


- Accueil
- Volume 17 (2013)
- numéro 3
- Impact des changements climatiques sur la répartition géographique des aires favorables à la culture et à la conservation des fruitiers sous-utilisés : cas du tamarinier au Bénin


Visualisation(s): 6356 (46 ULiège)
Téléchargement(s): 827 (1 ULiège)

Impact des changements climatiques sur la répartition géographique des aires favorables à la culture et à la conservation des fruitiers sous-utilisés : cas du tamarinier au Bénin

Notes de la rédaction
Reçu le 12 septembre 2012, accepté le 8 mai 2013
Résumé
L’intégration des fruitiers autochtones sous-utilisés dans les systèmes agroforestiers traditionnels, comme une des options de diversification de la production agricole en vue d’améliorer le revenu des ménages ruraux en Afrique, pourrait être compromise par les changements climatiques. La présente étude se propose d’analyser l’impact potentiel des changements climatiques sur la répartition géographique des habitats favorables au tamarinier (Tamarindus indica L.), une espèce agroforestière à fort potentiel économique mais sous-utilisée. Le principe d’entropie maximale (MAXENT) en combinaison avec un Système d’Information Géographique (SIG) a été utilisé pour modéliser les habitats favorables à la culture et à la conservation de l’espèce, sous les conditions climatiques actuelles et futures (horizon 2050). Les données de présence de l’espèce ont été collectées et combinées avec des données bioclimatiques dérivées de la base de données Worldclim et des données pédologiques. Trois modèles climatiques ont été utilisés pour les projections du futur (les modèles CCCMA, HadCM3 et CSIRO) sous le scénario A2 du GIEC (IPCC). Sous les conditions actuelles, 65 % du territoire et 87 % du réseau national d’aires protégées seraient très favorables respectivement à la culture et à la conservation des écotypes locaux de tamarinier. Par contre, une augmentation des précipitations (modèles CCCMA et HadCM3) pourrait convertir les zones actuellement très favorables à leur culture et à leur conservation (semi-aride et subhumide sèche) en zones peu favorables à l’horizon 2050. Une diminution des précipitations (modèle CSIRO) pourrait convertir les zones qui leur sont actuellement peu favorables (subhumides humides) en zones très favorables. Dans le schéma d’une aridification (CSIRO), la culture et la conservation du tamarinier seront possibles sur toute l’étendue du Bénin et/ou pourront nécessiter l’introduction d’écotypes originaires des zones plus arides. De telles études doivent être menées au fur et à mesure que les pouvoirs prédictifs des modèles climatiques seront améliorés, pour une plus grande pertinence des prises de décisions en vue d’une valorisation optimale des fruitiers autochtones sous-utilisés.
Abstract
Impact of climate change on the geographical distrubution of suitable areas for cultivation and conservation of underutilized fruit trees: case study of the tamarind tree in Benin. Climate change may limit integration of Underutilized Agroforestry Fruits Trees (UAFT) into formal cropping systems as a strategy to increase rural household income in Africa. The present study analyzed the potential impact of climate change on the geographical distribution of suitable areas for tamarind (Tamarindus indica L.), an economically important UAFT species. Presence records of the species were collected and combined with bioclimatic data derived from the Worldclim data base and from soil type data. The Maximum Entropy Modeling principle (MAXENT) was used in combination with a Geographic Information System (GIS) to forecast current and future (horizon 2050) suitable habitats for the cultivation and conservation of the species. Three different climate models were used for future predictions (the CCCMA, HadCM3 and CSIRO models) under IPCC scenario A2. Under current conditions, 65% of the national area and 87% of the national protected area network were found to be highly suitable for the cultivation and conservation of local tamarind ecotypes, respectively. It is possible that an increase in rainfall (CCCMA and HadCM3 models) will convert the currently highly suitable zones (semi-arid and subhumid dry) into poorly suitable areas at horizon 2050. A decline in precipitation (CSIRO model) could also convert the currently poorly suitable zones (sub-humid humid) into highly suitable zones. In the case of an aridification of the climate (CSIRO), cultivation and conservation of tamarind could become possible all over Benin and/or require the introduction of ecotypes from more arid areas. The predictive capacity of climatic models is constantly improving and it is therefore recommended that such studies be undertaken in order to better inform decision making for the optimum use of UAFT species.
Table des matières
1. Introduction
1Les changements climatiques sont aujourd’hui reconnus comme l'une des principales menaces pour la survie des espèces et l'intégrité des écosystèmes partout dans le monde. La connaissance des propriétés spécifiques de ces changements, susceptibles d'avoir un impact sur les espèces ou leurs habitats, constitue un élément central des stratégies d’adaptation (Heller et al., 2009). Ces changements du climat constituent une question environnementale qui mérite une attention particulière en matière de planification de la production agricole, de la diversification des productions agricoles et de la conservation des espèces. En effet, il est de plus en plus probable que les fluctuations des variables climatiques telles que les précipitations et la température auront une incidence sur la diversité biologique et sur la répartition géographique des habitats favorables aux espèces (IPCC, 2007).
2En Afrique, 25 à 42 % des espèces végétales pourraient être menacées d’extinction du fait d’une perte de 81 à 97 % des habitats favorables d’ici 2085 (Boko et al., 2007). D’après les projections, 20 à 30 % des espèces végétales et animales feront face à un plus grand risque d’extinction si le réchauffement global excède 1,5 °C à 2,5 °C en Afrique (IPCC, 2007 ; Busby et al., 2010).
3Les dernières décennies ont été témoins de l’incapacité des systèmes de production agricole africains (basés sur des espèces essentiellement exotiques) à améliorer durablement les conditions de vie des populations rurales. Pour remédier à cela, les initiatives futuristes suggéraient déjà à la fin du siècle échu, la domestication et la promotion des fruitiers agroforestiers autochtones sous-utilisés comme l’une des alternatives pouvant permettre d’atteindre le premier objectif du millénaire : éradiquer l’extrême pauvreté et la faim (Leakey et al., 1998). Dans cette optique, de grands efforts ont été consentis en termes de financement et de recherches scientifiques pour construire une grande base de données sur les espèces concernées (ex., www.worldagroforestry.org/resources/databases/agroforestree). Les données disponibles couvrent une gamme assez diversifiée de domaines (sylviculture, diversité génétique, qualité organoleptique, productivité, statut écologique, etc.). L’une des étapes clés importantes à franchir est l’intégration de ces espèces dans les politiques formelles de production agricole. Toutefois, en dépit des efforts de recherches scientifiques, très peu d’études à ce jour se sont intéressées à l’impact potentiel des changements climatiques sur la répartition géographique des habitats favorables à ces espèces. Or, ce type d’information s’avère important pour mieux raisonner les choix des zones où ces espèces peuvent être cultivées aujourd’hui et dans le futur. Ceci se justifie par le fait que les changements climatiques pourraient engendrer une dynamique spatiale de la distribution géographique des habitats favorables à ces espèces et rendre ainsi certaines régions aujourd’hui favorables, très peu favorables dans le futur et vice versa (Hannah et al., 2002). La négligence de cette dimension pourrait hypothéquer les efforts de domestication et avoir un impact dramatique sur les systèmes de production.
4De même, une grande partie de l’impressionnante biodiversité de l'Afrique est à ce jour conservée dans les aires protégées au moyen d'un très vaste système de différents types de bio-réserves. Bien que certaines études se soient récemment penchées sur l’efficacité des aires protégées à conserver certains de ces fruitiers autochtones dont Tamarindus indica L. (Fandohan et al., 2011a) et Sclerocarya birrea (A.Rich.) Hochst. (Gouwakinnou, 2011) en Afrique sub-saharienne, les investigations sur l’impact potentiel des changements climatiques sur l’efficacité des aires protégées à conserver les espèces autochtones demeurent assez rares. Or, les changements climatiques représentent une menace fondamentalement différente des autres menaces car jamais, par le passé, un stress écologique n’a remis en cause l’efficacité des réseaux d’aires protégées à conserver des échantillons représentatifs des espèces (Scott, 2005) ou à couvrir les habitats qui leurs sont favorables. En un mot, il y a de grandes incertitudes (à l’échelle des pays et des régions) sur comment ces aires statiques continueront à conserver les espèces concernées ou à leur offrir des habitats favorables, en raison des changements climatiques.
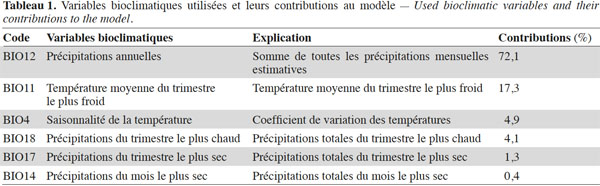
5Les changements climatiques constituent donc un vrai défi pour l’élaboration de politiques et options futures en agriculture et en conservation, du fait des incertitudes qui leurs sont associées : Que peut-on produire et où peut-on produire ? Que peut-on conserver et où peut-on conserver ? Ces différents questionnements encore sujets à de grands débats pourraient également introduire un décalage entre l'adoption des mesures d'adaptation et leur mise en œuvre, ce qui augmente le besoin de stratégies d'adaptation. Étant donné que les ressources naturelles au centre des stratégies d’adaptation (ex. les fruitiers autochtones sous-utilisés) pourraient être sensiblement affectées par les changements climatiques, une évolution vers une gestion adaptative des espèces et des habitats est nécessaire.
6Cette étude se propose d’évaluer l’impact potentiel des changements climatiques sur la distribution géographique des aires favorables à la culture et à la conservation du tamarinier (T. indica), un fruitier autochtone sous-utilisé.
7Spécifiquement, l’étude s’est articulée autour des questions suivantes. D’un point de vue bioclimatique, quelle est l’étendue des aires favorables à la culture et à la conservation du tamarinier au Bénin ? Au regard des projections climatiques, quel est l’effet potentiel des changements climatiques sur l’étendue de ces aires et leur distribution géographique à l’horizon 2050 ? Quelles sont les implications de ces résultats pour l’élaboration de politiques futures de culture et de conservation du tamarinier ?
2. Matériel et méthodes
2.1. Milieu d’étude
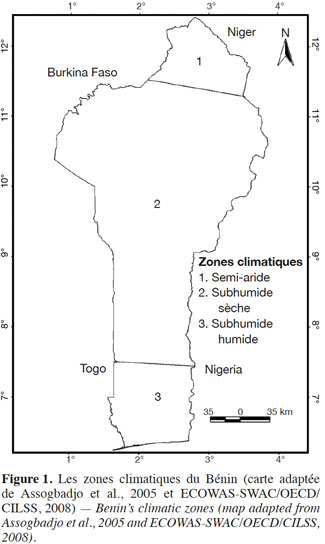
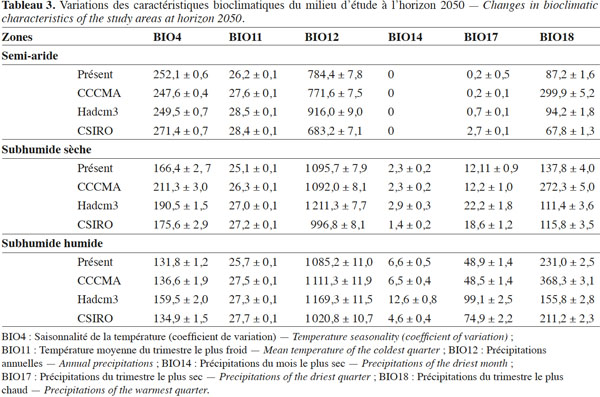
8La présente étude a été conduite en République du Bénin (6°-12,25°N; 0,40°-3°E). Le Bénin est situé dans le Dahomey Gap, un corridor d’écosystèmes savanicoles interrompant le bloc Ouest Africain de forêts sempervirentes (0°-3°E), suite aux changements climatiques intervenus au cours de l’Holocène (Salzmann et al., 2005). L’étude couvre trois zones climatiques du Bénin : la zone semi-aride (11°5’ N et 12° 25’ N), la zone subhumide sèche (7° 30’ N et 11°5’ N) et la zone subhumide humide (6° 25’ N et 7° 30’ N) (Figure 1).
2.2. Espèce modèle et collecte des données
9Le tamarinier (T. indica L., ex Tamarindus occidentalis [Gaertn], Tamarindus officinalis [Hook]) est une espèce semi-sempervirente de la famille des Fabaceae (El-Siddig et al., 2006). Originaire d’Afrique de l’Est ou de l’Ouest, de Madagascar ou d’Asie, le tamarinier est aujourd’hui très répandu sous les tropiques (Diallo et al., 2010). Selon les mêmes auteurs, le tamarinier a une grande importance dans les systèmes agroforestiers traditionnels au Sud du Sahara et présente un fort potentiel économique. Pour les populations rurales du Bénin, c’est une espèce essentiellement utilisée à des fins alimentaires, médicinales, culturelles et magico-thérapeutiques (Fandohan et al., 2010a). Elle fait partie des espèces autochtones sous-utilisées classées prioritaires en fonction de leur disponibilité sur le marché et de la facilité d’accès à leurs produits par les populations du Bénin, Burkina Faso, Mali, Sénégal et Niger, pour la diversification des productions agricoles et le développement socio-économique de la région (Akinnifesi et al., 2008). Hormis les usages alimentaires de sa pulpe connus sur tous les continents (El-Siddig et al., 2006), les graines du tamarinier sont riches en acides aminés et constituent une bonne source de protéines pouvant être utilisées pour prévenir la malnutrition infantile. En industrie, il a été établi que les graines peuvent substituer 30 % de la ration céréalière destinée aux bovins et les feuilles sont utilisées pour la préparation de divers produits pharmaceutiques comprenant des antibiotiques (Jama et al., 2008).
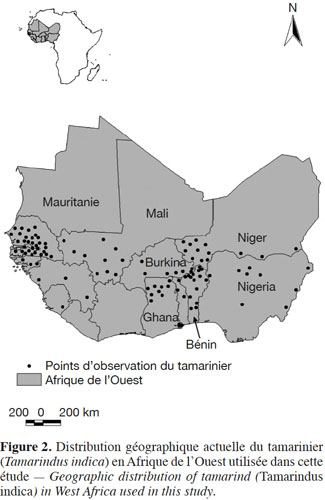
10Les coordonnées géographiques (longitude et latitude) de présence du tamarinier ont été enregistrées à partir de travaux de terrain dans les aires protégées et les systèmes agroforestiers dans les 77 communes du Bénin (Fandohan et al., 2011b). Afin de maximiser la précision des résultats de la modélisation, il est recommandé de veiller à ce que les données de présence (coordonnées) de l’espèce étudiée couvrent le plus possible la région où elle est influencée par les mêmes facteurs climatiques (Fitzpatrick et al., 2009). De ce fait, les coordonnées de présence de l’espèce hors du milieu d’étude, mais dans les mêmes zones climatiques couvertes, ont été collectées à travers son aire de distribution en Afrique de l’Ouest. Ces données additionnelles ont été obtenues en explorant des bases de données de biodiversité en ligne telles que le GBIF (Global Biodiversity Information Facility : www.gbif.org) et des articles publiés sur l’espèce (Bowe et al., 2010 ; Bourou et al., 2012).
11Les données climatiques actuelles et futures de l’aire globale considérée ont été téléchargées du site Internet de Worldclim (www.worldclim.org). Dix-neuf variables bioclimatiques (Busby, 1991) ont été générées à partir des données climatiques brutes (précipitations et température) préparées et mises sous formats compatibles avec le programme MaxEnt. À cet effet, le logiciel DIVA-GIS 7.5 a été utilisé.
12Les données sur les conditions climatiques actuelles ont été dérivées des données climatiques de 1950-2000, téléchargées de la base de données Worldclim version 1.4. (Hijmans et al., 2005; Hijmans et al., 2006). Pour les projections climatiques futures, trois modèles de circulation générale (Global Circulation Models : GCMs) parmi les plus recommandés (IPCC, 2007) ont été utilisés : les modèles CCCMA (Canadian Centre for Climate Modeling and Analysis), HadCM3 (Hadley Centre for Coupled Model version 3) et CSIRO (Commonwealth Scientific and Industrial Research Organization). Pour tous ces modèles, les projections faites pour 2050 ont été utilisées sous le scénario d’émission A2. Ce scénario a été utilisé de préférence car il prédit une situation considérée plus probable pour l’Afrique à l’horizon 2050 (Williams et al., 2007). Il décrit un monde très hétérogène, une population à fort taux de croissance avec un faible niveau technologique et de développement. Les couches climatiques utilisées ont une résolution de 0,05° (une grille de résolution d’approximativement 4,62 km x 4,62 km). Les modèles strictement bioclimatiques ont souvent été utilisés pour modéliser les habitats favorables aux espèces (ex., niche environnementale) parce que le climat est le principal déterminant de la niche écologique des espèces, à large échelle (ex., continent) (Parviainen et al., 2008). Cependant, les autres facteurs environnementaux seraient aussi déterminants à des échelles plus fines (ex., locale). Il est donc souvent recommandé de prendre en compte les facteurs pédologiques lorsque la zone d’étude s’étend sur une distance inférieure à 2 000 km (Pearson et al., 2003). Les données sur les types de sols du Bénin ont donc été combinées aux données bioclimatiques pour affiner les résultats (Faure et al., 1998).
2.3. Modélisation et validation du modèle
13Diverses méthodes statistiques sont utilisées pour modéliser la distribution des espèces, la distribution des habitats qui leur sont favorables ou encore pour estimer la probabilité de présence/absence d’une espèce donnée à une position géographique donnée (Guisan et al., 2000). MaxEnt (maximum entropy modeling) constitue l’une des méthodes de modélisation les plus puissantes susceptibles de générer de très bonnes informations biogéographiques tout en offrant une bonne discrimination des habitats favorables et non favorables à une espèce d’un point de vue bioclimatique (Phillips et al., 2006). L’intérêt de cette méthode pour cette étude est qu’elle combine les données de présence observées d’une espèce donnée avec les caractéristiques bioclimatiques actuelles aux points d’observation pour générer :
14– une carte globale des habitats potentiels de l’espèce dans l’aire considérée ;
15– une carte globale de la distribution future de ces habitats favorables vis-à-vis des projections climatiques faites pour la zone étudiée.
16Le programme MaxEnt (Phillips et al., 2006) a donc été préféré dans le cadre de cette étude.
17Au total, 108 points d’observations (dont 16 points au Bénin) ont été utilisés pour la modélisation (Figure 2). Les variables bioclimatiques ont été soumises à un test de corrélation pour sélectionner celles les moins corrélées (r < 0,85) compte tenu des biais que les corrélations ont sur les prédictions futures (Elith et al., 2011). Un test de Jackknife a été ensuite effectué sur les variables bioclimatiques considérées pour déterminer celles qui contribuent le plus à la modélisation. Pour évaluer le modèle, 25 % des points d’observation de l’espèce ont été utilisés pour tester le modèle et 75 % des points ont été utilisés pour calibrer le modèle. La validation croisée du modèle a été répétée quatre fois pour produire des estimations robustes des performances du modèle. La performance du modèle a été évaluée en utilisant la statistique AUC (Area Under the Curve) (Phillips et al., 2006). Un modèle est dit de bonne qualité si la valeur de l’AUC est supérieure à 0,90 (Swets, 1988). Un test binomial unilatéral a aussi été exécuté pour vérifier si la prédiction des habitats favorables à l’espèce est significativement différente d’une prédiction aléatoire (Anderson et al., 2002).
2.4. Analyse des données
18Les résultats de la modélisation ont été importés dans le logiciel ArcGIS 9.3 pour cartographier la distribution géographique actuelle des habitats favorables de l’espèce et celles du futur suivant chacun des scénarios utilisés. Pour évaluer la capacité actuelle et future du réseau national d’aires protégées à conserver l’espèce, une analyse des lacunes de représentation (gap analysis) des habitats très favorables à l’espèce dans les aires protégées a été effectuée en superposant chaque résultat issu de la modélisation à la carte du réseau d’aires protégées. Pour ce faire, la carte du réseau national d’aires protégées du Bénin a été extraite de la carte globale du réseau mondial des aires protégées (IUCN et al., 2009). La distribution de probabilité logistique d’occurence de l’espèce, générée par les modèles, a été considérée comme indicatrice de la qualité des habitats pour l’espèce. Pour une probabilité d’occurrence inférieure à la valeur du « maximum training sensitivity and specificity threshold », l’habitat est dit peu favorable à la culture ou à la conservation de l’espèce ; entre cette valeur et le « equal training sensitivity and specificity threshold », l’habitat est dit relativement favorable ; au-dessus de cette dernière, l’habitat est dit très favorable (Liu et al., 2005). L’étendue de chaque type d’habitat (aire et pourcentage) a été estimée en utilisant l’outil « spatial analyst » du logiciel ArcGIS 9.3 (Martínez et al., 2006). Les proportions d’habitats actuellement très favorables et susceptibles de devenir peu favorables dans le futur et vice versa ont été également estimées pour chaque modèle climatique. Le tamarinier étant une espèce affectionnant les sols profonds et bien drainés (El-Siddig et al., 2006), toutes les zones à sols hydromorphes ont été considérées comme non favorables à la culture de l’espèce et donc soustraites des résultats. Dans la même optique, nous avons enlevé les sols bruts, les sols modaux peu évolués et les sols lithiques. Par contre, compte tenu du fait que l’espèce pourrait trouver refuge sur ces derniers types de sols (ex. les collines), les zones ayant ces types de sols n’ont pas été soustraites de la carte des habitats favorables à la conservation de l’espèce. De même, comme l’espèce affectionne bien les forêts galeries où les sols sont souvent à hydromorphie temporaire (Fandohan et al., 2011a), ces types de sols n’ont pas été soustraits de la carte.
3. Résultats
3.1. Contribution des variables et validation du modèle
19L’analyse des corrélations et le test de Jackknife ont permis d’identifier six variables moins corrélées (r < 0,85) et contribuant le plus à la modélisation. Les précipitations annuelles (BIO12) et la température du trimestre le plus froid (BIO11) sont les variables ayant le plus contribué au modèle (Tableau 1). La modélisation a donné un AUC sensiblement égal à 0,91 avec une projection significativement différente d’une projection aléatoire (p < 0,0001 ; test binomial unilatéral), ce qui suggère une bonne capacité du modèle à prédire les habitats favorables à l’espèce. Les valeurs du « maximum training sensitivity plus specificity » et du « equal training sensitivity and specificity » étaient respectivement de 0,33 et 0,47.
3.2. Impact des changements climatiques sur l’étendue des habitats favorables à la culture du tamarinier
20D’après les résultats de la modélisation, près de 65 % du territoire national béninois (non compris les iles sur le fleuve Niger) sont actuellement très favorables à la culture du tamarinier (Figure 3a, tableau 2). Ces habitats très favorables à la culture de l’espèce sont essentiellement compris entre 12° et 7° de latitude Nord, ce qui correspond à la zone semi-aride et à la zone subhumide sèche. Les habitats peu favorables à l’espèce se situent essentiellement dans la zone subhumide sèche (entre 10° et 9° de latitude Nord) et la zone subhumide humide (en dessous de 7°5’ de latitude Nord).
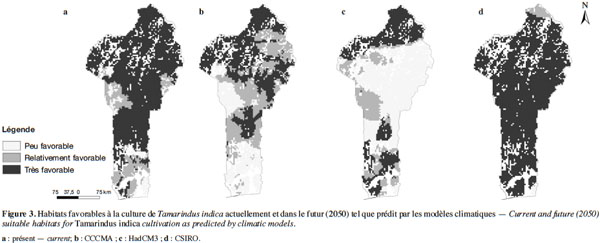
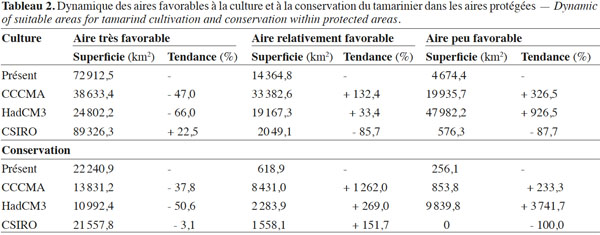
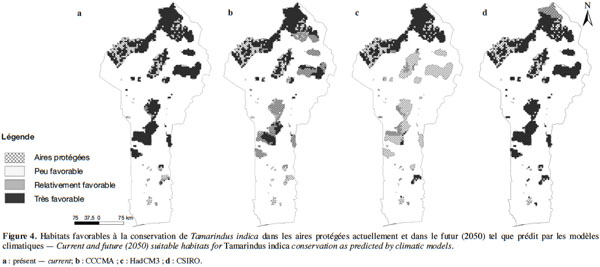
21Selon les projections bioclimatiques du modèle CCCMA, le tamarinier perdra près de 50 % des habitats qui sont actuellement très favorables à sa culture d’ici l’horizon 2050 (Tableau 2). Ces habitats seront essentiellement convertis en habitats relativement favorables à sa culture (Figure 3b). Ce modèle prédit également un accroissement de près de 327 % des habitats peu favorables à l’espèce, essentiellement par conversion des habitats actuellement relativement favorables. Le modèle HadCM3 donne des tendances similaires. Cependant, comparé au modèle précédent, ce modèle prédit un plus grand taux de conversion (près de trois fois plus élevé) d’habitats très favorables en habitats peu favorables à la culture du tamarinier, notamment entre 10°2’ et 7°5’ de latitude Nord (Tableau 2, figure 3c). Néanmoins, le modèle HadCM3 prédit également la conversion d’habitats actuellement relativement favorables à l’espèce en habitats très favorables (entre 6° et 7°5’ Nord). Contrairement aux autres modèles, le modèle CSIRO prédit une extension des habitats très favorables à la culture du tamarinier (de près de 23 % de l’aire actuelle) par conversion des habitats peu et relativement favorables (Tableau 2, figure 3d). Toutefois, le modèle prédit aussi une conversion d’une portion des habitats actuellement très favorables à l’espèce en habitats relativement favorables, entre 11° et 12°25’ Nord (Figure 3d).
3.3. Impact des changements climatiques sur l’aire occupée par les habitats favorables à la conservation du tamarinier dans les aires protégées
22Près de 87 % de la surface couverte par le réseau national d’aires protégées du Bénin serait actuellement très favorable à la conservation du tamarinier (Tableau 2, figure 4a). D’après les projections du modèle CCCMA, 62 % de cette superficie restera très favorable à la conservation de l’espèce à l’horizon 2050, soit une perte de 38 % des habitats très favorables (Tableau 2). On note également la conversion d’une portion des habitats actuellement très favorables à la conservation de l’espèce, en habitats relativement favorables, dans l’une des réserves les mieux protégées du pays (le Parc W), entre 11°5’ et 11° de latitude Nord (Figure 4b). Le modèle HadCM3 prédit une conversion plus élevée (une réduction de plus de 50 %) des habitats très favorables à la conservation de l’espèce en habitats peu et relativement favorables (Tableau 3). Il prédit notamment une extension importante des habitats peu favorables à la conservation du tamarinier. Toutefois, ce modèle suggère que la plupart des habitats très favorables à la conservation de l’espèce dans les parcs nationaux du pays (11°6’ à 12°2’ Nord) demeureront très favorables à l’horizon 2050 (Figure 4c). Le modèle CSIRO prédit une perte moins importante des habitats très favorables à la conservation du tamarinier (Tableau 2). Il prédit une extension des habitats très favorable à l’espèce vers les aires protégées en dessous de 7° Nord (Figure 4d). Cependant, il prédit également une conversion des habitats très favorables notamment dans le Parc W (entre 11°8’ et 12°2’ Nord) en habitats relativement favorables (Figure 4d).
4. Discussion
4.1. Modélisation et fiabilité du modèle
23La modélisation des niches écologiques a souvent été citée comme un puissant outil pour cartographier la distribution actuelle et future des espèces et prédire l’impact des changements climatiques sur leur distribution (Van Zonneveld et al., 2009 ; Nakao et al., 2010). Cependant, ces modèles ont aussi été très critiqués compte tenu de leurs faiblesses quant à prédire l’impact des changements climatiques sur la répartition géographique des espèces. Au nombre de ces faiblesses, on peut citer les incertitudes liées aux modèles utilisés, les difficultés à paramétrer les interactions écologiques, les réponses idiosyncratiques individuelles des espèces aux changements climatiques, les limitations de disséminations spécifiques à chaque espèce, la plasticité des limites physiologiques et les réponses adaptatives des agents disséminateurs (Elith et al., 2006 ; Schwartz, 2012). Par ailleurs, le postulat de base selon lequel le climat actuel où une espèce est rencontrée (sa niche réalisée actuelle) est sa niche originelle, est également discutable. Il est en effet possible qu’au moment de l’établissement de l’espèce dans ses zones d’occurrence actuelles, le climat ait été très différent (plus humide ou plus sec) et que sa présence actuelle implique plusieurs millénaires d’adaptation à différents changements climatiques. Dans le cas du tamarinier, l’apparition de sa tribu en Afrique (les Detarieae) remonte à 25 millions d’années (Pan et al., 2010). Il serait donc hasardeux de prédire une disparition de l’espèce de son aire actuelle à l’horizon 2050. Malgré ces faiblesses, ces modèles procurent des informations bioclimatiques très importantes en matière de prise de décisions, notamment pour identifier de nouvelles zones potentiellement favorables à la culture (Cuni-Sanchez et al., 2010) ou à la conservation d’une espèce donnée (Schwartz, 2012). De même, les cas de mortalité de fruitiers agroforestiers observés pendant les fortes poches de sècheresse dans le Sahel au cours du siècle passé (Maranz, 2009) montrent l’intérêt des projections climatiques pour de futures prises de décisions (ex., Quelles espèces agroforestières prioriser dans les politiques de diversification agricole et/ou de conservation ?). En effet, il a été récemment mis en évidence que certains fruitiers agroforestiers à fortes valeurs économiques ont été introduits dans le Sahel à la faveur des périodes où les conditions climatiques y étaient plus humides (Maranz, 2009). Selon le même auteur, la migration des isohyètes pluviométriques vers le Sud (parfois en dessous de 600 mm pour la pluviométrie annuelle) aurait eu des effets néfastes sur ces espèces. Les modèles climatiques peuvent donc être utilisés pour identifier les zones potentielles où les variables bioclimatiques pourraient significativement changer de valeurs et ainsi influencer la physiologie, la productivité et la dynamique des populations des espèces agroforestières. Ces informations pourront ensuite être utilisées pour l’élaboration de politiques adéquates de production et de conservation.
24Dans le cadre de cette étude, en général, les modèles prédisent une élévation des températures à l’horizon 2050 dans l’aire d’étude (ex., une augmentation d’un à deux degrés selon la zone et selon le modèle, pour la température du trimestre le plus froid ; tableau 3). Les résultats varient donc selon qu’un modèle prédise une augmentation ou une diminution des précipitations et selon les modifications dans la distribution des précipitations.
4.2. Culture du tamarinier
25Le tamarinier peut être cultivé dans des régions où les températures annuelles minima sont comprises entre 9,5 °C et 20 °C et les maxima entre 33 °C et 37 °C, avec une pluviométrie annuelle de 500 mm à 1 500 mm (El-Siddig et al., 2006). Il pourrait supporter les climats marqués par une saison sèche prononcée (six mois) grâce à un profond système d’enracinement. Les adultes pourraient faire face à des extrêmes de -3 °C et 47 °C sans dégâts majeurs. L’espèce peut aussi être plantée sur des sols pauvres. Selon les mêmes auteurs, de fortes pluviométries de l’ordre de 4 000 mm pourraient permettre une bonne croissance de l’espèce, mais empêcher la floraison et donc la fructification. Bien que les prédictions climatiques (Tableau 3) des modèles utilisés demeurent toutes dans les fourchettes favorables à l’espèce, les écotypes de la zone d’étude pourraient, comme l’indiquent les résultats, être affectés par les changements de valeurs et de magnitudes des variables bioclimatiques.
26Les variations des caractéristiques bioclimatiques peuvent significativement affecter les rendements des fruitiers (Challinor et al., 2006). En considérant la situation actuelle comme celle de base, et la physiologie de reproduction de l’espèce, les zones dites très favorables et converties en zones relativement ou peu favorables à l’horizon 2050 sont donc des zones où les individus de l’espèce pourraient connaitre une réduction de leur niveau actuel de production et vice versa.
27D’après nos résultats, la culture du tamarinier serait actuellement possible sur environ 65 % du territoire béninois, notamment en zones semi-aride et subhumide sèche. Cela confirme les observations antérieures sur la niche réalisée du tamarinier au Bénin (Fandohan et al., 2011b). Ces zones offrent à l’espèce une saison sèche caractérisée par l’harmattan et une grande réduction de l’hygrométrie dont elle aurait besoin pour une bonne maturation des fruits et un bon rendement (El-Siddig et al., 2006). Les modèles CCCMA et HadCM3 prédisent une grande réduction des aires favorables à la culture de l’espèce, contrairement au modèle CSIRO. À l’opposé de ce dernier, ces modèles prédisent une augmentation des précipitations dans les habitats actuellement très favorables à l’espèce (Tableau 3). Néanmoins, le modèle CSIRO prédit également une petite réduction des habitats favorables à l’espèce, mais plutôt par réduction de la pluviométrie. Bien que ces trois modèles fassent partie des plus recommandés (IPCC, 2007), ils montrent des désaccords et incertitudes sur l’évolution des précipitations en Afrique de l’Ouest. Ce sont ces différences qui expliquent les variations observées au niveau des prédictions (Christensen et al., 2007 ; Pittock, 2009). Ces auteurs suggèrent d’ailleurs que les incertitudes sont inévitables, mais que les risques sont certains. Plutôt que de prédire ce qui arrivera, ces modèles donnent donc une meilleure idée de ce qui pourrait arriver et offrent des alternatives d’adaptation en fonction de différents scenarios. Ainsi, l’augmentation des précipitations, notamment pendant le trimestre le plus chaud (tableau 3, CCCMA et HadCM3) qui correspond (dans les zones semi-aride et subhumide sèche) au pic de maturation des fruits du tamarinier (respectivement février-mars-avril et décembre-janvier-février), pourrait effectivement avoir des effets dommageables sur la production en cours et celle de l’année suivante. De même, la réduction des précipitations en interaction avec les autres variables bioclimatiques pourrait soit étendre les conditions favorables à la culture du tamarinier, soit en dessous d’un seuil donné, les réduire (tableau 3, CSIRO).
28En résumé, on peut dire que deux des modèles (CCCMA et HadCM3) prédisent une régression significative des habitats où les écotypes locaux de tamarinier conserveront leur niveau actuel de production à l’horizon 2050, alors que le modèle CSIRO prédit une extension de ces habitats. Toutefois, chez un grand nombre d’espèces tropicales, il a été observé un ajustement de la physiologie de reproduction en fonction des conditions climatiques (Yadav et al., 2008 ; Pittock, 2009). Ainsi, pour autant que les limites de tolérance des facteurs écologiques le permettront, les écotypes locaux de l’espèce pourraient être amenés à s’ajuster aux changements.
4.3. Conservation du tamarinier
29Nos résultats suggèrent que le réseau actuel d’aires protégées est très favorable à la conservation des écotypes locaux de tamarinier. Les modèles CCCMA et HadCM3 prédisent une diminution des habitats très favorables à l’espèce à l’intérieur du réseau d’aires protégées du Bénin (par augmentation des précipitations), alors que le modèle CSIRO prédit leur extension. Bien que les individus adultes de l’espèce soient tolérants à des extrêmes de température, les juvéniles sont assez sensibles aux stress hydriques des régions semi-arides et arides, ce qui limite la régénération naturelle chez l’espèce (Diallo, 2001). Les galeries forestières seraient très importantes dans la conservation de l’espèce compte tenu du fait que ces habitats lui offrent des conditions microclimatiques et pédologiques plus favorables à la survie des juvéniles et à la multiplication par voie végétative (Fandohan et al., 2010b). Ces habitats constituent ainsi les seules zones où les individus de l’espèce vivent en peuplement.
30L’augmentation des précipitations (CCCMA et HadCM3) pourrait entrainer une augmentation de l’écoulement et du débit des rivières, entrainer des débordements/inondations, et augmenter le déracinement des arbres dans les galeries forestières (Primack, 2000). La diminution des précipitations et l’augmentation des températures pourraient aussi significativement perturber la micro-écologie de ces habitats (ex. le régime hydrologique assurant l’équilibre), augmenter l’évapotranspiration et avoir des répercussions néfastes sur la physiologie des individus, la régénération et la dynamique des populations de l’espèce (Hulme, 2005 ; Serrat-Capdevila et al., 2007). Le devenir de l’espèce dans les aires protégées sera donc lié à celui de ces habitats et à sa capacité à supporter les extrêmes de température et de précipitation.
31En dépit de la réduction d’habitats très favorables prédite par deux des modèles, aucun des modèles n’a prédit une incapacité du réseau national d’aires protégées à offrir des habitats très favorables à l’espèce à l’horizon 2050.
4.4. Implications de l’étude
32Les conditions climatiques prévalant dans les zones semi-aride et subhumide sèche actuelles sont très favorables à la culture des écotypes locaux du tamarinier. En considérant les conditions actuelles comme optimales pour ces écotypes, en cas d’augmentation des précipitations (modèles CCCMA et HadCM3), les zones actuellement très favorables à leur culture (semi-aride et subhumide sèche) pourraient devenir peu favorables (ex., probable réduction des rendements des arbres) à l’horizon 2050. En cas de diminution des précipitations (modèle CSIRO), les zones qui leur sont actuellement peu favorables (subhumide humide) pourraient devenir très favorables. Dans le schéma d’une aridification (CSIRO), la culture et la conservation des écotypes locaux de l’espèce seraient possibles sur toute l’étendue de la zone d’étude et/ou nécessiteraient l’introduction d’écotypes originaires des zones plus arides (ex., Soudan, Mali, etc.). De même, le réseau national d’aires protégées offre des conditions très favorables à la conservation des écotypes locaux de l’espèce. En dépit des réductions projetées, quelles que soient les projections, ce réseau reste efficace pour fournir des habitats très favorables aux écotypes locaux de l’espèce à l’horizon 2050.
33Pour faciliter une gestion adaptative des fruitiers agroforestiers sous-utilisés et leur intégration dans les systèmes de production, des études approfondies doivent être menées sur l’impact potentiel des changements climatiques sur ces espèces (habitats favorables, phénologie et productivité), notamment, dans les pays sahéliens plus arides que le Bénin (Mauritanie, Niger, Mali, Burkina Faso, Sénégal, etc.) où les variations des variables bioclimatiques seront plus sévères, et pour des espèces agroforestières ayant une distribution moins large que celle du tamarinier (ex. le dattier du désert, Balanites aegyptiaca (L.) Del. ; le palmier dattier, Phoenix dactylifera L. ; le marula, Sclerocarya birrea (A.Rich.) Hochst. ; le karité, Vitellaria paradoxa Gaertn. ; le jujubier, Ziziphus mauritiana (Lam.). La constitution de bases de données sur l’impact potentiel des changements climatiques sur les fruitiers agroforestiers sous-utilisés en fonction des différents scenarios disponibles pourrait accélérer les prises de décisions adaptatives dans le cadre de leur intégration dans les systèmes formels de production (ex. élaborer des modèles de prise de décisions intégrant les changements climatiques pour identifier les fruitiers les plus éligibles à promouvoir). De telles études doivent être menées au fur et à mesure que les pouvoirs prédictifs des modèles climatiques seront améliorés, pour une plus grande pertinence des prises de décisions car de mauvaises options de gestion peuvent s’avérer encore plus dramatiques que les changements climatiques. Les variations des résultats des différents modèles permettent d’avoir une meilleure idée des incertitudes et appellent à en tenir compte dans les planifications. Cela appelle aussi à la prise en compte de la physiologie de reproduction des espèces étudiées et de leur base génétique pour une meilleure interprétation des résultats.
34Remerciements
35Nous remercions la Chinese Academy of Sciences et la National Natural Sciences Foundation of China qui ont soutenu cette étude grâce à deux financements accordés à B. Fandohan (respectivement CAS-Fellowship n°2012Y1ZA0009 et NSFC-Grant n°312111172). Nous remercions également les lecteurs qui ont aidé à améliorer la qualité scientifique du manuscrit. Enfin, nous remercions Messiah Fandohan et Bright Fandohan qui ont également consenti à des sacrifices au cours de la rédaction du manuscrit.
Bibliographie
Akinnifesi F.K. et al., 2008. Indigenous fruit trees in the tropics: domestication, utilization and commercialization. London: CAB International.
Anderson R.P., Gómez-Laverde M. & Peterson A.T., 2002. Geographical distributions of spiny pocket mice in South America: insights from predictive models. Global Ecol. Biogeogr., 11, 131-141.
Assogbadjo A.E., Sinsin B. & Van Damme P., 2005. Caractères morphologiques et production des capsules de baobab (Adansonia digitata L.) au Bénin. Fruits, 60(5), 327-340.
Boko M. et al., 2007. Africa. In: Parry M.L. et al., eds. Climate change 2007: impacts, adaptation and vulnerability. Contribution of working group II to the 4th assessment report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 433-467.
Bourou S., Bowe C., Diouf M. & Van Damme P., 2012. Ecological and human impacts on stand density and distribution of tamarind (Tamarindus indica L.) in Senegal. Afr. J. Ecol., 50, 253-265.
Bowe C. & Haq N., 2010. Quantifying the global environmental niche of an underutilized tropical fruit tree (Tamarindus indica) using herbarium records. Agric. Ecosyst. Environ., 139(1-2), 51-58.
Busby J.R., 1991. BioClim – a bioclimatic analysis and prediction system. In: Margules C.R. & Austin M.P., eds. Nature conservation: cost effective biological surveys and data analysis. Canberra: CSIRO, 64-68.
Busby J.W., Smith T.G., White K.L. & Strange S.M., 2010. Locating climate insecurity: where are the most vulnerable places in Africa? Austin, TX, USA: University of Texas, The Robert Strauss Center for International Security and Law, Climate Change and African Political Stability (CCAPS) Programme.
Challinor A.J., Wheeler T.R., Osborne T.M. & Slingo J.M., 2006. Assessing the vulnerability of crop productivity to climate change thresholds using an integrated crop-climate model. In: Schellnhuber J. et al., eds. Avoiding dangerous climate change. Cambridge, UK: Cambridge University Press, 187-194.
Christensen J.H. et al., 2007. Regional climate projections. In: Solomon S. et al., eds. Climate change 2007: the physical science basis. Contribution of working group I to the 4th assessment report of the Intergovernmental Panel on Climate Change. Cambridge, UK; New York, USA: Cambridge University Press, 847-940.
Cuni-Sanchez A., Osborne P.E. & Haq N., 2010. Identifying the global potential for baobab tree cultivation using ecological niche modeling. Agroforest. Syst., 80(2), 191-201.
Diallo B.O., 2001. Biologie de la reproduction et évaluation de la diversité génétique chez une légumineuse : Tamarindus indica L. (Caesalpinioideae). Thèse de doctorat : Université Montpellier II (France).
Diallo B.O. et al., 2010. Variation des caractères biométriques des graines et des plantules de neuf provenances de Tamarindus indica L. (Caesalpinioideae). Fruits, 65(3), 153-167.
ECOWAS-SWAC/OECD/CILSS, 2008. Climate and climate change. The atlas on regional integration in West Africa, http://www.atlas-westafrica.org, (15/10/12).
Elith J. et al., 2006. Novel methods improve prediction of species’ distributions from occurrence data. Ecography, 29, 129-151.
Elith J., Kearney M. & Philips S., 2011. The art of modeling range-shifting species. Methods Ecol. Evol., 1, 330-342.
El-Siddig K. et al., 2006. Tamarind, Tamarindus indica L. Southampton, UK: Southampton Centre for Underutilised Crops.
Fandohan B. et al., 2010a. Women’s traditional knowledge, use value and the contribution of tamarind (Tamarindus indica L.) to rural households’ cash income in Benin. Econ. Bot., 64(3), 248-259.
Fandohan B. et al., 2010b. Impact of habitat type on the conservation status of tamarind (Tamarindus indica L.) populations in the W National Park of Benin. Fruits, 65(1), 11-19.
Fandohan B., Assogbadjo A.E., Glèlè Kakaï R. & Sinsin B., 2011a. Effectiveness of a protected areas network in the conservation of Tamarindus indica L. (Leguminosae-Caesalpinioideae) in Benin. Afr. J. Ecol., 49, 40-50.
Fandohan B., Assogbadjo A.E., Glèlè Kakaï R. & Sinsin B., 2011b. Geographical distribution, tree density and fruit production of Tamarindus indica (Fabaceae) across three ecological regions in Benin. Fruits, 66(2), 53-62.
Faure P. & Volkoff B., 1998. Some factors affecting regional differentiation of the soils in the Republic of Benin (West Africa). Catena, 32, 281-306.
Fitzpatrick M.C. & Hargrove W.W., 2009. The projection of species distribution models and the problem of non-analog climate. Biodivers. Conserv., 18, 2255-2261.
Gouwakinnou N.G., 2011. Population ecology, uses and conservation of Sclerocarya birrea (A.Rich) Hocchst. (Anacardiaceae) in Benin, West Africa. Phd thesis: University of Abomey Calavi (Republic of Benin).
Guisan A. & Zimmermann N.E., 2000. Predictive habitat distribution models in ecology. Ecol. Model., 135, 147-186.
Hannah L., Midgley G.F. & Millar D., 2002. Climate change integrated conservation strategies. Global Ecol. Biogeogr., 11, 485-495.
Heller N.E. & Zavaleta E.S., 2009. Biodiversity management in the face of climate change: a review of 22 years of recommendations. Biol. Conserv., 142, 14-32.
Hijmans R.J. et al., 2005. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol., 25, 1965-1978.
Hijmans R.J. et al., 2006. The WorldClim Interpolated Global Terrestrial Climate Surfaces, version 1.4., http://biogeo.berkeley.edu/, (08/01/12).
Hulme P.E., 2005. Adapting to climate change: is there scope for ecological management in the face of a global threat? J. Appl. Ecol., 42, 784-794.
IPCC, 2007. Climate change: synthesis report. New York, USA: Cambridge University Press.
IUCN, UNEP, 2009. The world database on protected areas (WDPA). Cambridge, UK: UNEP-WCMC, http://protectedplanet.net/, (08/01/12).
Jama B.A., Mohamed A.M., Mulatya J. & Njui A.N., 2008. Comparing the ‘‘Big Five’’: a framework for the sustainable management of indigenous fruit trees in the drylands of East and Central Africa. Ecol. Indic., 8, 170-179.
Leakey R.R.B. & Simons A.J., 1998. The domestication and commercialisation of indigenous trees in agroforestry for the alleviation of poverty. Agroforest. Syst., 38, 165-176.
Liu C., Berry P.M., Dawson T.P. & Pearson R.G., 2005. Selecting thresholds of occurrence in the prediction of species distributions. Ecography, 28, 385-393.
Maranz S., 2009. Tree mortality in the African Sahel indicates an anthropogenic ecosystem displaced by climate change. J. Biogeogr., 36, 1181-1193.
Martínez I., Carreño F., Escudero A. & Rubio A., 2006. Are threatened lichen species well-protected in Spain? Effectiveness of a protected areas network. Biol. Conserv., 133, 500-511.
Nakao K. et al., 2010. Assessing the impact of land use and climate change on the evergreen broad-leaved species of Quercus acuta in Japan. Plant Ecol., 212, 229-243.
Pan A.D., Jacobs B.F. & Herendeen P.S., 2010. Detarieae sensulato (Fabaceae) from the Late Oligocene (27.23 Ma) Guang River flora of north-western Ethiopia. Bot. J. Linn. Soc., 163(1), 44-54.
Parviainen M., Luoto M., Ryttari T. & Heikkinen R.K., 2008. Modelling the occurrence of threatened plant species in taiga landscapes: methodological and ecological perspectives. J. Biogeogr., 35, 1888-1905.
Pearson R.G. & Dawson T.P., 2003. Predicting the impacts of climate change on the distribution of species: are bioclimatic envelope models useful? Global Ecol. Biogeogr., 12, 361-371.
Phillips S.J., Anderson R.P. & Schapire R.E., 2006. Maximum entropy modelling of species geographic distributions. Ecol. Model., 190, 231-259.
Pittock B.A., 2009. Climate change: the science, impacts and solutions. Collingwood, Australia: CSIRO Publishing; London: Earthscan.
Primack A.G.B., 2000. Simulation of climate-change effects on riparian vegetation in the Pere Marquette River, Michigan. Wetlands, 20(3), 538-547.
Salzmann U. & Hoelzmann P., 2005. The Dahomey Gap: an abrupt climatically induced rain forest fragmentation in West Africa during the late Holocene. Holocene, 15(2), 190-199.
Schwartz M.W., 2012. Using niche models with climate projections to inform conservation management decisions. Biol. Conserv., 155, 149-156.
Scott D., 2005. Integrating climate change into Canada’s national park system. In: Lovejoy T.E. & Hannah L., eds. Climate change and biodiversity. New Haven, CT, USA: Yale University Press, 342-345.
Serrat-Capdevila A. et al., 2007. Modeling climate change impacts – and uncertainty – on the hydrology of a riparian system; San Pedro basin (Arizona/Sonora). J. Hydrol., 347(1–2), 48-66.
Swets J.A., 1988. Measuring the accuracy of diagnostic systems. Science, 240, 1285-1293.
Van Zonneveld M., Koskela J., Vinceti B. & Jarvis A., 2009. Impact of climate change on the distribution of tropical pines in Southeast Asia. Unasylva, 231/232, 24-29.
Williams J.W., Jackson S.T. & Kutsbach J.E., 2007. Projected distribution of novel and disappearing climates by 2100 AD. Proc. Natl. Acad. Sci. U.S.A., 104, 5738-5742.
Yadav R.K. & Yadav A.S., 2008. Phenology of selected woody species in a tropical dry deciduous forest in Rajasthan, India. Trop. Ecol., 49(1), 25-34.
Pour citer cet article
A propos de : Belarmain Fandohan
Université d’Abomey-Calavi. École Nationale Supérieure des Sciences et Techniques Agronomiques. Laboratoire d’Écologie Appliquée. BP 95. Kétou (Bénin). E-mail : bfandohan@gmail.com – Chinese Academy of Sciences. Institute of Geographic Sciences and Natural Resources Research. International Ecosystem Management Partnership (IEMP). United Nations Environment Programme. No. 11A Datun Rd. Beijing 100101 (China).
A propos de : Gerard N. Gouwakinnou
Université d’Abomey-Calavi. École Nationale Supérieure des Sciences et Techniques Agronomiques. Laboratoire d’Écologie Appliquée. BP 95. Kétou (Bénin).
A propos de : Noël H. Fonton
Université d’Abomey-Calavi. Laboratoire d’Étude et de Recherche en Statistique Appliquée et Biométrie. 01 BP 526. Cotonou (Bénin).
A propos de : Brice Sinsin
Université d’Abomey-Calavi. École Nationale Supérieure des Sciences et Techniques Agronomiques. Laboratoire d’Écologie Appliquée. BP 95. Kétou (Bénin).
A propos de : Jian Liu
Chinese Academy of Sciences. Institute of Geographic Sciences and Natural Resources Research. International Ecosystem Management Partnership (IEMP). United Nations Environment Programme. No. 11A Datun Rd. Beijing 100101 (China).
