


- Accueil
- Volume 18 (2014)
- numéro 4
- Étude des possibilités d’amélioration de la finesse de la fibre de coton (synthèse bibliographique)


Visualisation(s): 4744 (38 ULiège)
Téléchargement(s): 295 (2 ULiège)

Étude des possibilités d’amélioration de la finesse de la fibre de coton (synthèse bibliographique)

Notes de la rédaction
Reçu le 27 janvier 2014, accepté le 8 septembre 2014
Résumé
Description du sujet. Le coton est la première fibre textile naturelle produite dans le monde. Sa qualité doit sans cesse être améliorée pour faire face à la concurrence des fibres synthétiques et pour l’adapter à la constante modernisation des systèmes de filature. Bien qu’elle ne soit pas prise en compte au plan commercial, la finesse de la fibre constitue, avec la longueur, la résistance, la couleur et la maturité, un élément essentiel de la qualité de la fibre de coton. La filature de fibres fines réduit les pertes de production et permet la fabrication de fils résistants, uniformes, brillants et doux au toucher.
Littérature. En agissant sur la sélection variétale, les pratiques culturales, la récolte, l’égrenage et le stockage de la fibre, des améliorations notables ont pu être obtenues sur la longueur, la maturité et la résistance de la fibre. Cependant, les sélectionneurs se sont peu intéressés à la finesse car les premiers travaux sur ce caractère, au contraire d’études plus récentes, avaient conclu à son association avec un rendement réduit. Nous présentons dans cette synthèse les facteurs qui influencent la finesse de la fibre du cotonnier et les possibilités d’améliorer celle-ci en exploitant la variabilité du genre Gossypium.
Conclusion. La finesse de la fibre est influencée par tout facteur perturbant la croissance et le développement de la fibre. Cependant, les facteurs génétiques ont un effet plus important que les facteurs environnementaux sur la finesse. Les différentes espèces du genre Gossypium représentent une source importante de variabilité génétique. Dès lors, l’approche génétique semble la voie la plus intéressante pour l’amélioration de la finesse de la fibre de l’espèce la plus cultivée, G. hirsutum.
Abstract
Study of possible ways to improve cotton fiber fineness. A review.
Description of the subject. Cotton is the most important textile fiber worldwide. Its quality has to be constantly improved to increase its competitive advantage over synthetic fibers and to keep up with the continuous modernization of spinning factories. Even though it is not a property of commercial value, fineness constitutes, along with length, strength, color and maturity, a major trait of fiber quality. The spinning of fine fiber results in reduced losses and the production of strong, uniform, lustrous, and soft-touch yarn.
Literature. Genetic improvement combined with improved cultural practices: harvesting, ginning and storage have led to significant improvements in cotton fiber length, maturity and strength. To date, cotton producers have not usually been interested in fineness because earlier studies, contrary to the more recent ones, have identified its correlation with reduced yield. In this review we present the factors that influence cotton fiber fineness and show possible ways to improve it.
Conclusion. Any factor interfering with fiber growth and fiber development can influence fiber fineness. However, genetic influence is more important than the impact of the environment on fineness. So considering that the other species of the Genus Gossypium represent an important source of genetic variability, the genetic approach seems to be the most beneficial way to improve the fineness of upland cotton fiber.
Table des matières
1. Introduction
1Le genre Gossypium L. regroupe une cinquantaine d’espèces de cotonnier (Fryxell, 1979). Ces espèces représentent une large diversité, notamment au plan morphologique. Ainsi, certaines produisent des graines glabres quand d’autres produisent des graines portant des poils cellulosiques. Chez quatre espèces cultivées de cotonnier, ces poils atteignent une taille suffisante pour être filés. Il s’agit des deux espèces allotétraploïdes, Gossypium hirsutum L. et Gossypium barbadense L. et de deux espèces diploïdes, Gossypium herbaceum L., exploitée en Inde et Gossypium arboretum L., cultivée dans les régions arides d’Afrique et d’Asie. L’espèce G. barbadense est caractérisée par des fibres longues, fines et résistantes, tandis que les espèces asiatiques G. herbaceum et G. arboretum présentent des fibres courtes et grossières. Gossypium hirsutum produit des fibres de qualité intermédiaire avec un rendement au champ élevé. Gossypium hirsutum est la plus importante espèce de cotonnier cultivée, elle fournit 90 % de la production mondiale, tandis que G. barbadense assure 5 % de cette production. La fibre représente à elle seule plus de 95 % de la valeur de la récolte du cotonnier (Cotton Incorporated, 20051 cité par Braden, 2005). Le prix d’achat de la fibre dépend de sa qualité. Avec un marché mondial en pleine expansion et la concurrence des textiles synthétiques, il est nécessaire d’améliorer sans cesse la qualité du coton. Cela passe principalement par l’amélioration de composantes intrinsèques, telles que la finesse, la longueur, la résistance, la maturité et la coloration, et extrinsèques, telles les contaminants. Si un grand nombre de travaux se sont intéressés à l’amélioration de la longueur, de la maturité et de la résistance, en agissant à travers la sélection variétale, les pratiques culturales, la récolte, l’égrenage et le stockage de la fibre, peu de travaux ont concerné la finesse. Cependant, la modernisation des techniques de filature fait de ce caractère un paramètre fondamental à prendre en compte. La filature de fibres fines permet la fabrication de fils résistants et uniformes. Cela s’accompagne d’une augmentation de la productivité par augmentation de la vitesse de la filature et la réduction des pertes de production (May, 1999).
2Dans cette étude, nous passons en revue les différents facteurs qui influencent la finesse de la fibre du cotonnier et les voies et moyens disponibles pour l’améliorer. Nous insisterons particulièrement sur les possibilités qu’offre l’exploitation de la variabilité intra- et interspécifique.
2. La fibre de cotonnier
2.1. Morphologie de la fibre
3Ce qui est appelé fibre chez le cotonnier est un poil unicellulaire présentant l’aspect d’un ruban vrillé dont la longueur peut dépasser 3 cm. La fibre a une section variant de 10 à 30 µm (Roehrich, 1947). Large à sa base, la fibre présente une forme effilée vers son extrémité. La coupe transversale d’une fibre montre une structure en plusieurs couches (Figure 1). De l’extérieur vers l’intérieur, la fibre est formée d’une cuticule constituée de cires recouvrant une paroi primaire mince ou paroi d’étirement composée de pectine, de cellulose, de phosphatides et de lipides disposés en bandes. À cette paroi primaire fait suite une paroi secondaire plus épaisse formée de trois couches de cellulose disposées en spirale. La paroi secondaire est composée de lamelles de 0,25 µm d’épaisseur décomposées en fibrilles. Ces fibrilles sont réparties en microfibrilles de 80 à 150 Å d’épaisseur et 0,25 µm de longueur. Le sens d’enroulement des fibrilles de cellulose autour de l’axe de la fibre peut s’inverser, créant des points de retournements. Le centre de la fibre est constitué du cytoplasme et d’une vacuole renfermant les constituants cellulaires. Au cours de la maturation de la fibre, le cytoplasme et la vacuole se résorbent et laissent un canal : le lumen.
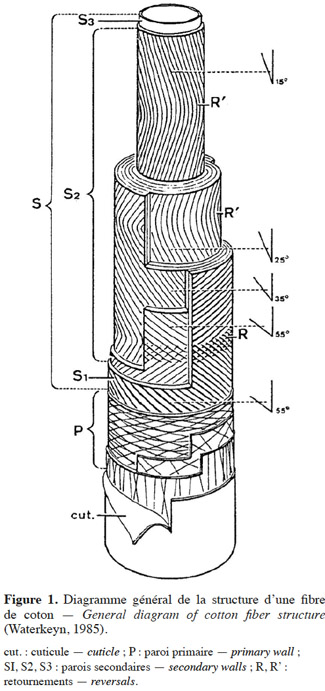
2.2. Développement de la fibre
4Il existe deux types de fibres de coton : des fibres moyennes et longues utilisées pour la filature et des fibres courtes. L’ensemble de ces fibres résulte de l’expansion des cellules épidermiques de la graine de cotonnier (Roehrich, 1947). Les fibres moyennes et longues appelées soies ou lint peuvent dépasser 30 mm de longueur. Les fibres courtes, appelées duvet ou linter font, par définition, moins d’un ½ pouce (soit 1,27 cm) de longueur (Lang, 1938). Le développement des fibres de « lint » est amorcé dès l’ouverture de la fleur, tandis que le développement des fibres de « linter » commence 5 à 10 jours après l’anthèse. Le développement des fibres longues dure 60 jours et se déroule suivant quatre phases qui se chevauchent (Guo et al., 2003). La première phase de développement est l’initiation ; elle commence un jour avant l’anthèse, d’abord du côté de la chalaze, puis elle se poursuit vers le micropyle. Elle se caractérise par le gonflement et l’expansion sphérique de quelques cellules épidermiques de la membrane de la graine. En s’allongeant, les cellules forment un tube cylindrique au-dessus de la surface épidermique. Les fibres initiées se développent de façon isodiamétrique à partir de la surface de l’épiderme et s’allongent de façon anisotropique. Elles sont visibles 24 h après la pollinisation. À l’initiation succède une phase d’élongation des cellules qui commence généralement le jour de l’anthèse et dure 20 jours. Cette étape est déterminée par des facteurs génétiques et environnementaux. À ce stade, la fibre n’est formée que d’une fine cuticule entourant une paroi primaire qui contient le reste de la cellule composée d’une vacuole et d’une paroi cytoplasmique (Seagull et al., 2001). La paroi primaire, alors mince, s’épaissit au cours de cette phase. La fibre, qui a une grande activité métabolique, fait face à un important besoin en nutriments et en eau. La longueur de la fibre augmente de 1 000 à 3 000 fois son diamètre au cours de l’élongation. C’est la pression de turgescence qui conduit l’expansion des cellules, en longueur et en diamètre, laquelle est limitée par l’extensibilité de la paroi primaire. La pression de turgescence est modifiée par le potentiel osmotique qui est déterminé par les concentrations en malate et en K+. La longueur moyenne des fibres varie en fonction de leur position sur la graine : elle décroît lorsqu’on passe de la chalaze vers le micropyle. Les fibres situées au niveau de la chalaze ont un plus large diamètre que les fibres de la région micropylaire (Davidonis et al., 1994). La troisième phase de développement est la phase d’épaississement de la fibre, elle correspond à la formation de la paroi secondaire. Elle débute entre le 16e et le 19e jour après l’anthèse et se poursuit 40 voire 50 jours après l’anthèse (Guo et al., 2003). Au cours de cette étape, des microfibrilles de cellulose se disposent de façon hélicoïdale sur la paroi primaire. Ces dépôts de couches pecto-cellulosiques se produisent avec des inversions de l’angle de dépôt. L’orientation des microfibrilles de cellulose est déterminée par l’orientation des microtubules dans le cytosquelette des cellules (Haigler, 2010). D’abord disposés de façon aléatoire lors de l’initiation, les microfibrilles s’orientent en cerceaux parallèles. La synthèse des microfibrilles des couches les plus basales commence autour de la circonférence de la fibre lors de son allongement. En même temps que se dépose la cellulose, le cytoplasme et la vacuole disparaissent peu à peu. Selon Haigler (2010), les microtubules seraient importants dans le contrôle du périmètre de la fibre. Les microtubules contribuent à restreindre l’expansion des fibres, probablement à cause de leur impact sur l’orientation des microfibrilles de cellulose. Seagull et al. (2000) ont rapporté que la fibre de cotonnier se développe par croissance diffuse. La majorité des cellules qui présente ce type de croissance se développe en longueur et en diamètre. Chez les cellules à croissance diffuse, la production de la paroi secondaire marque la fin de la croissance cellulaire. La paroi secondaire des cellules végétales renferme habituellement, en plus des constituants de la paroi primaire, des insertions de lignine. La lignine est le constituant qui donne à la paroi secondaire sa rigidité. La paroi secondaire des fibres de coton formée essentiellement de cellulose ne contient pas de lignine (Kim et al., 2001). L’absence de lignine dans la paroi de la fibre lui confère sa flexibilité et lui permet de croître à la fois en diamètre et en longueur. En cela, la fibre de cotonnier est un système unique car l’augmentation du diamètre continue pendant sa phase d’épaississement, qui correspond à la synthèse de la paroi secondaire (Seagull et al., 2007). Le diamètre de la fibre n’est pas fixe et maximal à la fin de la phase d’initiation, comme cela a été longtemps admis, il évolue tout au long des trois premières phases de son développement. Les travaux de Seagull et al. (2000) sur quatre variétés de cotonnier (G. hirsutum MD51ne, G. hirsutum DP50, G. hirsutum DP90 et G. barbadense) l’étayent. Ces auteurs ont rapporté à partir de l’analyse du diamètre et de la biréfringence de la paroi que le diamètre augmente avec la croissance de la fibre et le développement de la paroi secondaire. Le diamètre croît jusqu’à 30 jours après l’anthèse pour les quatre génotypes analysés et cette augmentation se poursuit au-delà de 40 jours après l’anthèse pour G. hirsutum DP90 et G. barbadense.
5À la phase d’épaississement fait suite la dernière étape de développement de la fibre : la maturation. Elle est déclenchée par l’ouverture des capsules entre 45 à 60 jours après l’anthèse. Les fibres cylindriques exposées à l’air et au soleil se déshydratent et s’affaissent pour donner des structures vrillées caractéristiques. Des fibres issues d’un même génotype peuvent présenter des finesses différentes, cette variation dépendrait des conditions de croissance, de la position de la capsule, de la position de la graine dans la capsule et de la position de la fibre sur la graine (Davidonis et al., 1994). Les fibres situées au niveau de la chalaze auraient un plus large diamètre que les fibres de la région micropylaire.
3. La finesse de la fibre
3.1. Importance de la finesse de la fibre pour l’industrie textile
6Le tableau 1 reprend, par ordre d’importance et en fonction du système de filature, les principaux caractères technologiques de la fibre. Si, dans les systèmes de filature classique, l’importance de la finesse de la fibre apparait moindre, elle est, dans le système de filature le plus moderne (filature à jet d’air), le caractère primordial à prendre en compte pour la transformation de la fibre en fil. La finesse détermine le nombre de fibres présentes dans la section transversale du fil. Plus les fibres sont fines, plus leur nombre est important dans la section du fil et, par conséquent, plus le fil est résistant. La finesse de la fibre influence à la fois la vitesse de la filature, la résistance, l’uniformité, la brillance, le toucher du fil et la productivité de la filature en réduisant les pertes de production. Les fibres les plus fines produisent par ailleurs des fils et des tissus plus souples et plus agréables au toucher (Lawrence, 2003).
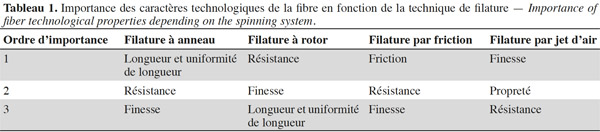
3.2. Mesures de la finesse
7La finesse est un caractère lié au diamètre, à la section transversale, au poids par unité de longueur et à la surface spécifique de la fibre (Haigler, 2010). Elle peut être estimée à l’aide de diverses méthodes directes ou indirectes. Les premières estimations directes de la finesse de la fibre étaient basées sur des méthodes gravimétriques. Pour ce faire, un ensemble de fibres de longueur standard est pesé. Le nombre de fibres constituant le faisceau est évalué au microscope. De ces mesures est déduite la densité linéaire qui est obtenue en divisant le poids du faisceau par le nombre de fibres. Cette mesure représente la masse par unité de longueur exprimée en mg pour 1 000 m ou millitex (Fournier et al., 1991). Le diamètre des fibres peut être évalué en observant la section transversale d’un grand nombre de fibres. Cependant, l’irrégularité de la section des fibres de coton, l’importance du travail et du temps que requièrent les méthodes gravimétriques et microscopiques limitent leur utilisation. De nouvelles méthodes ont vu le jour avec le développement des instruments qui utilisent le principe de la résistance au passage d’un flux d’air pour une estimation indirecte de la finesse. L’aréalomètre développé par Hertel et al. (1951) est un instrument à flux d’air qui donne une estimation de la superficie d’une masse spécifique de fibre, c’est la surface externe des fibres par unité de volume. De cette surface spécifique est déduit, par une formule empirique, le périmètre moyen des fibres. L’expérimentation nécessite 4 à 5 répétitions et est sensible aux variations de l’échantillonnage.
8La mesure de la finesse la plus répandue consiste à mesurer l’indice micronaire avec des appareils à flux d’air. Cet indice est basé sur la résistance à l’air d’un échantillon de fibre de cotonnier. Sur une échelle graduée en fonction des variations du débit d’air ou de la pression, la résistance au passage d’un flux d’air d’un échantillon de fibre de masse connue est estimée par l’appareil de mesure. De cette résistance découle l’indice micronaire qui est proportionnel à la surface spécifique des fibres. Le nombre de fibres pour une masse donnée sera plus élevé pour des fibres fines que pour des fibres épaisses. Un ensemble de fibres fines offrant une plus grande résistance au passage de l’air qu’un ensemble de fibres épaisses, toutes autres caractéristiques égales par ailleurs, il aura un indice micronaire plus faible. De nombreux instruments exploitant le principe de la résistance au flux d’air des fibres ont été développés pour estimer l’indice micronaire. Ces mesures peuvent être faites sur un appareil individuel : AFIS® (Advanced Fibre Information System), FMT (Fineness and Maturity Tester, ASTM D 3818-92, [American Society for Testing and Materials, 1994]), Micromat (Montalvo et al., 2002) ou sur les chaines de mesures intégrées (CMI). L’indice micronaire des fibres de coton peut aussi être évalué en réalisant des analyses thermogravimétriques des fibres et en utilisant la spectroscopie infrarouge par transformation de Fourier (Abidi et al., 2007). Bien que très courante, notamment dans l’industrie textile, l’utilisation de l’indice micronaire comme mesure de la finesse est erronée. En effet, l’indice micronaire mesure simultanément la finesse et la maturité, de sorte qu’un indice micronaire faible peut indiquer la présence de fibres à la fois fines et mûres ou celle de fibres épaisses et immatures. Montalvo (2005) note, dans l’étude des relations entre le micronaire, la finesse et la maturité des fibres de coton, que le micronaire est plus sensible aux variations de l’épaisseur de la fibre qu’à celles de son périmètre. Dans le cas de fibres immatures, l’indice micronaire ne serait pas affecté par le périmètre et sa valeur serait d’autant plus sensible aux variations du périmètre que la fibre est mûre. Les techniques citées ci-dessus donnent toutes une estimation de la finesse à partir de faisceaux de fibres. D’autres outils plus modernes permettent d’établir expérimentalement la finesse de fibres individuelles. Elles s’appuient sur des systèmes d’analyses d’images et de diffusion de la lumière (Buco et al., 1997 ; Huang et al., 2002 ; Aslan et al., 2003 ; Adedoyin et al., 2010 ; Rodgers et al., 2012).
3.3. Facteurs influençant la finesse de la fibre
9La qualité de la fibre d’une plante est certes déterminée par le potentiel génétique de la plante, mais les variations de son environnement et l’interaction du génotype avec cet environnement influencent également celle-ci (Bradow et al., 2010). La qualité peut être affectée par tous les facteurs qui retardent la croissance et le développement du cotonnier : manque d’eau, d’ensoleillement, de nutriments. Contrairement à la finesse, la longueur, la ténacité et l’uniformité de longueur peuvent être modifiés pendant la récolte mécanique du coton-graine, l’égrenage et le peignage de la fibre. Comme rappelé ci-dessus, les 40 premiers jours du développement de la fibre sont déterminants pour sa finesse parce qu’elle croît en diamètre durant toute cette période. La croissance se déroule selon un mécanisme de turgescence cellulaire qui requiert une disponibilité de potassium, d’eau et de chaleur. L’indisponibilité d’un de ces éléments conduit à un moindre développement de la fibre. La disponibilité en eau est particulièrement déterminante. Le stress hydrique de la plante affecte la photosynthèse au cours de laquelle est produit le glucose qui est converti en amidon, en lignine et en cellulose. Cette dernière est le constituant majeur de la fibre de coton. Tout effet sur la photosynthèse se répercute sur la production de cellulose et, par conséquent, sur la formation de la paroi secondaire et influence donc la maturité et le diamètre de la fibre.
10Bauer et al. (1998) ont observé que les fibres issues de cotonniers plantés tardivement ont un micronaire réduit par rapport aux fibres issues de plantes cultivées en période normale. Une insolation insuffisante durant la synthèse de la paroi secondaire altère l’accumulation de cellulose et réduit la maturité de la fibre et le micronaire (Wang et al., 2005). Les mécanismes qui gouvernent l’expansion cellulaire sont donc impliqués dans la détermination du diamètre.
11De nombreux caractères quantitatifs présentent des interactions entre les effets génétiques (G) et environnementaux (E). La présence d’interactions GxE signifie que l’environnement peut influencer de façon variable l’expression d’un caractère en activant ou désactivant des gènes contrôlant ce caractère et en modifiant leur niveau d’expression. Pour Aktar et al. (2008), l’effet de l’environnement sur l’héritabilité de la finesse et l’effet de la sélection sur ce caractère seraient non significatifs. Selon May (1999), l’analyse statistique de mesures de la finesse estimée par la densité linéaire et l’AFIS indique une plus grande influence des facteurs génétiques sur la finesse. Cependant, d’autres études remettent en cause la prédominance du contrôle génétique de la finesse. Campbell et al. (2005) rapportent que l’influence des facteurs environnementaux sur la mesure de la finesse en termes de micronaire est plus importante que celle des facteurs génétiques. La variation de l’importance du contrôle génétique de la finesse entre les différents travaux cités ci-dessus peut s’expliquer par le fait que chaque étude concernait du matériel végétal et des environnements différents. Récemment, Zhao et al. (2013) ont proposé un modèle de prédiction de la finesse, de la maturité et de l’indice micronaire de la fibre de G. hirsutum.
4. Amélioration génétique de la finesse
4.1. Déterminisme génétique de la finesse
12Les caractères quantitatifs comme la finesse de la fibre sont contrôlés par de nombreux gènes. Plus le nombre de gènes gouvernant un caractère est élevé, plus la probabilité est faible d’isoler des individus présentant les allèles intéressants pour tous les loci.
13L’une des exigences liées à l’étude du déterminisme d’un caractère quantitatif est l’estimation de son héritabilité qui renseigne sur le degré d’influence probable des facteurs génétiques. Une faible héritabilité signifie une faible variation génétique dans une population et/ou une forte variation de l’environnement. Les caractères hautement héritables sont moins difficiles à améliorer que les caractères à faible héritabilité. Les travaux de Bishr2 (1954) cité par May (1999) sur des cultivars de G. hirsutum ont montré que le périmètre et l’épaississement de la paroi de fibres mesurés par l’aréalomètre sont modérément transmissibles avec une héritabilité respective de 0,51 et 0,43. Les mêmes conclusions ont été faites par Aktar et al. (2008) à propos de la surface de la fibre. Liu et al. (2011) ont trouvé au sein d’une population issue du croisement entre G. hirsutum variété Guazuncho 2 (GUA) et G. barbadense accession VH8-4602 (VH8) une héritabilité de 0,4 pour le micronaire et de 0,3 pour le périmètre. Pour Ulloa (2006), l’héritabilité de la densité linéaire et du micronaire est de 0,6 et 0,58 chez des populations F2 issues du croisement entre les cultivars FiberMax 832 et MD51ne de G. hirsutum. Il ressort de ces études que la finesse estimée en termes d’indice micronaire ou de densité linéaire est modérément à hautement transmissible (May, 1999 ; Aktar, 2008). La majorité des caractères quantitatifs ne sont pas indépendants les uns des autres. Selon Frelichowski et al. (2006) et An et al. (2010), l’indice micronaire est positivement corrélé au rendement et à la ténacité et négativement corrélé à la longueur de la fibre. Le périmètre est corrélé positivement à la longueur de la fibre et à la ténacité (Ulloa et al., 2000). Selon Zhang et al. (2011), la finesse mesurée en termes d’indice micronaire serait liée à la longueur et à l’uniformité de longueur. Selon ces auteurs, il existe une corrélation entre les QTL liés à l’élongation et ceux liés à la finesse. De nombreux écrits ont rapporté que la finesse est sous le contrôle de gènes additifs. May (1999) relate que l’indice micronaire et la surface spécifique de la fibre, estimés par l’aréalomètre, sont influencés par une variance génétique additive. Wu et al. (2009) et Zhang et al. (2012) ont également conclu que la finesse est sous le contrôle de gènes additifs. À l’opposé, pour Ahuja et al. (2007), le micronaire est sous l’influence de gènes non additifs. An et al. (2010) ont trouvé que la finesse dépend de l’action de gènes dominants tandis que pour Kohel et al. (2001), ce caractère est sous l’influence de gènes récessifs. Selon Wang et al. (2007), l’effet des gènes sur la finesse est à la fois additif et dominant. Il apparait que selon le type d’instruments utilisés pour son estimation, les composants du déterminisme de la finesse de la fibre sont variables.
4.2. Amélioration génétique
14Les méthodes d’amélioration conventionnelles basées sur la sélection phénotypique ont permis d’améliorer la qualité de la fibre. C’est ainsi que la domestication, la sélection massale et la sélection généalogique, entre autres, ont permis d’obtenir des cotonniers présentant des fibres longues résistantes, blanches, brillantes et fines. Ces processus ont cependant réduit la diversité génétique existante par la perte de gènes intéressants (May, 1999). L’ensemble des espèces du genre Gossypium constitue un stock de variabilité génétique considérable pour l’amélioration de la fibre. Ces espèces originaires d’un même phylum sont classées au sein de neuf groupes génomiques. Les espèces diploïdes (2n = 2x = 26) appartiennent aux génomes A, B, C, D, E, F, G ou K et les espèces tétraploïdes (2n = 4x = 52) du genre sont attribuées au génome AD (Ndungo et al., 1988). En fonction de la perméabilité génétique qui existe entre leur génome et celui de G. hirsutum, les espèces de cotonnier sont classées en trois catégories : les réservoirs primaire, secondaire et tertiaire (Stewart, 1995). Le pool génique primaire est constitué de toutes les formes cultivées subspontanées et sauvages de G. hirsutum et de G. barbadense, ainsi que des trois autres espèces tétraploïdes (Gossypium tomentosum Nutt. ex Seem., Gossypium mustelinum Miers ex G.Watt et Gossypium darwinii G.Watt). Les croisements avec ces espèces sont faciles et les fréquences de recombinaison sont élevées. La diversité génétique qu’offrent les cultivars de G. hirsutum est considérée comme très étroite. Le pool génétique secondaire comprend les espèces diploïdes de génome A, D, B ou F (Stewart, 1995). Le réservoir génétique tertiaire renferme les espèces des génomes E, C, G ou K. L’utilisation de ces espèces est plus difficile à cause de la nécessité de doubler le nombre chromosomique des formes hybrides intermédiaires (diploïdes ou triploïdes) et surtout en raison de la faiblesse des taux de recombinaison avec les chromosomes de l’espèce tétraploïde. Vu l’étroitesse de la variabilité existant au sein de G. hirsutum, les améliorateurs ont rapidement fait appel à G. barbadense, l’autre espèce cultivée de cotonnier tétraploïde. Les premiers travaux d’amélioration de G. hirsutum par hybridation interspécifique avaient pour but d’associer les qualités technologiques de la fibre de G. barbadense aux qualités agronomiques de G. hirsutum. Cependant, même si ces deux espèces s’hybrident facilement, il est généralement observé un retour rapide vers le parent récurent (Schwendiman, 1978). Lacape et al. (2010) ont noté que des lignées recombinantes issues du croisement entre G. hirsutum et G. barbadense présentaient des caractères technologiques intermédiaires entre les deux parents, mais plus proches de G. hirsutum que de G. barbadense.
15Les espèces diploïdes sauvages représentent une importante source potentielle de variabilité. Quoique ces espèces diploïdes produisent des fibres impropres à la filature, elles possèdent des caractères d’intérêt pouvant être transmis à la principale espèce de cotonnier cultivé pour l’amélioration de sa fibre. En effet, la majorité des espèces diploïdes peuvent être croisées avec G. hirsutum en contournant les barrières d’incompatibilité qui existent entre ces espèces et le cotonnier cultivé. Deux principales voies sont utilisées : une voie tri-spécifique indirecte et une voie bi-spécifique directe (Mergeai, 2006). La voie bi-spécifique ou méthode aphylétique d’introgression aboutit à la formation de lignées monosomiques d’addition qui ne contiennent qu’un seul chromosome de l’espèce donneuse. Cette voie consiste à réaliser un croisement direct de G. hirsutum avec une espèce diploïde porteuse de caractères d’intérêt. La voie tri-spécifique consiste à utiliser une espèce pont appartenant au génome A ou D qui est croisée soit avec l’espèce donneuse pour créer un tétraploïde artificiel, soit avec l’espèce cultivée pour créer un allohexaploïde transitoire. La première stratégie est qualifiée de voie paraphylétique d’introgression et la seconde de voie pseudophylétique d’introgression. Toutes les méthodes qui visent à exploiter une espèce diploïde passent par la création d’un hybride stérile (diploïde ou triploïde) dont il faut rétablir la fertilité par doublement de son nombre chromosomique. Le choix du schéma d’hybridation interspécifique dépend du déterminisme génétique du caractère à introgresser, sachant que la voie bi-specifique est plus appropriée pour l’introgression de caractères monogéniques, tandis que la voie trispécifique convient mieux à l’introgression de gènes localisés sur plusieurs chromosomes (Mergeai et al., 2008). Le croisement direct des espèces diploïdes requis par la méthode paraphylétique est restreint à l’utilisation d’espèces donneuses du génome A ou D et, dans une moindre mesure, aux espèces de génome B ou F. L’obtention d’un hybride fertile est plus difficile lorsque les espèces diploïdes des autres génomes sont croisées avec une espèce appartenant au génome A ou D. La voie pseudophylétique permet de contourner la difficulté du croisement direct des deux espèces diploïdes. À ce jour, très peu d’études ont porté sur l’utilisation d’espèces diploïdes pour l’amélioration de la finesse de G. hirsutum. Demol et al. (1976) et Demol et al. (1978) ont étudié l’influence des génomes des espèces sauvages sur la fibre de G. hirsutum en estimant les caractères technologiques d’hybrides allohexaploïdes issus du croisement entre cotonnier cultivé et cotonniers sauvages. Il en ressort que chez les hybrides interspécifiques comparés, les espèces sauvages Gossypium anomalum Wawra & Peyr., Gossypium stocksii Mast., Gossypium areysianum Deflers et Gossypium longicalyx J.B.Hutch. & B.J.S.Lee améliorent la longueur de la fibre, tandis que Gossypium thurberi Tod., Gossypium karknesii Brandegee, Gossypium raimondii Ulbr. et Gossypium aridum (Rose & Standl.) Skovst. améliorent sa résistance. Gossypium longicalyx et G. thurberi améliorent la finesse de la fibre. Les hexaploïdes obtenus avec ces espèces produisent des fibres plus fines que celles de G. barbadense. Mergeai et al. (2010) ont observé une réduction drastique de la largeur du ruban chez l’hexaploïde (G. hirsutum x G. longicalyx)² et l’hybride ([G. hirsutum x G. thurberi]² x G. longicalyx), ainsi que chez des plantes obtenues par back-cross BC1 et BC2 issues de l’hybride trispécifique. Ces observations prouvent que G. longicalyx est une source intéressante pour l’amélioration de la finesse de G. hirsutum.
16Une fois le matériel hybride obtenu, l’ADN étranger peut être identifié dans le fond génétique de G. hirsutum à l’aide d’observations morphologiques, cytogénétiques et moléculaires. Les marqueurs moléculaires sont un moyen de choix pour le suivi de l’introgression. Ils permettent de mettre en évidence la position de loci liés à des caractères quantitatifs appelés QTL (Quantitative Trait Loci) sur une carte génique et de suivre ces QTL tout au long de la sélection. Les travaux visant l’identification de QTL liés à la qualité de la fibre se sont multipliés au cours des dernières années. Des QTL ont été mis en évidence chez des populations issues de croisements impliquant les espèces tétraploïdes G. hirsutum, G. barbadense et G. tomentosum (Tableau 2). Les QTL identifiés contrôlent de 4,5 à 43,9 % de la variation phénotypique, suggérant que la finesse serait contrôlée par des gènes mineurs et majeurs. La majorité de ces QTL a de petits effets car ils contrôlent moins de 20 % de la variation phénotypique. Certains QTL ont été détectés dans les mêmes régions chromosomiques. Il peut s’agir d’un unique QTL à effet pléiotropique ou de deux QTL génétiquement liés. Zhang et al. (2005) ont détecté la présence de QTL liés au micronaire, au pourcentage de fibre, à la résistance et à l’élongation sur le chromosome 5. Des QTL liés à l’élongation de la fibre (Shen et al., 2005 ; Chen et al., 2009), à la longueur de la fibre (Shen et al., 2005), au micronaire (Lin et al., 2005 ; Shen et al., 2005 ; He et al., 2007) et à la ténacité (Chen et al., 2009) ont été identifiés sur les mêmes régions des chromosomes 23 et 24. Un certain nombre de QTL associés à la qualité de la fibre ont été isolés sur le sous-génome D (Rong et al., 2007). Bien que les espèces du génome D produisent des fibres courtes et ne soient pas cultivées, ce génome contiendrait des gènes contrôlant les caractères technologiques de la fibre (Jiang et al., 1998). Xu et al. (2012) rapportent la localisation de deux QTL associés au chromosome C7 au sein de populations issues du croisement entre G. hirsutum et Gossypium klotzschianum Andersson, une espèce diploïde sauvage du génome D. Ces QTL qFMIC-7-1 et qFMIC-7-2 situés à proximité du marqueur SSR NAU1362 ont pour effet de réduire le micronaire. Après la détection des QTL associés à la finesse de la fibre, il s’agit par la suite de créer par croisement un génotype portant tous les allèles favorables à l’expression de ce caractère.
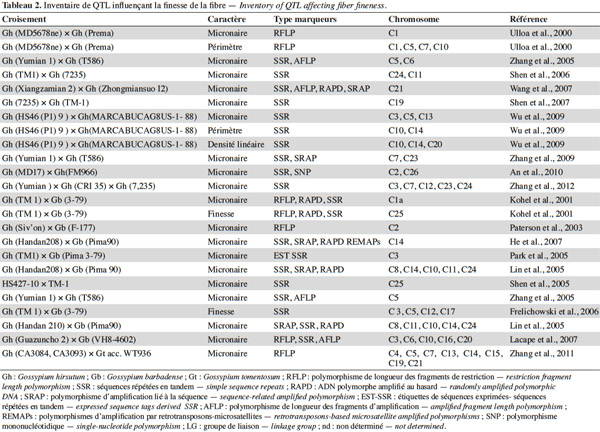
5. Conclusions
17La finesse est un composant essentiel de la qualité de la fibre de cotonnier. Elle est évaluée à l’aide de méthodes directes et indirectes qui l’estiment en termes de densité linéaire, de diamètre, de micronaire, etc. La mesure la plus utilisée est l’indice micronaire qui est une évaluation composite de la finesse et de la maturité. Qu’il s’agisse de densité linéaire, de diamètre ou de micronaire, de nombreuses études ont montré que la finesse présente une hérédité modérée à élevée et est transmissible par sélection. Ce caractère n’est pas uniquement déterminé par les facteurs génétiques, comme cela avait été longtemps considéré, mais est également influencé par les conditions environnementales qui prévalent pendant la croissance et le développement de la fibre.
18Bien que le déterminisme génétique de la finesse de la fibre soit à ce jour inconnu, de nombreuses études ont détecté des QTL liés à ce caractère sur des chromosomes différents (1 à 8 chromosomes, selon les études). Ceci indique que la finesse de la fibre, comme tous les caractères quantitatifs, seraient sous le contrôle de plusieurs gènes. Dans ce contexte, les méthodes pseudophylétique et paraphylétique d’introgression sont les plus pertinentes pour le transfert de caractères polygéniques. Les espèces sauvages G. longicalyx, G. raimondii, G. anomalum sont potentiellement exploitables pour le transfert de gènes susceptibles d’améliorer la finesse de la fibre chez G. hirsutum. Toutes ces espèces présentent des affinités relativement importantes vis-à-vis des chromosomes des génomes A (G. anomalum et G. longicalyx) et D (G. raimondii et G. thurberi) constitutifs de l’amphidiploïde cultivé.
191 Cotton Incorporated, 2005. Personal communication with Roy Cantrell.
Bibliographie
Abidi N. & Hequet E., 2007. Characterization of cotton fibers using TGA and FTIR. In: Proceedings of the World Cotton Research Conference, September 10-14, 2007, Lubbock, Texas.
Adedoyin A. & Li C., 2010. Single cotton fiber diameter determination by using Fraunhofer diffraction. In: Proceedings of Beltwide Cotton Conferences, January 4-7, 2010, New Orleans, Louisiana. Memphis, TN, USA: National Cotton Council of America.
AhujaS.L. & Dhayal L.S., 2007. Combining ability estimates for yield and fibre quality traits in 4 x 13 line x tester crosses of Gossypium hirsutum. Euphytica, 153, 87-98.
Akhtar M.M., Azhar F.M. & Ali Z., 2008. Genetic basis of fiber quality attributes in upland cotton (Gossypium hirsutum) germplasm. Int. J. Agric. Biol., 10(2), 217-220.
American Society for Testing and Materials, 1994. Standard test method for linear density and maturity index of cotton fibers (IIC-Shirley Fineness/Maturity Tester). In: Annual Book of ASTM Standards, 07.02: ASTM Standard D 3818-92, 143-146.
An C. et al., 2010. Use of fiber and fuzz mutants to detect QTL for yield components, seed, and fiber traits of upland cotton. Euphytica, 172(1), 21-34.
Aslan M., Yamada J., Menguëc M. & Thomasson J.A., 2003. Characterization of individual cotton fibers via light-scattering experiments. J. Thermophys. Heat Transfer, 17(4), 442-449.
Bauer P.J., May O.L. & Camberato J.J., 1998. Planting date and potassium fertility effects on cotton yield and fiber properties. J. Prod. Agric., 11, 415-420.
Bradow J.M. & Davidonis G.H., 2010. Effects of environment on fiber quality. In: Stewart J. McD., Oosterhuis D., Heitholt J.J. & Mauney J.R., eds. Physiology of cotton. Dordrecht, The Netherlands: Springer, 229-245.
Bragg C.K. & Shofner F.M., 1993. A rapid, direct measurement of short fiber content. Text. Res. J., 63(3), 171-176.
Buco S.M. et al., 1997. Fast determination of maturity and fineness by NIR with a diode-array HVI. Part 1. Data analysis. In: Proceedings of the Beltwide Cotton Conferences, January 9-12, 1996, Nashville, USA. Memphis, TN, USA: National Cotton Council of America, 552-553.
Campbell B.T. & Jones M.A., 2005. Assessment of genotype environment interactions for yield and fiber quality in cotton performance trials. Euphytica, 144(1-2), 69-78.
Chen H. et al., 2009. Using three overlapped RILs to dissect genetically clustered QTL for fiber strength on Chro.D8 in upland cotton. Theor. Appl. Genet., 119, 605-612.
Davidonis G. & Hinojosa O., 1994. Influence of seed location on cotton fiber development in planta and in vitro. Plant Sci., 203, 107-113.
Demol J., Verschraege L. & Maréchal R., 1976. Utilisation des espèces sauvages en amélioration cotonnière. Caractéristiques technologiques des formes allo-hexaploïdes. Coton Fibres Trop., 31(4), 381-389.
Demol J., Verschraege L. & Maréchal R., 1978. Utilisation des espèces sauvages en amélioration cotonnière. Observations sur les caractéristiques technologiques des nouvelles formes allohexaploïdes. Coton Fibres Trop., 33(3), 327-333.
Deussen H., 1992. Improved cotton fiber properties - the textile industry’s key to success in global competition. In: Proceedings of the Conference Cotton fiber cellulose: structure, function, and utilization, October 28-31, 1992, Savannah, GA, USA, 43-63.
Fournier J. & Gutknecht J., 1991. Variabilité et héritabilité de la finesse standard Hs de la fibre de 6 cultivars de coton d'Afrique francophone. Coton Fibres Trop., 46(2), 143-158
Frelichowski Jr J.E., Palmer M.B. & Main D., 2006. Cotton genome mapping with new microsatellites from Acala ‘Maxxa’ BAC-ends. Mol. Genet. Genomics, 275(5), 479-491.
Fryxell P.A., 1979. The natural history of the cotton tribe. College Station, TX, USA: Texas A&M University Press.
Guo W., Sun J. & Zhang T., 2003. Gene cloning and molecular breeding to improve fiber qualities in cotton. Chin. Sci. Bull., 48(8), 709-717
Haigler C.H., 2010. Physiological and anatomical factors determining fiber structure and utility. In: Stewart J. McD., Oosterhuis D., Heitholt J.J. & Mauney J.R., eds. Physiology of cotton. Dordrecht, The Netherlands: Springer, 33-47.
He D.H. et al., 2007. QTL mapping for economic traits based on a dense genetic map of cotton with PCR-based markers using the interspecific cross of Gossypium hirsutum × Gossypium barbadense. Euphytica, 153, 181-197.
Hertel K.L. & Craven C.J., 1951. Fineness and immaturity as measured by the arealometer. Text. Res. J., 21(11), 765-774.
Huang Y. & Xu B., 2002. Image analysis for cotton fibers. Part I: longitudinal measurements. Text. Res. J., 72(8), 713-720.
Jiang C., Wright R.J., El-Zik K.M. & Paterson A.H., 1998. Polyploid formation created unique avenues for response to selection in Gossypium (cotton). Proc. Natl. Acad. Sci. U.S.A., 95, 4419-4424.
Kim H.J. & Triplett B.A., 2001. Cotton fiber growth in planta and in vitro. Models for plant cell elongation and cell wall biogenesis. Plant Physiol., 127, 1361-1366.
Kohel R.J., Yu J., Park Y.H. & Lazo G.R., 2001. Molecular mapping and characterization of traits controlling fiber quality in cotton. Euphytica, 121(2), 163-172.
Lacape M., Nguyen T.-B., Hau B. & Giband M., 2007. Targeted introgression of cotton fibre quality quantitative trait loci using molecular markers. In: Guimarães E. et al., eds. Market-assisted selection: current status and future perspectives in crops, livestock, forestry and fish. FAO: Roma, 67-80.
Lacape J.M. et al., 2010. Meta-analysis of cotton fiber quality QTL across diverse environments in an interspecific Gossypium hirsutum x G. barbadense RIL population. BMC Plant Biol., 10(132).
Lang A.G., 1938. The origin of lint and fuzz hair of cotton. J. Agric. Res., 56(7), 507-521.
Lawrence C.A., 2003. Fundamentals of spun yarn technology. Boca Raton, FL, USA: CRC Press.
Lin Z. et al., 2005. Linkage map construction and mapping QTL for cotton fibre quality using SRAP, SSR and RAPD. Plant Breeding, 124, 180-187.
Liu S. et al., 2011. Heritability and predicted selection response of yield components and fibre properties in an inter-specific derived RIL population of cotton. Euphytica, 178, 309-320.
May O.L., 1999. Genetic variation in fiber quality. In: Basra A.S., ed. Cotton fibers - Developmental biology, quality improvement, and textile processing. New York, USA: Food Products Press Imprint of Haworth Press, 183-229.
Mergeai G., 2006. Introgressions interspécifiques chez le cotonnier. Cah. Agric., 15, 135-143.
Mergeai G., Sarr D., Diouf F.H. & Baudoin J.-P., 2008. Comparison of the efficacy of the bispecific and trispecific pathways to exploit Australian wild diploid species for the genetic improvement of upland cotton. In: Proceedings of the Beltwide Cotton Conferences, January 8-11, 2008, Nashville, Gaylord Opryland Resort and Convention Center, Tennessee. Memphis, TN, USA: National Cotton Council of America.
Mergeai G., Baudoin J.-P. & Konan O.N., 2010. Improvement of upland cotton through interspecific hybridization: analyze of the fibre fineness of bi- and trispecific hybrids involving G. hirsutum. In: Proceedings of the Beltwide Cotton Conferences, January 4-7, 2010, New Orleans. Memphis, TN, USA: National Cotton Council of America.
Montalvo J.G. Jr., 2005. Relationship between micronaire, fineness and maturity. Part I. Fundamentals. J. Cotton Sci., 9, 81-88.
Montalvo J.G. Jr., Faught S.E. & Buco S.M., 2002. Sensitivity of the Shirley developments Ltd. Micromat tester to operators and sample preparation. J. Cotton Sci., 6(3), 133-142.
Ndungo V., Demol J. & Maréchal R., 1988. Lʼamélioration du cotonnier Gossypium hirsutum L. par hybridation interspécifique : 3. Application et résultats obtenus. Bull. Rech. Agron. Gembloux, 23(3), 283-316.
Park Y.H., Alabady M.S. & Ulloa M., 2005. Genetic mapping of new cotton fiber loci using EST-derived microsatellites in an interspecific recombinant inbred line cotton population. Mol. Genet. Genomics, 274(4), 428-441.
Paterson A.H. et al., 2003. QTL analysis of genotype × environment interactions affecting cotton fiber quality. Theor. Appl. Genet., 106(3), 384-396.
Rodgers J.E., Delhom C., Fortier C. & Thibodeaux D., 2012. A rapid measurement for cotton breeders of maturity and fineness from developing and mature fibers. Text. Res. J., 82(3).
Roehrich O., 1947. Méthode générale d'étude des caractères technologiques des fibres textiles végétales (1ère partie). Coton Fibres Trop., 2(2), 37-50.
Rong J. et al., 2007. Meta-analysis of polyploid cotton QTL shows unequal contributions of subgenomes to a complex network of genes and gene clusters implicated in lint fiber development. Genetics, 176(4), 2577-2588.
Schwendiman J., 1978. L’amélioration du cotonnier Gossypium hirsutum par hybridation interspécifique : utilisation des espèces G. barbadense et G. stocksii. Coton Fibres Trop., 33, 239-255.
Seagull R.W. et al., 2000. Cotton fiber growth and development. 2. Changes in cell diameter and wall birefringence. J. Cotton Sci., 4, 97-104.
Seagull R.W. & Alspaugh P., 2001. Cotton fibre development and processing. Lubbock, TX, USA: Cotton Incorporated, International Textile Center, Texas Tech University.
Seagull R.W., Russell C. & Gould P., 2007. Effects of exogenous application of GA3 on fiber morphology and secondary wall synthesis. In: Proceedings of the World Cotton Research Conference-4, September 10-14, 2007, Lubbock, Texas, USA. Washington, DC: International Cotton Advisory Committee.
Shen X. et al., 2005. Molecular mapping of QTL for fiber qualities in three diverse lines in upland cotton using SSR markers. Mol. Breeding, 15, 169-181.
Shen X. et al., 2006. Mapping fiber and yield QTLs with main, epistatic, and QTL x environment interaction effects in recombinant inbred lines of upland cotton. Crop Sci., 46(1), 61-66.
Shen X. et al., 2007. Genetic mapping of quantitative trait loci for fiber quality and yield trait by RIL approach in upland cotton. Euphytica, 155(3), 371-380.
Stewart J.M., 1995. Potential for crop improvement with exotic germplasm and genetic engineering. In: Constable G.A. & Forrester N.W., eds. Proceedings of the World Cotton Research Conference-1, Challenging the future, February 14-17, 1994, Brisbane, Australia. Melbourne, Australia: CSIRO, 313-327.
Ulloa M., 2006 . Heritability and correlations of agronomic and fiber traits in an okra-leaf upland cotton population. Crop Sci., 46, 1508-1514.
Ulloa M. & Meredith Jr W.R., 2000. Genetic linkage map and QTL analysis of agronomic and fibre quality traits in an intraspecific population. J. Cotton Sci., 4, 161-170.
Wang B. et al., 2007. QTL analysis and epistasis effects dissection of fiber qualities in an elite cotton hybrid grown in second generation. Crop Sci., 47(4), 1384-1392.
Wang Q.C. et al., 2005. Effects of shading at blossoming and boll-forming stages on cotton fiber quality. Chin. J. Appl. Ecol., 16, 1465-1468 (en chinois).
Waterkeyn L., 1985. Light microscopy of the cotton fibre. In: Cotton fibres: their development and properties. A technical monograph from the Belgian Cotton Research Group. Manchester, UK: International Institute for Cotton, 17-22.
Wu J. et al., 2009. Quantitative analysis and QTL mapping for agronomic and fiber traits in an RI population of upland cotton. Euphytica, 165(2), 231-245.
Xu P. et al., 2012. Molecular mapping and identification of QTLs for fiber micronaire on chromosome 7 from Gossypium klotzschianum. Acta Agron. Sinica, 38(03), 447-453 (en chinois).
Zhang K., Zhang J., Ma J. & Liu S.T., 2012. Genetic mapping and quantitative trait locus analysis of fiber quality traits using a three-parent composite population in upland cotton (Gossypium hirsutum L.). Mol. Breeding, 29(2), 335-348.
Zhang Z. et al., 2011. QTL alleles for improved fiber quality from a wild Hawaiian cotton, Gossypium tomentosum. Theor. Appl. Genet., 123(7).
Zhang Z.S., Xiao Y.H. & Luo M., 2005. Construction of a genetic linkage map and QTL analysis of fiber-related traits in upland cotton Gossypium hirsutum L. Euphytica, 144(1-2), 91-99.
Zhang Z.S., Hu M.C. & Zhang J., 2009. Construction of a comprehensive PCR-based marker linkage map and QTL mapping for fiber quality traits in upland cotton (Gossypium hirsutum L.). Mol. Breeding, 24(1), 49-61.
Zhao W.-Q. et al., 2013. Modeling fiber fineness, maturity, and micronaire in cotton (Gossypium hirsutum L.). J. Integr. Agric., 12(1), 101-108.
Notes
2 Bishr M.A., 1954. Inheritance of perimeter and wall thickness of fiber in a cross between two varieties of upland cotton. Ph.D. dissertation: Baton Rouge, Louisiana State University (USA).
