


- Accueil
- volume 9 (2005)
- Observations préliminaires de la variabilité entre quelques morphotypes de voandzou (Vigna subterranea L. Verdc., Fabaceae) de Côte d’Ivoire


Visualisation(s): 9639 (62 ULiège)
Téléchargement(s): 483 (5 ULiège)

Observations préliminaires de la variabilité entre quelques morphotypes de voandzou (Vigna subterranea L. Verdc., Fabaceae) de Côte d’Ivoire

Notes de la rédaction
Reçu le 15 avril 2004, accepté le 26 mai 2005
Résumé
Le voandzou (Vigna subterranea L. Verdc.) est une légumineuse alimentaire cultivée principalement par les femmes pour lesquelles il assure une source de revenus pour le ménage. En Côte d’Ivoire, la culture du voandzou est localisée à l’ouest et dans la partie nord du pays. Ces zones sont caractérisées par des conditions agroécologiques contrastées incluant la forêt tropicale humide et la savane plus sèche. Dans ces zones, le voandzou joue un rôle important dans l’alimentation et la culture des populations. Quatre morphotypes de Côte d’Ivoire (ICU, BPR, RBU, NFU) ont été utilisés dans un essai préliminaire pour évaluer la variabilité phénotypique entre morphotypes. Pour chaque morphotype, 100 individus ont été utilisés pour analyser 26 caractères agromorphologiques sélectionnés parmi la liste des descripteurs du voandzou. Les résultats de l’analyse statistique ont montré une importante variabilité entre les morphotypes, suggérant ainsi que 22 des caractères peuvent servir valablement à distinguer la variabilité entre les morphotypes de voandzou de Côte d’Ivoire. Trois morphotypes (ICU, BPR et RBU) ont une phase reproductive plus courte que l’autre (NFU). Les morphotypes à phase reproductive courte ont donné un pourcentage plus élevé de gousses matures (87 à 95 %), par comparaison avec le morphotype NFU (69 %) dans nos conditions d’expérimentation. Le morphotype ICU est particulièrement précoce, atteignant la maturité 90 jours après semis (JAS), tandis que le morphotype à phase reproductive longue (NFU) nécessite 137 jours. A partir des caractères agronomiques analysés, les possibilités d’amélioration du rendement du voandzou et la vulgarisation de sa culture en Côte d’Ivoire sont discutées.
Abstract
Preliminary observations of variability between some morphotypes of bambara groundnut (Vigna subterranea L. Verdc., Fabaceae) from Côte d’Ivoire. Bambara groundnut (Vigna subterranea L. Verdc.), is a food legume mainly cultivated by women for whom it represents a source of income for the household. In Côte d’Ivoire, the cultivation of bambara groundnut is located in the western and northern parts of the country. These zones are characterised by contrasted agroecology including tropical rain forest and dry savanna. In these zones, bambara groundnut plays a key role in both food and culture of peoples. Four morphotypes of Côte d’Ivoire (ICU, BPR, RBU, NFU) were used in a preliminary study to assess the phenotypic variability between morphotypes. For each morphotype, 100 individuals were sampled to analyse 26 agromorphological traits selected from the list of bambara groundnut descriptors. Results of statistical analyses showed an important variability among morphotypes suggesting that 22 of these characters could be powerful to distinguish diversity among bambara groundnut morphotypes of Côte d’Ivoire. Three morphotypes (ICU, BPR and RBU) show a shorter reproductive cycle than the other (NFU). In our experimental conditions, morphotypes with a shorter reproductive cycle give a higher percentage of matured pods (87 to 95%), compared to morphotype NFU (60%). The morphotype ICU was particularly earlier, maturing 90 days after sowing (DAS), whereas the long reproductive cycle morphotype (NFU) required about 137 days. Based on the analysed agronomic traits, possibilities to improve bambara groundnut yield and to promote its cultivation in Côte d’Ivoire are discussed.
Table des matières
1. Introduction
1Pour assurer leur développement, de nombreux pays africains ont massivement investi dans la promotion des cultures industrielles au détriment des cultures vivrières dont certaines sont à l’abandon et menacées de disparition (Aké Assi, 1988). Les plantes alimentaires marginales ont de faibles rendements mais elles sont bien adaptées à leur milieu de culture et sont très rustiques (Altieri, 1986). De plus, elles revêtent, souvent, pour les populations qui les cultivent, un important aspect social et culturel.
2Le voandzou, [Vigna subterranea (L.) Verdc.], encore appelé pois bambara ou pois de terre, est une légumineuse alimentaire mineure dont la culture est restée dans les limites de l’Afrique sub-saharienne où il est adapté à diverses conditions climatiques et écologiques (steppe, savane et forêt). Les graines servent à l’alimentation humaine, seules ou mélangées à d’autres aliments. Les feuilles, riches en phosphore, servent à l’alimentation du bétail. Les graines et les feuilles sont utilisées dans la médecine traditionnelle (Nacoulma-Ouédraogo, 1996). Le voandzou est une plante hautement calorique (387 kcal/100 g), riche en vitamines et en éléments minéraux et très équilibrée en protéines (Ucciani, Busson, 1963 ; Oliveira, 1976 ; Oniwamo et al., 1998 ; Minka, Bruneteau, 2000). En dépit de ces nombreux avantages, le voandzou a été peu étudié et est très peu représenté dans les collections mondiales des ressources phytogénétiques (Eyzaguirre et al., 1997 ; Azam-Ali et al., 2001).
3En Côte d’Ivoire, le voandzou est cultivé dans la partie centrale et septentrionale du pays, caractérisées par trois zones agroécologiques différentes. Le climat du centre de la Côte d’Ivoire est caractérisé par quatre saisons (deux saisons sèches et deux saisons des pluies) et une pluviométrie de 1000 à 1700 mm/an. Le sol est peu profond avec une végétation de savane dominée par les arbustes et arbres de moyenne taille et quelques vestiges de forêts. Dans cette zone, le voandzou est une plante secondaire cultivée seule ou en association avec l’igname. La seconde région de culture est l’ouest de la Côte d’Ivoire, dominée par un paysage de montagne de moyenne altitude (entre 1000 et 1800 m d’altitude) et une pluviométrie de 1400 à 2300 mm/an. Le climat est caractérisé par l’existence de 2 saisons : une longue saison des pluies (mars à octobre) et une courte saison sèche (novembre à février). Le sol est lourd, profond, riche en matière organique en décomposition et est recouvert d’une végétation de type tropical humide constituée de forêts denses. Le voandzou y est cultivé sur de petites parcelles en culture pure ou en association avec le manioc et le bananier plantain. La principale zone de production de la Côte d’Ivoire est le nord. La pluviométrie est de 1000 à 1700 mm/an et le climat est caractérisé par une longue saison sèche (octobre à mars) et une saison des pluies (avril à septembre) marquée par des pluies irrégulières. Le paysage est dominé par les hautes herbes avec quelques arbustes. Le sol est sableux, superficiel parfois latéritique et pauvre. Dans le nord, les cultures pures ou en association avec les céréales (maïs, sorgho et mil) occupent des superficies plus importantes. La production du voandzou y dépasse la demande locale et l’excédent est acheminé dans la capitale par les grossistes où il est vendu sur les marchés par des détaillantes. Les classifications traditionnelles des variétés dans les différentes régions sont établies principalement sur la base des couleurs du tégument des graines, et secondairement selon l’habitus de croissance de la plante et la durée du cycle. On parle alors de types morphologiques ou morphotypes. Certains morphotypes sont spécifiques de régions données alors que d’autres sont distribués dans l’ensemble des régions de production.
2. Etat de la recherche sur le voandzou en Afrique
4Les études sur le voandzou en Afrique portent sur des aspects phytotechniques, biochimiques et de diversité génétique. Des études phytotechniques ont été menées notamment en ce qui concerne les amendements en engrais et le meilleur conditionnement des graines pour faciliter la germination (Massawe et al., 1999 ; Collinson et al., 2000). L’influence de la température et de la durée de l’éclairement sur la germination, la croissance et le développement du voandzou a été étudiée pendant plusieurs années et une modélisation a même été élaborée (Harris, Azam-Ali, 1993 ; Brink, 1997 ; 1999 ; Kocabas et al., 1999 ; Brink et al., 2000). Ces études ont montré que le voandzou est une plante de jour court qui fleurit mieux et donne un meilleur rendement en graines sous une photopériode de 10 à 12 heures par jour. La plante supporte des températures variant entre 16 et 35 °C (Linnemann, 1993 ; 1994 ; Linnemann, Craufurd, 1994 ; Linnemann et al., 1995). En raison de la sensibilité à la photopériode, Collinson et al. (1996) conseillent de sélectionner les types locaux insensibles à la photopériode pour le développement des gousses car ils peuvent être cultivés toute l’année. Le voandzou tolère assez bien les sols acides et légèrement pauvres, possède une bonne résistance au stress hydrique et se caractérise par un lien étroit entre une bonne capacité de fixation symbiotique de l’azote et un rendement élevé en graines. Les potentialités de cette légumineuse expliqueraient sa présence commune dans les régions semi arides du continent africain (Kishinevsky et al., 1996 ; Collinson et al., 1996 ; 1997 ; Uguru, Ezeh, 1997 ; Azam-Ali, 1998).
5Les études biochimiques confirment les remarquables propriétés nutritives du voandzou, notamment en tant que supplément protéique. Son addition dans les préparations locales permet d’équilibrer l’alimentation quotidienne des populations et de réduire ainsi les carences alimentaires en protéines très fréquentes sous les tropiques.
6Le voandzou est préférentiellement autogame mais des fécondations croisées sont observées entre certaines variétés. La période de fécondation est marquée par l’intervention de deux espèces de Formicidae (Pheidole megacephala Fabricius et Monomorium pharaonis Linnaeus) qui facilitent la nouaison et la pénétration des gousses dans le sol. Le développement des gousses précède la maturation des graines ; le rendement du voandzou est réduit par une trop forte humidité du sol au cours de cette phase (Doku, 1968 ; Doku, Karikari, 1970 ; 1971 ; Ameyaw, Doku, 1983). Bien que plus résistant aux maladies que les autres légumineuses, le voandzou montre des baisses de rendement suite aux attaques par les insectes (Dike, 1997). Le traitement des graines par des huiles essentielles permet de réduire les dégâts causés par les ravageurs de denrées stockées (Ajayi, Lale, 2001).
7Une importante collection de voandzou (2008 accessions) est conservée à l’IITA (International Institute of Tropical Africa), situé à Ibadan au Nigeria. Les principaux pays d’où sont originaires les accessions sont par ordre d’importance le Nigeria (310), la Zambie (284), le Zimbabwe (245), le Cameroun (207), le Togo (139), le Ghana (120) et la République de Centrafrique (103). La variabilité morphologique et agronomique observée entre les accessions de voandzou est grande (Goli et al., 1997 ; Karikari, 2000) mais elle n’apparaît pas dans les études de diversité génétique réalisées à l’aide de marqueurs neutres (protéines de réserves, enzymes, RAPD et AFLP) (Odeigah, Osayinpeju, 1998 ; Pasquet et al., 1999 ; Amadou et al., 2001 ; Massawe et al., 2002 ; Ntundu et al., 2004). Le voandzou originaire de la Côte d’Ivoire est très peu représenté dans les collections africaines. On compte seulement quatre accessions originaires de la Côte d’Ivoire sur les 2008 accessions conservées à l’IITA (Eyzaguirre et al., 1997). A notre connaissance, aucune étude n’a été faite sur le voandzou en Côte d’Ivoire. Pour remédier à cette lacune portant sur une plante bien connue des populations rurales et dans le but de promouvoir et vulgariser la culture, le laboratoire de Génétique de l’Université d’Abobo-Adjamé (UAA) a entrepris de collecter les morphotypes de voandzou rencontrés en Côte d’Ivoire afin d’étudier la diversité génétique, la phytotechnie et l’amélioration de cette légumineuse.
8Ce travail présente les résultats d’une étude de variabilité portant sur 26 caractères et 4 morphotypes de la collection de l’Université d’Abobo-Adjamé (Côte d’Ivoire). Les objectifs sont d’évaluer la diversité existant entre ces quatre morphotypes et de sélectionner les caractères pouvant servir lors des évaluations de routine impliquant l’ensemble des morphotypes de la collection de l’UAA.
3. Matériel et méthodes
3.1. Matériel végétal et description des sites d’étude
9Dans la région nord de la Côte d’Ivoire, le voandzou est une plante couramment cultivée à côté des cultures principales que sont le cotonnier et l’anacardier (pour l’exportation), les céréales (maïs, sorgho, mil) et l’igname pour l’alimentation de base. Le voandzou est caractérisé par des graines avec des colorations très diversifiées, un habitus de croissance à port érigé ou rampant, des gousses d’aspect lisse ou rugueux contenant une ou deux graines et un cycle cultural de quatre à huit mois. Le nord de la Côte d’Ivoire est la seule région où la production est excédentaire et le voandzou régulièrement consommé. Après la récolte (décembre à février), les productrices du nord, regroupées ou non en coopératives, font acheminer leur production sur certains marchés à Abidjan à destination des populations originaires du nord pour lesquelles cette légumineuse fait partie intégrante du régime alimentaire. C’est auprès de ces productrices que quatre morphotypes originaires de la région de Korhogo (5 °37’ longitude ouest, 9 °25’ latitude nord) ont été sélectionnés pour cette étude de variabilité. Les graines des quatre morphotypes utilisés présentaient les caractéristiques suivantes (Figure 1) :
10– graines de couleur beige panachée de rayures rouges (BPR) ;
11– graines de couleur ivoire crème unie (ICU) ;
12– graines de couleur rouge bordeaux unie (RBU) ;
13– graines de couleur noire foncée unie (NFU).

14La zone des essais est située dans la région agroécologique du sud de la Côte d’Ivoire dans la ville d’Abidjan (5 °17’- 5 °31’longitude ouest, 3 °45’- 4 °22’ latitude nord). Le climat de la région, localement appelé attiéen se caractérise par quatre saisons comme dans le centre du pays. La végétation est constituée de forêt tropicale dense et la pluviométrie est abondante (1400 à 2400 mm/an). La zone se caractérise par deux saisons pluvieuses (avril à juillet et octobre à novembre) espacées de deux saisons sèches (décembre à mars et août à septembre). La température est peu variable au cours de l’année (entre 25 et 30 °C) et l’humidité relative est très élevée (>80 %). Les principales cultures de rente sont le caféier, le cacaoyer et l’hévéa. Les cultures vivrières traditionnelles sont le manioc et le bananier plantain. Le sol du site (ferme expérimentale de l’Université d’Abobo-Adjamé) est profond, riche en matière organique et en humus, ce qui lui confère une teinte noire. Le semis a eu lieu au cours de la grande saison des pluies le 15 juillet 2002. A titre de comparaison, les périodes de semis dans le nord (juin à août) correspondent à des jours longs (13 à 14 h d’éclairage/jour) et les récoltes se font en pleine saison sèche (décembre à janvier) durant les jours courts (11 h d’éclairage/jour). Les températures moyennes à Abidjan étaient respectivement de 27,1 °C et 28,4 °C en 2001 et 2002 ; celles de Korhogo (une ville du nord) de 27,2 °C et 27,5 °C aux mêmes périodes. L’humidité relative est plus importante à Abidjan (79 et 85 % respectivement en 2001 et 2002) qu’à Korhogo (64 et 63 % respectivement en 2001 et 2002).
3.2. Méthodes expérimentales
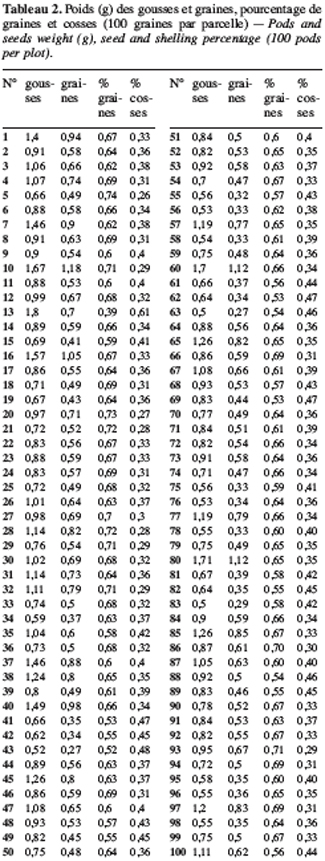
15Les graines ont été semées en culture pure sur une ancienne jachère. Le dispositif expérimental est constitué de blocs aléatoires complets avec deux blocs et chaque morphotype est répliqué deux fois par bloc, ce qui donne au total 16 parcelles. Les dimensions des parcelles sont de 1,2 m1,2 m et elles sont séparées de 0,5 m. Chaque parcelle porte cinq rangées de cinq plantes soit une densité de 17,36 plantes/m2, 100 individus par morphotype et un total de 400 individus mis en culture. Toutes les graines sont semées à 3 cm de profondeur avec un espacement de 25 cm entre deux poquets. Nous avons examiné 26 caractères recensés dans la liste des descripteurs du voandzou (IPGRI/IITA/BAMNET, 2000). On distingue parmi ces caractères 23 variables quantitatives (date d’émergence, taux de germination, longueur et largeur de la foliole terminale, longueur du pétiole et de l’entrenœud, nombre de feuilles, envergure de la plante, hauteur de la plante, nombre de nodosités, nombre de nœuds par tige et nombre de tiges par pied, habitus de croissance, longueur de l’étendard de la fleur, date d’apparition de la première fleur et de 50 % de floraison, poids de 100 graines, pourcentage de gousses matures, pourcentage de cosses, nombre de gousses par plante, durée du cycle, longueur et largeur des graines) et trois variables qualitatives (forme de la foliole terminale, texture et forme de la gousse). Toutes les mesures sont faites au niveau de la parcelle sur cinq ou dix plantes en fonction des indications de la liste des descripteurs du voandzou (IPGRI/IITA/BAMNET, 2000). La date d’émergence et le taux de germination concernent tous les individus de chaque parcelle. Les variables quantitatives sont mesurées à l’aide d’un mètre ruban (en cm) ou d’un pied à coulisse à affichage digital (en mm). Le rapport entre la longueur du pétiole (P en mm) et la longueur de l’entrenoeud (I en mm) permet d’obtenir la valeur de l’habitus de croissance (P/I) ; la plante est dite rampante si la valeur est faible (P/I< 7), semi érigée si elle est moyenne (P/I = 7-9) et érigée si elle est élevée (P/I> 9) (IPGRI/IITA/BAMNET, 2000). A la récolte, les gousses immatures se distinguent par une coloration blanchâtre et après la période de séchage, elles présentent un aspect étiolé et ne contiennent qu’une ébauche de graine. Au niveau de chaque parcelle, un lot de 100 gousses matures, séchées dans les mêmes conditions au soleil pendant deux mois après la récolte, est pesé à l’aide d’une balance de précision (Melter Toledo). Chaque gousse est cassée puis la cosse et la graine sont pesées. Pour le poids de 100 graines, cinq lots de 100 graines sont pesés au niveau de chaque parcelle.
3.3. Analyses statistiques
16Pour chacun des caractères étudiés, nous avons procédé à une comparaison des moyennes entre morphotypes par l’analyse de la variance (ANOVA). Lorsqu’une différence significative est révélée entre morphotypes pour un caractère donné, l’ANOVA est complétée par le test de la plus petite différence significative (ppds). Ce test nous permet d’identifier le ou les morphotype(s) qui diffère(nt) significativement des autres (Dagnelie, 1998). Tous les tests statistiques ont été réalisés à l’aide du logiciel Minitab 12.0 version Windows.
4. Résultats
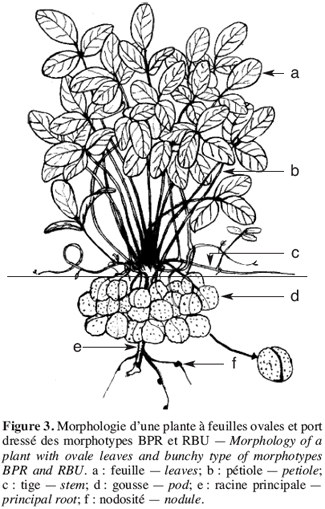
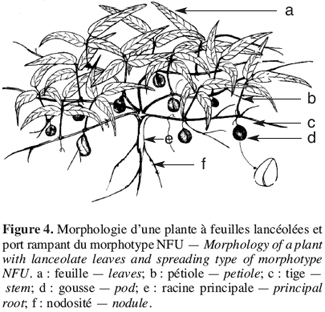
17Les caractères qualitatifs n’ont pas été pris en compte dans l’analyse statistique. Pour les trois caractères qualitatifs, nous observons que les morphotypes se regroupent en deux classes. Les morphotypes ICU et NFU possèdent une foliole terminale de forme lancéolée et des gousses lisses présentant une pointe à l’extrémité distale (Figures 2 et 4). A l’inverse, les morphotypes BPR et RBU ont une foliole terminale de forme ronde et des gousses rugueuses présentant une pointe aux extrémités distale et apicale (Figure 3).
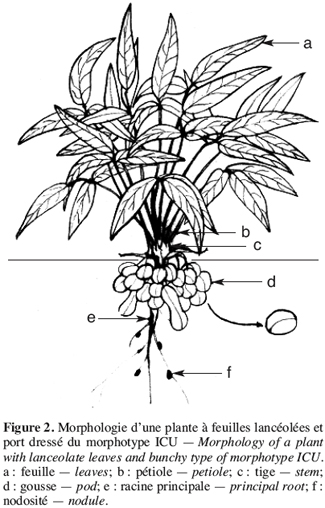
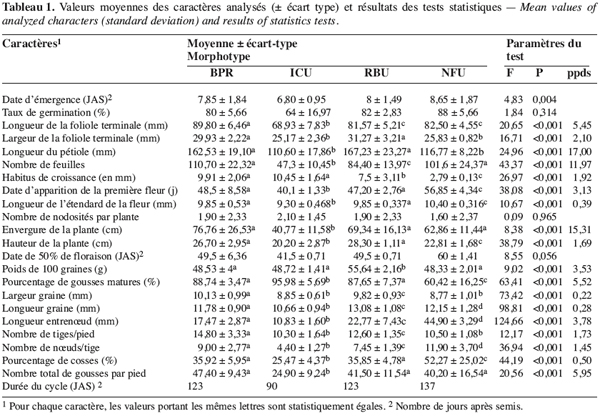
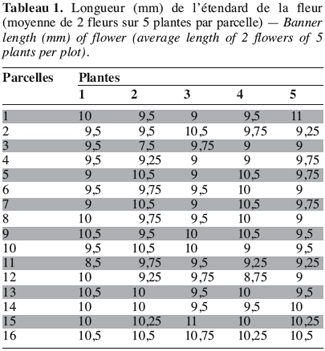
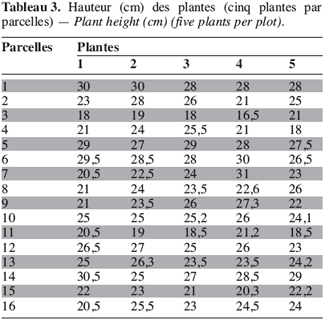
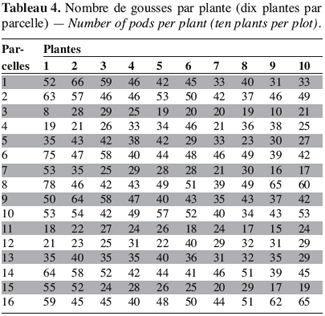
18Sur les 23 caractères quantitatifs utilisés dans l’analyse statistique, les différences entre les valeurs des quatre caractères suivants : la durée de la période d’émergence, le taux de germination, le nombre de nodosités et la date de 50 % de floraison, sont non significatives. Pour les 19 autres caractères, les valeurs observées présentent une différence hautement significative entre les morphotypes étudiés (Tableau 1). Les périodes d’émergence varient entre six et huit jours et les taux de germination sont de 64 à 88 % par morphotype au niveau des parcelles. Le nombre de nœuds par tige, la longueur des entrenœuds et la longueur de la graine sont significativement différents entre morphotypes (ppds = 1,45 ; 3,78 et 0,28, respectivement). Pour les caractères suivants : l’envergure de la plante, le nombre de gousses par pied et le poids de 100 graines, la différence hautement significative observée entre les valeurs est due à un morphotype, les trois autres ayant des valeurs statistiquement identiques. Ainsi, le poids de 100 graines plus élevé du morphotype RBU (55,64 ± 2,16 g), le faible nombre de gousses par pied (24,90 ± 9,24) et la petite envergure (40,77 ± 11,58 cm) du morphotype ICU sont responsables des différences hautement significatives observées pour ces trois caractères. On remarque un regroupement des morphotypes BPR et RBU d’une part, et d’autre part, des morphotypes ICU et NFU pour la largeur de la foliole terminale et la longueur du pétiole. Pour chacun des cinq caractères suivants : la longueur de l’étendard de la fleur, la date d’apparition de la première fleur, la hauteur des plantes, le nombre de gousses matures et le pourcentage de cosse, aucune différence statistique n’est observée entre les morphotypes RBU et BPR. à l’inverse les morphotypes ICU et NFU ont des valeurs statistiquement différentes pour ces caractères. Les données concernant la hauteur de la plante, la longueur de l’étendard de la fleur, le nombre de gousses par plante et le pourcentage de cosse sont présentées en annexe (Tableaux 1 à 4). La fin de la phase reproductive est marquée par l’allongement du pédoncule floral qui s’enfonce dans le sol marquant ainsi le début de la formation des gousses. Cette phase de développement commence assez vite (10 à 21 jours plus tard) après l’apparition de la première fleur chez les morphotypes ICU, BPR et RBU. Chez le morphotype NFU, des nouvelles fleurs naissent et s’épanouissent encore plus de 50 jours après l’apparition de la première fleur, la phase de maturation des gousses est ainsi étalée dans le temps pour ce morphotype.
19Les morphotypes BPR et ICU sont de type érigé (P/I = 9,91 ± 2,06 et 10,45 ± 1,64, respectivement) ; le morphotype RBU est semi érigé (P/I = 7,5 ± 3,11) (IPGRI/IITA/BAMNET, 2000). Ces trois morphotypes ont leurs gousses groupées autour de la racine principale (Figures 2 et 3). Le morphotype NFU est rampant (P/I = 2,79 ± 0,13) et produit des gousses sur toute la surface occupée par ses nombreuses tiges rampantes (Figure 4).
5. Discussion
20Chez les morphotypes de voandzou étudiés, l’émergence des plantules s’étale entre sept et dix jours après le semis comme cela a été noté par Linnemann (1993). La hauteur des plantes (20–29 cm) est similaire à celle obtenue par Goli et al. (1997) dont l’étude a été réalisée dans le sud du Nigeria dans une zone climatique similaire à la nôtre (forêt tropicale humide) à partir d’accessions maintenues ex situ. L’habitus de croissance est un caractère important à prendre en considération dans les études phytotechniques, notamment pour ce qui concerne les études de densité de semis. Les morphotypes à croissance érigée (ICU et BPR) ou semi érigée (RBU) permettent des densités de culture élevées ; le morphotype rampant NFU nécessite des densités plus faibles.
21Nous avons observé un faible nombre de nodosités chez les plantes, ce qui pourrait s’expliquer par la méthode de comptage utilisée qui a consisté à dégager la terre sous chaque plante dix semaines après semis pour observer les nodosités (IPGRI/IITA/BAMNET, 2000). Ce caractère doit faire l’objet de mesures plus appropriées telle que le dosage de la quantité d’azote symbiotique fixée dans les nodules (Uguru, Ezeh, 1997 ; Gueye et al., 1998). La capacité de fixation de l’azote chez le voandzou étant maximale pendant la phase reproductive (Kumaga et al., 1994), les analyses sur les nodosités devront être réalisées au cours de cette phase.
22L’amplitude du temps d’apparition de la première fleur (41–56 jours) est moins grande que celle observée par Goli et al. (1997) qui ont rapporté une période de 38 à 66 jours. Cela pourrait s’expliquer par le nombre important d’accessions étudiées par ces auteurs (1384) et leur origine géographique très large. Les morphotypes ICU, BPR et RBU ont une floraison groupée et le morphotype NFU présente une floraison échelonnée. Doku et Karikari (1970) ont observé ces deux types de floraison (groupée ou échelonnée) chez les variétés de voandzou originaires du Ghana. Ce caractère semble influencer le rendement chez le voandzou. Le pourcentage de gousses matures est très élevé chez les morphotypes à courte période de floraison (87 à 95 %) et faible (60 %) chez le morphotype à floraison échelonnée. Selon Karikari (2000), les morphotypes à floraison groupée disposeraient de suffisamment de temps pour la maturation des gousses et des graines avant l’apparition de la mauvaise saison et cela leur confère un intérêt agronomique certain dans l’optique d’une amélioration de la culture en milieu paysan. Le poids de 100 graines que nous avons obtenu, est plus faible que celui de la majorité des accessions étudiées par Goli et al. (1997) ; cela pourrait s’expliquer par la culture des morphotypes dans une zone forestière humide aux conditions climatiques et écologiques différentes des zones de production (notamment une forte humidité du sol consécutive à de fortes pluies en novembre). Doku et Karikari (1970) ont rapporté que la pluie entraîne une diminution du nombre de fleurs. La floraison des quatre morphotypes étudiés a débuté pendant la courte saison sèche (septembre) et s’est poursuivie pendant la courte saison des pluies (octobre à novembre), ce qui a entraîné la chute de nombreuses fleurs avant leur fécondation et a réduit le nombre de gousses par pied. Les durées de cycle signalées par Goli et al. (1997) et Karikari (2000) sont respectivement de 100 à 160 JAS et de 120 à 165 JAS. Le morphotype ICU, avec une durée de cycle de 90 JAS, est très précoce. Cette précocité permet d’envisager plus d’une culture par an, cela fait de ce morphotype un matériel de choix dans les programmes d’amélioration et de vulgarisation du voandzou en Afrique de l’Ouest.
6. Conclusions et perspectives
23Sur l’ensemble des 26 caractères évalués dans cette étude, seuls quatre n’ont pas permis de différencier les morphotypes. Les caractères analysés peuvent ainsi constituer des critères de base pour différencier les morphotypes et servir pour une étude de variabilité entre les morphotypes de voandzou de Côte d’Ivoire. Les morphotypes ICU, BPR, RBU et NFU ont manifesté une importante variabilité. La mise en place d’une banque de graines rassemblant le maximum de morphotypes ivoiriens est essentielle pour l’amélioration génétique du voandzou. Ces morphotypes seront analysés pour estimer leur diversité morphologique et agronomique sur la base des caractères les plus variables observés dans cette étude. Les caractères quantitatifs étant influencés par le milieu, il conviendrait de réaliser des essais agronomiques multilocaux dans différentes zones agroécologiques sur des saisons variables (deux cultures par an) pour mieux apprécier les morphotypes les plus performants. Par ailleurs, l’utilisation de morphotypes à maturité précoce et une meilleure adéquation des cultures avec les saisons peuvent aider à améliorer les rendements et faciliter une vulgarisation de la culture en milieu paysan.
24Remerciements
25Ce travail entre dans le cadre d’un programme d’études sur le voandzou financé par le Service de Coopération et d’Action Culturelle (SCAC) de l’Ambassade de France en Côte d’Ivoire : Programme d’Appui à la Recherche Agronomique (PARA), convention n° 2002 935, et la fondation internationale pour la science IFS (Grant number C/7434).
26Annexes
Bibliographie
Ajayi FA., Lale NES. (2001). Susceptibility of unprotected seeds and seeds of local bambara groundnut cultivars protected with insecticidal essential oils to infestation by Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). J. Stored Products Res. 37, p. 47–62.
Aké Assi L. (1988). La diversité génétique des espèces végétales sous-exploitées d'Afrique. In P. Perrino (Ed.) Crop genetic resources of Africa, vol. 1. Nairobi, Kenya : IBPGR/IITA/UNEP, p. 53–88.
Altieri MA. (1986). L'agroécologie : bases scientifiques d’une agriculture alternative. Paris : Debard, 237 p.
Amadou HI., Bebeli PJ., Kaltsikes PJ. (2001). Genetic diversity in bambara groundnut (Vigna subterranea L.) germplasm revealed by RAPD markers. Genome 44, p. 995–999.
Ameyaw GCE., Doku EV. (1983). Effect of soil moisture stress on the reproductive efficiency and yield of the bambara groundnut (Voandzeia subterranea). Crop Sci. 28, p. 23–29.
Azam-Ali SN. (1998). Evaluating the potential for bambara groundnut as a food crop in semi-arid Africa. An approach for assessing the yield potential and ecological requirement of an underutilised crop. Trop. Subtrop. Agric., p. 134-136.
Azam-Ali SN., Sesay A., Karikari KS., Massawe FJ., Aguilar-Manjarrez J., Bannayan M., Hampson KJ. (2001). Assessing the potential of an underutilized crop - a case study using bambara groundnut. Exp. Agric. 37, p. 433–472.
Brink M. (1997). Rates of progress towards flowering and podding in bambara groundnut (Vigna subterranea) as a function of temperature and photoperiod. Ann. Bot. 80, p. 505–513.
Brink M. (1999). Development, growth and dry matter partitioning in bambara groundnut (Vigna subterranea) as influenced by photoperiod and shading. J. Agric. Sci. 133, p. 159–166.
Brink M., Sibuga KP., Tarimo AJP., Ramolemana GM. (2000). Quantifying photothermal influences on reproductive development in bambara groundnut (Vigna subterranea): models and their validation. Field Crops Res. 66, p. 1–14.
Collinson ST., Azam-Ali SN., Chavula KM., Hodson DA. (1996). Growth, development and yield of bambara groundnut (Vigna subterranea) in response to soil moisture. J. Agric. Sci.126, p. 307–318.
Collinson ST., Clawson EJ., Azam-Ali SN., Black CR. (1997). Effects of soil moisture deficits on the water relations of bambara groundnut (Vigna subterranea L. Verdc.). J. Exp. Bot. 48, p. 877–884.
Collinson ST., Sibuga KP., Tarimo AJP., Azam-Ali SN. (2000). Influence of sowing date on the growth and yield of bambara groundnut landraces in Tanzania. Exp. Agric. 36, p. 1–13.
Dagnelie P. (1998). Statistique théorique et appliquée (Tome 1). Bruxelles : De Boeck & Larcier.
Dike MC. (1997). Effect of insect pests on pod and seed yields of bambara groundnut, Vigna subterranea L. verde in Nigeria. Int. J. Pest Manage. 43, p. 191–192.
Doku EV. (1968). Flowering, pollination and pod formation in bambara groundnut (Voandzeia subterranea) in Ghana. Exp. Agric. 4, p. 41–48.
Doku EV., Karikari S. (1970). Fruit development in bambara groundnut (Voandzeia subterranea). Ann. Bot. 34, p. 951–957.
Doku EV., Karikari S. (1971). The role of ants in pollination and pod production of bambarra groundnut. Econ. Bot. 25, p. 357–362.
Eyzaguirre P., Thormann I., Goli AE., Attere AF. (1997). Strategies for the conservation and use of neglected and under-utilized crops in the African savannah: fonio and bambara groundnut. In A. Begic (Ed). Actes du colloque “Gestion des ressources génétiques des plantes en Afrique des savanes”, 24–28 février, 1997, IER-BRG-Solagral, Bamako, Mali, p. 315–322.
Goli AE., Begemann F., Ng NQ. (1997). Characterization and evaluation of IITA’s bambara groundnut collection. In Heller J., Begemann E., Mushonga J. (Eds). Promotion of the conservation and use of underutilized and neglected crops. 9. Proceedings of the workshop on conservation and improvement of bambara groundnut (Vigna subterranea (L.) Verdc.), 14-16 November 1995, IPGRI, Harare, Zimbabwe, p. 101–118.
Gueye M., James EK., Kierans M., Sprent JI. (1998). The development of root nodules on bambara groundnut (Voandzeia (Vigna) subterranea). World J. Microbiol. Biotechnol. 14, p. 177–184.
Harris D., Azam-Ali SN. (1993). Implications of day length sensitivity in bambara groundnut (Vigna subterranea) for production in Botswana. Agric. Sci. 120, p. 75–78.
IPGRI/IITA/BAMNET. (2000). Descriptors for bambara groundnut: 48, Rome, Italy.
Karikari KS. (2000). Variability between local and exotic bambara groundnut landraces in Botswana. Afr. Crop Sci. J. 8, p. 153–157.
Kumaga S., Danso SKA., Zapata F. (1994). Time-course of nitrogen fixation in two bambara groundnut (Vigna subterranea L. Verdc.) cultivars. Biol. Fertil. Soils 18, p. 231–236.
Kishinevsky BD., Zur M., Friedman Y., Meromi G., Ben-Moshe E., Nemas C. (1996). Variation in nitrogen fixation and yield in landraces of bambara groundnut (Vigna subterranea L.). Field Crops Res. 48, p. 57–64.
Kocabas Z., Craignon J., Azam-Ali SN. (1999). The germination response of bambara groundnut (Vigna subterranea (L.) Verdc.) to temperature. Seed Sci. Technol. 27, p. 303–313.
Linnemann AR. (1993). Phenological development in bambara groundnut (Vigna subterranea) at constant exposure to photoperiods of 10 to 16 h. Ann. Bot. 71, p. 5–452.
Linnemann AR. (1994). Phenological development in bambara groundnut (Vigna subterranea) at alternate exposure to 12 and 14 h photoperiods. J. Agric. Sci. 123, p. 40.
Linnemann A., Craufurd P. (1994). Effects of temperature and photoperiod on phenological development in three genotypes of bambara groundnut (Vigna subterranea). Ann. Bot. 74, p. 675–681.
Linnemann AR., Westphal E., Wessel M. (1995). Photoperiod regulation of development and growth in bambara groundnut (Vigna subterranea). Field Crops Res. 40, p. 39–47.
Massawe FJ., Collinson ST., Roberts JA., Azam-Ali SN. (1999). Effect of pre-sowing hydration on germination, emergence and early growth of bambara groundnut (Vigna subterranea L. Verdc.). Seed Sci. Technol. 27, p. 893–905
Massawe FJ., Dickinson M., Roberts JA., Azam-Ali SN. (2002). Genetic diversity in bambara groundnut (Vigna subterranea (L.) Verdc.) landraces revealed by AFLP markers. Genome 45, p. 1175–1180.
Minka DR., Bruneteau M. (2000). Partial chemical composition of bambara pea (Vigna subterranea L. Verdc.). Food Chem. 68, p. 273–276.
Nacoulma-Ouédraogo O. (1996). Plantes médecinales et pratiques traditionnelles au Burkina-Faso : cas du plateau central. Thèse de doctorat des Sciences Naturelles, Université de Ouagadougou. Ouagadougou, Burkina-Faso.
Ntundu W., Bach I., Christiansen J., Andersen S. (2004). Analysis of genetic diversity in bambara groundnut [Vigna subterranea (L.) Verdc] landraces using amplified fragment length polymorphism (AFLP) markers. Afr. J. Biotechnol. 3, p. 220–225.
Odeigah PGC., Osanyinpeju AO. (1998). Evaluating the genetic diversity of bambara groundnut accessions from Nigeria using SDS-polyacrylamide gel electrophoresis. Gen. Res. Crop Evol. 45, p. 451–458.
Oliveira J. (1976). Grain legumes of Mozambique. Trop. Grain Legume Bull. 3, p. 13–15.
Onimawo IA., Momoh AH., Usman A. (1998). Proximate composition and functional properties of four cultivars of bambara groundnut (Voandzeia subterranea). Plant Foods Hum. Nutr. 53, p. 153–158.
Pasquet RS., Schwedes S., Gepts P. (1999). Isosyme diversity in bambara groundnut. Crop Sci. 39, p. 1228–1236.
Ucciani E., Busson F. (1963). Contribution à l'étude des corps gras de Voandzeia subterranea Thouars (Papilionnacées). Oléagineux 18, p. 45–48.
Uguru MI., Ezeh NE. (1997). Growth, nodulation and yield of bambara groundnut (Vigna subterranea (L) Verdc.) on selected Nigerian soils. J. Food Agric. 73, p. 377–382.
