


- Accueil
- Volume 28 (2025)
- number 3-4
- First report on the occurrence of the echinoderm classes Soluta and Stylophora in a new Lower Devonian (Pragian) Konservat-Lagerstätte from southern Belgium


Visualisation(s): 1011 (24 ULiège)
Téléchargement(s): 455 (5 ULiège)

First report on the occurrence of the echinoderm classes Soluta and Stylophora in a new Lower Devonian (Pragian) Konservat-Lagerstätte from southern Belgium

Abstract
In the Neufchâteau–Eifel Synclinorium (southern Belgium), remnants of the spoil heap of the disused and now flooded underground Sainte-Barbe Quarry (Warmifontaine) yielded an invertebrate Pragian assemblage occurring in the dark monotonous slates of the La Roche Formation (Martelange Member). This locality is considered as a new Belgian Konservat-Lagerstätte (fossil conservation deposit) due to the presence of both biomineralised and soft-bodied organisms. Two specimens from the Sainte-Barbe Quarry Biota are described and assigned to two distinct classes of extinct echinoderms. The first one corresponds to an almost complete, flattened and distorted dendrocystitid solutan, which is tentatively identified as Dehmicystis? sp., based on similarities with D. globulus from the Hunsrück Slate (Emsian) of Germany. The second echinoderm specimen is a well-preserved, flattened, fully articulated and almost complete mitrate stylophoran, morphologically close to Mitrocystites? styloideus-like peltocystidans from the Lower and Middle Devonian of Germany. Taphonomic attributes of the two echinoderms from Warmifontaine (e.g. both fully articulated; mitrate with its aulacophore preserved in feeding position) suggest they were probably killed and entombed by obrution deposits in an otherwise quiet and deep setting. Other echinoderms (asteroids, crinoids, blastoids, cyclocystoids, ophiuroids) reported from the Lower Devonian of Belgium are also briefly discussed and illustrated (e.g. holotypes).
Table des matières
1. Introduction
1Relatively similar morphologies were acquired independently and convergently in at least three groups of epibenthic, free-living (unattached) Palaeozoic echinoderms (pleurocystitids, solutans, and stylophorans), very likely as adaptations to a comparable ‘homalozoan’ mode of life on soft substrates (Parsley, 1972; Lefebvre, 2007; Parsley & Sumrall, 2007). These three groups are characterised by a reduced number of ambulacra (one or two), a flattened, more or less bilaterally symmetrical theca, and the possession of few, if any, respiratory structures (e.g. rhombs). Unsurprisingly, pleurocystitids, solutans and stylophorans are often co-occurring in the same assemblages, and their fossil records are very similar.
2All three groups underwent a significant increase in diversity and palaeobiogeographic distribution during the Ordovician, with an acme in the Sandbian–Katian interval (Sprinkle & Guensburg, 2004; Lefebvre et al., 2013). Pleurocystitids, solutans and stylophorans were strongly affected by the Late Ordovician Mass Extinction (LOME) in the Hirnantian. Rare occurrences of Silurian stylophorans have been documented in Australia (Gill & Caster, 1960; Philip, 1981; Caster, 1983; Ruta, 1997; Ruta & Jell, 1999a, b), Canada (Lefebvre & Ausich, 2021), Mongolia (Rozhnov, 1990), Sweden (Lindström, 1888; Regnéll, 1945, 1960; Reich et al., 2007), and the United Kingdom (de Koninck, 1869; Jefferies & Lewis, 1978). The Silurian record of pleurocystitids is totally cryptic, while solutans are only known by two occurrences: in Spain (Zamora & Gutiérrez-Marco, 2023) and in the United Kingdom (Nohejlová & Lefebvre, 2022). In contrast, all three groups are characterised by a second (lower) peak of diversity in the Early Devonian (McIntosh & Macurda, 1979). In particular, pleurocystitids, solutans and stylophorans are co-occurring in the Lower Devonian of Australia (Humevale Formation; Gill & Caster, 1960; Jell, 1983), Germany (Hunsrück Slate; Dehm, 1932, 1934), and the USA (Bois d’Arc Formation; Parsley & Sumrall, 2007).
3The aim of this paper is to report the first occurrence of two of these groups (solutans and stylophorans) in the Lower Devonian (Pragian) of southern Belgium, and to discuss their palaeobiogeographic affinities.
2. Geological setting
4The two studied specimens were collected in June 2021 (RBINS a14020) and 2022 (RBINS a14021) at Warmifontaine (Fig. 1), a small village of the commune of Neufchâteau that is established on the northern flank of the Neufchâteau–Eifel Synclinorium (Asselberghs, 1946). This major tectonic unit belongs to the Ardenne Allochthon that was overthrust on the Brabant Parautochthon during the Variscan orogeny (e.g. Belanger et al., 2012). The echinoderms were found on the remnants of the spoil heap of the disused and now flooded underground Sainte-Barbe Quarry situated in the centre of the village (e.g. Asselberghs, 1924; Watteyne, 2016); consequently, no information about their precise position in the lithological column is available. The residual spoil heap, which was particularly huge in the past (see Watteyne, 2016, fig. 193), was converted over the last decade into slate chippings for garden, and the chances of making new discoveries will become slim in the future. According to Ghysel (2022), the Sainte-Barbe Quarry intensively exploited the dark grey to black, monotonous slates of the local Martelange Member of the La Roche Formation (Fig. 2) until the early 2000s. This basal member is distinguished from the rest of the La Roche Formation (see description in Denayer & Mottequin, 2024) by the absence of sandy laminae and sandstone beds (e.g. Ghysel, 2022), which allowed the production of high-quality slates at Warmifontaine as reflected by the awards obtained at the Liège (1905) and Brussels International expositions (1910) (Watteyne, 2016). The thickness of the Martelange Member is comprised between 100 and 150 m in its type area (Ghysel, 2023) on the Belgian–Luxembourg border, but that of the La Roche Formation is highly variable from the southern margin of the Dinant Synclinorium up to the Neufchâteau–Eifel Synclinorium (see Denayer & Mottequin, 2024) (Fig. 1): from 215 m south of Couvin (Stainier, 1994) to more than 800 m in the Ardenne Anticlinorium (Dejonghe & Hance, 2001), whereas Ghysel (2022) estimated its minimal thickness at 700 m (300 m for the Martelange Member) in the western part of the Neufchâteau–Eifel Synclinorium (Fig. 1). The La Roche Formation (Fig. 2) is mostly Pragian (Siegenian) in age, but it is probable that its upper part, in the Couvin area, is early Emsian in age based on the presence of the conodont Caudicriodus celtibericus (Bultynck et al., 2000). Macrofossils are rare and poorly diverse within the Martelange Member (Fig. 2) as shown by the few available faunal lists (Dewalque, 1891; Maillieux, 1940; Asselberghs, 1946) that were based notably on the collections of the Royal Belgian Institute of Natural Sciences and of the Liège University. All these old collections essentially include solitary rugose corals, bivalves, orthoconic cephalopods and trilobites as is the case of those recently discovered at the Centre Grégoire Fournier of the Maredsous Abbey (Fig. 3), where Mottequin (2021) also pointed out the existence of an enigmatic fossil from Warmifontaine, which was considered as a putative early chordate by Olive et al. (2022). The specimens are compressed, a characteristic inherent to fossils preserved in slate, and sometimes elongated, due to the tectonic strains they have undergone. Despite our local requests, we were unable to find any other Warmifontaine collections assembled by amateur geologists of the Neufchâteau area.
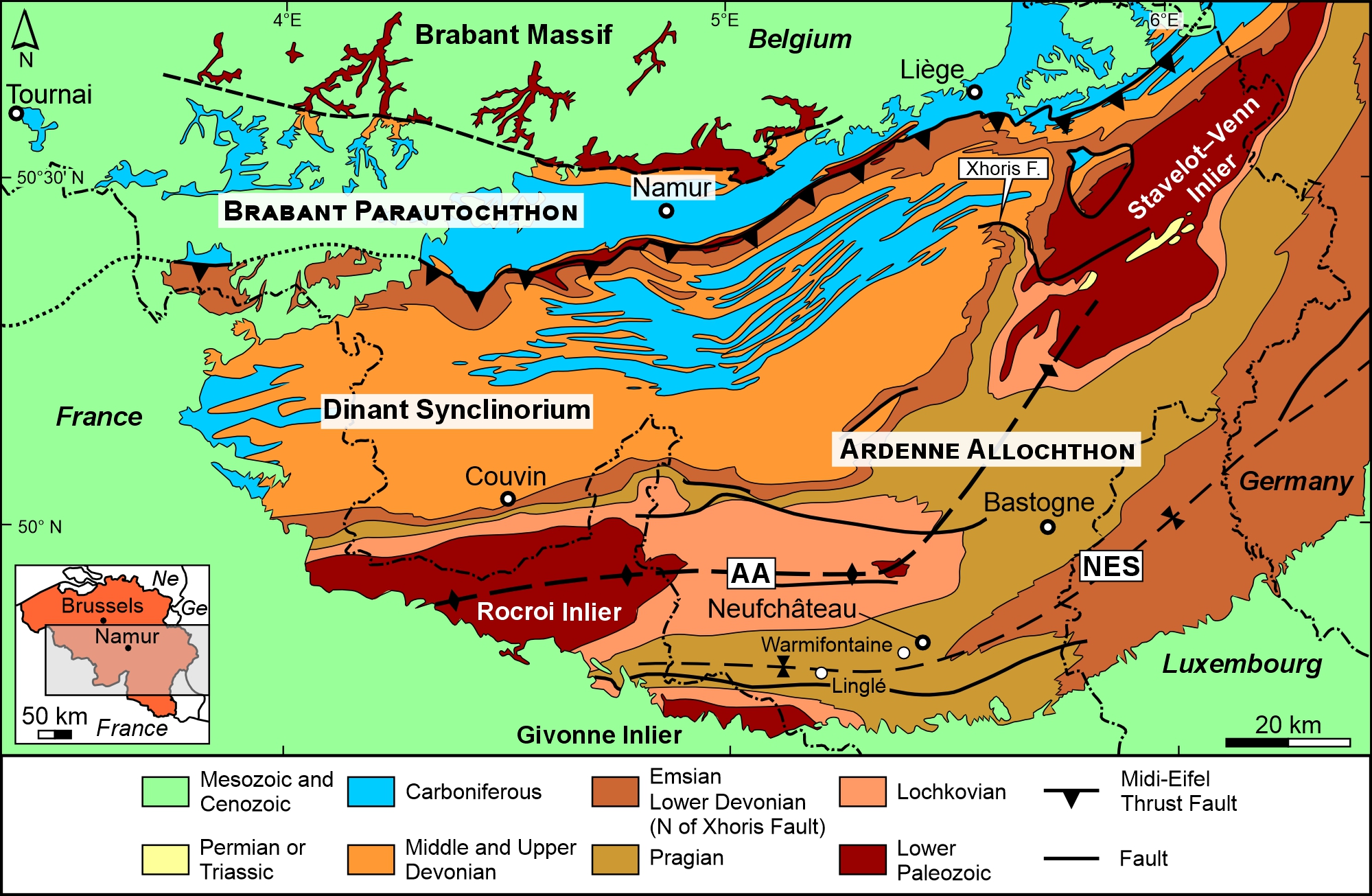
Figure 1. Simplified geological map of southern Belgium (modified from de Béthune, 1954 and Candela & Mottequin, 2022) with the location of the village of Warmifontaine (Sainte-Barbe Quarry) and the hamlet of Linglé (Mortehan). Abbreviations: AA, Ardenne Anticlinorium; F., Fault; NES, Neufchâteau–Eifel Synclinorium. Abbreviations in the insert: Ge, Germany; Ne, the Netherlands.
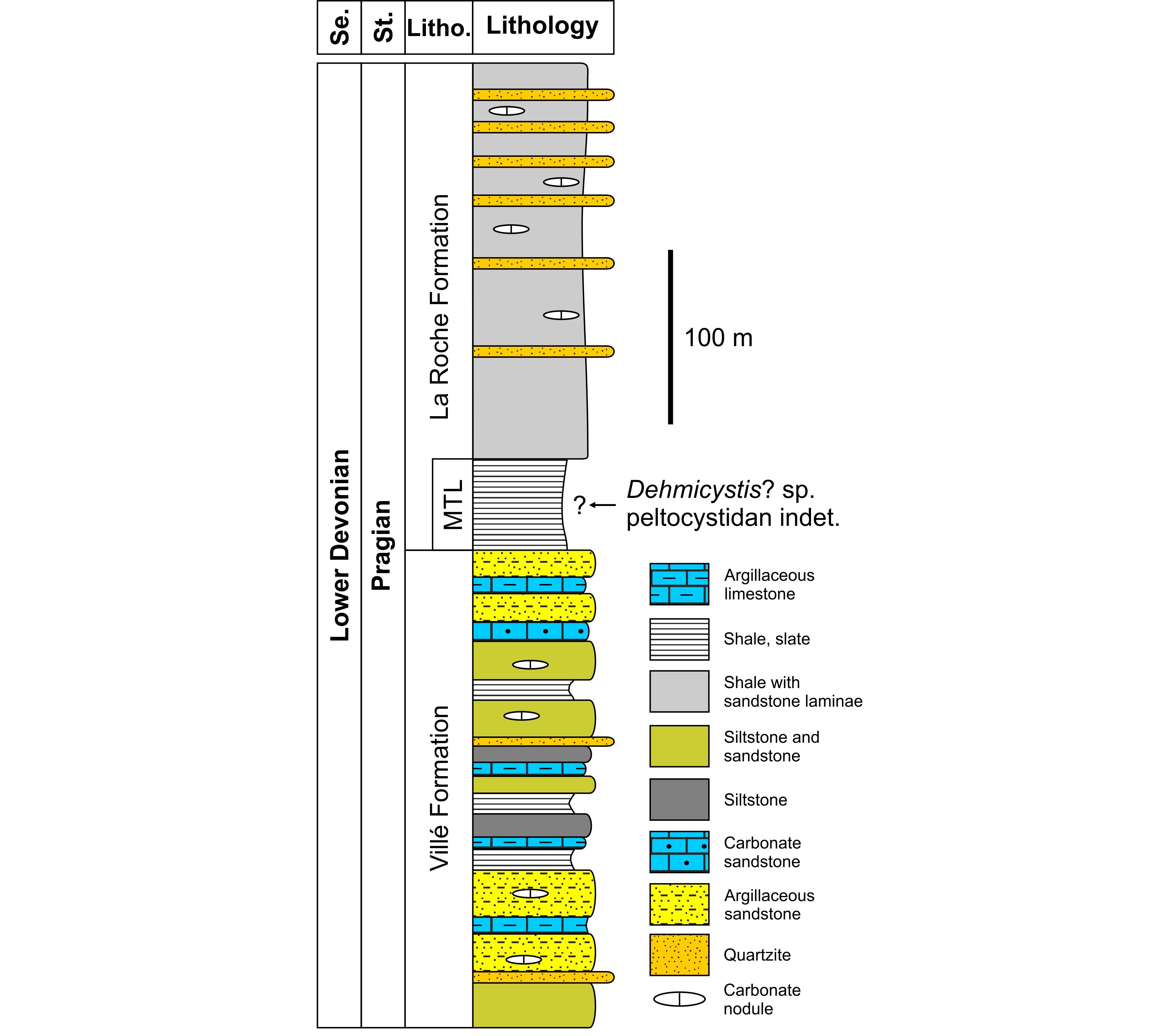
Figure 2. Schematic lithological column of the Pragian lithostratigraphic units (pars) encountered in the western part of the Neufchâteau–Eifel Synclinorium (modified from Ghysel, 2022). The range of the two studied echinoderms within the lower part of the La Roche Formation is unknown. Abbreviations: Litho., Lithostratigraphy; MTL, Martelange Member; Se., Series; St., Stage.
5Recent fieldwork at Warmifontaine in 2021 and 2022 yielded hundreds of fossils that are generally poorly preserved and quite difficult to identify, but some of which have preserved soft parts (Cincotta et al., 2022, 2023). Besides the two echinoderms, the fauna includes arthropods, molluscs, corals, unidentified fossils and specimens probably similar to that mentioned by Mottequin (2021) and Olive et al. (2022). However, the fossils recently collected on the spoil heap of the Sainte-Barbe Quarry are markedly different from those present in the old aforementioned collections, by their preservation and their taxonomic identification. Either they come from a horizon that was not exploited during the first decades of slate mining, or there was a strong collection bias in the past.
6The high degree of disarticulation of the fossils and the scarcity of large specimens indicate that they would have been transported over an unknown distance, but it is not excluded that some were buried in situ, notably the echinoderms described in this paper, as is the case of the fossils of the Lower Devonian Hunsrück Slate deposits of Germany (Brett & Seilacher, 1991; Bartels et al., 1998; Sutcliffe et al., 1999). The Belgian locality constitutes a new Lower Devonian Konservat-Lagerstätte (fossil conservation deposit sensu Shields, 1998), with the preservation of both biomineralised and soft-bodied organisms. However, further studies are required for identifying problematic taxa that would shed more light on the biodiversity of this new fossil locality.

Figure 3. Pyritised and distorted invertebrates from the Sainte-Barbe Quarry at Warmifontaine, La Roche Formation (Martelange Member), Pragian. A. Orthoconic cephalopod (identified as Kionoceras rhysum in Maillieux, 1940), RBINS a14160. B. Bivalve, CGF II. 232. C. Bivalve (identified as Puella cf. elegantissima in Maillieux, 1940), RBINS a14161. D. Homalonotid trilobite in dorsal view (pygidium and thorax), CGF II. 69.
3. Material and methodology
7The two echinoderm specimens from Warmifontaine are deposited at the Royal Belgian Institute of Natural Sciences, Brussels (prefixed RBINS). The other specimens illustrated here are deposited at the Liège University (prefixed PA.ULg), the Centre Grégoire Fournier of the Maredsous Abbey (prefixed CGF), and the Naturhistorisches Museum Mainz (prefixed NHMMZ PWL).
8The specimens from Warmifontaine and Mortehan (RBINS) and Bundenbach (NHMMZ PWL) were photographed using a Nikon D850 digital camera, equipped with a 70mm f/2.8 Sigma macro lens and a Canon EOS 750D digital camera, equipped with a Canon EF 100mm f/2.8L macro IS USM macro lens, respectively. The other echinoderms were coated with ammonium chloride and photographed using an Olympus OM-D E-M10 Mark II digital camera, equipped with an Olympus M.Zuiko Digital ED 60 mm macro lens, with the exception of the single blastoid (PAULg) selected for scanning electron microscopy that was uncoated and imaged with an ESEM FEI Quanta 200, under low vacuum.
9The morphology of one of the specimens (RBINS a14021) was further documented using synchrotron-based micro-X-ray fluorescence (µXRF) major-to-trace elemental and laboratory-based X-ray microcomputed tomography (µCT).
10µXRF was performed at the PUMA beamline of the SOLEIL synchrotron source (France) (Schöder et al., 2024), using a monochromatic beam of 18.5 keV, selected for excitation of K-lines from phosphorus to zirconium and L-lines from cadmium to uranium. The incoming X-ray beam was focused using Kirkpatrick-Baez mirrors down to a spot size of ~7 × 5 μm2 (H × V, full width at half maximum). The specimen was mounted on a scanner stage allowing 150 mm and 100 mm movements (in horizontal and vertical directions respectively) with micrometre accuracy, and orientated at 30° to the incident beam, producing an effective beam size of ~8 × 5 μm2 (H × V) on the specimen. XRF was collected using a SiriusSD silicon drift detector (SGX Sensortech Ltd, 80 mm2 active area) oriented at 90° to the incident beam, in the horizontal plane. Two-dimensional spectral images, i.e. images for which each pixel is characterized by a full XRF spectrum, were collected on the fly over the specimen at a 100 µm lateral resolution with a 40 ms dwell time (effective counting time was 90% of the dwell time). We present herein a false-colour RGB overlay of the distributions of Fe (red), Pb (green), and Ni (blue), reconstructed from integrated intensities in spectral regions of interest (peaks) using the PyMCA data-analysis freeware (Solé et al., 2007). Elemental distributions are presented using a logarithmic colour scale that goes from dark to light, corresponding respectively to lower and higher concentrations.
11µCT was carried out at the RBINS, using a RX EasyTom instrument (RX Solutions, Chavanod, France; https://www.rx-solutions.com/ ). The tube power, voltage, and current were 30 W, 150 kV, and 200 µA, respectively, with a 0.4 mm copper filter. The scan parameters included a frame rate of 4.9, an average of four frames per image, a horizontal shift of 2, and a vertical stack of 4, capturing 2880 individual radiographs per 360° rotation, in middle spot mode, and an isotropic voxel size of 48.86 μm. The resulting 16-bit TIFF stack was inspected in 2D and 3D with Dragonfly ORS software (from Object Research Systems Inc, Montreal, Canada). Since the slab containing the specimen was not unidimensional, beam hardening was mitigated by scanning it alongside three other Sainte-Barbe slabs of similar dimensions.
4. Systematic palaeontology
12Phylum Echinodermata Klein, 1778
13Subphylum Blastozoa Sprinkle, 1973
14Remarks. The feeding appendages of solutans and stylophorans are considered here as free (erect) ambulacral structures, i.e. brachioles (see e.g. Haugh & Bell, 1980; Lefebvre, 2022; Guensburg et al., 2023). The possession of brachioles represents the defining apomorphy of blastozoans (Sprinkle, 1973; David et al., 2000; Nardin et al., 2017). Solutans and stylophorans are therefore interpreted here as two distinct clades belonging to this subphylum (Mooi et al., 2024).
15Class Soluta Jaekel, 1901
16Order Dendrocystitida Noailles et al., 2014
17Family Dendrocystitidae Bassler, 1938
18Genus Dehmicystis Caster, 1967
19Type species. Dendrocystites (Dendrocystoides?) globulus Dehm, 1934 from the Lower Devonian (lower Emsian) of Germany.
20Remarks. The possession of large, well-defined suranal plates (‘sugar loaf’ plates) strongly supports the assignment of Dehmicystis to the family Dendrocystitidae (Dehm, 1934; Caster, 1967; Noailles et al., 2014; Zamora & Gutiérrez-Marco, 2023). However, several aspects of its morphology remain poorly known, due to the limited number of available specimens and tectonic distortion (Dehm, 1934; Rahman & Lintz, 2012). The type species, D. globulus, was originally described from the Hunsrück Slate (Emsian) of Rhineland-Palatinate, Germany, based on its holotype and only known specimen (Dehm, 1934; Caster, 1967). More recently, three additional individuals, all located on a same slab, were documented (Rahman & Lintz, 2012). All other figured specimens assigned to D. globulus correspond to peltocystidan mitrates (see below). Dehmicystis was recently also reported from the Silurian (Ludlow Series) Llagarinos Formation of the Central Iberian Zone, Spain (Zamora & Gutiérrez-Marco, 2023), therefore considerably extending its stratigraphic range and palaeobiogeographic distribution. The holotype and only known specimen of the Spanish Dehmicystis (D. ariasi) is particularly well-preserved and makes it possible to document several poorly known morphological features of the genus (Zamora & Gutiérrez-Marco, 2023). For example, the brachiole is not inserted in apical position, but more posteriorly, on the lower (plano-concave) thecal surface. Moreover, the periproct is not located in lateral position, at the posterior extremity of the anal lobe, but displaced on the upper (plano-convex) thecal surface (Zamora & Gutiérrez-Marco, 2023).
21Dehmicystis? sp.
22(Fig. 4A–B)
23Material. RBINS a14020, part and counterpart.
24Description. Part and counterpart of strongly flattened organism consisting of two distinct parts: elongate, pear-shaped ‘body’ (~6.0 cm long, up to 2.5 cm wide), and ~8.8 cm long, narrow (~0.1–0.15 cm wide), segmented ‘appendage’ (Fig. 4A–B).
25Body outline nearly bilaterally symmetrical, with two opposite, unequal, rounded extremities, and elongate, gently convex lateral sides. Lateral edges of body apparently slightly thickened, into narrow (0.2–0.25 cm wide) ‘marginal’ frame, best visible on counterpart (Fig. 4B). Body consisting of numerous, small, polygonal, smooth, poorly distinct elements (plates). Next to appendage insertion, more or less circular structure consisting of a ~1.0 cm wide ring, enclosing few, large plates (Fig. 4A). At opposite, narrowest extremity of body (apex), short segmented(?) extension (~1.0 cm long, 0.3 cm wide) protruding next to appendage (Fig. 4A–B).
26Appendage inserted in poorly distinct, segmented(?) area at largest body extremity (opposite the apex), and forming long, slender structure extending along the entire length of the body, and ~3.0 cm beyond the apex (Fig. 4A–B). Appendage distally curved, with shorter (~3.0 cm long), straight, distal-most portion forming an angle of ~45° with long (~5.0 cm), straight to sinuous, more proximal part. Proximal-most portion of appendage very short, straight, possibly broken. Appendage consisting of numerous elongate segments, some of them, apparently slightly displaced. Distinct longitudinal groove along the whole length of appendage (Fig. 4A–B).
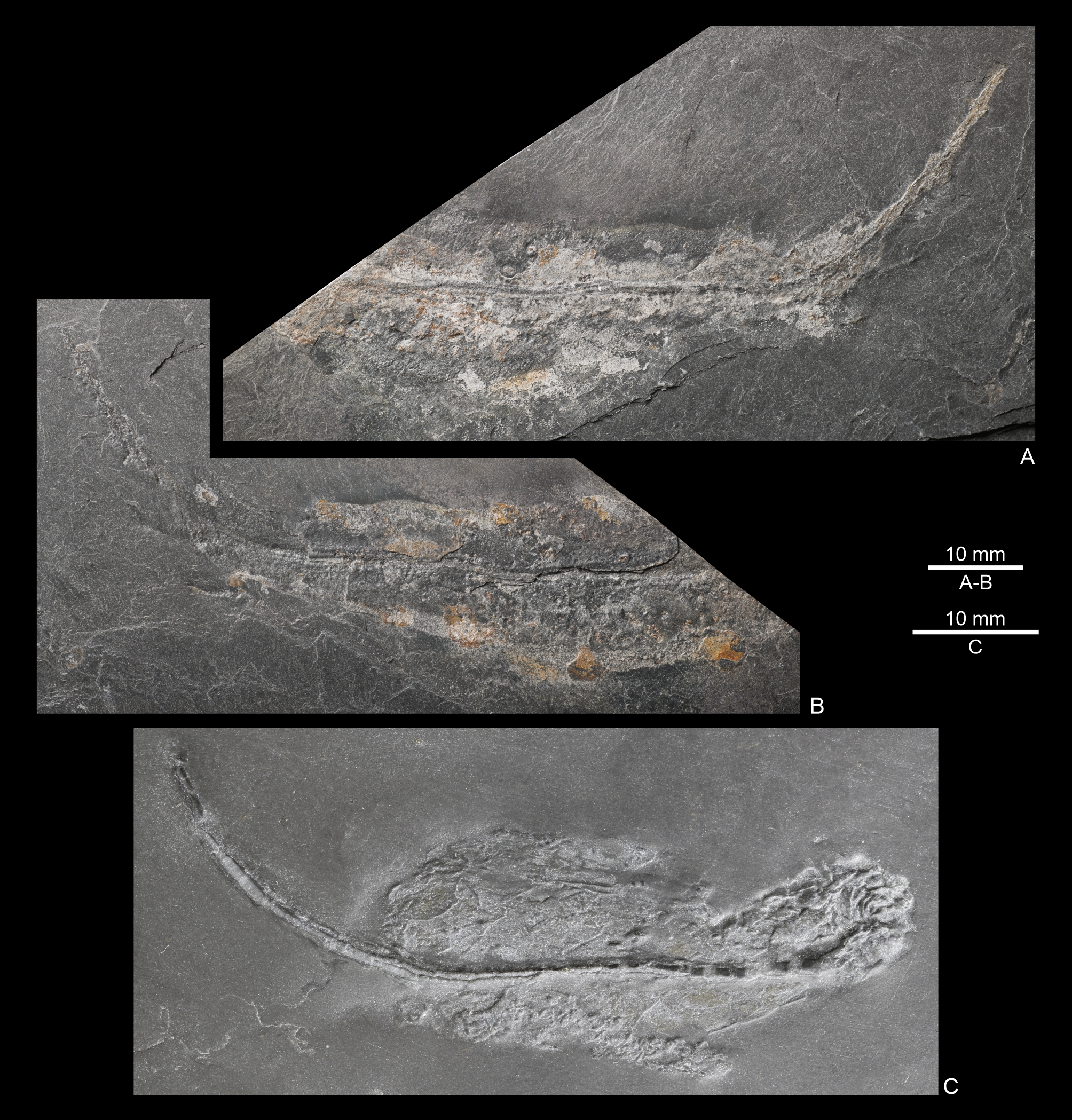
Figure 4. A–B. Dehmicystis? sp., RBINS a14020, part and counterpart; Warmifontaine, spoil heap of the Sainte-Barbe Quarry, La Roche Formation (Martelange Member), Pragian. C. Dehmicystis globulus (Dehm, 1934), NHMMZ PWL 2000/90-LS; pit near Bundenbach (Germany), Kaub Formation (Emsian).
27Remarks. The identification of specimen RBINS a14020 is particularly difficult, because of the strong flattening and tectonic distortion affecting it. However, the preservation of several morphological features, such as a bipartite organisation, with a ‘body’ composed of numerous plates and a very long and narrow appendage consisting of articulated segments, makes it possible to rule out its assignment to most metazoan phyla, with the exception of echinoderms. Within this phylum, affinities with most Devonian groups can be excluded (e.g. asterozoans, blastoids, crinoids, echinoids, edrioasteroids, holothurians, ophiocistioids) because of major differences in general organisation. The morphology of RBINS a14020 is only compatible with a limited number of echinoderm clades: pleurocystitids, stylophorans, and solutans.
28RBINS a14020 shares with pleurocystitids the possession of a bipartite body, consisting of a multi-element stem inserted into a flattened, elongate, nearly bilaterally symmetrical theca. However, a detailed comparison with the morphology of Devonian pleurocystitids (e.g. Henicocystis, Regulaecystis, Turgidacystis) makes it possible to identify several important differences. For example, the stem of pleurocystitids is made of numerous, relatively short, ring-shaped elements (columnals), forming a wide, regularly tapering structure (Dehm, 1932, 1934; Jell, 1983; Parsley & Sumrall, 2007). In marked contrast, the appendage of RBINS a14020 is a narrow, elongate structure, remaining of the same width all along its length, and made of one (possibly two, if the longitudinal groove corresponds to plate sutures) series of elongate elements (Fig. 4A–B). In Devonian pleurocystitids, the upper thecal surface consists of a limited number of stout, usually ornamented (with strong ridges) plates, while the opposite (lower) thecal side comprises a large, polyplated anal area (Dehm, 1932, 1934; Jell, 1983; Parsley & Sumrall, 2007). The theca of RBINS a14020 does not show any evidence of contrasted plate patterns between its two opposite faces, both apparently made exclusively of small, polygonal, non-ornamented elements (Fig. 4A–B).
29As is the case in RBINS a14020, the long feeding appendage of mitrate stylophorans (aulacophore) is frequently preserved in recurved position over the body (theca). In mitrates, this preferential taphonomic posture is generally interpreted as resulting from a post-mortem contraction (Ruta & Bartels, 1998; Lefebvre, 2003; Lefebvre & Ausich, 2021). In such preserved mitrates, portions of the particularly long distal aulacophore can also extend far beyond the posterior extremity of the theca (see e.g. Ubaghs, 1979). However, distal portions of mitrate aulacophores are not bent (or hooked), as this is the case in RBINS a14020. Another intriguing aspect of this specimen is that its appendage is visible all along its length in both the part and the counterpart, i.e. even when it extends below the theca (Fig. 4A–B). This implies that the body wall was made of very thin elements. This interpretation is in good accordance with the occurrence of thickened lateral edges (Fig. 4B). The apparent marginal frame is probably taphonomic in origin, and results from the flattening of a globose, polyplated theca made of numerous small plates (see comparable examples in e.g. Dupichaud et al., 2023a; Zamora & Gutiérrez-Marco, 2023). Such a body organisation departs from the situation in Devonian mitrates, all characterised by a theca made of a limited number of thick plates (see e.g. Ruta & Bartels, 1998; Ruta, 1999; Lefebvre & Ausich, 2021). There is no known example of a mitrate stylophoran with its recurved aulacophore leaving an imprint on the opposite (lower) surface of the theca (see e.g. Dehm, 1932, 1934; Gill & Caster, 1960; Jefferies & Lewis, 1978; Ubaghs, 1979; Ruta & Bartels, 1998; Ruta & Jell, 1999a, b; Lefebvre & Ausich, 2021).
30Devonian solutans share with RBINS a14020 the possession of a very long, stem-like appendage (homoiostele), inserted in a more or less globose theca made of numerous, unorganised plates (see e.g. Dehm, 1934; Gill & Caster, 1960; Caster, 1967; Parsley & Sumrall, 2007; Rahman & Lintz, 2012; Zamora & Gutiérrez-Marco, 2023). Similarly to the situation in RBINS a14020, solutans can be preserved with their homoiostele recurved over the theca (Fig. 4C; see also individual 3 in Rahman & Lintz, 2012, figs 4, 7). In solutans, the homoiostele is made of two parts: a long and narrow distal portion (dististele), and a wider, shorter proximal region, made of telescopic rings (proxistele) (Caster, 1967; Ubaghs, 1981; Noailles et al., 2014). This organisation is compatible with the situation in RBINS a14020, with a narrow, elongate appendage corresponding to the dististele, and the wider, segmented region at its proximal extremity, to the proxistele (Fig. 4A). Comparison with the solutan homoiostele also suggests that the longitudinal groove running over the length of the appendage in the Warmifontaine specimen could correspond to the suture between the two sets of upper and lower series of ossicles forming the dististele (Fig. 4A–B). Moreover, as in RBINS a14020, the distal extremity of the homoiostele is frequently bent or hooked in solutans (see e.g. Rozhnov & Jefferies, 1996; Noailles et al., 2014; Dupichaud et al., 2023b). If the interpretation of RBINS a14020 as a solutan is correct, the short, segmented (?) structure protruding next to its dististele, beyond the apex of the theca, could then very likely correspond to the poorly preserved distal extremity of its unique brachiole (Fig. 4A–B). Finally, if this interpretation is correct, the large, rounded structure near the homoiostele insertion of RBINS a14020 could then represent the periproct, and the large associated skeletal elements, ‘sugar loaf’ anal plates. In solutans, large suranal plates are an apomorphy of the family Dendrocystitidae (Noailles et al., 2014). Consequently, although poorly preserved, strongly flattened and distorted, all available morphological features observed in RBINS a14020 are compatible with its assignment to dendrocystitid solutans.
31Comparison of RBINS a14020 with the three Siluro–Devonian solutan genera known so far suggests closer affinities with Dehmicystis and Rutroclypeus, both characterised by large rounded thecae made of numerous small plates (Dehm, 1934; Gill & Caster, 1960; Caster, 1967; Rahman & Lintz, 2012), than with Claritacarpus, possessing a more quadrangular theca, consisting of few, large elements (Parsley & Sumrall, 2007). Rutroclypeus differs from both Dehmicystis and the Warmifontaine solutan by the possession of ornamented thecal plates (all of them bearing a small central tubercle) and the more lateral position of its periproct, at the posterior extremity of the anal lobe (Gill & Caster, 1960; Jell & Holloway, 1982). Consequently, RBINS a14020 is tentatively assigned to Dehmicystis. Its morphology is too poorly known and tectonically distorted to suggest an identification at species level.
32Occurrence. Warmifontaine, disused underground Sainte-Barbe Quarry; La Roche Formation (Martelange Member), Pragian (Fig. 2).
33Class Stylophora Gill & Caster, 1960
34Order Mitrata Jaekel, 1918
35Suborder Peltocystida Jefferies, 1973
36Remarks. Peltocystidan mitrates are the most long-ranging clade of stylophorans, with representatives known at least from the Tremadocian (Early Ordovician) to the Bashkirian (Early Pennsylvanian). They are characterised by the possession of a single posterior spine (glossal) and two particularly large adorals on the upper thecal surface. Although peltocystidans were highly diverse and palaeogeographically widespread during the Ordovician, their younger record is particularly cryptic (Lefebvre, 2007; Lefebvre & Ausich, 2021). They are so far undocumented in the Silurian, and are known from a handful of later occurrences: in the Lower and Middle Devonian of Germany (Haude, 1983; Südkamp, 2017) and the Carboniferous (Pennsylvanian) of Oklahoma (Kolata et al., 1991; Domínguez et al., 2002). Another occurrence of post-Ordovician peltocystidans is reported here from the Lower Devonian (Pragian) of southern Belgium.
37The stratigraphic range of the family Kirkocystidae was recently tentatively extended to the Early Devonian by Südkamp (2017), so as to include all specimens from the Hunsrück Slate (lower Emsian) of Rhineland-Palatinate (Germany) regularly mentioned as Mitrocystites? styloideus (see e.g. Dehm, 1934; Bartels & Brassel, 1990; Bartels et al., 1998; Rahman & Lintz, 2012), Dehmicystis globulus (Bartels & Brassel, 1990, fig. 162; Bartels et al., 1998, fig. 139), or homalozoans indet. (Bartels et al., 1998, fig. 140). The family Kirkocystidae traditionally comprises the two Ordovician genera Anatifopsis and Balanocystites (Lefebvre, 1999; Lee et al., 2004, 2006). Both taxa are characterised by the possession of two particularly extensive, subequal adorals, forming together the whole upper thecal surface and the lateral edges of the lower side. The lower thecal surface comprises five marginals and, posteriorly, a relatively wide left infracentral area (Jefferies, 1981; Lefebvre, 1999, 2001; Lee et al., 2004, 2006). Marginals are more or less overlapped by adorals, some of them, completely. The anal opening is slit-like and located along the posterior margin of a wide infracentral element modified into a rounded anal plate. A single elongate spine (glossal) is articulated to the posterior extremity of the marginal frame, right of the anal opening. The aulacophore of kirkocystids is characterised by particularly spinose stylocone and ossicles.
38Within peltocystidan mitrates, kirkocystids differ from peltocystids mainly by the possession of more extensive adorals, a single infracentral area (instead of two), and of a well-defined rounded anal plate on the lower thecal surface (instead of a transverse slit-like orifice located within the supracentral area) (Lefebvre, 1999, 2001; Lee et al., 2006; Lefebvre & Botting, 2007). Jaekelocarpids differ from kirkocystids mainly by the loss of the left infracentral area, and the modification of the anal plate into a large element inserted medially in between the posterior extremities of two less extensive adorals, restricted to the upper thecal surface (Kolata et al., 1991; Domínguez et al., 2002).
39Südkamp (2017) considered that Hunsrück peltocystidans show all apomorphies of the family Kirkocystidae (e.g. large adorals extending to and overlapping part of the lower thecal surface, single posterior spine, rounded anal plate, large left infracentral area, spinose aulacophore) and, therefore, he assigned them to the Ordovician genus Anatifopsis. However, the detailed morphology of Mitrocystites? styloideus and slightly younger peltocystidans from the Middle Devonian of Rhenish Massif (Germany) figured by Haude (1983, fig. 3) remains imperfectly known. In particular, the precise extension of their adorals and exact number of thecal plates are not documented so far. Moreover, Devonian peltocystidans from Germany seem to be characterised by an unusually ornamented stylocone bearing a strongly spinose transverse crest on its lower surface (Dehm, 1934; Haude, 1983). Therefore, their precise taxonomic assignment to any peltocystidan family (Jaekelocarpidae or Kirkocystidae) would require a detailed investigation of their morphology, which is beyond the scope of this paper.
40Peltocystidan indet.
41(Figs 5, 6)
42Material. RBINS a14021, part and counterpart.
43Description. Flattened fossil consisting of two distinct parts (Figs 5, 6): broad heart-shaped body, almost as long as wide (~2.4 cm) and, articulated to it, elongate, straight appendage (~3.8 cm long and 0.4–0.5 cm wide).
44Pentagonal, slightly asymmetrical body outline, with deep, median V-shaped concavity along ‘proximal’ margin at appendage insertion (Fig. 5). Lateral margins unequal in length: one nearly straight, the other longer and strongly bent. Lateral sides converging to relatively short, truncated, oblique distal extremity. Very faint, gently curved longitudinal median line extending from distal tip of body to opposite extremity (at appendage insertion), and dividing body into two subequal halves (Fig. 5). Narrow, slightly thickened U-shaped ‘collerette’ framing more or less regularly curved lateral and distal body margins, and distally delimiting faint, more or less rounded area (Fig. 6A). Two other lines, subequal in length, extending from median part of each lateral side, converging proximally, and forming together faint V-shaped structure directed towards appendage insertion (Fig. 6). No other structure or limit clearly visible on body.
45Proximal-most portion of appendage relatively short and wide, inserted into deep, concave body cavity (Figs 5, 6). Distal-most portion very long, almost straight, composed of numerous (at least 20) well-defined segments, all of relatively comparable width, but decreasing in length distally (Fig. 6B). Each segment made of well-defined, massive central element and, at least in distal part of appendage, two smaller V-shaped plates (one of the left, one of the right) articulated to lateral margins of each central element (Fig. 6B–C).

Figure 5. A–B. Peltocystidan indet., RBINS a14021, part and counterpart; Warmifontaine, spoil heap of the Sainte-Barbe Quarry, La Roche Formation (Martelange Member), Pragian.
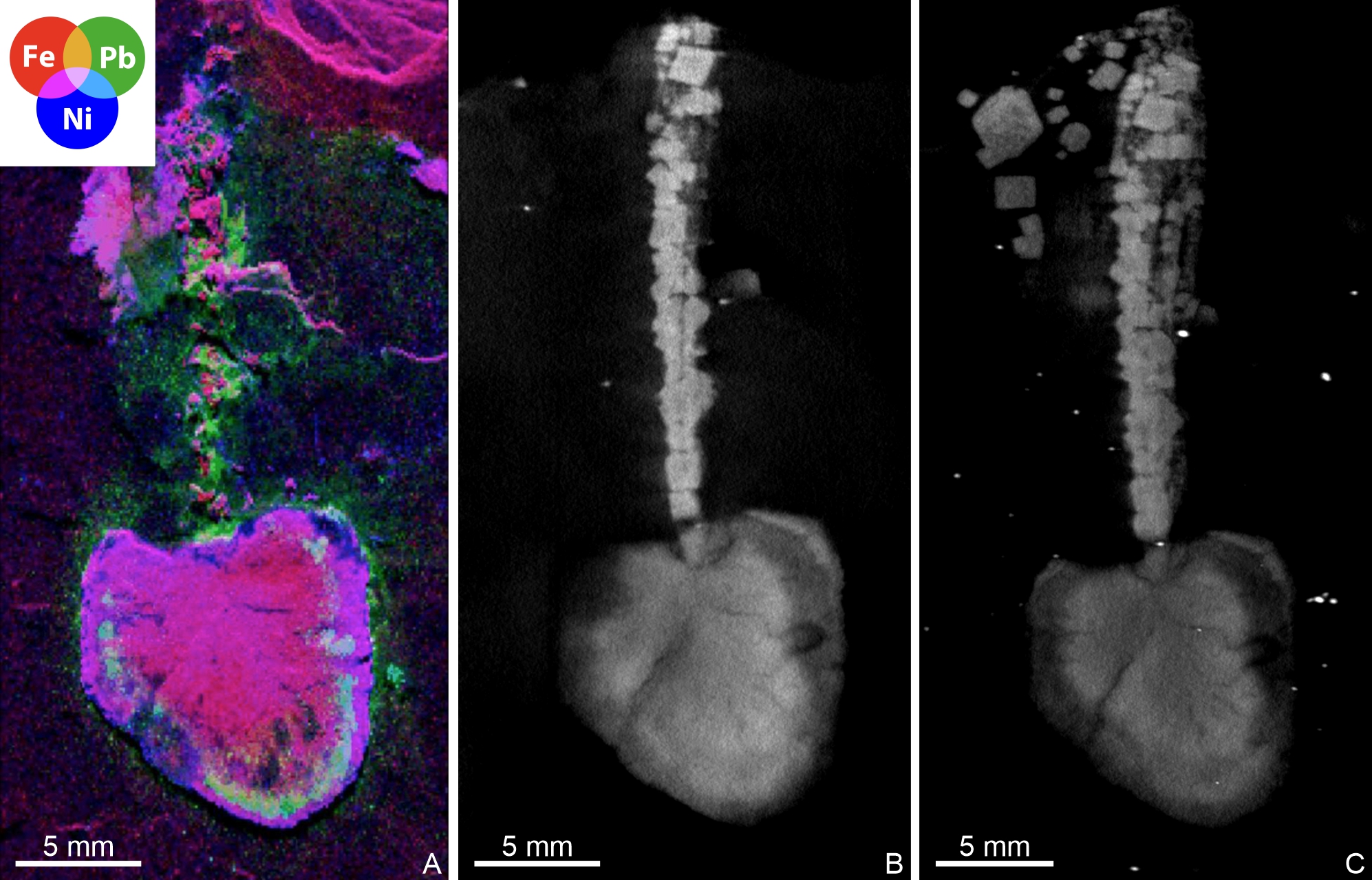
Figure 6. A–C. Peltocystidan indet., RBINS a14021; Warmifontaine, spoil heap of the Sainte-Barbe Quarry, La Roche Formation (Martelange Member), Pragian. A. μXRF overlay of iron (red), lead (green) and nickel (blue) distributions. B. Tomogram parallel to the slate surface. C. Maximum intensity projection of 46 tomograms parallel to the slate surface.
46Remarks. Specimen RBINS a14021 is better preserved and less tectonically distorted than RBINS a14020 (Dehmicystis? sp., see above). Its identification is therefore more straightforward. The bipartite organisation of this specimen, with a long, delicate, segmented appendage inserted into a more massive, asymmetrical and flattened body strongly supports echinoderm affinities and, more precisely, its assignment to either pleurocystitids, solutans, or stylophorans. In the distal part of the appendage, the occurrence of two series of small V-shaped plates articulated on both sides (left and right) of large central ossicles is incompatible with the identification of this appendage as a stem-like structure (pleurocystitid stem, solutan homoiostele). Stem-like appendages are cylindrical, closed structures made of tightly sutured elements (David et al., 2000; Lefebvre et al., 2019). On the other hand, the organisation of the distal appendage of RBINS a14021 is typical of ambulacral structures in echinoderms and, more precisely, the stylophoran aulacophore (Lefebvre et al., 2019, 2022). In this context, the large central ossicles can be identified as uniserial ossicles, and the smaller articulated elements as left and right series of ambulacral cover plates. This identification makes it possible to identify which views of RBINS a14021 are exposed in lower (Figs 5A, 6) and upper aspects (Fig. 5B).
47Within stylophorans, only three clades of mitrates survived the LOME and eventually diversified in the Early Devonian: anomalocystitids, paranacystids, and peltocystidans (Lefebvre, 2007; Lefebvre & Ausich, 2021). Devonian anomalocystitids and paranacystids share the same basic (mitrocystitidan) organisation for their lower thecal surface, with two central elements surrounded by several, relatively large marginal plates (Lefebvre, 1999, 2001). The lower thecal side of RBINS a14021 does not show any evidence of such a plate pattern. In contrast, it is very likely that the faint longitudinal line extending from the distal extremity of the theca to the aulacophore insertion corresponds to the imprint of the suture between two very large adorals (on the opposite, upper surface), therefore clearly suggesting peltocystidan affinities. This identification is supported by the pentagonal thecal outlines of RBINS a14021, identical to those of Devonian peltocystidans from the Hunsrück Slate (see e.g. Bartels et al., 1998, fig. 139). At the distal extremity of the lower thecal surface, the occurrence of a rounded element, morphologically identical to the anal plate of Mitrocystites? styloideus, further supports the assignment of RBINS a14021 to this family. Comparison with other peltocystidans suggests that the faint V-shaped structure on the lower thecal surface of the Warmifontaine specimen very likely corresponds to the strong, U- to V-shaped ridge occurring in some taxa (e.g. Anatifopsis trapeziiformis, Jaekelocarpus oklahomaensis; see Haude, 1983; Kolata et al., 1991; Lefebvre, 1999; Lee et al., 2004). No posterior spine (glossal) is present in RBINS a14021, but this plate is seldom preserved in Devonian peltocystidans (see e.g. Dehm, 1934; Haude, 1983; Bartels & Brassel, 1990; Bartels et al., 1998; but see Südkamp, 2017). Consequently, all available morphological features displayed by RBINS a14021 support its identification as a stylophoran (aulacophore with two series of cover plates articulated to uniserial ossicles) and more precisely as a peltocystidan mitrate (large adorals, rounded anal plate). The morphology of the Warmifontaine specimen seems to be very close to that of Mitrocystites? styloideus from the Hunsrück Slate, which was tentatively placed within the genus Anatifopsis by Südkamp (2017). However, a precise taxonomic assignment of the Warmifontaine mitrate remains difficult in the absence of a proper (re)description of the more abundant and better preserved peltocystidans from the Devonian of Germany, including Mitrocystites? styloideus.
48RBINS a14021 is preserved with its aulacophore in extended position (Figs 5, 6). This posture is unusual in mitrates: they are generally preserved with their aulacophore flexed and recurved above the upper thecal surface (Parsley, 1991; Ruta & Bartels, 1998; Lefebvre, 2003; Lefebvre & Ausich, 2021). This posture is interpreted as resulting from the post-mortem contraction of the appendage (Dehm, 1932; Ruta & Bartels, 1998; Lefebvre, 2003; Lefebvre & Ausich, 2021; Lefebvre et al., 2022; Saleh et al., 2023). The preservation of the aulacophore in extended position, generally interpreted as corresponding to its life (feeding) posture, implies that the organism was very likely buried alive by a sudden influx of sediments (Parsley & Gutiérrez-Marco, 2005; Lefebvre & Botting, 2007; Lefebvre & Ausich, 2021; Lefebvre et al., 2022; Saleh et al., 2023).
49Occurrence. Warmifontaine, disused underground Sainte-Barbe Quarry; La Roche Formation (Martelange Member), Pragian.
5. Discussion
50Uppermost Silurian (Pridoli) and Lower Devonian invertebrate faunas from the Ardenne Allochthon are strongly similar to those from the contemporaneous siliciclastic succession of the nearby Rhenish Massif (Germany), but it has to be said that palaeontologists essentially paid attention to the brachiopods during the last decades (see references in Mottequin, 2019 and Mottequin & Jansen, 2025). These benthic organisms proliferated in the ‘Rhenish’ (rhenotypic) facies (e.g. Erben, 1962; Jansen, 2016), which would represent shallow-marine, nearshore palaeoenvironments with turbulent, turbid water and essentially coarse siliciclastic sedimentation in contrast with the ‘Hercynian’ (hercynotypic) or ‘Bohemian’ facies that chiefly includes pelagic faunas reflecting offshore palaeoenvironments with calm water and predominantly argillaceous and calcareous sedimentation (Jansen, 2016).
51Our knowledge of echinoderms from the rhenotypic facies of southern Belgium is restricted essentially to the crinoids that can be abundant as columnals or stem fragments in some levels of the thick, essentially siliciclastic Lochkovian (~ Gedinnian), Pragian (~ Siegenian), and Emsian successions (e.g. Leriche, 1912; Asselberghs, 1930, 1946; Maillieux, 1931, 1933, 1938, 1940, 1941). Nevertheless, they remain inadequately known notably due to the rarity of well-preserved specimens (Figs 7, 8A–C). Indeed, calyxes are particularly scarce as reflected by the few discoveries reported in the literature (Schmidt in Maillieux, 1936; Maillieux, 1938 [note that Platycrinus lommanus Maillieux, 1938 (Fig. 8C) could be late Emsian or early Eifelian in age]; Ubaghs, 1945a, 1945b, 1947; Closset, 1970). It is obvious that the taxonomic revision of the Early Devonian crinoids from Belgium is needed to better assess their diversity and their close relationships with those from the contemporaneous successions of the nearby Rhenish Massif. This is notably the case for those from the Longlier Formation (Pragian), which were identified by Schmidt (in Maillieux, 1936) (Fig. 7A), who noted strong similarities with the German species, but they mostly remain undescribed. Pyritised crinoids are also known from the Herbeumont slate underground quarries west of Warmifontaine (RBINS collections). Apart from crinoids, reports of other echinoderms are scarce in the literature. A single blastoid (Pentremitidea) occurrence is known from the upper Emsian–lower Eifelian Moulin de la Foulerie Formation of the southern margin of the Dinant Synclinorium (Fraipont, 1884; Etheridge & Carpenter, 1886) (Fig. 8D–I). Asterozoans (asteroids and ophiuroids) can be counted on the fingers of the two hands (Leriche, 1912; Asselberghs, 1930; Ubaghs, 1942, 1943) (Fig. 8J–K). To this point, as is the case for the aforementioned pyritised crinoids, large asteroids are known from the Linglé slate underground quarry (Fig. 1) at Mortehan (RBINS collections) (Fig. 8K). A single specimen (holotype) of the cyclocystoid Sievertsia concava Smith & Paul, 1982 (Fig. 8L–M), first reported by Maillieux (1926) and illustrated by Sieverts-Doreck (1951), is known from the upper Emsian Hierges Formation. Consequently, the first reports of Dehmicystis? sp. and a peltocystidan mitrate in the Belgian Lower Devonian deserved to be thoroughly documented.
52The identification of the representatives of the two echinoderm classes Soluta (Dehmicystis? sp.) and Stylophora (peltocystidan indet.) significantly increases the taxonomic diversity of invertebrate marine assemblages from the Martelange Member (La Roche Formation), which was up to now considered as yielding poorly diverse invertebrate fossils (e.g. Asselberghs, 1946). The two echinoderms identified in Warmifontaine suggest strong affinities with slightly younger taxa from the Hunsrück Slate of Germany (early Emsian): Dehmicystis globulus and Mitrocystites? styloideus. Faunal affinities between these two regions, palaeogeographically close to each other in the Early Devonian, are not surprising and were already suggested (see references in Mottequin & Jansen, 2025). However, as far as solutans and stylophorans are concerned, palaeobiogeographic implications remain limited, because of the scarcity of other occurrences worldwide. Similarities with Devonian taxa from Germany may also simply reflect the existence of relatively similar environmental and/or taphonomic conditions, associated with relatively deep settings.

Figure 7. Crinoids from the Lower Devonian of Belgium; both are negatives of pictures of the external moulds that were coated with ammonium chloride sublimate. See Denayer & Mottequin (2024) for the lithostratigraphic units. A. Diamenocrinus stellatus Jaekel, 1895 (fide Schmidt in Maillieux, 1936), RBINS a1057 (= specimen D1 in Ubaghs, 1947); Martelange (Fauvillers 7a), Longlier Formation, Pragian. B. Ctenocrinus pyramidalis Ubaghs, 1945a, RBINS a7679 (holotype); south of Petigny, Hierges Formation, upper Emsian.

Figure 8. Crinoids (A–C), blastoid (D–I), ophiuroids (J–K), and cyclocystoid (L–M) from the Lower Devonian of Belgium (except otherwise stated). See Denayer & Mottequin (2024) for the lithostratigraphic units. All specimens are coated with ammonium chloride sublimate (except otherwise stated). A. Ctenocrinus aculeatus Ubaghs, 1945a, RBINS a7674 (holotype), artificial cast; railway trench between Grupont and Mirwart, Ruisseau de la Forge Formation (Pèrnelle Member), upper Pragian. B. Ctenocrinus arduennensis Closset, 1970, RBINS a172 (holotype), artificial cast; railway trench between Grupont and Mirwart, Ruisseau de la Forge Formation (Pèrnelle Member), upper Pragian. C. Platycrinus lommanus Maillieux, 1938, RBINS (holotype), artificial cast; Rochefort 31(2), Moulin de la Foulerie Formation (Eau Noire Member), upper Emsian or lower Eifelian (see fig. 2 in Denayer et al., 2024). D–I. Pentremitidea fraiponti (‘Fraipont, 1884’), PA.ULg.20250109-1 (holotype by monotypy), calyx in lateral (D–G), aboral (H) and oral (I) views (SEM); Champlon-Famenne, Moulin de la Foulerie Formation (Saint-Joseph Member or Eau Noire Member), upper Emsian or lower Eifelian (see fig. 2 in Denayer et al., 2024). J. Bohemura constellata (Thorent, 1839), RBINS a7666, artificial cast; Brûly-de-Pesche, Mondrepuis Formation, Lochkovian. K. Unidentified pyritised ophiuroid, RBINS a14162; Linglé slate underground quarry (Mortehan), La Roche Formation, Pragian. L–M. Sievertsia concava Smith & Paul, 1982, RBINS a9588 (holotype), external mould and artificial cast; Rochefort, 8649, Hierges Formation, upper Emsian.
Acknowledgements
53This paper is part of the project B2/202/P1/VERTIGO of the Belgian Science Policy Office (Belspo). We thank Herman Borré for authorising the collection of samples on his property, Guy Conard the neighbour of the quarry who provided assistance, our numerous colleagues for taking part in the field work, Laetitita Despontin (RBINS) for the SEM photographic work, and Thierry Hubin (RBINS) for photographing the echinoderm, mollusc and trilobite specimens from Warmifontaine and Bertrix. We are grateful to Manuela Aiglstorfer, David Kuhlmann and Kai Nungesser (Naturhistorisches Museum Mainz), who warmly welcomed us in the collections in their care and provided us with photographs of specimens. We thank A. Folie (RBINS), J. Lalanne (RBINS) and V. Fischer (ULiège) for access to the collections under their care. We thank SOLEIL synchrotron for provision of beamtime (proposal 20220721). Object Research Systems (ORS; Montreal, Canada, https://www.theobjects.com/dragonfly/index.html) and GOM (https://www.gom.com/) are thanked for allowing the use of their Dragonfly ORS (Version 4.1.0.647 for Windows) and GOM Inspect softwares. We are grateful to M. Reich (Germany) and an anonymous reviewer for their comments and suggestions.
Author contribution
54All the authors contributed to the writing of the paper, but BL is responsible for the systematic description and interpretation.
Data availability
55All studied specimens are housed in an official repository ensuring their long-term safekeeping and availability to other researchers for future studies.
References
56Asselberghs, E., 1924. Ardoisières du Dévonien de l’Ardenne. Annales des Mines de Belgique, 25/4, 1037–1098.
57Asselberghs, E., 1930. Description des faunes marines du Gedinnien de l’Ardenne. Mémoires du Musée royal d’Histoire naturelle de Belgique, 41, 1–73.
58Asselberghs, E., 1946. L’Éodévonien de l’Ardenne et des régions voisines. Mémoires de l’Institut géologique de l’Université de Louvain, 14, 1–598.
59Bartels, C. & Brassel, G., 1990. Fossilien im Hunsrückschiefer. Dokumente des Meereslebens im Devon. Museum Idar-Oberstein, Idar-Oberstein, 232 p.
60Bartels, C., Briggs, D.E.G. & Brassel, G., 1998. The Fossils of the Hunsrück Slate: Marine Life in the Devonian. Cambridge University Press, Cambridge, 324 p.
61Bassler, R.S., 1938. Pelmatozoa Palaeozoica (Generum et Genotyporum Index et Bibliographia). In Quenstedt, W. (ed.), Fossilium Catalogus (I: Animalia), pars 83. Verlag für Naturwissenschaften, ’s-Gravenhage, 1–194.
62Belanger, I., Delaby, S., Delcambre, B., Ghysel, P., Hennebert, M., Laloux, M., Marion, J.-M., Mottequin, B. & Pingot, J.-L., 2012. Redéfinition des unités structurales du front varisque utilisées dans le cadre de la nouvelle Carte géologique de Wallonie (Belgique). Geologica Belgica, 15/3, 169–175.
63Brett, C.E. & Seilacher, A. 1991. Fossil Lagerstätten: a taphonomic consequence of event sedimentation. In Einsele, G., Ricken, W. & Seilacher, A. (eds), Cycles and Events in Stratigraphy. Springer, Berlin, 283–297.
64Bultynck, P., Coen-Aubert, M. & Godefroid, J., 2000. Summary of the state of correlation in the Devonian of the Ardennes (Belgium-NE France) resulting from the decisions of the SDS. In Bultynck, P. (ed.), Subcommission on Devonian Stratigraphy, Recognition of Devonian series and stage boundaries in geological areas. Courier Forschungsinstitut Senckenberg, 225, 91–114.
65Candela, Y. & Mottequin, B., 2022. Tremadocian and Floian (Ordovician) linguliformean brachiopods from the Stavelot–Venn Massif (Avalonia; Belgium and Germany). Geologica Belgica, 25/1-2, 1–15. https://doi.org/10.20341/gb.2021.007
66Caster, K.E., 1967. Homoiostelea. In Moore, R.C. (ed.), Treatise on Invertebrate Paleontology. Pt. S. Echinodermata 1(2). Geological Society of America and University of Kansas Press, Lawrence, S581–S627.
67Caster, K.E., 1983. A new Silurian carpoid echinoderm from Tasmania and a revision of the Allanicytidiidae. Alcheringa, 7, 321–335. https://doi.org/10.1080/03115518308619615
68Cincotta, A., Mottequin, B., Guériau, P. & Olive, S., 2022. Taphonomy of a unique assemblage of putative early chordates from the Lower Devonian of Belgium. The Palaeontological Association, 66th Annual Meeting, 18th–24th July 2022, University College Cork, Ireland. Program and abstracts, 63.
69Cincotta, A., Mottequin, B., Guériau, P. & Olive, S., 2023. A new Konservat-Lagerstätte with putative early chordate from the Lower Devonian of Belgium. 4th International Congress on Stratigraphy, Strati 2023, 11th–13th July 2023, Lille, France. Abstract book, 271.
70Closset, J., 1970. Ctenocrinus arduennensis (Crinoidea Camerata), espèce nouvelle du Dévonien inférieur de la Belgique. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 46/29, 1–17.
71David, B., Lefebvre, B., Mooi, R. & Parsley, R., 2000. Are homalozoans echinoderms? An answer from the extraxial-axial theory. Paleobiology, 26, 529–555. https://doi.org/10.1666/0094-8373(2000)026<0529:AHEAAF>2.0.CO;2
72de Béthune, P., 1954. Carte géologique de Belgique. 1/500 000. Atlas de Belgique, planche 8. In Fourmarier, P. (ed.), Prodrome d’une description géologique de la Belgique. Liège, Société géologique de Belgique, Annexe 1.
73Dehm, R., 1932. Cystoideen aus dem rheinischen Unterdevon. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilage-Band (B: Geologie und Paläontologie), 69, 63–93.
74Dehm, R., 1934. Untersuchungen an Cystoideen des rheinischen Unterdevons. Sitzungsberichte der Bayerischen Akademie der Wissenschaften zu München, Mathematisch-naturwissenschaftliche Abteilung, 1934/I, 19–43.
75Dejonghe, L. & Hance, L., 2001. Carte géologique de Wallonie : Champlon – La Roche-en-Ardenne 60/1-2. 1/25 000. Ministère de la Région wallonne, Direction générale des Ressources naturelles et de l’Environnement, Namur, with an explanatory booklet of 44 p.
76de Koninck, L.-G., 1869. Sur quelques échinodermes remarquables des terrains paléozoïques. Bulletin de l’Académie royale des Sciences, des Lettres et des Beaux-Arts de Belgique, 28, 544–552.
77Denayer, J. & Mottequin, B., 2024. Lower Devonian lithostratigraphy of Belgium. Geologica Belgica, 27/3-4, 115–154. https://doi.org/10.20341/gb.2024.004
78Denayer, J., Coen-Aubert, M., Marion, J.-M. & Mottequin, B., 2024. Middle Devonian lithostratigraphy of Belgium. Geologica Belgica, 27/3-4, 155–192. https://doi.org/10.20341/gb.2024.009
79Dewalque, G., 1891. Sur quelques fossiles des ardoises de Warmifontaine (Neufchâteau). Annales de la Société géologique de Belgique, 18, Bulletin, 61–62.
80Domínguez, P., Jacobson, A.G. & Jefferies, R.P.S., 2002. Paired gill slits in a fossil with a calcite skeleton. Nature, 417, 841–844. https://doi.org/10.1038/nature00805
81Dupichaud, C., Lefebvre, B., Milne, C.H., Mooi, R., Nohejlová, M., Roch, R., Saleh, F. & Zamora, S., 2023a. Solutan echinoderms from the Fezouata Shale Lagerstätte (Lower Ordovician, Morocco): diversity, exceptional preservation, and palaeoecological implications. Frontiers in Ecology and Evolution, 11, 1290063. https://doi.org/10.3389/fevo.2023.1290063
82Dupichaud, C., Lefebvre, B. & Nohejlová, M., 2023b. Solutan echinoderms from the Lower Ordovician of the Montagne Noire (France): new data and palaeobiogeographic implications. Estonian Journal of Earth Sciences, 72, 26–29. https://doi.org/10.3176/earth.2023.80
83Erben, H.K., 1962. Zur Analyse und Interpretation der rheinischen und herzynischen Magnafazies des Devons. In Erben, H.K. (ed.), Internationale Arbeitstagung über die Silur/Devon-Grenze und die Stratigraphie von Silur und Devon. Bonn-Bruxelles 1960, Symposiums Band 2. E. Schweizerbart, Stuttgart, 42–61.
84Etheridge, R. & Carpenter, P.H., 1886. Catalogue of the Blastoidea in the Geological Department of the British Museum (Natural History), with an account of the morphology and systematic position of the group, and a revision of the genera and species. Trustees of British Museum, London, 322 p. https://doi.org/10.5962/bhl.title.45858
85Fraipont, J., 1884. Recherches sur les crinoïdes du Famennien (Dévonien supérieur) de Belgique. III. Annales de la Société géologique de Belgique, 11, Mémoires, 105–118.
86Ghysel, P., 2022. Carte géologique de Wallonie : Herbeumont - Suxy 67/3-4. 1/25 000. Service public de Wallonie, Agriculture, Ressources naturelles et Environnement, Namur, with an explanatory booklet of 50 p.
87Ghysel, P., 2023. Carte géologique de Wallonie : Neufchâteau – Juseret 65/5-6. 1/25 000. Service public de Wallonie, Agriculture, Ressources naturelles et Environnement, Namur, with an explanatory booklet of 50 p.
88Gill, E.D. & Caster, K.E., 1960. Carpoid echinoderms from the Silurian and Devonian of Australia. Bulletins of American Paleontology, 41, 1–71.
89Guensburg, T.E., Mooi, R., & Mongiardino Koch, N., 2023. Crinoid calyx origin from stem radial echinoderms. Journal of Paleontology, 97/5, 1092–1115. https://doi.org/10.1017/jpa.2023.14
90Haude, R., 1983. Kaum bekannte und seltene Echinodermen aus dem Mitteldevon des rechtsrheinischen Schiefergebirges. Der Aufschluss, 34, 101–110.
91Haugh, B.N. & Bell, B.M., 1980. Classification schemes. Studies in Geology, Notes for a Short Course, 3, 94–105. https://doi.org/10.1017/S0271164800000129
92Jaekel, O., 1895. Beiträge zur Kenntniss der palaeozoischen Crinoiden Deutschlands. Palaeontologische Abhandlungen, Neue Folge, 3/1, 1–116.
93Jaekel, O., 1901. Ueber Carpoideen; eine neue Classe von Pelmatozoen. Zeitschrift der Deutschen Geologischen Gesellschaft, 52, 661–677.
94Jaekel, O., 1918. Phylogenie und System der Pelmatozoen. Paläontologische Zeitschrift, 3/1, 1–128. https://doi.org/10.1007/bf03190413
95Jansen, U., 2016. Brachiopod faunas, facies and biostratigraphy of the Pridolian to lower Eifelian succession in the Rhenish Massif (Rheinisches Schiefergebirge, Germany). Geological Society, London, Special Publications, 423, 45–122. https://doi.org/10.1144/sp423.11
96Jefferies, R.P.S., 1973. The Ordovician fossil Lagynocystis pyramidalis (Barrande) and the ancestry of amphioxus. Philosophical Transactions of the Royal Society (B: Biological Sciences), 265, 409–469. https://doi.org/10.1098/rstb.1973.0032
97Jefferies, R.P.S., 1981. In defence of the calcichordates. Zoological Journal of the Linnean Society, 73, 351–396. https://doi.org/10.1111/j.1096-3642.1981.tb01601.x
98Jefferies, R.P.S. & Lewis, D.N., 1978. The English Silurian fossil Placocystites forbesianus and the ancestry of the vertebrates. Philosophical Transactions of the Royal Society, (B: Biological Sciences), 282, 205–323. https://doi.org/10.1098/rstb.1978.0013
99Jell, P.A., 1983. Early Devonian echinoderms from Victoria (Rhombifera, Blastoidea and Ophiocistioidea). Memoirs of the Association of Australasian Palaeontologists, 1, 209–235.
100Jell, P.A. & Holloway, D., 1982. Anal structure in Rutroclypeus Withers, 1933. Alcheringa, 6, 42. https://doi.org/10.1080/03115518208565418
101Klein, J.T., 1778. Naturalis Dispositio Echinodermatum. Accesserunt Lucubratiuncula de Aculeis Echinorum Marinorum et Specilegium de Belemnitis. Edita et Descriptionibus Novisque Inventis et Synonymis Auctorum Auca a Nathanaele Godofredo Leske. Officina Gleditschiana, Lipsiae (Leipzig), 278 p.
102Kolata, D.R., Frest, T.J. & Mapes, R.H., 1991. The youngest carpoid: occurrence, affinities, and life mode of a Pennsylvanian (Morrowan) mitrate from Oklahoma. Journal of Paleontology, 65/5, 844–855. https://doi.org/10.1017/s0022336000037811
103Lee, S.B., Lefebvre, B. & Choi, D.K., 2004. Morphometric analysis of Tremadocian (earliest Ordovician) kirkocystid mitrates (Echinodermata, Stylophora) from the Taebaeksan Basin, Korea. Geobios, 37, 731–748. https://doi.org/10.1016/j.geobios.2003.05.012
104Lee, S.B., Lefebvre, B. & Choi, D.K., 2006. Tremadocian stylophoran echinoderms from the Taebaeksan Basin, Korea. Journal of Paleontology, 80, 1072–1086. https://doi.org/10.1666/0022-3360(2006)80[1072:TSEFTB]2.0.CO;2
105Lefebvre, B., 1999. Stylophores (Cornuta, Mitrata) : situation au sein du phylum des échinodermes et phylogenèse. Unpublished Ph.D. Thesis, Université Claude Bernard Lyon 1, Villeurbanne, 630 p.
106Lefebvre, B., 2001. A critical comment on ‘ankyroids’ (Echinodermata, Stylophora). Geobios, 34, 597–627. https://doi.org/10.1016/S0016-6995(01)80024-3
107Lefebvre, B., 2003. Functional morphology of stylophoran echinoderms. Palaeontology, 46, 511–555. https://doi.org/10.1111/1475-4983.00309
108Lefebvre, B., 2007. Early Palaeozoic palaeobiogeography and palaeoecology of stylophoran echinoderms. Palaeogeography, Palaeoclimatology, Palaeoecology, 245/1–2, 156–199. https://doi.org/10.1016/j.palaeo.2006.02.021
109Lefebvre, B., 2022. La diversification des échinodermes au Paléozoïque inférieur : l’apport des gisements à préservation exceptionnelle. Dédale Editions, Lyon, 337 p.
110Lefebvre, B. & Ausich, W.I., 2021. New Siluro-Devonian anomalocystitids (Echinodermata, Stylophora) from Bolivia and Canada, and a reevaluation of skeletal homologies in mitrates. Paleontological Journal, 55/9, 932–965. https://doi.org/10.1134/S0031030121090070
111Lefebvre, B. & Botting, J.P., 2007. First report of the mitrate Peltocystis cornuta Thoral (Echinodermata, Stylophora) in the Lower Ordovician of central Anti-Atlas (Morocco). Annales de Paléontologie, 93, 183–198. https://doi.org/10.1016/j.annpal.2007.06.003
112Lefebvre, B., Sumrall, C.D., Shroat-Lewis, R.A., Reich, M., Webster, G.D., Hunter, A.W., Nardin, E., Rozhnov, S.V., Guensburg, T.E., Touzeau, A., Noailles, F. & Sprinkle, J., 2013. Palaeobiogeography of Ordovician echinoderms. In Harper, D.A.T & Servais, T. (eds), Early Palaeozoic Biogeography and Palaeogeography. Geological Society, London, Memoirs, 38, 173–198. https://doi.org/10.1144/M38.14
113Lefebvre, B., Guensburg, T.E., Martin, E.L.O., Mooi, R., Nardin, E., Nohejlová, M., Saleh, F., Kouraïss, K., El Hariri, K. & David, B., 2019. Exceptionally preserved soft parts in fossils from the Lower Ordovician of Morocco clarify stylophoran affinities within basal deuterostomes. Geobios, 52, 27–36. https://doi.org/10.1016/j.geobios.2018.11.001
114Lefebvre, B., Nohejlová, M., Martin, E.L.O., Kašička, L., Zicha, O. & Gutiérrez-Marco, J.C., 2022. New Middle and Late Ordovician cornute stylophorans (Echinodermata) from Morocco and other peri-Gondwanan areas. In Hunter, A.W., Álvaro, J.J., Lefebvre, B., Van Roy, P. & Zamora, S. (eds), The Great Ordovician Biodiversification Event: Insights from the Tafilalt Biota, Morocco. The Geological Society, London, Special Publications, 485, 345–522. https://doi.org/10.1144/SP485-2021-99
115Leriche, M., 1912. La faune du Gedinnien inférieur de l’Ardenne. Mémoires du Musée royal d’Histoire naturelle de Belgique, 6, 1–58.
116Lindström, G., 1888. List of the Fossil Faunas of Sweden, Part 2, Upper Silurian. Norstedt & Söner, Stockholm, 29 p.
117Maillieux, E., 1926. Présence du genre “Cyclocystoides” Salter et Billings dans l’Emsien supérieur de l’Ardenne. Bulletin de la Société belge de Géologie, de Paléontologie et d’Hydrologie, 35/1 (pro 1925), Procès-verbaux, 93–95.
118Maillieux, E., 1931. La faune des grès et schistes de Solières (Siegenien moyen). Mémoires du Musée royal d’Histoire naturelle de Belgique, 51, 1–90.
119Maillieux, E., 1933. Terrains, roches et fossiles de la Belgique. Deuxième édition revue et corrigée. Patrimoine du Musée royal d’Histoire naturelle de Belgique, Bruxelles, 217 p.
120Maillieux, E., 1936. La faune et l’âge des Quartzophyllades siegeniens de Longlier. Mémoires du Musée royal d’Histoire naturelle de Belgique, 73, 1–140.
121Maillieux, E., 1938. Le Couvinien de l’Ardenne et ses faunes. Mémoires du Musée royal d’Histoire naturelle de Belgique, 83, 1–57.
122Maillieux, E., 1940. Le Siegenien de l’Ardenne et ses faunes. Bulletin du Musée royal d’Histoire naturelle de Belgique, 16/5, 1–23.
123Maillieux, E., 1941. Note sur la faune de quelques gisements fossilifères éodévoniens de l’Ardenne. Bulletin du Musée royal d’Histoire naturelle de Belgique, 17/9, 1–7.
124McIntosh, G.C. & Macurda, D.B., 1979. Devonian echinoderm biostratigraphy. Special Papers in Palaeontology, 23, 331–334.
125Mooi, R., Lefebvre, B., Guensburg, T.E., Nohejlová, M. & Dupichaud, C., 2024. Approaches to understanding echinoderm origins. Part 2: Questioning conceptual models. Cahiers de Biologie Marine, 65, 463–490.
126Mottequin, B., 2019. An annotated catalogue of types of Silurian–Devonian brachiopod species from southern Belgium and northern France in the Royal Belgian Institute of Natural Sciences (1870–1945), with notes on those curated in other Belgian and foreign institutions. Geologica Belgica, 22/1-2, 47–89. https://doi.org/10.20341/gb.2019.005
127Mottequin, B., 2021. Earth science collections of the Centre Grégoire Fournier (Maredsous) with comments on Middle Devonian–Carboniferous brachiopods and trilobites from southern Belgium. Geologica Belgica, 24/1-2, 33–68. https://doi.org/10.20341/gb.2020.028
128Mottequin, B. & Jansen, U., 2025. Revisiting the Silurian–Lower Devonian spiriferide and spiriferinide brachiopods from the Condroz Inlier and Ardenne Allochthon (Belgium): current data and perspectives. Palaeobiodiversity and Palaeoenvironments, 105, 229–264. https://doi.org/10.1007/s12549-025-00645-6
129Nardin, E., Lefebvre, B., Fatka, O., Nohejlová, M., Kašička, L., Šinágl, M. & Szabad, M., 2017. Evolutionary implications of a new transitional blastozoan echinoderm from the middle Cambrian of the Czech Republic. Journal of Paleontology, 91, 672–684. https://doi.org/10.1017/jpa.2016.157
130Noailles, F., Lefebvre, B. & Kašička, L., 2014. A probable case of heterochrony in the solutan Dendrocystites Barrande, 1887 (Echinodermata: Blastozoa) from the Upper Ordovician of the Prague Basin (Czech Republic) and a revision of the family Dendrocystitidae Bassler, 1938. Bulletin of Geosciences, 89, 451–476. https://doi.org/10.3140/bull.geosci.1475
131Nohejlová, M. & Lefebvre, B., 2022. Late Ordovician solutan echinoderms from the western Tafilalt, Morocco. In Hunter, A.W., Álvaro, J.J., Lefebvre, B., Van Roy, P. & Zamora, S. (eds), The Great Ordovician Biodiversification Event: Insights from the Tafilalt Biota, Morocco. The Geological Society, London, Special Publications, 485, 523–539. https://doi.org/10.1144/SP485-2021-55
132Olive, S., Guériau, P., Janvier, P. & Mottequin, B., 2022. Enigmatic early chordates from the Lower Devonian of Belgium. In Paredes-Aliaga, M.V., Manzanares, E., Mondéjar Fernández, J., Ros-Franch, S., Botella, H. & Martínez-Pérez, C. (eds), 16th International Symposium on Early and Lower Vertebrates, Valencia, Spain, 2022. Ichtyolith Issues, Special Publication, 15, 51.
133Parsley, R.L., 1972. The Belemnocystitidae: solutan homeomorphs of the Anomalocystitidae. Journal of Paleontology, 46, 341–347.
134Parsley, R.L., 1991. Review of selected North American mitrate stylophorans (Homalozoa: Echinodermata). Bulletins of American Paleontology, 100, 5–57.
135Parsley, R.L. & Gutiérrez-Marco, J.C., 2005. Stylophorans in middle Arenig shallow water siliciclastics: Vizcainocarpus from the Imfout syncline in Morocco’s western Meseta. Bulletin of Geosciences, 80, 185–192.
136Parsley, R.L. & Sumrall, C.D., 2007. New recumbent echinoderm genera from the Bois d’Arc Formation: Lower Devonian (Lochkovian) of Coal County, Oklahoma. Journal of Paleontology, 81/6, 1486–1493. https://doi.org/10.1666/04-072.1
137Philip, G.M., 1981. Notocarpos garratti gen. et sp. nov., a new Silurian mitrate carpoid from Victoria, Alcheringa, 3, 29–38. https://doi.org/10.1080/03115518108565431
138Rahman, I.A. & Lintz, H., 2012. Dehmicystis globulus, an enigmatic solute (Echinodermata) from the Lower Devonian Hunsrück Slate, Germany. Paläontologische Zeitschrift, 86, 59–70. https://doi.org/10.1007/s12542-011-0116-y
139Regnéll, G., 1945. Non-crinoid Pelmatozoa from the Paleozoic of Sweden. Meddelanden från Lunds Geologisk-Mineralogiska Institution, 108, 1–255.
140Regnéll, G., 1960. The Lower Palaeozoic echinoderm faunas of the British Isles and Balto-Scandia. Palaeontology, 2, 161–179.
141Reich, M., Lefebvre, B., Franzen, C. & Kutscher, M., 2007. Mitrates (Echinodermata: Stylophora) from the Silurian of Gotland, Sweden. In Elicki, O. (ed.), Fossile Ökosysteme: 77. Jahrestagung der Paläontologischen Gesellschaft. Kurzfasungen der Vorträge und Poster. Wissenschaftliche Mitteilungen des Institutes für Geologie der TU Bergakademie Freiberg, 36, 109.
142Rozhnov, S.V., 1990. New representatives of the class Stylophora (echinoderms). Paleontological Journal, 90/4, 27–38.
143Rozhnov, S.V. & Jefferies, R.P.S., 1996. A new stem-chordate solute from the Middle Ordovician of Estonia. Geobios, 29, 91–109. https://doi.org/10.1016/S0016-6995(96)80074-X
144Ruta, M., 1997. Redescription of the Australian mitrate Victoriacystis with comments on its functional morphology. Alcheringa, 21, 81–101. https://doi.org/10.1080/03115519708619177
145Ruta, M., 1999. A cladistic analysis of the anomalocystitid mitrates. Zoological Journal of the Linnean Society, 127, 345–421. https://doi.org/10.1111/j.1096-3642.1999.tb00681.x
146Ruta, M. & Bartels, C., 1998. A redescription of the anomalocystitid mitrate Rhenocystis latipedunculata from the Lower Devonian of Germany. Palaeontology, 41, 771–806.
147Ruta, M. & Jell, P.A., 1999a. Adoketocarpus gen. nov., a mitrate from the Ludlovian Kilmore Formation and Lochkovian Humevale Formation of central Victoria. Memoirs of the Queensland Museum, 43, 377–398.
148Ruta, M. & Jell, P.A., 1999b. Revision of Silurian and Devonian Allanicytidiidae (Anomalocystitida: Mitrata) from southeastern Australia, Tasmania and New Zealand. Memoirs of the Queensland Museum, 43, 431–451.
149Saleh, F., Lefebvre, B., Dupichaud, C., Martin, E.L.O., Nohejlová, M. & Spaccesi, L., 2023. Skeletal elements controlled soft-tissue preservation in echinoderms from the Early Ordovician Fezouata Biota. Geobios, 81, 51–66. https://doi.org/10.1016/j.geobios.2023.08.001
150Schöder, S., Müller, K., Tranchant, L., Rouquié, A., Gueriau, P., Thoury, M., Bérard, E., Okbinoglu, T., Berenguer, F., Iacconi, C., Robbiola, L., Moreno, T., Cohen, S.X. & Bertrand, L., 2024. Heritage research at the PUMA beamline. Applied Physics A, 130, 848. https://doi.org/10.1007/s00339-024-08026-0
151Shields, G., 1998. What are Lagerstätten? Lethaia, 31/2, 124. https://doi.org/10.1111/j.1502-3931.1998.tb00498.x
152Sieverts-Doreck, H., 1951. Über Cyclocystoides Salter & Billings und eine neue Art aus dem belgischen und rheinischen Devon. Senckenbergiana, 32, 9–30.
153Smith, A.B. & Paul, C.R.C, 1982. Revision of the Class Cyclocystoidea (Echinodermata). Philosophical Transactions of the Royal Society of London, (B: Biological Sciences), 296, 577–679. https://doi.org/10.1098/rstb.1982.0031
154Solé, V.A., Papillon, E., Cotte, M., Walter, P. & Susini, J., 2007. A multiplatform code for the analysis of energy-dispersive X-ray fluorescence spectra. Spectrochimica Acta Part B: Atomic Spectroscopy, 62/1, 63–68. https://doi.org/10.1016/j.sab.2006.12.002
155Sprinkle, J., 1973. Morphology and Evolution of Blastozoan Echinoderms. Museum of Comparative Zoology Harvard University, Cambridge (Mass.), 283 p. https://doi.org/10.5962/bhl.title.66379
156Sprinkle, J. & Guensburg, T.E., 2004. Crinozoan, blastozoan, echinozoan, asterozoan, and homalozoan echinoderms. In Webby, B.D., Paris, F., Droser, M.L. & Percival, I.G. (eds), The Great Ordovician Biodiversification Event. Columbia University Press, New York, 266–280. https://doi.org/10.7312/webb12678-027
157Stainier, P., 1994. LAR - Formation de La Roche. In Godefroid, J., Blieck, A., Bultynck, P., Dejonghe, L., Gerrienne, P., Hance, L., Meilliez, F., Stainier, P. & Steemans, P., Les formations du Dévonien inférieur du Massif de la Vesdre, de la Fenêtre de Theux et du Synclinorium de Dinant (Belgique, France). Mémoires pour servir à l’Explication des Cartes géologiques et minières de la Belgique, 38, 53–57.
158Südkamp, W. 2017. Leben im Devon. Bestimmungsbuch Hunsrückschieferfossilien. Pfeil, Munich, 176 p.
159Sutcliffe, O.E., Briggs, D.E.G. & Bartels, C., 1999. Ichnological evidence for the environmental setting of the Fossil-Lagerstätten in the Devonian Hunsrück Slate, Germany. Geology, 27/3, 275–278. https://doi.org/10.1130/0091-7613(1999)027%3C0275:ieftes%3E2.3.co;2
160Thorent, A., 1839. Mémoire sur la constitution géologique de la partie nord du département de l’Aisne, touchant au royaume de Belgique, et de l’extrémité sud du département du Nord. Mémoires de la Société géologique de France, 3/7, 239–260.
161Ubaghs, G., 1942. Bohemura constellata (Thorent) et Drepanaster sp., ophiuroïdes du Dévonien inférieur de la Belgique et du Nord de la France. Bulletin du Musée royal d’Histoire naturelle de Belgique, 18/7, 1–19.
162Ubaghs, G., 1943. Le genre Spaniaster Schöndorf dans le Dévonien inférieur de la Belgique. Bulletin du Musée royal d’Histoire naturelle de Belgique, 19/52, 1–4.
163Ubaghs, G. 1945a. Contribution à la connaissance des crinoïdes de l’Éodévonien de la Belgique. I. – Révision systématique des Melocrinitidae. Bulletin du Musée royal d’Histoire naturelle de Belgique, 21/15, 1–24.
164Ubaghs, G., 1945b. Contribution à la connaissance des crinoïdes de l’Éodévonien de la Belgique. II. – La morphologie des bras chez Ctenocrinus Bronn. Bulletin du Musée royal d’Histoire naturelle de Belgique, 21/16, 1–24.
165Ubaghs, G., 1947. Contribution à la connaissance des crinoïdes de l’Éodevonien de la Belgique. III. – L’appareil brachial d’Acanthocrinus Roemer et de Diamenocrinus Oehlert. Bulletin du Musée royal d’Histoire naturelle de Belgique, 23/4, 1–31.
166Ubaghs, G., 1979. Trois Mitrata (Echinodermata: Stylophora) nouveaux de l’Ordovicien de Tchécoslovaquie. Paläontologische Zeitschrift, 53, 98–119. https://doi.org/10.1007/BF02987791
167Ubaghs, G., 1981. Réflexions sur la nature et la fonction de l’appendice articulé des carpoïdes Stylophora (Echinodermata). Annales de Paléontologie, 67, 33–48.
168Watteyne, D., 2016. Les ardoisières de Warmifontaine. In Cattelain, P. (ed.), Au fil de l’ardoise… Au fil de l’eau. Quelques ardoisières ardennaises : Fumay, Haybes, L’Escaillère, Oignies-en-Thiérarche, Rimogne, Warmifontaine... DIRE, Treignes, 127–131.
169Zamora, S. & Gutiérrez-Marco, J.C., 2023. Filling the Silurian gap of solutan echinoderms with the description of new species of Dehmicystis from Spain. Acta Palaeontologica Polonica, 68/2, 185–192. https://doi.org/10.4202/app.01054.2023
170Manuscript received 04.03.2025, accepted in revised form 02.07.2025, available online 12.11.2025.
