


- Portada
- Volume 18 (2014)
- numéro 4
- Effet des polyamines sur la réduction du chrome hexavalent par des souches bactériennes et leur résistance


Vista(s): 1554 (8 ULiège)
Descargar(s): 122 (1 ULiège)

Effet des polyamines sur la réduction du chrome hexavalent par des souches bactériennes et leur résistance

Notes de la rédaction
Reçu le 14 octobre 2013, accepté le 7 octobre 2014
Résumé
Les polyamines sont impliquées dans plusieurs fonctions chez les bactéries. Dans ce travail, on a étudié le rôle de ces composés dans la réduction du chrome hexavalent (Cr[VI]) par trois souches bactériennes isolées à partir des sites contaminés par les effluents des tanneries. Les souches sont identifiées comme étant Serratia proteamaculans, Leucobacter chromiireducens et Brevibacterium frigoritolerans. L’inhibition de la synthèse des polyamines par l’α-difluorométhylornitine (DFMO) a provoqué une diminution de la tolérance des souches bactériennes au Cr(VI), ce qui montre le rôle des polyamines endogènes dans la résistance au Cr(VI). L’application exogène des polyamines (putrescine, spermidine, cadavérine) a permis de stimuler la croissance bactérienne et la réduction du Cr(VI) par les souches bactériennes, en milieu Luria-Bertani. Les résultats montrent l’importance des polyamines dans la réponse bactérienne au stress métallique causé par le Cr(VI).
Abstract
Effect of polyamines on the reduction of hexavalent chromium by bacterial strains and their resistance. Polyamines are involved in several functions in bacteria. In this study, we examined the role of polyamines in hexavalent chromium (Cr[VI]) reduction by three bacterial strains isolated from sites contaminated by tannery effluents. The strains were identified as Serratia proteamaculans, Leucobacter chromiireducens and Brevibacterium frigoritolerans. The inhibition of polyamine synthesis by α-difluoromethylornithine (DFMO) caused a decrease in Cr(VI) tolerance in the bacterial isolates, indicating the role of endogenous polyamines in resistance to Cr(VI). The exogenous application of polyamines (putrescine, spermidine, cadaverine) was found to stimulate growth and Cr(VI) reduction by the bacterial strains in Luria-Bertani medium. The results show the importance of polyamines in response to heavy metal stresses, especially Cr(VI) toxicity.
Tabla de contenidos
1. Introduction
1Le chrome est utilisé à travers le monde dans plusieurs activités industrielles (Lauwerys et al., 2007). Il se retrouve sous plusieurs états de valences. Dans l’environnement, il est particulièrement présent sous deux formes, la forme hexavalente (Cr[VI]) et la forme trivalente (Cr[III]), avec des propriétés physico-chimiques et biologiques différentes. L’élimination du Cr(VI) par les micro-organismes peut être significativement influencée par des facteurs tels que la biomasse cellulaire, la concentration initiale du Cr(VI), la source de carbone, le pH, la température, l’oxygène dissous, le potentiel d’oxydo-réduction et la présence d’anions et de cations (Somasundaram et al., 2009). Cependant, l’effet des polyamines sur la réduction bactérienne du Cr(VI) n’a pas été évoqué dans la littérature.
2Les polyamines (PAs) sont des molécules issues du métabolisme des acides aminés et du cycle de l’urée, naturellement présentes dans toutes les cellules animales, végétales ou bactériennes. Ces composés servent de facteurs essentiels ou de stimulateurs de la croissance pour plusieurs micro-organismes (De Bandt et al., 2006), pour la biosynthèse de sidérophores (Brian et al., 2007), la résistance à l’acidité et l’élimination des radicaux libres (Brian et al., 2007) et la formation de biofilms (Wortham et al., 2007). Les PAs sont aussi reconnues pour leur rôle dans la résistance bactérienne aux antibiotiques (Kwon et al., 2006). Les polyamines agissent en tant qu’antioxydants en réponse au stress métallique causé par le cadmium et le cuivre chez le blé (Groppa et al., 2007). Johnson et al. (2012) ont montré que les PAs peuvent jouer un rôle dans la protection de la membrane extérieure des bactéries contre les antibiotiques et les dégâts oxydatifs. Cependant, leur implication dans la réponse au stress chromique chez les bactéries n’a pas été étudiée. Dans ce travail, nous nous sommes intéressés à étudier l’effet de ces polyamines (exogènes) notamment (putrescine, spermidine et cadavérine) en réponse au stress métallique causé par le Cr(VI) sur la croissance et la réduction du Cr(VI) par des souches bactériennes. Nous avons aussi testé l’effet d’un inhibiteur de la synthèse des polyamines endogènes, l’ α-difluorométhylornitine (DFMO), sur la résistance de ces souches au Cr(VI).
2. Matériel et méthodes
2.1. Isolement et identification des souches bactériennes
3L’isolement des bactéries résistantes et réductrices du Cr(VI) est réalisé comme décrit par Tahri Joutey et al. (2011). Les isolats sont identifiés par la méthode de biologie moléculaire par analyse de l’ADNr 16S (Tahri Joutey et al., 2013).
2.2. Évaluation de l’effet de la DFMO sur la résistance au Cr(VI)
4La détermination de la concentration minimale inhibitrice (CMI) au Cr(VI) est réalisée sur milieu Luria-Bertani (LB) solide. Un aliquot de 100 µl d’une pré-culture est étalé sur des boites contenant le milieu LB-agar et additionné de concentrations croissantes de Cr(VI), sous forme de K2Cr2O7, de 100 à 1 000 mg·l-1. Pour étudier l’effet des polyamines endogènes sur la tolérance bactérienne au Cr(VI), l’α-difluorométhylornitine (DFMO) (1 mM), inhibiteur de synthèse des polyamines par l’ornithine décarboxylase (ODC), est ajouté au milieu LB-agar contaminé par les différentes concentrations de Cr(VI). La mesure de la croissance bactérienne est réalisée après 48 h d’incubation à 30 °C.
2.3. Effet des polyamines sur la réduction du Cr(VI)
5Le milieu LB est réparti dans des erlenmeyers de 100 ml après stérilisation à l’autoclave (121 °C pendant 20 min). Le Cr(VI) est ajouté pour obtenir des concentrations de 1 et 2 mM. Les polyamines putrescine, spermidine et cadavérine sont ajoutées à des concentrations de 1 et 10 mM. Les essais sont réalisés en trois répétitions. Des témoins non inoculés sont préparés de la même façon pour tenir compte de la réduction abiotique du Cr(VI), s’il y en a. Après 24 h d’incubation à 30 °C sous agitation orbitale (150 rpm), le Cr(VI) est dosé dans les surnageants des cultures par la méthode colorimétrique au diphényl-carbazide (DPC) (Thacker et al., 2006).
3. Résultats et discussion
3.1. Identification moléculaire des isolats bactériens
6Les souches bactériennes isolées CRB1, CRB2 et CRB3 sont identifiées respectivement comme étant Serratia proteamaculans, Leucobacter chromiireducens et Brevibacterium frigoritolerans. Les séquences nucléotidiques de leur ADNr 16S sont déposées dans la base de données GenBank (http://www.ncbi.nlm.nih.gov) sous les numéros d’accession : HF913431, HE963772 et HF937250, respectivement.
3.2. Effet de la DFMO sur la résistance au Cr(VI)
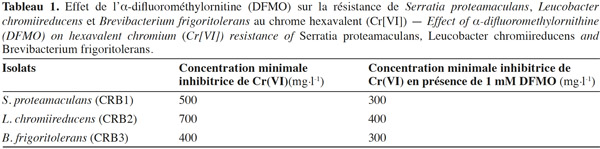
7Les résultats montrent que la tolérance des isolats bactériens au Cr(VI) a diminué en présence de 1 mM du DFMO (Tableau 1), ce qui montre le rôle des polyamines endogènes dans la résistance au Cr(VI).
8L’implication des polyamines dans la gestion du stress a été largement rapportée chez les végétaux (Liu et al., 2007) et attribuée essentiellement à leur capacité à éliminer les radicaux libres et les ROS (Reactive Oxygen Species) (Ha et al., 1998). Chez les micro-organismes, Johnson et al. (2012) ont montré que les PAs sont produites en présence de Mg2+ comme polycations pour lier les lipopolysaccharides (LPS) et protéger la membrane extérieure des bactéries contre les antibiotiques et les dégâts oxydatifs. Par ailleurs, Sharma et al. (2006) ont rapporté que le Cr(VI) peut être spontanément oxydé pour produire les ROS, mais sa réduction par deux électrons catalysée par la chromate réductase minimise la possibilité de produire les radicaux libres nocifs et augmente la résistance bactérienne au Cr(VI). Parmi les composants non enzymatiques impliqués dans l’élimination des ROS figurent les polyamines. On peut donc suggérer que les polyamines augmentent la résistance de nos souches au stress métallique à travers l’élimination des ROS.
3.3. Effet des polyamines sur la réduction du Cr(VI)
9L’effet des PAs exogènes sur la réduction du Cr(VI) varie d’un isolat à l’autre, d’une concentration du Cr(VI) à l’autre et selon la quantité ajoutée de ces polyamines (Figure 1). Ces résultats suggèrent que les mécanismes impliqués dans la résistance au stress métallique sont complexes, ce qui rend difficile l’explication des résultats obtenus, surtout qu’il s’agit du premier travail effectué dans ce domaine. Cependant, puisque les courbes de croissance et de réduction du Cr(VI) varient dans le même sens, on peut suggérer que c’est la stimulation de la croissance cellulaire par ces PAs qui serait responsable de la stimulation de la capacité des micro-organismes étudiés à réduire le Cr(VI). Les PAs agissent comme des facteurs de croissance intracellulaires. La théorie « polyamine modulon » explique que les polyamines stimulent la synthèse de plusieurs facteurs clés tels que les protéines de synthèse des oligopeptides, l’adénylate cyclase, l’ARN polymérase et plusieurs facteurs de transcription (Terui et al., 2004 ; Igarashi et al., 2006).
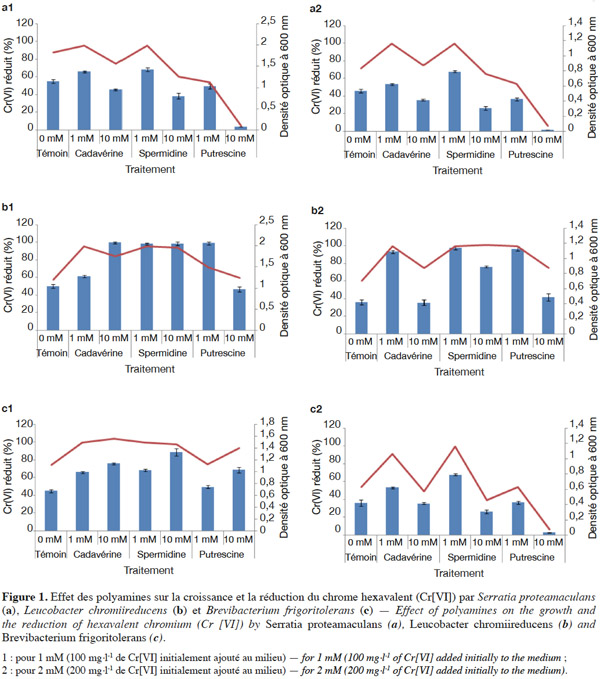
10Par ailleurs, plusieurs auteurs ont rapporté que la réduction du Cr(VI) est stimulée par des cations, notamment le Cu2+ chez de nombreuses bactéries (Ibrahim et al., 2011 ; Sayel et al., 2012). Les polyamines se retrouvent sous forme de cations (putrescine2+, spermidine3+ et spermine4+) au pH cytoplasmique et peuvent agir en tant que cofacteurs enzymatiques dans la réaction de réduction du Cr(VI).
11Les études de l’effet des polyamines sur la réponse au stress métallique (Cr[VI]) chez les bactéries devraient être poursuivies en vue de vérifier leur implication éventuelle en tant que facteurs de croissance intracellulaires, antioxydants (élimination des radicaux libres et des ROS), bloqueurs des porines bactériennes (diminution de la perméabilité membranaire vis-à-vis du Cr[VI]) ou cofacteurs enzymatiques de la chromate réductase.
Bibliographie
Brian W., Marcos O. & Chandra P., 2007. Polyamines in bacteria: pleiotropic effects yet specific mechanisms. Adv. Exp. Med. Biol., 603, 106-115.
De Bandt J.-P., Moinard C. & Cynober L., 2006. Métabolisme et fonctions des polyamines. Immunoanalyse Biol. Spécialisée, 21, 333-341.
Groppa M.D., Tomaro M.L. & Benavides M.P., 2007. Polyamines and heavy metal stress: the antioxidant behavior of spermine in cadmium- and copper-treated wheat leaves. Biometals, 20(2), 185-195.
Ha H.C. et al., 1998. The natural polyamine spermine functions directly as a free radical scavenger. Proc. Natl. Acad. Sci., 95, 11140-11145.
Ibrahim A.S.S., Mohamed A., Elbadawi E.Y.B. & Al-Salamah A.A., 2011. Bioreduction of Cr(VI) by potent novel chromate resistant alkaliphilic Bacillus sp. strain KSUCr5 isolated from hypersaline Soda lakes. Afr. J. Biotechnol., 10(37), 7207-7218.
Igarashi K. & Kashiwagi K., 2006. Polyamine modulon in Escherichia coli: genes involved in the stimulation of cell growth by polyamines. J. Biochem., 139, 11-16.
Johnson L. et al., 2012. Surface-localized spermidine protects the Pseudomonas aeruginosa outer membrane from antibiotic treatment and oxidative stress. J. Bacteriol., 194(4), 813-826.
Kwon D.H. & Lu C.-D., 2006. Polyamines increase antibiotic susceptibility in Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 50(5), 1623-1627.
Lauwerys R., Haufroid V., Hoet P. & Lison D., 2007. Toxicologie industrielle et intoxications professionnelles. 5e éd. Issy-les-Moulineaux, France : Elsevier Masson.
Liu J.H. et al., 2007. Polyamines and their ability to provide environmental stress tolerance to plants. Plant Biotechnol., 24, 117-126.
Sayel H. et al., 2012. Cr(VI) reduction by Enterococcus gallinarum isolated from tannery waste-contaminated soil. Ann. Microbiol., 62, 1269-1277.
Sharma S.S. & Dietz K.J., 2006. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot., 57(4), 711-726.
Somasundaram V., Philip L. & Murty Bhallamudi S., 2009. Experimental and mathematical modeling studies on Cr(VI) reduction by CRB, SRB and IRB, individually and in combination. J. Hazard. Mater., 172(2-3), 606-617.
Tahri Joutey N. et al., 2011. Remediation of hexavalent chromium by consortia of indigenous bacteria from tannery waste-contaminated biotopes in Fez, Morocco. Int. J. Environ. Stud., 1, 1-12.
Tahri Joutey N. et al., 2013. Hexavalent chromium removal by a novel Serratia proteamaculans isolated from the bank of Sebou River (Morocco). Environ. Sci. Pollut. Res. Int., 21(4), 3060-3072.
Terui Y. et al., 2004. Enhancement of the synthesis of RpoN, Cra, and H-NS by polyamines at the level of translation in Escherichia coli cultured with glucose and glutamate. J. Bacteriol., 189, 2359-2368.
Thacker U., Parikh R., Shouche Y. & Madamwar D., 2006. Hexavalent chromium reduction by Providencia sp. Proc. Biochem., 41, 1332-1337.
Wortham B.W., Patel C.N. & Oliveira M.A., 2007. Polyamines in bacteria: pleiotropic effects yet specific mechanisms. Adv. Exp. Med. Biol., 603, 106-115.
