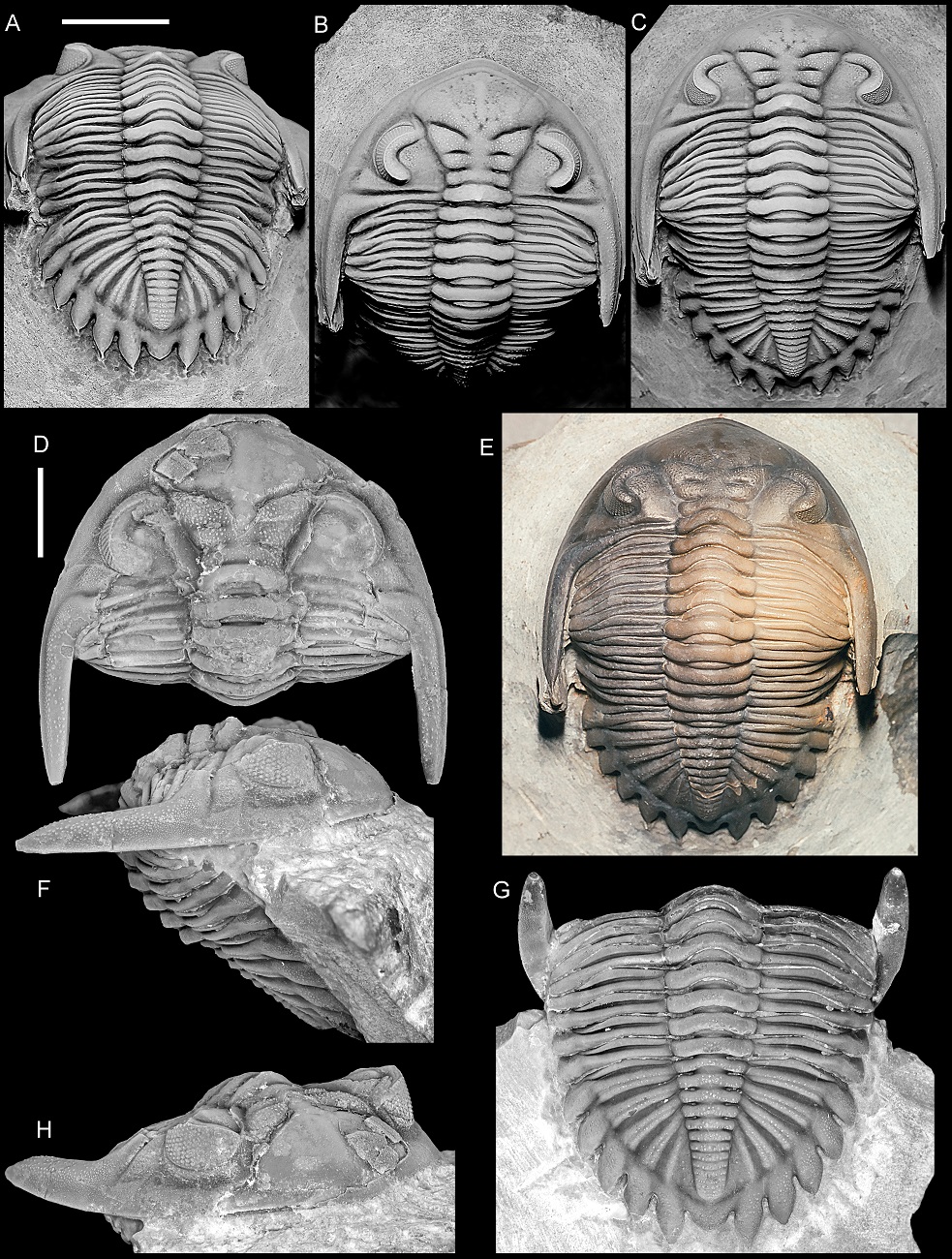
Figure 15. Hollardops hyfinkeli (Lieberman & Kloc, 1997), Devonian of Morocco. A–C, E. Holotype, AMNH 45259, in dorsal views of pygidium (A) and cephalon (B), and dorsal view (C, E). Paratype, AMNH 45265, in dorsal view of cephalon (D), lateral view (F), dorsal view of pygidium (G), and anterolateral view (H).
Scale bars are 5 mm.
135Hollardops burtandmimiae (Lieberman & Kloc, 1997)
136(Figs 8L–Q, 11D, 16, 17H–L)
137*v 1997 Philipsmithiana burtandmimiae Lieberman & Kloc, pp. 68, 69, figs 7.2–7.6, 7.8, 21.1, 21.3, 21.4, 22.3, 22.4, 22.7, 22.10, 22.12, 22.13.
138non 2001 Hollardops burtandmimiae; Morzadec, pl. 8, fig. 8, pl. 9, figs 5–8 [= H. spp.].
139non 2010 Hollardops burtandmimiae; Chatterton & Gibb, figs 6.4, 6.9 [= H. sp.].
140non 2014 Hollardops burtandmimiae; Lawrance & Stammers, unnumb. fig.
1412017 Hollardops burtandmimiorum (sic); van Viersen et al., p. 17 [undue subsequent spelling].
142Material. AMNH 45266, holotype specimen and AMNH 45267, 45271, 45272, paratype specimens from the Devonian of Morocco. NHMM 2021041, ventrally prepared specimen from the Devonian of Morocco.
143Discussion. Lieberman & Kloc (1997) provided an extensive description of H. burtandmimiae and contrasted it with H. hyfinkeli. Figure 11D shows the cephalic tuberculation of H. burtandmimiae to be restricted to the glabella between S0 and S3, and laterally on the genal spine where it drops in height towards the longitudinal furrow.
144Morzadec (2001) revised H. burtandmimiae in assigning to it a range of morphologically disparate specimens from various localities in the Ma’der and Tindouf basins. Although Morzadec (2001) did not emend the diagnosis to accommodate the other forms, he provided characters to discriminate H. burtandmimiae and H. mesocristatus, which he considered to be closely similar species: the former has the front of the cephalon less rounded, 27/8 (rarely 11) lenses as against 8 to 9 in H. mesocristatus, a longer pygidial axis comprising a larger number (15–16) of rings as against 14 in H. mesocristatus, longer and more pointed pygidial spines, and a shorter pygidial median spine. For reasons already provided above, this comparison can no longer be retained.
145New photos of some of the specimens described by Lieberman & Kloc (1997), including the holotype, are provided here. Characteristics are the high-parabolic cephalic outline with long genal spines, high towering eyes, large tubercles restricted to the glabella between S0–S3 and laterally on the genal spine where it drops in height towards the longitudinal furrow, very short but firmly impressed pseudo-S4 furrow incising the anterior glabellar lobe just anterior to γ, markedly ventrally deflected and anteriorly protruding cephalic border, and comparatively long and slender pygidial pleural lappets.
146Hollardops burtandmimiae was described by Lieberman & Kloc (1997) based on the types from the “Middle Devonian of Morocco, near Mt. Issomour” (sic). As explained above, no Hollardops is known to occur in those levels. Chatterton & Gibb (2010, pp. 1191, 1200, figs 6.4, 6.9) identified a damaged incomplete specimen and a pygidium from the Erbenochile couche (basal, upper Emsian part of the upper Emsian to lowermost Eifelian El Otfal Formation) in Jbel Issoumour (‘CG2010’ locality in Fig. 1), as H. burtandmimiae. Their incomplete specimen is dissimilar from H. burtandmimiae in having a different angle and curvature of the S3 furrows, more and larger tubercles on the palpebral area and L1–L2, and very different lengths and shapes of the pygidial pleural lappets. Chatterton & Gibb (2010) only provided low-resolution photos in dorsal view. They did not describe their specimens and used Morzadec’s (2001) obsolete differential diagnosis to discriminate H. burtandmimiae and H. mesocristatus. Chatterton & Gibb (2010) suggested that H. burtandmimiae may range from the upper Emsian into the lower Eifelian but the reasons for their statement are unclear. Morzadec (2001) believed it to be restricted to the upper Emsian. We have seen photos on the Internet of specimens claimed to have been collected from the El Otfal Formation in Jbel Ofaten, that appear to belong to H. burtandmimiae. Some of these were associated with Coltraneia oufatenensis Morzadec, 2001 and an undescribed Koneprusia species on the same rock slabs. Morzadec (2001) reported C. oufatenensis only in his level 25 in the lower (upper Emsian) part of the El Otfal Formation in Jbel Ofaten. Van Viersen et al. (2017) reported the co-occurrence of C. cf. oufatenensis with H. burtandmimiae in the upper Emsian part of the El Otfal Formation just West of Jbel Issoumour, but this information was based exclusively on observations communicated by local miners.
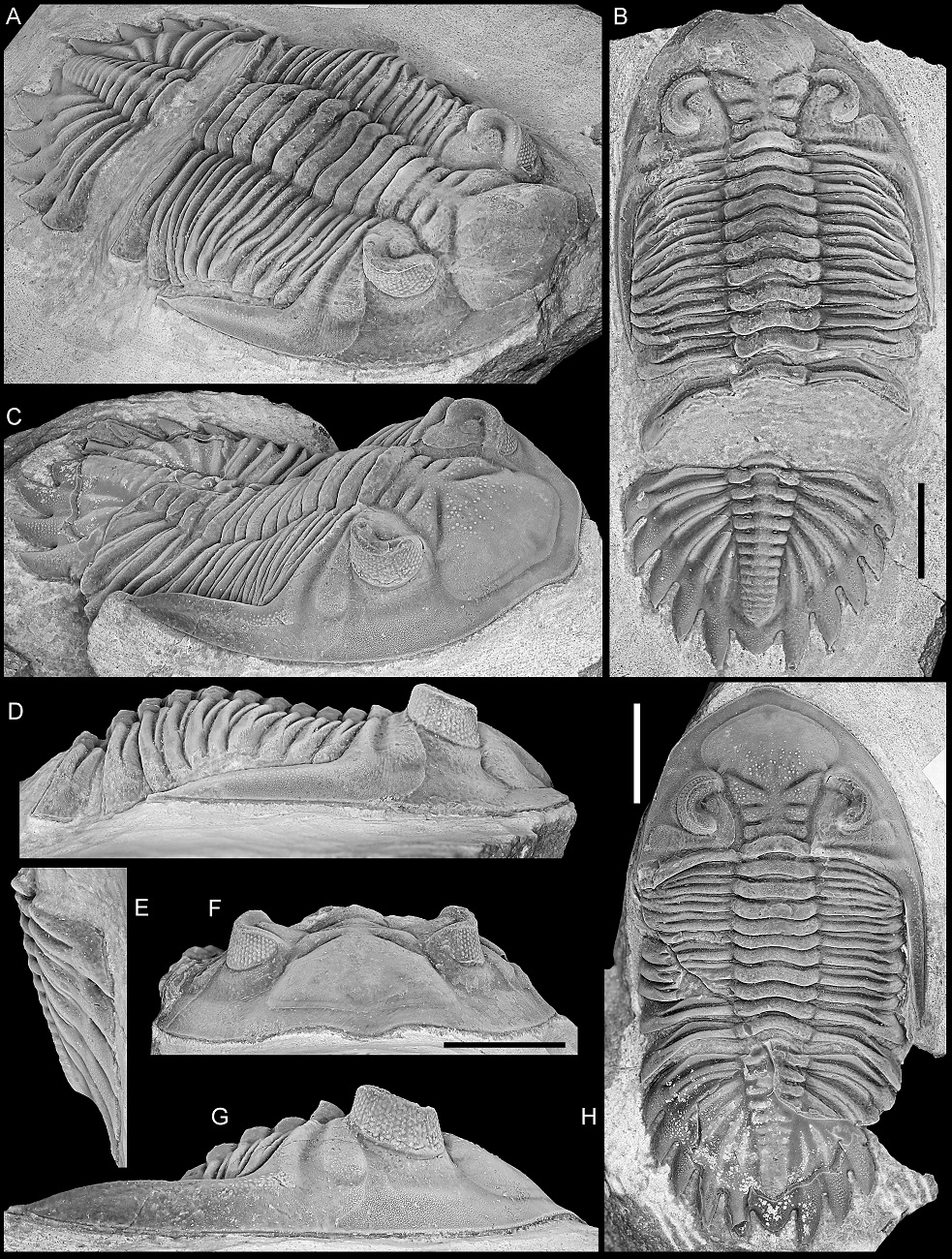
Figure 16. Hollardops burtandmimiae (Lieberman & Kloc, 1997), Devonian of Morocco. A, B, D, E. Holotype, AMNH 45266, in oblique anterolateral (A), dorsal (B) and lateral (D, E) views. C, G, H. Paratype, AMNH 45267, in oblique anterolateral (C), lateral (G) and dorsal (H) views. F. AMNH 45272, in anterior view.
Scale bars are 10 mm.
147Hollardops boudibensis Morzadec, 2001
148(Figs 9H–Q, 10F, 11G, 17A–G)
149* 2001 Hollardops boudibensis Morzadec, pp. 70, 71, pl. 8, figs 1, 2, 4, 5, 7, 10.
150Material. IRSNB a13571, specimen with broken left genal spine tip, from the Tazoulaït Formation in the Jbel Issoumour1 locality. NHMM 2021042, coiled specimen from the Devonian of Morocco.
151Discussion. Morzadec (2001) characterised H. boudibensis as having a flat border, anterior half of the anterior glabellar lobe smooth, genal spines with ducted tips, short and rounded pygidial lappets, and median lappet as long as pleural lappets. Morzadec (2001) did not compare his new species to the exceedingly similar H. lemaitreae Morzadec, 1997 from the upper Emsian in Algeria. The Algerian material generally has more pronounced sculpture although one cephalon (Morzadec, 1997, pl. 4, fig. 4) has an anteriorly smooth anterior glabellar lobe, and the pygidial pleural spines of some (e.g. ibid., pl. 5, fig. 5) but not all (e.g. ibid., pl. 5, fig. 2) specimens are narrower than the Moroccan material. Morzadec (1997) did not provide a description of H. lemaitreae and his specimens are not as well-preserved as the available material of H. boudibensis. A more detailed comparison of these species must await re-examination of the Algerian material. According to Morzadec (2001) the types of H. boudibensis come from a level near the summit of the Er Remlia Formation. We have assigned our specimen from the lower part of the overlying Tazoulaït Formation to H. boudibensis and not to H. lemaitreae, based on its similarities to the types of Morzadec (2001) and the geographic proximity to the type locality, near Bou Dib. Better preserved specimens of both species from their type localities will be needed to carry out a comprehensive comparison.
152Hollardops boudibensis shares similarities with H. burtandmimiae such as the uniformly sized tubercles centrally on the glabella but it is principally different from the latter in having S2 and S3 straight, fading well before reaching the axial furrow; long (sag., exsag.), weakly differentiated preglabellar field; palpebral lobe posteriorly merged with palpebral field because of fading palpebral furrow here; narrow (tr.) librigenal field; groove-like posterior border furrow of cephalon; lack of a ventrally deflected anterior border; more widely rounded cephalic outline; shorter pygidial lappets.

Figure 17. A–G. Hollardops boudibensis Morzadec, 2001, level just below Psychopyge couche, Tazoulaït Formation, Jbel Issoumour, Morocco. IRSNB a13571, in dorsal view of cephalon (A), lateral view (B), dorsal view of pygidium (C), oblique anterolateral (D), anterior (E), oblique posterolateral (F) and posterior (G) views.
H–L. Hollardops burtandmimiae (Lieberman & Kloc, 1997), Devonian of Morocco. H. Ventrally prepared specimen, NHMM 2021041. I, K. AMNH 45271, in dorsal (I) and oblique anterior (K) views. J, L. AMNH 45272, in dorsal (J) and lateral (L) views.
M, N. Hollardops multatuli sp. nov., Cyphaspis couche, Hollardops Limestone Member of the Khebchia Formation, Tadachacht, Morocco. Paratype, NHMM 2022001, close-up of third thorax segment (M) and anterior view of cephalon (N).
Scale bars are 5 mm.
153Hollardops kyriarchos sp. nov.
154(Figs 8E, F, 10D, 18A–J)
155Etymology. Derived from the Greek κυρίαρχος (ascendant), for being the stratigraphically earliest Hollardops species in the Khebchia Formation; appositionally.
156Holotype. IRSNB a13569, complete specimen (Figs 8E, F, 10D, 18A–J).
157Type locality and horizon. Bou Tserfine locality; basal part of Hollardops Limestone Member, Khebchia Formation.
158Diagnosis. Central part of glabella, palpebral area and palpebral lobe in the same subhorizontal plane; anterior glabellar lobe downward sloped, running straight towards anterior margin (sag.); rudimentary preglabellar field and furrow; broad (tr.), slender (sag., exsg.) occipital ring, slightly wider than glabella across L2, without median node; wide thoracic axis comprised of elongate (tr.), weakly W-shaped rings; comparatively weakly demarcated pleural fields by distally shallow pleural and interpleural furrows.
159Description. Cephalon is of rounded subtriangular outline. Glabella anterior to S0 is moderately weakly vaulted (tr.); centrally slightly higher than palpebral area; slightly wider (tr.) than long (sag.). Axial furrows more divergent near L2 than near L1; straight near L1 and L2; weakly curved abaxially near L3. Occipital ring as wide (tr.) as glabella across L2; ascending slightly above the rest of the glabella; lacking a median node on its posterior margin. L1 is comparatively weakly vaulted; of subquadratical outline. L2 and L3 are of rounded trapezoidal outline. S0 is narrow (sag., exsag.); slightly shallower medially than exsagitally. S1 deep; adaxially slightly forward flexed. S2 moderately deep. S3 weakly curved; divergent at around 135°; junction between S3 and axial furrow lies lateral to γ. Smoothly diamond-shaped anterior glabellar lobe. Anterior border furrow very shallow in front of the glabella, indicated by a change of inclination. Eye comprising 29 dorsoventral files with maximally nine lenses per file (29/9). Librigenal field is narrow and very steeply inclined (tr.) lateral of eye. Lateral border furrow is shallow; mostly recognisable as a change of inclination (tr.) from the librigenal field towards the lateral border. Concave, medially slightly anteroventrally deflected anterior to lateral cephalic border; of equal width (sag., exsag.) in front of glabella. Posterior border runs subtransversally until posterior to ω; from there slightly broadening and curving posteriorly abaxially. Genal spines not fully known. Dorsal sculpture consists of densely spaced, moderately small tubercles that become smaller towards the cephalic borders.
160Thorax comprises 10 segments. Dorsal sculpture consists of close-set tubercles, similar to cephalon and pygidium.
161Pygidium is widely rounded triangular; length about 0.70 of width. Comparatively broad axis comprised of 15 axial rings, the first five of which are weakly W-shaped; the anterior eight rings are straightened and laterally well-demarcated by deep furrows. Axial furrows slightly more convergent near anterior five rings than posterior to these. Five pairs of gently posteriorly curved pleurae; each pair slightly shorter than the previous. Moderately broad pleural furrows, about as wide as associated anterior pleural bands. Posterior pleural bands are slightly narrower than anterior bands. Broad pleural spines with slightly pointed tips. Terminal lappet is broad and dorsally about as high as adjacent pleural spines. Dorsal sculpture is similar to thorax, especially on anterior part of the pygidium; consisting of densely spaced, small tubercles except medially (exsag.) on the lateral parts of the anterior eight axial rings; a median node is never developed.
162Discussion. The beds that yielded our specimens of Hollardops kyriarchos and Hollardops multatuli in the Hollardops Limestone Member in the Assa area are separated by only several metres. Both species share a number of overall similarities with in particular the well-developed subocular ridge overhanging the librigenal field and the rudimentary abaxial ends of S2 and S3. Hollardops multatuli is principally different in having a more angular cephalic outline; narrower glabella; S3 furrows less divergent (115° instead of 135°); junction between S3 and axial furrow opposite γ; narrower (tr.) occipital ring, narrower (tr.) than glabella across L2, longer (sag.) medially than abaxially (exsag.), with a weakly defined median node; more cephalic relief with a strongly vaulted (tr.), well-demarcated glabella; larger eyes (31/11) closer to axial furrow anteriorly; sparser, larger tubercles all over the pygidium. Length/width ratios of the cephalic outline are the same for specimens of both species but the widths across the occipital ring relative to cephalic widths are 0.45 and 0.50, and maximum widths across the glabella relative to cephalic widths are 0.25 and 0.29 for H. multatuli and H. kyriarchos, respectively; i.e., the occipital ring and widest point of glabella are comparatively wide in the latter species.
163The abaxially weakly impressed S2, occipital ring as wide (tr.) as glabella across L2, and the smooth anterior expansion (tr.) of the glabella and very weakly indicated preglabellar field, are shared with H. lemaitreae and H. boudibensis but the cephala of those species tend to be more trapezoidally outlined and the pygidia are very different.
164Hollardops hyfinkeli also has weakly impressed S2 as well as many overall similarities to H. kyriarchos yet it differs principally in having a posteriorly dorsally convex occipital ring carrying a median tubercle; palpebral lobe higher than palpebral area and glabella; larger distance between eye and lateral border; sparser tubercles on the pygidium.

Figure 18. A–J. Hollardops kyriarchos sp. nov., base of Hollardops Limestone Member, Khebchia Formation, Bou Tserfine, Morocco. Holotype, IRSNB a13569, anterior cephalic border and posterior pygidial border (digitally modified version of E as to appear in near-full coaptation) (A), dorsal view (digitally modified version of E as to appear fully enrolled) (B), dorsal view of pygidium (C), oblique anterolateral view (D), dorsal view of cephalon (unmodified; note the slight left rotation of the cephalon) (E), lateral (F), oblique anterior (G), oblique lateral (H), posterior (I), and anterior (J) views.
K. Hollardops multatuli sp. nov., Cyphaspis couche, Hollardops Limestone Member of the Khebchia Formation, Tadachacht, Morocco. Paratype, NHMM 2022001, in oblique lateral view.
Scale bar is 10 mm.
165Hollardops multatuli sp. nov.
166(Figs 3P, 5, 6, 7B, C, E, F, J-N, 10A–C, 11E, 17M, N, 18K, 19)
167Etymology. Named after Eduard Douwes Dekker, a 19th-century Dutch writer, known under the pseudonym Multatuli (“I have suffered much”, derived from the Latin multa tulit), in reference to the pathologies shown by the large paratype of this species.
168Holotype. IRSNB a13567, complete specimen (Figs 3P, 7B, E, 10B, 19A–G, J, M).
169Paratypes. IRSNB a13568, deformed complete specimen and NHMM 2022001, nearly complete specimen, from type locality and horizon.
170Type locality and horizon. Tadachacht locality; Cyphaspis couche, upper part of Hollardops Limestone Member, Khebchia Formation.
171Diagnosis. S3 furrows divergent at around 115°, curved posteriorly abaxially where it is indicated by absence of sculpture; junction between S3 and axial furrow lies opposite γ; palpebral area weakly inflated (exsag.) adaxially; posterior pygidial border furrow accentuated by deep pit-like depressions. Numerous tubercles on entire glabella, thoracic axis, and an obliquely running strip including the palpebral lobe, anterior part of palpebral area, librigena lateral to L1–L2, anterior part of fixigena adaxial to border furrow.
172Discussion. This species is similar to H. kyriarchos and the two are discriminated as explained under that species. The holotype is used for comparative purposes because paratype NHMM 2022001 is very large and distorted, and paratype IRSNB a13568 is longitudinally truncated (digitally enhanced in Fig. 19I to demonstrate what it looked like in undeformed condition).
173Hollardops hyfinkeli is principally different in having smaller eyes with maximally 10 lenses per dorsoventral file; larger distance between eye and lateral border; more anteriorly located junction between S3 and axial furrow; wider cephalic outline (length/width ≈ 0.50 in H. hyfinkeli and around 0.57 in H. multatuli); well-developed, kite-shaped FAI system.
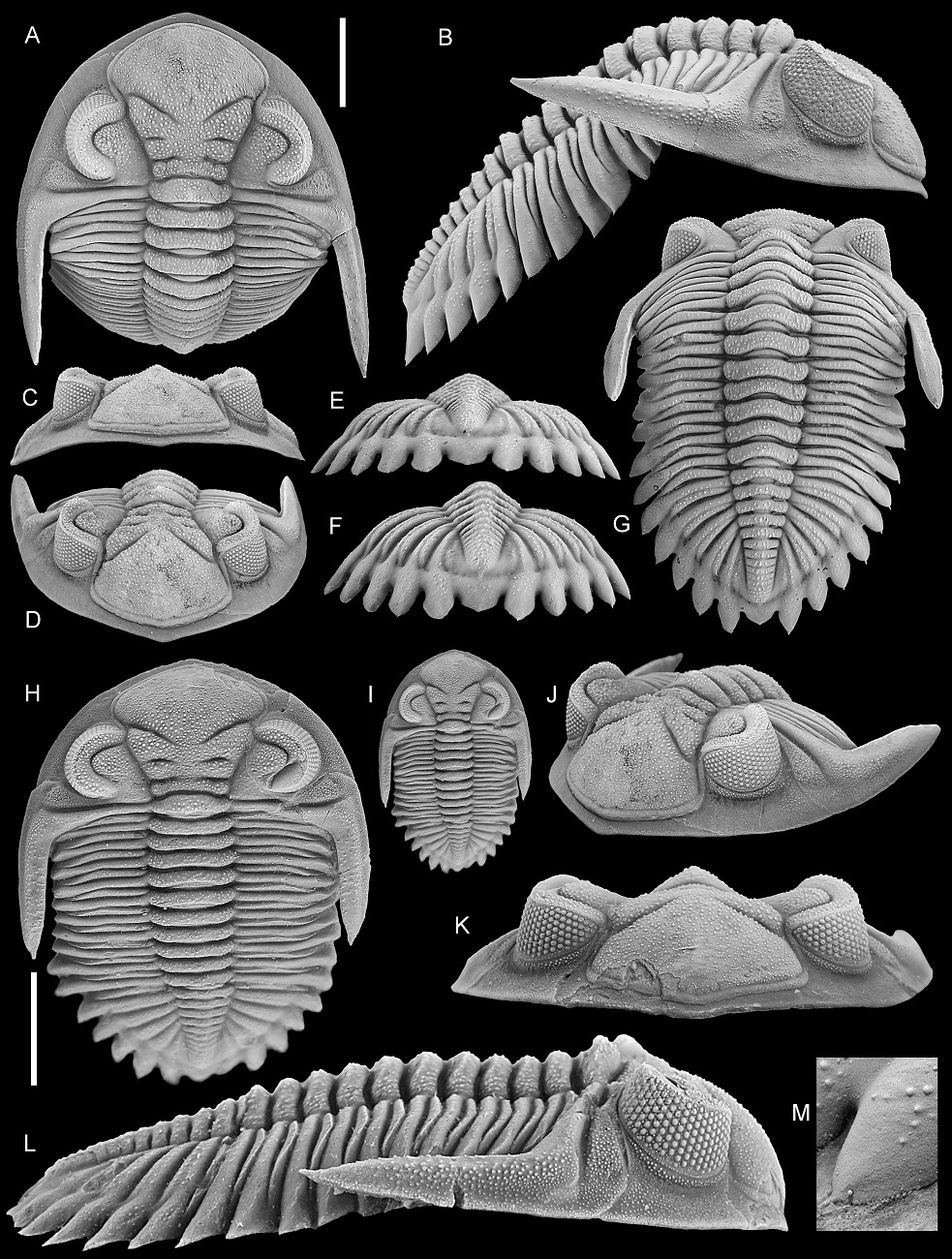
Figure 19. Hollardops multatuli sp. nov., Cyphaspis couche, Hollardops Limestone Member of the Khebchia Formation, Tadachacht, Morocco. A–G, J, M. Holotype, IRSNB a13567, in dorsal (A), lateral (B), anterior (C), oblique anterior (D), posterior (E), oblique posterior (F) views, dorsal view of pygidium (G), oblique anterolateral view (J), and close-up of P2 second right pygidial lappet (M). H–L. Paratype slightly deformed (sag.) specimen, IRSNB a13568, in dorsal view of cephalon (H), same as H but elongated (digitally modified as to appear undeformed) (I), anterior (K), and lateral (L) views.
Scale bars are 5 mm.
174Hollardops angustifrons sp. nov.
175(Figs 7H, 10E, 11A, 20)
176Etymology. Derived from the Latin angustus (narrow) and frons (forehead), in reference to the narrow glabella.
177Holotype. IRSNB a13570, complete specimen (Figs 7H, 10E, 20).
178Type locality and horizon. Jbel Khraid locality; Timrhanrhart Formation (ZGEE2 equivalent).
179Diagnosis. Glabella of narrow subpentagonal outline; anterior glabellar lobe strongly vaulted (sag.), higher than central area of glabella; lateral border furrow reaches glabella anterior to β; distance between eye and lateral border furrow equals distance between border furrow and margin; occipital ring as wide (tr.) as glabella across L1. Numerous coarse tubercles all over the thorax, pygidium and cranidium, with the exception of the genal spine and its base.
180Description. Cephalon is of rounded subtriangular outline. Glabella anterior to S0 is moderately weakly vaulted (tr.); centrally about as high as palpebral area; as long (sag.) as wide (tr.). Axial furrows more divergent near L2 than near L1; straight near L1 and L2; weakly curved abaxially near L3. Occipital ring is about as wide (sag.) as glabella across L1; ascending high above the rest of the glabella; bearing a median node on its posterior margin. L1 is comparatively weakly vaulted; of subquadratical outline. L2 and L3 are of rounded trapezoidal outline. S0 is narrow (sag., exsag.); well-developed throughout. S1 and S2 transversely running; S1 shallow medially; S2 short. S3 straight; divergent at around 120°; junction between S3 and axial furrow lies anterior of the visual surface. Smoothly diamond-shaped anterior glabellar lobe. Preglabellar furrow shallow, especially medially. Moderately narrow (exsag.), weakly inflated preglabellar field which is broadest medially; running parallel to facial suture and anterior border in front of the glabella. Anterior border furrow shallow in front of the glabella. Librigenal field is broad and very steeply inclined (tr.) lateral of eye. Eye is moderately small for a Hollardops species; comprising 28 dorsoventral files with maximally seven lenses per file (28/7). Lateral border furrow is shallow; mostly recognisable as a change of inclination (tr.) from the librigenal field towards the lateral border. Concave, medially slightly anteroventrally deflected anterior to lateral cephalic border; of equal width (sag., exsag.) in front of glabella; broadest (tr.) lateral to β. Posterior border runs subtransversally until posterior to ω; from there slightly broadening and curving posteriorly abaxially. Genal spines are moderately long (more or less equalling sagittal length of cephalon). Dorsal sculpture consists of densely spaced, moderately small tubercles.
181Thorax comprises 10 segments. Thoracic axis comparatively narrow (tr.) for genus; widths of the axial rings little differentiated. Dorsal sculpture is similar to that on occipital ring and posterior cephalic border except that a median node is never developed.
182Pygidium is widely rounded triangular; length about 0.75 of width. Axis comprised of 13 axial rings, the first five of which are weakly W-shaped; the remaining rings are straightened (tr.) and demarcated by shallow, transverse furrows. Axial furrows slightly more convergent near anterior five rings than posterior to these. Five pairs of weakly curved pleurae; each pair slightly shorter than the previous. Broad pleural furrows, slightly wider than associated anterior pleural bands. Posterior pleural bands are as wide as anterior bands. Subtriangular pleural spines with pointed tips. Terminal lappet is comparatively short, subtriangular, and dorsally lower than adjacent pleural spines. Dorsal sculpture is similar to thorax, especially on anterior part of the pygidium; consisting of densely spaced, moderately small tubercles; a median node is never developed.
183Discussion. Hollardops burtandmimiae is principally different in having eyes towering high above the glabella; sparse tubercles restricted to the posterior two-thirds of glabella; markedly ventrally deflected anterior border; longer preglabellar field (sag., exsag.); larger (tr.) librigenal field; genal spines longer than glabella; wider thoracic to pygidial axis; sculpture on pygidium largest on median and pleural lappets.
184Hollardops hyfinkeli has occipital ring wider than L1 (tr.); moderately closely spaced tubercles on posterior two-thirds of glabella and sparse tubercles on anterior third of glabella; longer preglabellar field (sag., exsag.); well-developed FAI system; larger eyes closer to lateral border, comprising maximally 10 lenses per dorsoventral file; wider thoracic to pygidial axis; sparser, larger tubercles on the pygidial axis and pleurae.
185Hollardops multatuli has S3 curved abaxially; junction between S3 and axial furrow lies opposite γ; wider glabella, especially across occipital ring, L1 and L2; occipital ring with (weakly defined) lateral lobes; well-developed subocular ridge overhanging the librigenal field; smooth transition from librigenal field towards lateral border; broader, more inflated preglabellar field; larger eyes; less ventrally deflected anterior border; generally larger sized and less densely spaced tubercles dorsally on the exoskeleton.
186Hollardops kyriarchos has an anteriorly wider, more weakly differentiated glabella; widely rounded subtriangular cephalic outline; larger eyes; occipital ring as wide as glabella across L2; wider thoracic to pygidial axis; broader pygidial lappets.
187For a comparison with Hollardops luscus, see below.
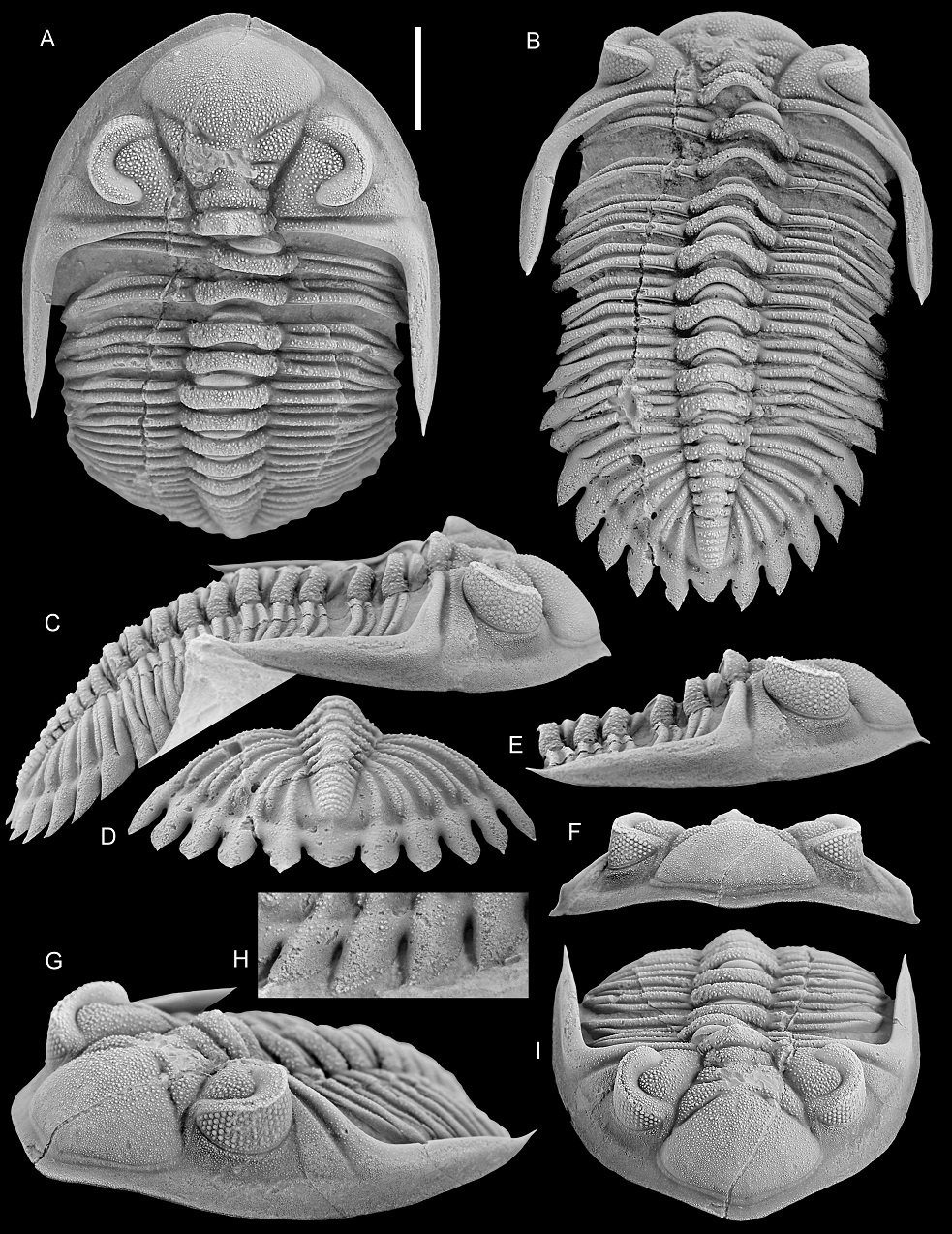
Figure 20. Hollardops angustifrons sp. nov., Timrhanrhart Formation (ZGEE2 equivalent), Jbel Khraid, Morocco. Holotype, IRSNB a13570, in dorsal view of cephalon (A), dorsal view of pygidium (B), oblique lateral (C), oblique posterior (D), lateral (E), anterior (F), oblique anterolateral (G), and oblique anterior (I) views, and close-up of right pygidial lappets 2 to 5 (H).
Scale bar is 5 mm.
188Hollardops luscus sp. nov.
189(Figs 11J, 21A–G)
190? e.p. 2006 Hollardops mesocristata; Chatterton et al., pp. 28, 29, pl. 14, figs 1, 2, 4–6, pl. 15, figs 1–7, pl. 16, figs 1–6, pl. 20, figs 8, 12, 15 [non pl. 14, fig. 3].
191v 2019 Hollardops; van Viersen & Lerouge, p. 230.
192Etymology. Derived from the Latin luscus (one-eyed), alluding to this condition of the type.
193Holotype. NHMM 2021043, complete specimen (Fig. 21A–G).
194Type locality and horizon. Timerzit locality; Walliserops couche, lower part of El Otfal Formation.
195Diagnosis. Cephalic sculpture excluding genal spines comprised of densely spaced granules, alternated on central part of cranidium by small tubercles; anterior glabellar lobe comparatively long (sag., exsag.); long distance (exsag.) between anteriormost point of visual surface and anterior border; smoothly curved preocular suture, especially between β and γ.
196Discussion. Several specimens of this species were recovered from the type locality (AV, pers. comm. with I. Kesselaer and F. Lerouge in 2022). Even though only the damaged type is currently fully prepared and available for deposition, the material is so morphologically distinctive that we could not place it in any of the Hollardops species formally recognised at present. Perhaps the most similar is H. angustifrons from which the new species differs in having finer, more densely packed dorsal sculpture; comparatively weakly anteriorly and ventrally deflected anterior cephalic border; less vaulted (sag.) anterior glabellar lobe; smoothly curved (as against sigmoidal) preocular suture, especially between β and γ; lateral border furrow reaches glabella posterior to β (instead of anterior to β in H. angustifrons) (this is well visible in anterior view); visual surface with 28/8 lenses (instead of 28/7); librigenal field narrower (tr.) than lateral border here; 15 pygidial axial rings (instead of 13). Like most asteropygines, the fifth pygidial pleural pair is connected to the fifth axial ring. The additional axial rings of H. luscus require the posterior pleural pairs to cross a longer distance toward the border compared to H. angustifrons.
197Chatterton et al. (2006) recorded specimens from an upper Emsian level in their Zguilma locality and assigned these to H. mesocristatus. We have not examined the original specimens and we are unsure whether they all belong to a single species; some of them may turn out to be H. luscus.

Figure 21. A–G. Hollardops luscus sp. nov., Walliserops couche, El Otfal Formation, Timerzit, Morocco. Holotype, NHMM 2021043, in dorsal (A), lateral (B), anterior (C) and oblique anterior (D) views of cephalon, dorsal (E) and lateral (F) views of pygidium, and close-up (G) of third right pygidial pleural lappet.
H–N. Hollardops klugi sp. nov., Seheb El Rhassel Group (Faunule 1 of Klug et al., 2008), Ouidane Chebbi, Morocco. Holotype, PIMUZ 7296, in lateral (H), anterior (I) views, and dorsal views of pygidium (J) and cephalon (K). Paratype, PIMUZ 7297, in dorsal view of pygidium (L), lateral view (M) and dorsal view of cephalon (N).
LO is occipital ring.
O–R. Minicryphaeus suavius sp. nov., Seheb El Rhassel Group (Faunule 1 of Klug et al., 2008), Ouidane Chebbi, Morocco. Holotype, PIMUZ 7291, in anterior (O), lateral (P) and dorsal (Q, R) views.
Scale bars are 5 mm.
198Hollardops klugi sp. nov.
199(Figs 11I, 21H–N)
2002008 Pilletina zguidensis; Klug et al., p. 142, pl. 11, figs 4–7, 13–16.
201Etymology. Named after Christian Klug, in recognition of his important contributions to palaeontology in Morocco.
202Holotype. PIMUZ 7296, incomplete enrolled specimen (Fig. 21H–K).
203Paratype. PIMUZ 7297, incomplete, enrolled, partially exfoliated specimen, from type locality and horizon.
204Type locality and horizon. Ouidane Chebbi locality; Faunule 1, kitabicus Conodont Zone, Seheb El Rhassel Group.
205Diagnosis. Facial sutures parallel to slightly convergent from ε to β; anterior border weakly dorsally arched anterior to glabella; small eyes with 26?/9 lenses; thoracic and anterior seven pygidial axial rings carry a short, spiny median node; small pygidium relative to cephalon along with very narrow pleural furrows; each consecutive pygidial pleural pair shorter posteriorly.
206Discussion. The phylogenetic affinities of H. klugi are discussed above. This species is not particularly similar to congeners and appears to lack the distinctive marginal row of pore canals enclosed by supra- and submarginal tubercle rows. The uncertainties about the distribution of this feature within Asteropyginae cast some doubt on its phylogenetic importance. It is primarily the 10-segmented condition of the thorax that led us to assign this species to Hollardops. We might have chosen to erect a monotypic genus for H. klugi but considered this to be little helpful since accepting its position as ancestral to Hollardops would probably render it a paraphyletic fragment of the latter. More importantly, the potentially paedomorphic origin of H. klugi and the lack of data on the ontogenies of other Early Devonian asteropygines impede crucial comparisons at this time.
207Hollardops boudibensis has crowded tubercles medially throughout the axis that are potentially homologous with the median nodes of H. klugi.
208Genus Platykardiapyge nov.
209Etymology. Derived from the Greek πλατύς (wide), καρδιά (heart) and -pyge (a common suffix for trilobites), in reference to the widely heart-shaped pygidium.
210Type species. Metacanthina maderensis Morzadec, 2001 from the upper part of the Pragian Ihandar Formation, Jbel Issoumour.
211Diagnosis. Preglabellar field twice as long sagittally as exsagittally anterior to γ; concave epiborder furrow (type species only) running border parallel from point opposite β until junction with postocular suture, from there weakly impressed before merging with sigmoidally flexed lateral border furrow, continuing onto the genal spine, dividing the latter into an adaxial, only distally tapered part, and shorter (exsag.), more rapidly tapered abaxial part; posterior five thoracic axial rings carry a thick, short median spine; pygidial pleural fields horizontal adaxially, steep distally, their outline well demarcated by subvertical edge and discontinuation of the pleural and interpleural furrows on the pygidial border; six or seven pairs of nearly straight pygidial pleurae; anterior and posterior bands in each pygidial pleural pair are mostly similarly sized and shaped (posterior band is only proximally better developed than anterior band); each pleural lappet about as thick (exsag.) and as long as associated pleura; median pygidial lappet projecting slightly further posteriorly than fifth pleural lappet pair, with very large base and heart-shaped lateral to posterior outline.
212Species assigned. Metacanthina aequisulcata aequisulcata Gandl, 1972 (Pragian, Spain). Tentatively assigned are Metacanthina anatolica Haas, 1968 (Pragian, Türkiye), Cryphaeus asiaticus Verneuil, 1866 (Pragian, Türkiye), Metacanthina lips mozoensis Gandl, 1972 (lower Emsian, Spain), Asteropyge wallacei Termier & Termier, 1950 (Pragian, Morocco). These species are tentatively included in the new genus because their morphologies and especially their cephala and thoraxes are insufficiently known.
213Discussion. The type species, Platykardiapyge maderensis comb. nov. (Fig. 8C, D), was originally assigned to Metacanthina by Morzadec (2001, pp. 66, 67) based on its cephalic morphology, shape of the pygidial spines and its pygidial segmentation. Bignon & Crônier (2013) regarded Metacanthina as monotypic and in doing so, orphaned many previously included species. We agree with Bignon & Crônier that Metacanthina maderensis and Metacanthina issoumourensis Morzadec, 2001 do not belong in that genus. We are in doubt about what genus to place M. issoumourensis in. It lacks most of the diagnostic characters of Platykardiapyge. Basse & Müller (2012) provided reasons to include M. issoumourensis in Pilletina. Morzadec (2001, p. 67, pl. 6, figs 1–4) discussed and refigured the types of Metacanthina wallacei from the Pragian of the Draa Valley. A specimen from the lower Emsian of the Tafilalt Platform, figured by Klug et al. (2008, pl. 11, figs 8–12) and identified as M. wallacei, has a median node on the occipital ring, smaller eyes, and fewer pygidial axial rings and pleurae (five pairs), among other distinctive features; this particular specimen is referred to a new species of Minicryphaeus below.
214The sixth and seventh pygidial pleural pairs of Platykardiapyge are also shown by stratigraphically older acastids from the Silurian and the lowermost Devonian. This suggests that the condition in Platykardiapyge may be a symplesiomorphy. However, since the relatively high pleural pair count is a rare occurrence in Asteropyginae and therefore helpful in distinguishing Platykardiapyge from most other genera, we have included this character in our diagnosis. A well-developed sixth pair of pygidial pleurae is furthermore known e.g. in an undescribed species of Quadratispina from the Givetian or Frasnian of Belgium (see Basse, 2003, pl. 27, fig. 460) but, given the large stratigraphic interval to span the two occurrences, likely to be homoplastic. Platykardiapyge shows a furrowed lateral border of cephalon which is also known from Siluro-Devonian dalmanitids (e.g. Whiteley et al., 2002, pl. 91) and so this character, too, might be symplesiomorphic.
215Bignon & Crônier (2013) assigned both Metacanthina aequisulcata aequisulcata and Metacanthina aequisulcata matutina Gandl, 1972 to Pilletina with doubt. In the same paper Bignon & Crônier treated Metacanthina matutina as a distinct species which they included in their new genus Morzadecops (= Feruminops). The latter assignment is provisionally accepted pending the discovery of a cephalon; M. matutina is conditionally transferred to Feruminops accordingly.
216Genus Minicryphaeus Bignon & Crônier, 2013
217Type species. Pseudocryphaeus minimus Morzadec, 2001 from the Pragian Ihandar Formation, Ofaten, Morocco, by original designation.
218Species assigned. Minicryphaeus giganteus Bignon, Corbacho & López-Soriano, 2014 (Pragian, Morocco), Pseudocryphaeus quaterspinosus Morzadec, 2001 (Pragian, Morocco), Pseudocryphaeus sarirus Morzadec, 2001 (Pragian, Morocco), Minicryphaeus suavius sp. nov. (lower Emsian, Morocco).
219Minicryphaeus suavius sp. nov.
220(Fig. 21O–R)
221e.p. 2008 Metacanthina wallacei; Klug et al., pp. 141, 142, pl. 11, figs 8–11 [only PIMUZ 7291].
222Etymology. Derived from the Latin suavium (kiss), in reference to the pouting frontal spine of this species.
223Holotype. PIMUZ 7291, incomplete enrolled specimen (Fig. 21O–R).
224Type locality and horizon. Ouidane Chebbi locality; Faunule 1, kitabicus Conodont Zone, Seheb El Rhassel Group.
225Diagnosis. Cephalon of inverted heart-shaped outline; S1–S3 lateral glabellar furrows relatively short (adax.) for a Minicryphaeus species; large palpebral area; visual surface containing 26?/7 lenses; pygidium with anterior band of a sixth pleural pair.
226Discussion. Klug et al. (2008) reported 43 specimens from the type locality and provided a brief description. Minicryphaeus minimus has a less abaxially expanded anterior glabellar lobe, anteriorly convex occipital ring, S1 to S3 reaching farther adaxially, large palpebral lobe at the cost of palpebral area, visual surface containing 21/5 lenses, narrower lateral cephalic border, weak change of inclination from pygidial pleural field to border and lappets.
227Minicryphaeus sarirus has abaxially short (exsag.) L1, visual surface containing 24/8 lenses, large palpebral lobe, pygidium with 10 axial rings (as against eight axial rings in M. minimus and M. suavius).
228Minicryphaeus quaterspinosus has abaxially short (exsag.) L1, visual surface containing 22/6 lenses, large palpebral lobe, pygidium with seven axial rings, and four pleural and lappet pairs.
229Minicryphaeus giganteus has a large anterior cephalic projection, abaxially short (exsag.) L1, large palpebral lobe, higher librigenal field below the eye (lateral view), and large visual surface with 34/8 lenses. In the holotype of M. giganteus the pygidial spines appear to originate from the posterior pleural bands (Bignon et al., 2014, fig. 3.3) whereas this is not clear in the paratypes.
230Genus Bignonops nov.
231Etymology. Named after Arnaud Bignon, in recognition of his work on asteropygine trilobites.
232Type species. Kayserops tamnrhertus Chatterton, Fortey, Brett, Gibb & McKellar, 2006 from the upper Emsian part of the upper Emsian to Eifelian Timnrhanrhart Formation, Zguilma.
233Diagnosis. Small asteropygine with rounded rhombic, distinctly inflated anterior glabellar lobe; visual surface with relatively few, large lenses adjoining or overhanging lateral border; axial furrow convex near L3; transversely running posterior cephalic border furrow; wide (tr.) palpebral area; pygidium with indistinct border and border furrow; weakly vaulted (tr.) axis barely ascending above pleural fields.
234Species assigned. Greenops sp. of Morzadec (1997) (Emsian, Algeria), here referred to as Bignonops sp. nov.
235Discussion. Whilst describing Kayserops tamnrhertus Chatterton et al. (2006) expressed some doubts about the correct generic assignment of their new species although they preferred not to erect a monophyletic genus for it. Bignon & Crônier (2013) proposed Gandlops (type species: Kayserops brevispinosus Gandl, 1972) and added Pseudocryphaeus ogivalis Morzadec, 1976, Delocare? palenciae Smeenk, 1983, and Kayserops tamnrhertus. Van Viersen & Lerouge (2021) accepted Gandlops as a taxon of independent generic rank but underlined the need for a critical review in order to ascertain its monophyly. Van Viersen & Lerouge (2021) sustained the transfer of Delocare? palenciae to Dunopyge that was proposed by van Viersen (2013) and regarded Morzadec’s (1997, pl. 6, fig. 2) enrolled Greenops sp. specimen from the upper Emsian of El Kseib, Algeria, as a Gandlops species exhibiting exceeding similarities to K. tamnrhertus. In fact, the latter two species are so different from G. brevispinosus and G. ogivalis that we decided to remove them from that genus and propose Bignonops gen. nov.
236Morzadec (1997) only briefly commented on his Greenops sp. from Algeria which, despite remaining in open nomenclature here (we have not examined the single, incompletely preserved specimen), we believe represents an undescribed Bignonops species that enables a first assessment of potential synapomorphies shared with Bignonops tamnrhertus. Chatterton et al. (2006) noted the median axial nodes on most of the thorax segments of B. tamnrhertus and suggested that it descended from a more spinose ancestor. In this and other respects (e.g. swollen preglabellar field, far abaxially positioned eyes, prominent subocular ridge, significantly raised ventral pygidial rim) B. tamnrhertus is certainly a morphologically distinctive species. The Algerian B. sp. nov. lacks median nodes altogether except perhaps for a weak node posteriorly on the occipital ring, and also other marked features of B. tamnrhertus do not seem as well developed, if at all. Median nodes are retained into adulthood in some species of Greenops where they are considered to be a paedomorphic feature and this may also be the case for B. tamnrhertus. Thus, until more species are discovered to demonstrate otherwise, the distinguishing characters of B. tamnrhertus are best treated as autapomorphies. The incorrect count of 10 thorax segments reported for B. tamnrhertus by Chatterton et al. (2006) was dealt with above (see 7.2.). Like most asteropygines, this species has 11 thorax segments.
237Minicryphaeus suavius shares with B. tamnrhertus a small pygidium with distally steeply inclined pleurae which, during enrolment, rested against the inner margin of the lateral cephalic doublure, and small pleural spines of rounded section that project slightly more horizontally beyond the cephalic margin. But Bignonops tamnrhertus has a raised posterior pygidial rim and a V-shaped vincular furrow which tightly sealed the internal parts from the surroundings. This resembles a phacopid-type of enrolment and is clearly a convergence on members of that family. Although the posterior part of the pygidium of the B. sp. nov. specimen is damaged, the steep edges of the pleural region overlap the anterior cephalic border in frontal view (Morzadec, 1997, pl. 6, fig. 2b) in a way similar to the damaged B. tamnrhertus specimen figured by Chatterton et al. (2006, pl. 19, fig. 7).
238Species of Minicryphaeus also share a number of cephalic features with Bignonops, including a short (exsag.) anterolateral border, short but robust, proximally divergent and distally posteriorly directed genal spine with a short furrow, and steep eyes. If Minicryphaeus and Bignonops are truly phylogenetically closely related, the most significant character changes to reach the Bignonops states would be the increased convexity (tr.) of the axial furrow near L3, broader (tr.) palpebral field and lateral migration of the hypertrophied eye adjoining the more inclined lateral border, and the disappearance of the pygidial border furrow. The touting frontal process of M. suavius suggests the presence of a breathing device which would imply the absence of a V-shaped vincular furrow like that of B. tamnrhertus.
11. Conclusions
239Hollardops, including its junior subjective synonyms Philipsmithiana, Modellops and Pennarbedops, is much more diverse than previously assumed. Its species occur in the lower Emsian to lower Eifelian of southern Morocco and in the upper Emsian of western Algeria, Brittany and the Cantabrian Mountains. The known palaeogeographic distribution is thus exclusively northern to peri-Gondwanan (Fig. 22). The palaeogeographic dispersions of Platykardiapyge during the Pragian, Feruminops during the lower Emsian, and Hollardops during the upper Emsian are consistent with the proximity of peri-Gondwanan terranes to northern Gondwana, connected by open, shallow-water migration routes. The absence of these genera in Laurussia is taken to indicate a strong facies preference rather than questioning the proximity of both palaeocontinents during the Lower Devonian because other asteropygines (Comura, Delocare, Ganetops, Psychopyge, Treveropyge, etc.) and members of many other subfamilies roamed successfully in the Rheic Ocean. Cyphaspis, Delocare, Diademaproetus, Gerastos, Kettneraspis, Leonaspis and Morocops, among other trilobite genera, typically occur in the upper Emsian of southern Laurussia and northern to peri-Gondwana, having benefited from eustatic sea level rises in that time. These observations might be taken to suggest that the closure of the Rheic Ocean had already progressed far in the Lower Devonian, in accordance with the palaeogeographic maps of Scotese & McKerrow (1990) and Scotese (2001).
240
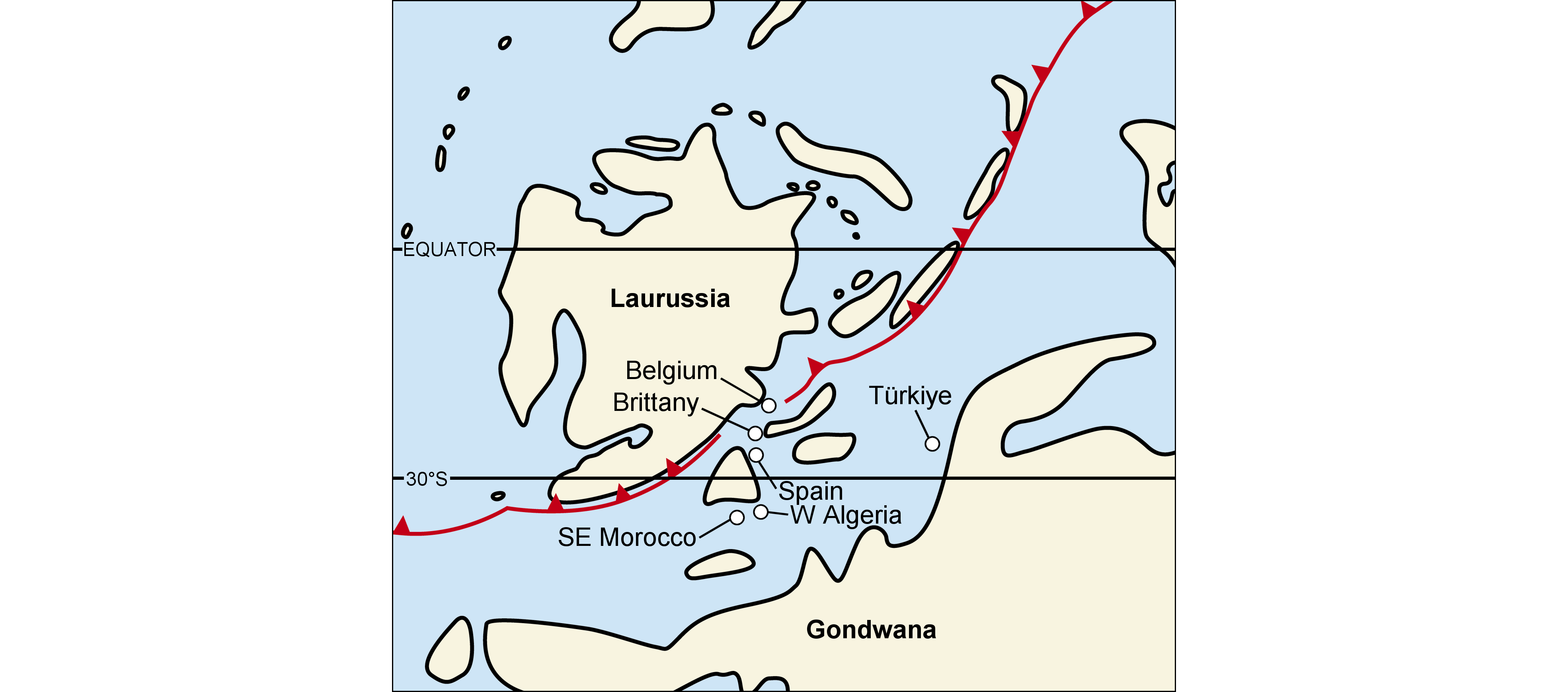
Figure 22. Palaeogeographic map of Laurussia and northern and peri-Gondwana during the Lower Devonian. Adopted from Scotese (2001) and modified.
241
242Hollardops is construed as a scavenger or predator with well-developed schizochroal eyes, that used its shovel-like cephalon to plough the top layers of the sediment in search of food. It is inferred, from large and exceptionally well-preserved specimens from Morocco, that Hollardops had a well-developed sensory system that allowed it to monitor its surroundings. A pseudomegalaspid type of enrolment, with the distinctive, ventrally deflected anterior border abutting the pygidial posterior rim, was practised by all of its species. Together with the robust genal spine bases, these features inhibited lateral shearing. Like many other asteropygines, Hollardops was able to retract its pygidium during coaptation to let water circulate internally and antennae protrude, and this may well have been the ‘normal resting’ mode. Many other members of the class, and particularly members of Phacopida, are also assumed to have practised dual coaptation. Other asteropygines, including Bignonops and Minicryphaeus, had different morphological adaptations that may have served as breathing devices.
243The 10-segmented thoracic condition of Hollardops is fairly unique among asteropygines and accepted as an apomorphy. The genus may have arisen from Feruminops through paedomorphic processes but without knowledge of the ontogenies of their species, this remains difficult to assess. A revised diagnosis of Hollardops is provided herein, provisionally, because H. klugi is in several aspects not representative of the stratigraphically younger, more derived species, and because its pleural projections are unknown. Additionally, we are aware of the occurrences of several Hollardops species in Morocco that remain to be documented fully.
244Certain morphological features of Hollardops need to be subjected to further study. These include the frontal auxiliary impressions which occur in all species and, like tubercles on the cephalon, could show distinctive distribution patterns of potential taxonomic value. Allometric changes are a barely touched upon topic and it is difficult to assess their importance to taxonomic descriptions. Future studies, based on more specimens collected under stratigraphic control, should lead to a better understanding of these aspects of the morphology of Hollardops, species definitions, ontogeny and biostratigraphy.
Acknowledgments
245We thank Bushra Hussaini and Melanie Hopkins of the American Natural History Museum, Annelise Folie and Alain Drèze of the Institut royal des Sciences naturelles de Belgique, and Rudy Lerosey-Aubril (then Senckenberg Naturmuseum) for tracing type material in their care, and Lisa Amati of the New York State Museum, Christian Klug of the Universität Zürich, and Andries Weug (then Senckenberg Naturmuseum) for providing photographs of trilobite specimens. Ivo Kesselaer (Palaeontologica Belgica), and the late Dieter Holland (Ilsede) and Harald Prescher (Kerpen) donated trilobite specimens for study and shared field information. Felix Collantes (Palencia) and Peter Taghon (Gent) allowed us to figure trilobite specimens in their collections. Stephan Helling of the Westfälische Wilhelms-Universität Münster, Martin Basse of the Senckenberg Naturmuseum, Said Ihmadi (Alnif), David Rueda (Madrid), David Holloway of the Museums Victoria, Barry van Bakel of the Oertijdmuseum De Groene Poort, and Frederik Lerouge of Palaeontologica Belgica, participated in helpful discussions. Scott Morrison of the University of Oregon helped us to find some of the literature used. Mirco Alberti (Hessisches Landesamt für Naturschutz, Umwelt und Geologie), Martin Basse, and Bruce Lieberman (University of Kansas) are thanked for their constructive review comments which helped to improve the manuscript.
Author contribution
246Both authors contributed equally to the data acquisition, their interpretation and the writing of this article.
Data availability
247Most of the studied specimens are housed in official repositories guaranteeing their long-term safekeeping and availability to other researchers for future studies. Several other specimens residing in private collections are included for illustrative purposes; none of these were designated types or used in the descriptions.
References
248Adrain, J.M., 1997. Proetid trilobites from the Silurian (Wenlock-Ludlow) of the Cape Phillips Formation, Canadian Arctic Archipelago. Palaeontographia Italica, 84, 21–111.
249Alberti, M., 2014. Im Leben und im Tod vereint: Trilobit mit Koralle. Fossilien, 2014/3, 23–25.
250Arbizu, M., 1977. Asteropyginae (Trilobita) du Dévonien des Montagnes cantabriques (Espagne). Bulletin de la Société géologique et minéralogique de Bretagne C, 9, 59–102.
251Asselberghs, E., 1930. Description des faunes marines du Gedinnien de l’Ardenne. Mémoires du Musée royal d’Histoire naturelle de Belgique, 41, 1–73.
252Babcock, L.E., 1993. Trilobite malformations and the fossil record of behavioral asymmetry. Journal of Paleontology, 67, 217–229. https://doi.org/10.1017/S0022336000032145
253Babcock, L.E., 2003. Trilobites in Paleozoic predator-prey systems, and their role in reorganization of Early Paleozoic ecosystems. In Kelley, P.H., Kowalewski, M. & Hansen, T.A. (eds), Predator-Prey Interactions in the Fossil Record. Kluwer Academic/Plenum Publishers, New York, 55–92. https://doi.org/10.1007/978-1-4615-0161-9_4
254Barrande, J., 1852. Systême Silurien du centre de la Bohême. 1ère Partie: Recherches paléontologiques. Vol. 1: Crustacés: Trilobites. J. Barrande, Prague and Paris, 935 p. https://doi.org/10.5962/bhl.title.14776
255Barrande, J., 1872. Systême silurien du centre de la Bohême, 1ère Partie: Recherches paléontologiques. Supplément au Vol. 1. Trilobites, crustacés divers et poissons. J. Barrande, Prague and Paris, 647 p.
256Basse, M., 2002. Eifel-Trilobiten. 1. Proetida. Goldschneck Verlag, Korb, 152 p.
257Basse, M., 2003. Eifel-Trilobiten. 2. Phacopida 1. Goldschneck Verlag, Korb, 198 p.
258Basse, M., 2017. Die Ventralseite des Trilobiten Rhenops – eine Terra incognita. Fossilien, 2017/2, 32–35.
259Basse, M. & Müller, P., 2004. Eifel-Trilobiten III. Corynexochida, Proetida (2), Harpetida, Phacopida (2), Lichida. Quelle & Meyer-Verlag, Wiebelsheim, 260 p.
260Basse, M. & Müller, P., 2012. Drei Arten der Asteropyginae aus den Seifen-Schichten, Westerwald (Trilobita; Mittel-Siegen-Unterstufe, Unter-Devon; Rheinisches Schiefergebirge). Geologica et Palaeontologica, 44, 9–26.
261Basse, M. & Müller, P., 2016. Trilobiten aus dem Ober-Emsium und frühen Eifelium der südlichen Lahnmulde (Rupbach-Schiefer, Leun-Schiefer und Ballersbach-Kalk). Abhandlungen der Senckenberg Gesellschaft für Naturforschung, 572, 1–329.
262Bauer, R.T., 1981. Grooming behavior and morphology in the Decapod Crustacea. Journal of Crustacean Biology, 1, 153–173. https://doi.org/10.2307/1548154
263Bauer, R.T., 2013. Adaptive modification of appendages for grooming (cleaning, antifouling) and reproduction in the Crustacea. In Watling, L. & Thiel, M. (eds), The natural History of the Crustacea, Volume 1: Functional Morphology and Diversity. Oxford University Press, New York, 337–375. https://doi.org/10.1093/acprof:osobl/9780195398038.003.0013
264Becker, K. & Wahl, M., 1996. Behaviour patterns as natural antifouling mechanisms of tropical marine crabs. Journal of Experimental Marine Biology and Ecology, 203, 245–258. https://doi.org/10.1016/0022-0981(96)02575-0
265Becker, R.T., Jansen, U., Plodowski, G., Schindler, E., Aboussalam, S.Z. & Weddige, K., 2004a. Devonian litho- and biostratigraphy of the Dra Valley – an overview. Documents de l’Institut Scientifique, Rabat, 19, 3–18.
266Becker, R.T., Aboussalam, S.Z., Bockwinkel, J., Ebbighausen, V., El Hassani, A. & Nübel, H., 2004b. Upper Emsian stratigraphy at Rich Tamelougou near Torkoz (SW Dra Valley, Morocco). Documents de l’Institut Scientifique, Rabat, 19, 85–89.
267Becker, R.T., Bockwinkel, J., Ebbighausen, V., Aboussalam, S.Z., El Hassani, A. & Nübel, H., 2004c. Lower and Middle Devonian stratigraphy and faunas at Bou Tserfine near Assa (Dra Valley, SW Morocco). Documents de l’Institut Scientifique, Rabat, 19, 90–100.
268Bergström, J., 1973. Organization, life, and systematics of trilobites. Fossils and Strata, 2, 1–69.
269Bergström, J. & Brassel, G., 1984. Legs in the trilobite Rhenops from the Lower Devonian Hunsrück Slate. Lethaia, 17, 67–72. https://doi.org/10.1111/j.1502-3931.1984.tb00668.x
270Bignon, A. & Crônier, C., 2013. The systematics and phylogeny of the Devonian subfamily Asteropyginae (Trilobita: Phacopida). Journal of Systematic Palaeontology, 12, 637–668. https://doi.org/10.1080/14772019.2013.804008
271Bignon, A., Corbacho, J. & López-Soriano, F.J., 2014. A revision of the first Asteropyginae (Trilobita; Devonian). Geobios, 47, 281–289. https://doi.org/10.1016/j.geobios.2014.09.002
272Birkenmajer, K. & Bruton, D.L., 1971. Some trilobite resting and crawling traces. Lethaia, 4, 303–319. https://doi.org/10.1111/j.1502-3931.1971.tb01926.x
273Bonino, E. & Kier, C., 2009. Trilobiti. Il libro del Museo. Back To The Past Museum, Puerto Morelos, 442 p.
274Brandt, D.S., 1996. Epizoans on Flexicalymene (Trilobita) and implications for trilobite paleoecology. Journal of Paleontology, 70, 442–449. https://doi.org/10.1017/S0022336000038373
275Brett, C.E. & Walker, S.E., 2002. Predators and predation in Paleozoic marine environments. Paleontological Society Papers, 8, 93–118. https://doi.org/10.1017/S1089332600001078
276Brett, C.E., Allison, P.A. & Hendy, J.W., 2010. Comparative taphonomy and sedimentology of small-scale mixed carbonate/siliciclastic cycles: Synopsis of Phanerozoic examples. In Allison, P.A. & Bottjer, D.J. (eds), Taphonomy: Process and Bias Through Time. Springer, Dordrecht, Topics in Geobiology, 32, 107–198. https://doi.org/10.1007/978-90-481-8643-3_4
277Brett, C.E., Zambito, J.J., Schindler, E. & Becker, R.T., 2012. Diagenetically-enhanced trilobite obrution deposits in concretionary limestones: The paradox of “rhythmic events beds”. Palaeogeography, Palaeoclimatology, Palaeoecology, 367, 30–43. https://doi.org/10.1016/j.palaeo.2011.12.004
278Bruton, D.L. & Haas, W., 1997. Functional morphology of Phacopinae (Trilobita) and the mechanics of enrolment. Palaeontographica (A), 245, 1–43. https://doi.org/10.1127/pala/245/1997/1
279Bultynck, P. & Hollard, H., 1980. Distribution comparée de conodontes et goniatites dévoniens des plaines du Dra, du Ma’der et du Tafilalt (Maroc). Aardkundige Mededelingen, 1, 9–73.
280Bultynck, P. & Walliser, O.H., 2000. Devonian boundaries in the Moroccan Anti-Atlas. In Bultynck, P. (ed.), Subcommision on Devonian Stratigraphy. Recognition of Devonian series and stage boundaries in geological areas. Courier Forschungsinstitut Senckenberg, 225, 211–226.
281Campbell, K.S.W., 1977. Trilobites of the Haragan, Bois d’Arc and Frisco formations (Early Devonian) Arbuckle Mountains Region, Oklahoma. Oklahoma Geological Survey Bulletin, 123, 1–227.
282Chatterton, B.D.E., 1971. Taxonomy and ontogeny of Siluro-Devonian trilobites from near Yass, New South Wales. Palaeontographica (A), 137, 1–108.
283Chatterton, B.D.E. & Campbell, M., 1993. Enrolling in trilobites: a review and some new characters. Memoirs of the Association of Australasian Palaeontologists, 15, 103–123.
284Chatterton, B.D.E. & Gibb, S., 2010. Latest Early to early Middle Devonian trilobites from the Erbenochile bed, Jbel Issoumour, southeastern Morocco. Journal of Paleontology, 84, 1188–1205. https://doi.org/10.1666/10-027.1
285Chatterton, B.D.E., Fortey, R.A., Brett, K.D., Gibb, S. & McKellar, R.C., 2006. Trilobites from the upper Lower to Middle Devonian Timrhanrhart Formation, Jbel Gara el Zguilma, southern Morocco. Palaeontographica Canadiana, 25, 1–177.
286Chen, G., Han, N. & Luo, R., 2011. A restudy of Coronocephalus gaoluoensis Wu, 1979. Acta Palaeontologica Sinica, 50, 284–303. [in Chinese, with English summary]. https://doi.org/10.19800/j.cnki.aps.2011.03.002
287Chlupáč, I., 1994. Devonian trilobites – Evolution and events. Geobios, 27, 487–505. https://doi.org/10.1016/S0016-6995(09)90029-8
288Chlupáč, I., Feist, R. & Morzadec, P., 2000. Trilobites and standard Devonian stage boundaries. Courier Forschungsinstitut Senckenberg, 220, 87–98.
289Clarkson, E.N.K. & Henry, J.-L., 1973. Structures coaptatives et enroulement chez quelques Trilobites ordoviciens et siluriens. Lethaia, 6, 105–132. https://doi.org/10.1111/j.1502-3931.1973.tb01186.x
290Clarkson, E.N.K. & Whittington, H.B., 1997. Enrollment and coaptative structures. In Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, 67–74.
291Corbacho, J. & Kier, C., 2013. Divesidad y distribución de Acanthopyge (Belenopyge), (Lichidae) en Marruecos; Devónico inferior y medio. Batalleria, 18, 3–14.
292Crônier, C., Oudot, M., Klug, C. & De Baets, K., 2018. Trilobites from the Red Fauna (latest Emsian, Devonian) of Hamar Laghdad, Morocco and their biodiversity. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 290, 241–276. https://doi.org/10.1127/njgpa/2018/0781
293Dalingwater, J.E., Siveter, D.J. & Mutvei, H., 1999. Cuticular microstructure of some Silurian homalonotid trilobites from Sweden. Journal of Paleontology, 73, 256–262. https://doi.org/10.1017/S002233600002775X
294Delo, D.M., 1940. Phacopid trilobites of North America. Geological Society of America Special Papers, 29, 1–132. https://doi.org/10.1130/SPE29-p1
295Dojen, C., Aboussalam, S. & Becker, R.T., 2011. Lowermost Emsian to lower Givetian ostracods from the Western Dra Valley (Morocco): First palaeoecological and event-stratigraphical data. Joannea Geologie und Paläontologie, 11, 53–54.
296Dowding, E.M. & Ebach, M.C., 2019. Evaluating Devonian bioregionalization: quantifying biogeographic areas. Paleobiology, 45, 636–651. https://doi.org/10.1017/pab.2019.30
297Edgecombe, G.D., 1991. Morocconites Struve, 1989, a Devonian acastine trilobite (Calmoniidae: Acastinae). American Museum Novitates, 2998, 1–7.
298Edgecombe, G.D. & Wright, A.J., 2004. Silicified Early Devonian trilobites from Brogans Creek, New South Wales. Proceedings of the Linnean Society of New South Wales, 125, 177–188.
299Eldredge, N., 1971. Patterns of cephalic musculature in the Phacopina (Trilobita) and their phylogenetic significance. Journal of Paleontology, 45, 52–67.
300Eldredge, N., 1973. Systematics of Lower and lower Middle Devonian species of the trilobite Phacops Emmrich in North America. Bulletin of the American Museum of Natural History, 151, 285–338.
301Eldredge, N., 1977. Trilobites and evolutionary patterns. In Hallam, A. (ed.), Patterns of Evolution as Illustrated by the Fossil Record. Elsevier, Amsterdam, Developments in Palaeontology and Stratigraphy, 5, 305–332. https://doi.org/10.1016/S0920-5446(08)70329-7
302Fatka, O. & Budil, P., 2021. Frontal auxiliary impressions in the Ordovician trilobite Dalmanitina Reed, 1905 from the Barrandian area, Czech Republic. Bulletin of Geosciences, 96, 481–491. https://doi.org/10.3140/bull.geosci.1792
303Feist, R., Talent, J.A. & Mawson, R., 2001. Devonian trilobites from the Shogrām Formation, Kurāgh, Chitral (NW Pakistan). Alcheringa, 25, 107–115. https://doi.org/10.1080/03115510108619216
304Feist, R., Lerosey-Aubril, R. & Johnson, R., 2010. Coaptative devices, enrolment, and life habits in Paralejurus, a particular case in scutelluid trilobites. Palaeobiodiversity and Palaeoenvironments, 90, 125–137. https://doi.org/10.1007/s12549-010-0025-1
305Fortey, R.A. & Owens, R.M., 1997. Evolutionary history. In Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, 249–287.
306Fortey, R.A. & Owens, R.M., 1999. Feeding habits in trilobites. Palaeontology, 42, 429–465. https://doi.org/10.1111/1475-4983.00080
307Franke, C., 2010. Marine Fauna der Wiltz-Schichten (Ober-Emsium, Unter-Devon) der Mulde von Wiltz und der Daleider Mulden-Gruppe (Luxemburg, Deutschland): Teil 1. In Franke (ed.), Beiträge zur Paläontologie des Unterdevons Luxemburgs (2). Ferrantia, 58, 5–62.
308Gandl, J., 1972. Die Acastavinae und Asteropyginae (Trilobita) Keltiberiens (NE-Spanien). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 530, 1–184.
309Ghobadi Pour, M., Popov, L.E., Hosseini, M., Adhamian, A. & Yazdi, M., 2013. Late Devonian (Frasnian) trilobites and brachiopods from the Soh area, Central Iran. Memoirs of the Association of Australasian Palaeontologists, 44, 149–158.
310Gibb, S. & Chatterton, B.D.E., 2010. Gerastos (Order Proetida; Class Trilobita) from the Lower to Middle Devonian of the southern Moroccan Anti-Atlas region. Palaeontographica Canadiana, 30, 1–89.
311Godefroid, J. & Cravatte, T., 1999. Les brachiopodes et la limite Silurien/Dévonien à Muno (sud de la Belgique). Bulletin de l’Institut royal des Sciences naturelles de Belgique, 69, 5–26.
312Guy, P.-M., 2020. Pérégrinations paléontologiques en Péninsule ibérique 6. Rencontres aussi inattendues qu’insolites à Polentinos. Fossiles, 42, 51–54.
313Haas, W., 1968. Trilobiten aus dem Silur und Devon von Bithynien (NW-Türkei). Palaeontographica (A), 130, 60–207.
314Haas, W., 1982. Preliminary notes of the Devonian SE of Istanbul (Turkey). In Sokolov, B.S. & Rzhonsnitskaya, M.A. (eds), Biostratigrafiya pogranichnykh otlozheniy nizhnego i srednego devona; Trudy polevoy sessii Mezhdunarodnoy podkomissii po stratigrafii devona; Samarkand, 1978. Akademiya Nauk SSSR, otdelenie geologii, geofiziki i geokhimii; Ministerstvo Geologii SSSR, Vsesoyuznyy Nauchno-Issledovatel’skiy Geologicheskiy Institut (VSEGEI), 144–147. [in Russian, with English summary].
315Haas, W. & Mensink, H., 1970 Asteropyginae aus Afghanistan (Trilobita). Abhandlungen der Mathematisch-Naturwissenschaftlichen Klasse, 1969, 1–60.
316Hammer, Ø., 2000. Spatial organization of tubercles and terrace lines in Paradoxides forchhammeri – evidence of lateral inhibition. Acta Palaeontologica Polonica, 45, 251–270.
317Harrington, H.J., 1959. General description of Trilobita. In Moore, R.C. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, O38–O117.
318Harrington, H.J., Moore, R.C. & Stubblefield, C.J., 1959. Morphological terms applied to Trilobita. In Moore, R.C. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, O117–O126.
319Hidalgo, F.J., Firstater, F.N., Lomovasky, B.J., Gallegos, P., Gamero, P. & Iribarne, O.O., 2010. Macroalgal fouling on the intertidal mole crab Emerita analoga facilitates bird predation. Helgoland Marine Research, 64, 367–376. https://doi.org/10.1007/s10152-010-0193-4
320Hoffman, M., Maguet, A. & Poullaouec, M., 2014. Les trilobites du Dévonien (Emsien-Famennien) de la rade de Brest, Finistère. Fossiles, 18, 50–62.
321Hollard, H., 1967. Le Dévonien du Maroc et du Sahara nord-occidental. In: Oswald, D.H. (ed.), International Symposium on the Devonian System, Calgary 1967. Vol. 1. Alberta Society of Petroleum Geologists, Calgary, 203–244.
322Hollard, H., 1974. Recherches sur la stratigraphie des formations du Dévonien moyen, de l’Emsien supérieur au Frasnien, dans le Sud du Tafilalt et dans le Ma’der (Anti-Atlas oriental). Notes du Service géologique de Maroc, 36/264, 7–68.
323Hollard, H., 1981. Tableaux de corrélations du Silurien et du Dévonien de l’Anti-Atlas. Notes du Service géologique de Maroc, 42/308, 23.
324Holloway, D.J., 1981. Silurian dalmanitacean trilobites from North America and the origins of the Dalmanitinae and Synphoriinae. Palaeontology, 24, 695–731.
325Holloway, D.J. & De Carvalho, M.G.P., 2009. The extraordinary trilobite Fenestraspis (Dalmanitidae, Synphoriinae) from the Lower Devonian of Bolivia. Palaeontology, 52, 933–949. https://doi.org/10.1111/j.1475-4983.2009.00878.x
326Holloway, D.J. & De Carvalho, M.G.P., 2010. The trilobite Chacomurus (Dalmanitidae, Synphoriinae) from the Lower Devonian of Bolivia. Memoirs of the Association of Australasian Palaeontologists, 39, 71–83.
327Hughes, N.C., Chapman, R.E. & Adrain, J.M., 1999. The stability of thoracic segmentation in trilobites: a case study in developmental and ecological constraints. Evolution & Development, 1, 24–35. https://doi.org/10.1046/j.1525-142x.1999.99005.x
328Hughes, N.C., Minelli, A. & Fusco, G., 2006. The ontogeny of trilobite segmentation: a comparative approach. Paleobiology, 32, 602–627. https://doi.org/10.1666/06017.1
329Jaanusson, V., 1975. Evolutionary processes leading to the trilobite suborder Phacopina. Fossils and Strata, 4, 209–218.
330Jansen, U., 2016. Brachiopod faunas, facies and biostratigraphy of the Pridolian to lower Eifelian succession in the Rhenish Massif (Rheinisches Schiefergebirge, Germany). In Becker, R.T., Königshof, P. & Brett, C.E. (eds), Devonian Climate, Sea Level and Evolutionary Events. Geological Society, London, Special Publications, 423, 45–122. http://doi.org/10.1144/SP423.11
331Jansen, U., Becker, G., Plodowski, G., Schindler, E., Vogel, O. & Weddige, K., 2004. Pragian and Emsian near Aouinet Torkoz (SW Dra Valley, Morocco). Documents de l’Institut Scientifique, Rabat, 19, 75–84.
332Jansen, U., Lazreq, N., Plodowski, G., Schemm-Gregory, M., Schindler, E. & Weddige, K., 2007. Neritic-pelagic correlation in the Lower and basal Middle Devonian of the Dra Valley (Southern Anti-Atlas, Moroccan Pre-Sahara). In Becker, R.T. & Kirchgasser, W.T. (eds), Devonian Events and Correlations. Geological Society, London, Special Publications, 278, 9–37. https://doi.org/10.1144/SP278.2
333Kácha, P. & Šarič, R., 2009. Host preferences in Late Ordovician (Sandbian) epibenthic bryozoans: example from the Zahořany Formation of Prague Basin. Bulletin of Geosciences, 84, 169–178. https://doi.org/10.3140/bull.geosci.1048
334Key, M.M., Volpe, J.W., Jeffries, W.B. & Voris, H.K., 1997. Barnacle fouling of the blue crab Callinectes sapidus at Beaufort, North Carolina. Journal of Crustacean Biology, 17, 424–439. https://doi.org/10.2307/1549437
335Key, M.M., Knauf, J.B. & Barnes, D.K.A., 2010. Epizoic bryozoans on predatory pycnogonids from the South Orkney Islands, Antarctica: “If you can’t beat them, join them”. In Ernst, A., Schäfer, P. & Scholz, J. (eds), Bryozoan Studies 2010. Springer, Berlin, Lecture Notes in Earth System Sciences, 143, 137–153. https://doi.org/10.1007/978-3-642-16411-8_10
336Khaldi, A.Y., Crônier, C., Hainaut, G., Abbache, A. & Ouali Mehadji, A., 2016. A trilobite faunule from the Lower Devonian of the Saoura Valley, Algeria: biodiversity, morphological variability and palaeobiogeographical affinities. Geological Magazine, 153, 357–387. https://doi.org/10.1017/S0016756815000345.
337Kloc, G.J., 1997. Epibionts on Dicranurus and some related genera. 2nd International Trilobite Conference. Brock University, St. Catharines, Ontario, August 22-24, 1997. Abstracts.
338Klug, C., Kröger, B., Korn, D., Rücklin, M., Schemm-Gregory, M., De Baets, K. & Mapes, R.H., 2008. Ecological change during the early Emsian (Devonian) in the Tafilalt (Morocco), the origin of the Ammonoidea, and the first African pyrgocystid edrioasteroids, machaerids and phyllocarids. Palaeontographica (A), 283, 83–176. https://doi.org/10.1127/pala/283/2008/83
339Kowalski, H., 1990. Trilobiten aus dem Devon der Eifel. Der Aufschluss, Sonderband, 33, 11–63.
340Laurencin, M. & Hoffman, M., 2012. Le Dévonien de la rade de Brest, Finistère. Fossiles, 10, 48–50.
341Lawrance, P. & Stammers, S., 2014. Trilobites of the world. Siri Scientific Press, Manchester, 416 p.
342Lebrun, P., 2002. Trilobites de France. Tome I – Généralités sur les trilobites, Massif armoricain (Bretagne, Normandie, Vendée). Minéraux & Fossiles, Hors-Série, 14, 1–131.
343Lebrun, P., 2018. Fossiles du Maroc. Tome I. Gisements emblématiques du Paléozoïque de l’Anti-Atlas. Les Editions du Piat, Saint-Julien-du-Pinet, 298 p.
344Le Maître, D., 1952. La faune du Dévonien inférieur et moyen de la Souara et des abords de l’Erg El Djemel (Sud Oranais). Matériaux pour la Carte géologique de l’Algérie. 1re série, paléontologie, 12, 1–170.
345Lerosey-Aubril, R. & Angiolini, L., 2009. Permian trilobites from Antalya Province, Turkey, and enrolment in Late Palaeozoic trilobites. Turkish Journal of Earth Sciences, 18, 427–448. https://doi.org/10.3906/yer-0801-5
346Lerosey-Aubril, R. & Peel, J.S., 2018. Gut evolution in Early Cambrian trilobites and the origin of predation on infaunal macroinvertebrates: evidence from muscle scars in Mesolenellus. Palaeontology, 61/5, 747–760. https://doi.org/10.1111/pala.12365
347Lerosey-Aubril, R., Hegna, T.A. & Olive, S., 2011. Inferring internal anatomy from the trilobite exoskeleton: the relationship between frontal auxiliary impressions and the digestive system. Lethaia, 44, 166–184. https://doi.org/10.1111/j.1502-3931.2010.00233.x
348Lespérance, P.J., 1975. Stratigraphy and paleontology of the Synphoriidae (Lower and Middle Devonian dalmanitacean trilobites). Journal of Paleontology, 49, 91–137.
349Levi-Setti, R., 2014. The trilobite book. The University of Chicago Press, Chicago, 273 p.
350Lieberman, B.L. & Kloc, G.J., 1997. Evolutionary and biogeographic patterns in the Asteropyginae (Trilobita, Devonian) Delo, 1935. Bulletin of the American Museum of Natural History, 232, 1–127.
351McKellar, R.C. & Chatterton, B.D.E., 2009. Early and Middle Devonian Phacopidae (Trilobita) of southern Morocco. Palaeontographica Canadiana, 28, 1–109.
352McKinney, M.L. & McNamara, K.J., 1991. Heterochrony: the Evolution of Ontogeny. Plenum Press, New York, 437 p. https://doi.org/10.1007/978-1-4757-0773-1
353McNamara, K.J. & Tuura, M.E., 2011. Evidence for segment polarity during regeneration in the Devonian asteropygine trilobite Greenops widderensis. Journal of Paleontology, 85, 106–110. https://doi.org/10.1666/10-049.1
354Miller, J., 1976 The sensory fields and life mode of Phacops rana (Green, 1832) (Trilobita). Transactions of the Royal Society of Edinburgh, 69, 337–367. https://doi.org/10.1017/S0080456800015350
355Minelli, A., Fusco, G. & Hughes, N.C., 2003. Tagmata and segment specification in trilobites. Special Papers in Palaeontology, 70, 31–43.
356Monseur, G., 1959. Sur un trilobite déterminé comme Asteropyge punctata (Steininger). Annales de la Société géologique de Belgique, 82, B89–97.
357Morris, S.F., 1988. A review of British trilobites, including a synoptic revision of Salter’s monograph. Monographs of the Palaeontographical Society, 140/574, 1–316. https://doi.org/10.1080/25761900.2022.12131758
358Morzadec, P., 1969. Le Dévonien de la rive nord de la Rivière du Faou (Finistère). Étude stratigraphique. Étude des trilobites. Bulletin de la Société minéralogique et géologique de Bretagne, 1969, 1–58.
359Morzadec, P., 1983. Trilobites du Dévonien (Emsien-Famennien) de la Rade de Brest (Massif Armoricain). Palaeontographica (A), 191, 103–184.
360Morzadec, P., 1990. Évolution, biozonation et biogéographie de Protacanthina Gandl, trilobite du Dévonien inférieur nord-gondwanien. Geobios, 23, 719–735. https://doi.org/10.1016/S0016-6995(06)80338-4
361Morzadec, P., 1992 Evolution des Asteropyginae (Trilobita) et variations eustatiques au Dévonien. Lethaia, 25, 85–96. https://doi.org/10.1111/j.1502-3931.1992.tb01793.x
362Morzadec, P., 1997. Les trilobites Asteropyginae du Dévonien de l’Ougarta (Algérie). Palaeontographica (A), 244, 143–158. https://doi.org/10.1127/pala/244/1997/143
363Morzadec, P., 2001. Les trilobites Asteropyginae du Dévonien de l’Anti-Atlas (Maroc). Palaeontographica (A), 262, 53–85. https://doi.org/10.1127/pala/262/2001/53
364Morzadec, P., 2002. Trilobites Asteropyginae dévoniens d’Iran. Geobios, 35, 411–427. https://doi.org/10.1016/S0016-6995(02)00037-2
365Müller, P., 2005. Revision der Gattung Scabrella (Trilobita; Unter-Devon). Geologica et Palaeontologica, 39, 1–27.
366Müller, P. & Alberti, M., 2010. Burmeisterella vixarmata aus der Klerf-Formation in Nord-Luxemburg (Trilobita, oberes Unter-Devon). In Franke (ed.), Beiträge zur Paläontologie des Unterdevons Luxemburgs (2). Ferrantia, 58, 63–71.
367Ormiston, A.R., 1976. New Middle Devonian trilobites from northwestern Canada. Journal of Paleontology, 50, 1162–1174.
368Pillet, J., 1961. Contribution à l’étude des faunes de trilobites du Zemmour (Mauritanie septentrionale). Annales de la Faculté des Sciences de l’Université de Dakar, 6, 93–113.
369Pillet, J., 1973. Les trilobites du Dévonien inférieur et du Dévonien moyen du Sud-Est du Massif armoricain. Société d’Études Scientifiques de l’Anjou, Mémoire, 1, 1–307.
370Richter, D., 1964. Der geologische Bau des südwestlichen Teiles des Massives von Stavelot (Belgien) unter besonderer Berücksichtigung einer tektonischen Prägung. Forschungsberichte des Landes Nordrhein-Westfalen, 1203, 5–83. https://doi.org/10.1007/978-3-663-06957-7
371Richter, R. & Richter, E., 1942. Die Trilobiten der Weismes-Schichten am Hohen Venn, mit Bemerkungen über die Malvinocaffrische Provinz. Senckenbergiana, 25, 156–179.
372Richter, R. & Richter, E., 1943. Studien im Paläozoikum der Mittelmeer-Länder. 4a. Trilobiten aus dem Devon von Marokko, mit einem Anhang über Arten des Rheinlands. Senckenbergiana, 26, 116–198.
373Richter, R. & Richter, E., 1954. Die Trilobiten des Ebbe-Sattels und zu vergleichende Arten. (Ordovizium, Gotlandium/Devon). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 488, 1–76.
374Rustán, J.J., Waisfeld, B.G. & Vaccari, N.E., 2020. The homalonotid trilobite Burmeisteria Salter, 1865 in the Lower Devonian of Argentina: new data in the context of southwestern Gondwana. Journal of Paleontology, 94, 498–512. https://doi.org/10.1017/jpa.2019.104
375Saleh, F., Vidal, M., Laibl, L., Sansjofre, P., Gueriau, P., Pérez-Peris, F., Lustri, L., Lucas, V., Lefebvre, B., Pittet, B., El Hariri, K. & Daley, A.C., 2020. Large trilobites in a stress-free Early Ordovician environment. Geological Magazine, 158, 261–270. https://doi.org/10.1017/S0016756820000448
376Schemm-Gregory, M. & Jansen, U., 2009. The Silurian of the Goldsteintal (Rheinisches Schiefergebirge, Germany). Rendiconti della Società Paleontologica Italiana, 3, 339–340.
377Schoenemann, B., Clarkson, E.N.K. & Høyberget, M., 2017. Traces of an ancient immune system–how an injured arthropod survived 465 million years ago. Scientific Reports, 7, 40330. https://doi.org/10.1038/srep40330
378Schraut, G., 1998. Trilobiten aus dem Unter-Devon des südlichen Antiatlas (Marokko) – ihre Bedeutung für die zeitliche Korrelation zwischen Rheinischer und Herzynischer Fazies. Senckenbergiana lethaea, 77, 61–69. https://doi.org/10.1007/BF03043733
379Schraut, G., 2000. Trilobiten aus dem Unter-Devon des südöstlichen Anti-Atlas, Süd-Marokko. Senckenbergiana lethaea, 79, 361–433. https://doi.org/10.1007/BF03043647
380Scotese, C.R., 2001. Atlas of Earth History. PALEOMAP Project. University of Texas at Arlington.
381Scotese, C.R. & McKerrow, W.S., 1990. Revised World maps and introduction. In McKerrow, W.S. & Scotese, C.R. (eds), Palaeozoic Palaeogeography and Biogeography. Geological Society, London, Memoirs, 12, 1–21. https://doi.org/10.1144/GSL.MEM.1990.012.01.01
382Smeenk, Z., 1983. Devonian trilobites of the southern Cantabrian Mountains (northern Spain) with a systematic description of the Asteropyginae. Leidse Geologische Mededelingen, 52, 383–511.
383Šnajdr, M., 1980. Bohemian Silurian and Devonian Proetidae (Trilobita). Rozpravy Ústředního ústavu geologického, 45, 1–324.
384Speyer, S.E., 1988. Biostratinomy and functional morphology of enrolment in two Middle Devonian trilobites. Lethaia, 21, 121–138. https://doi.org/10.1111/j.1502-3931.1988.tb02063.x
385Speyer, S.E., 1990. Enrollment in trilobites. In Boucot, A.J. (ed.), Evolutionary Paleobiology of Behavior and Coevolution. Elsevier, Amsterdam, 450–455. https://doi.org/10.1016/B978-0-444-88034-5.50021-2
386Storey, A.J., 2012. Late Silurian trilobite palaeobiology and biodiversity. Unpublished Ph.D. thesis, University of Birmingham, Birmingham, 389 p.
387Størmer, L., 1980. Sculpture and microstructure of the exoskeleton in chasmopinid and phacopid trilobites. Palaeontology, 23, 237–271.
388Straw, S.H., 1933. The fauna of the Palaeozoic rocks of the Little Missenden Boring. Summary of Progress of the Geological Survey of Great Britain, 1932, 112–142.
389Struve, W., 1959a. Subfamily Asteropyginae Delo, 1935. In Moore, R.C. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, O477–O483.
390Struve, W., 1959b. Suborder Phacopina nov. In Moore, R.C. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, O461, O462.
391Stürmer, W. & Bergström, J., 1973. New discoveries on trilobites by X-rays. Paläontologische Zeitschrift, 47, 104–141. https://doi.org/10.1007/BF02989565
392Teigler, D.J. & Towe, K.M., 1975. Microstructure and composition of the trilobite exoskeleton. Fossils and Strata, 4, 137–149.
393Thomas, A.T., Owens, R.M. & Rushton, A.W.A., 1984. Trilobites in British stratigraphy. Geological Society, London, Special Report, 16, 1–78.
394Thormann, F. & Weddige, K., 2001. Addendum zu Struve, W. (1992), Neues zur Stratigraphie und Fauna des rhenotypen Mittel-Devon: Abbildungen der Holotypen. Senckenbergiana lethaea, 81, 307–327. https://doi.org/10.1007/BF03042784
395Trilles, J.-P. & Hipeau-Jacquotte, R., 2012. Symbiosis and parasitism in the Crustacea. In Forest, J. & von Vaupel Klein, C. (eds), Treatise on Zoology - Anatomy, Taxonomy, Biology. The Crustacea, Volume 3. Brill, Leiden, 239–317. https://doi.org/10.1163/9789004188259_006
396Van Viersen, A.P., 2013. Latest Early to early Middle Devonian acastid trilobites from the eastern part of the Dinant Synclinorium, Belgium (Rhenohercynian Zone). Memoirs of the Association of Australasian Palaeontologists, 44, 1–10.
397Van Viersen, A.P., 2021a. Type and other species of Gerastos and allied genera (Trilobita, Proetinae) from the Siluro-Devonian. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 299, 185–217. https://doi.org/10.1127/njgpa/2021/0964
398Van Viersen, A.P., 2021b. Systematics of Devonian trochurine trilobites (Lichidae). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 300, 175–187. https://doi.org/10.1127/njgpa/2021/0983
399Van Viersen, A.P. & Bignon, A., 2011. Late Devonian (Frasnian) asteropygine trilobites from the Frasnes area, southern border of Dinant Synclinorium, Belgium. Geologica Belgica, 14, 109–128.
400Van Viersen, A.P. & Holland, D., 2016. Morphological trends and new species of Cyphaspis (Trilobita, Otarioninae) in the Devonian of Morocco, Turkey, Germany and Belgium. Geologica Belgica, 19, 251–271. http://dx.doi.org/10.20341/gb.2016.008
401Van Viersen, A.P. & Lerouge, F., 2019. Cornuproetine (proetide) trilobites with nine thorax segments from the Devonian of Morocco, Germany and the Czech Republic. PalZ., Paläontologische Zeitschrift, 94, 227–254. https://doi.org/10.1007/s12542-019-00491-w
402Van Viersen, A.P. & Lerouge, F., 2021. Timsaloproetus alissae sp. nov. (Trilobita: Proetidae) from the Lower Devonian of southern Morocco. PalZ., Paläontologische Zeitschrift, 95, 223–230. https://doi.org/10.1007/s12542-020-00543-6
403Van Viersen, A.P. & Vanherle, W., 2018. The rise and fall of Late Devonian (Frasnian) trilobites from Belgium: taxonomy, biostratigraphy and events. Geologica Belgica, 21, 73–94. https://doi.org/10.20341/gb.2018.005
404Van Viersen, A.P., Holland, D. & Koppka, J., 2017. The phacopine trilobite genera Morocops Basse, 2006 and Adrisiops gen. nov. from the Devonian of Morocco. Bulletin of Geosciences, 92, 13–30. https://doi.org/10.3140/bull.geosci.1610
405Van Viersen, A.P., Taghon, P. & Magrean, B., 2019. Early Middle Devonian trilobites and events in the Nismes - Vireux-Molhain area, southern border of the Dinant Synclinorium (Belgium, northern France). Geologica Belgica, 22, 7–33. https://doi.org/10.20341/gb.2019.001
406Waugh, D.A., Feldmann, R.M., Crawford, R.S., Jakobsen, S.L. & Thomas, K.B., 2004. Epibiont preservational and observational bias in fossil marine decapods. Journal of Paleontology, 78, 961–972. https://doi.org/10.1666/0022-3360(2004)078%3C0961:EPAOBI%3E2.0.CO;2
407Wendt, J., 2021. Middle and Late Devonian paleogeography of the eastern Anti-Atlas (Morocco). International Journal of Earth Sciences, 110, 1531–1544. https://doi.org/10.1007/s00531-021-02028-6
408Whiteley, N.M., 2011. Physiological and ecological responses of crustaceans to ocean acidification. Marine Ecology Progress Series, 430, 257–271. https://doi.org/10.3354/meps09185
409Whiteley, T.E., Kloc, G.J. & Brett, C.E., 2002. Trilobites of New York. Cornell University Press, Ithaca, 203 p.
410Whittington, H.B., 1997. Morphology of the exoskeleton. In Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, 1–85.
411Whittington, H.B. & Kelly, S.R.A., 1997. Morphological terms applied to Trilobita. In Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1. Trilobita, revised. Volume 1: Introduction, Order Agnostida, Order Redlichiida. University of Kansas Press, Lawrence, Kansas and Geological Society of America, Boulder, Colorado, 313–329.
412Yolkin, E.A. & Ormiston, A.R., 1985. Representatives of the genus Terranovia (Trilobita) from the Territory of the Soviet Union and Arctic North America. Journal of Paleontology, 59, 461–474.
413Zapalski, M. & Klug, C., 2018. Trilobite sclerites as attachment surface for Emsian tabulate corals of Hamar Laghdad (Anti-Atlas, Morocco). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 290, 111–115. https://doi.org/10.1127/njgpa/2018/0769
414Zhou, Z. & Campbell, K.S.W., 1990. Devonian phacopacean trilobites from the Zhusilenghaierhan region, Ejin Qi, western Inner Mongolia, China. Palaeontographica (A), 214, 57–77.
415Manuscript received 31.05.2022, accepted in revised form 30.09.2022, available online 05.12.2022.
