


- Portada
- volume 9 (2005)
- numéro 2
- A landscape approach for the study of calcareous grassland plant communities


Vista(s): 2330 (25 ULiège)
Descargar(s): 112 (4 ULiège)

A landscape approach for the study of calcareous grassland plant communities

Résumé
Une approche paysagère pour l’étude des communautés végétales des pelouses calcaires. La fragmentation des habitats est la menace principale pour la biodiversité des habitats semi-naturels. Les conséquences de la fragmentation sur la biodiversité peuvent être étudiées dans le contexte de l’écologie du paysage. Dans ce contexte trois paradigmes permettent de poser les hypothèses sur l’évolution de la biodiversité des habitats fragmentés : le paradigme de la biogéographie insulaire, le paradigme des petites populations et le paradigme des métapopulations. L’examen de deux cas d’étude montre que les pelouses calcaires en Belgique ont subi une fragmentation très importante. La taille des taches d’habitat est le facteur principal qui détermine à la fois la richesse des communautés, la composition spécifique des taches d’habitats et la probabilité de présence d’espèces caractéristique dans un système de pelouses calcaires fragmentées en Calestienne, Belgique.
Abstract
Habitat fragmentation is the principal threat on biodiversity in semi-natural habitats. The consequences of fragmentation for biodiversity may be studied within the framework of landscape ecology. In this context, three paradigms have generated hypotheses that need to be tested in field situations: the insular biogeography paradigm, the small population paradigm and the metapopulation paradigm. Examination of two case studies showed that Belgian calcareous grasslands have undergone a drastic fragmentation process during the last century. Size of patches is shown to be the most important determinant for species richness, species composition and probability of presence of characteristic species in a fragmented system of calcareous grasslands in Calestienne, Belgium.
Tabla de contenidos
1. Introduction
1Ecological systems are naturally dynamic at all spatio-temporal scales from geological phenomenon to individual migration. This is particularly relevant for ecological landscapes, defined as the level of organization that refers to an heterogeneous portion of territory, consisting of interacting ecosystems, similarly repeated in space (Forman, Godron, 1986). Besides this natural pattern, during the last centuries, on most of the earth surface, human activities have become the main cause of landscape dynamics (Baudry, Tatoni, 1993) in relation with modification of land use. This human-driven dynamics fostered the fragmentation of natural and semi-natural habitats. Fragmentation refers to the reduction of large continuous patches of habitat into small isolated patches. It results in habitat loss, decreasing patch size and greater spatial isolation. All these features may significantly affect communities diversity as well as population dynamics. Habitat destruction and fragmentation are considered the principal causes of species extinction worldwide (Opdam et al., 1993; Bruun, 2000).
2Habitat fragmentation is a prominent characteristic of semi-natural habitats traditionally managed by extensive agro-pastoral techniques, such as calcareous grasslands. Well developed throughout western Europe at the end of the nineteenth, those ecosystems have undergone a drastic fragmentation due to the abandonment of the traditional agropastoral practices responsible for their extension (Poschlod, WallisDeVries, 2002; Willems, 2001). Those communities have been whether directly replaced by intensive agricultural or sylvicultural systems, or have undergone spontaneous succession to forest (Poschlod, WallisDeVries, 2002; Bobbink, Willems, 1987).
3In this paper, we examine the basic paradigms adopted for studies on relationships between landscape evolution and biodiversity, and we illustrate those paradigms with results from an ongoing project on plant species conservation in Belgian calcareous grasslands.
2. Relations between landscape evolution and biodiversity: the paradigms
4The relations between landscape dynamics and biodiversity are currently studied within the context of the emerging science of landscape ecology. Landscape ecology is the science that examines the way in which landscape features (composition, spatial structure and dynamics of habitats) affect ecological patterns (i.e. composition of communities) and ecological processes (i.e. population dynamics). The study of biodiversity patterns and processes at the landscape level in regard to habitat fragmentation is based on three main paradigms.
5– From a community perspective, the insular biogeography paradigm postulates that the species richness of an island is related to its area and its distance to mainland (MacArthur, Wilson, 1963). Species richness on islands reaches a dynamic equilibrium regulated by species extinctions and immigrations from the mainland. Extinction rate is related to the island area, with larger islands displaying lower extinction rates, while immigration rate is related to the distance to mainland, with lower immigration rates on more distant islands. Applied to fragmented habitats, the insular biogeography paradigm states that small isolated patches of habitat host less species than large connected ones.
6– From a species perspective, the small population paradigm postulates that populations that have undergone a demographic bottleneck are more prone to extinction, due to genetic drift and increased inbreeding, higher demographic stochasticity and higher sensitivity to environmental stochasticity. It can be expected that the reduction of habitat size due to fragmentation would lead to a diminution of population size, and then, to higher extinction probabilities (Lande, 1988; Young et al., 1996; Morgan, 1999).
7– Again from a species perspective, the metapopulation paradigm states that, at the landscape level, populations are not isolated from each other, but linked by individuals (diaspores) or pollen exchanges and are characterized by extinction-recolonisation dynamics (Hanski, 1989). Metapopulation viability is influenced by local populations viability and by species ability to recolonize suitable habitat patches, depending on habitat spatial configuration and on dispersal capacities of the considered species.
8Based on these paradigms, the relations between landscape features and population demography may be examined in regard to the population growth equation :
9Nt+1 = Nt + B – D + I – E
10where
11Nt and Nt+1 represent the number of individuals in the population respectively at times t and t+1,
12B and D represent births and deaths that occurred between times t and t+1,
13I and E represent the number of individuals that immigrated within and emigrated from the population, respectively.
14Considering the influence of the spatial structure of habitats, birth and death rates will be influenced mainly by patch size (small population paradigm), while immigration and emigration rates will be influenced mainly, but not only, by distances between patches (metapopulation paradigm). Then, persistence of a species at the landscape scale depends on the interaction between species life history traits and landscape structure (Hanski, 1998).
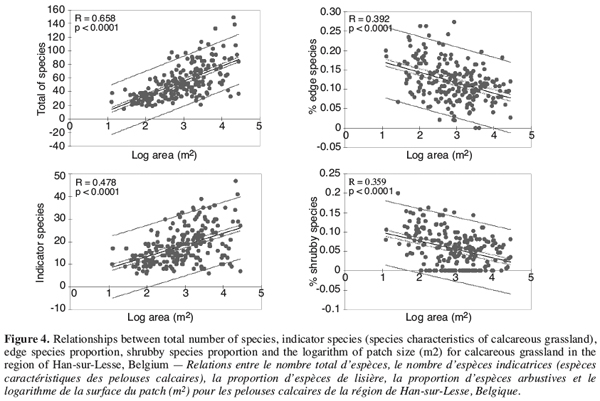
3. The fragmentation of calcareous grasslands in Belgium
15We have recently examined the spatio-temporal dynamics of calcareous grasslands fragmentation in Belgium with two studies (Leduc, Mahy, 2004; Dugnoille, 2003). Leduc and Mahy described the landscape evolution over a period of 250 years, in a rural area of 3500 ha, situated in the municipality of Viroinval. The study was based on historical maps and recent aerial pictures: Ferraris maps (1770, land-use maps drawn for military purpose), the ICM maps (1870, land-use maps), vegetation maps (1964, phytosociological associations) and recent aerial photographs (1998). Two main modifications of the landscape were pointed out:
16– the transition of the landscape matrix from cultivated land in the 1700s to forest nowadays,
17– a dramatic fragmentation of open semi-natural habitat, corresponding mostly to calcareous grasslands (Figure 1).
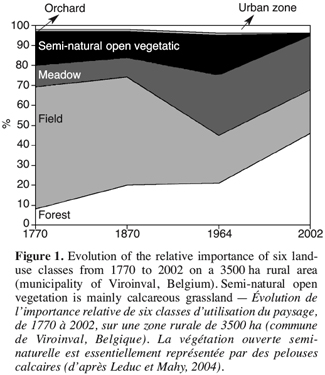
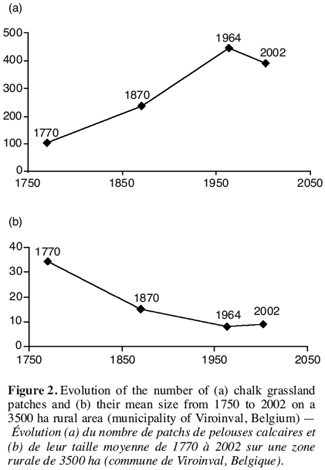
18This phenomenon is rather recent, with a diminution of 93% of the calcareous grasslands surface from 1964 to 2002. Their total area passed from 716 ha in 1964 to 51 ha in 2002, that is to say less than 2% of the considered territory surface. A high proportion of the historical calcareous grasslands has been afforested at the end of the nineteenth with pine (Pinus sylvestris and Pinus nigra). Figure 2 illustrates the consequences of calcareous grassland decrease at the scale of individual habitat patches. From 1750s to 1960s, the number of grassland patches increased while their mean size decreased regularly. This pattern reflects the division of large continuous patches into smaller ones and is typical for a fragmentation pattern. From 1960s up to now, the system collapsed dramatically with the disappearance of most of the remnant patches.
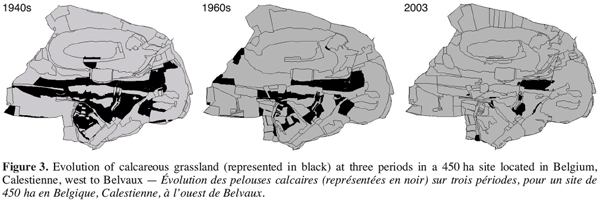
19The same pattern was reported by Dugnoille (2003) who described calcareous grasslands evolution at smaller spatial (450 ha) and temporal (60 years) scales in the region of Han-sur-Lesse. Calcareous grasslands were localized on aerial photographs dating from 1948, 1965 and 2002. Figure 3 shows that grasslands evolution was very similar to the pattern reported by Leduc and Mahy (2004). In Belgium, it has been estimated that surfaces covered by calcareous grasslands passed from thousands of hectares in the 19th century to hardly 200 ha currently (Delescaille, 2000).
20Those changes in calcareous grassland configuration in Belgium were due to various factors. Extensive grazing has been gradually abandoned at the end of the 19th. Wool produced overseas and new textile fibres appeared on the market as being more competitive. Use of chemical fertilizers became widespread, and enclosure techniques were largely adopted (Decocq et al., 1996). As a consequence, areas with low productivity, that did not fit intensive agriculture requirements, were progressively abandoned and underwent natural forest recolonization. In addition, massive reforestations with Austrian Black Pine took place in the 1930s in this region (Vandermotten, Decroly, 1995).
4. The influence of landscape structure on plant communities and species of calcareous grasslands in Calestienne, Belgium
21In an ongoing research program, our team is studying the relationships between landscape structure on the one hand, and communities composition, individual species distribution and population reproductive success on the other hand, in calcareous grasslands in Calestienne.
22The study site, about 2500 ha, is situated in the region of Han-sur-Lesse – Rochefort (Belgium) and corresponds to the Natura 2000 site BE35038. In 2003, the site has been intensively surveyed and all calcareous grassland patches localized. Any patch larger than 25 m2 was individually mapped. Localization and extension of calcareous grassland patches have been transferred in a GIS in Arc View 3.2. In each patch, we realized an exhaustive inventory of superior plants species, with a semi-quantitative evaluation of the population size for each species (<10 flowering heads, 10–50 flowering heads, 50–500 flowering heads, >500 flowering heads). A total of 216 calcareous grassland patches were found with surfaces ranging from 25 m2 to nearly 3 ha. The total surface of calcareous grassland was about 40 ha, that is less than 2% of the study site area.
4.1. Species richness
23Firstly, we examined the effect of the landscape structure on plant species richness and species composition of calcareous grassland. Species richness referred to the total number of species found in each patch. Species composition was assessed in regard to conservation objectives. Species were arranged in three groups:
24I) Species considered as characteristic of calcareous grassland phytosociological associations reporting to xeric grasslands (Xerobromion), mesophilous grasslands (Mesobromion) and open grasslands of the Alysso-Sedion and Sedo-Scleranthetea;
25II) Edge species;
26III) Shrubby species.
27Multiple regressions were used to test for significant relationships between i) the total number of species or the number of species of group I (characteristic of calcareous grasslands) in a patch, and ii) the spatial configuration of the patch, estimated by its size (logarithm of surface), its shape (perimeter to surface ratio) and its isolation (distance to the nearest neighbour). In the same way, multiple regressions were used to test for a significant relationship between i) the proportion of species of group II (edge) and III (shrub) in a patch and ii) the spatial configuration of the patch, estimated by its size (logarithm of surface), its shape (perimeter to surface ratio) and its isolation (distance to the nearest neighbour). We tested whether the floristic similarity among patches differed between pairs of small patches (<0,01 ha) and pairs of large patches (>0,5 ha). Significant difference for mean Jaccard similarity index between pairs of patches in the two groups of patches (small vs large) was tested using a one-way ANOVA.
28Patch area (logarithmic scale) was the only spatial parameter that explained a significant part of the variation among patches for the total number of species (r=0.699; P<0.01) and the number of grassland characteristic species (r=0.574; P<0.01), with larger patches containing more species (Figure 4). Patch area (logarithmic scale) was also the only spatial parameter that explained a significant part of the variation among patches for the proportion of edge species (r=-0.393; P<0.01) and shrubby species (r=-0.404; P<0.01), with smaller patches containing a higher proportion of edge and shrubby species (Figure 4). Isolation and shape of patches were not significantly related to species richness or species composition. Those preliminary analyses emphasized that, in a conservation and restoration strategy, priority should be given to the maintenance and restoration of large grassland patches, while connectivity seems to be, in a first approach, a less important factor for plant species richness. Other studies have reported that size is the main factor affecting species richness in fragmented grasslands (Krauss et al., 2004), but in some cases, isolation has also proven to be a significant factor (Bruun, 2000; Piessens et al., 2005). In our study, we must underline that we evaluated the degree of patch isolation on the basis of one single, very crude, parameter: the distance to the nearest neighbour. This measure may seldom reflect the real isolation of a single patch because it does not take into account the surface of similar habitat adjacent to the target patch (Moilanen, Nieminen, 2002). More developed indices of isolation should be used in refined analyses. In addition, most of calcareous grasslands species are perennials. For such species, fragmentation of calcareous grasslands may be too recent (about 40 years) to have developed all its effects on population viability. Delay to extinction has been reported by different studies on grassland fragmentation (Bruun, 2000; Krauss et al., 2004). Then, isolated fragments may still retain a significant species diversity despite the fact that individual species suffer from a reduction of their demographic performances. Mean Jaccard similarity was significantly higher among small patches than among large patches, indicating that small patches exhibited a more homogeneous floristic composition than large patches. This pattern may be partly explained by a lower quality of small patches for calcareous species and an homogenisation of the flora under the influence of shrubby colonization that was shown to be proportionally more important in small patches.
4.2. Species distribution and persistence
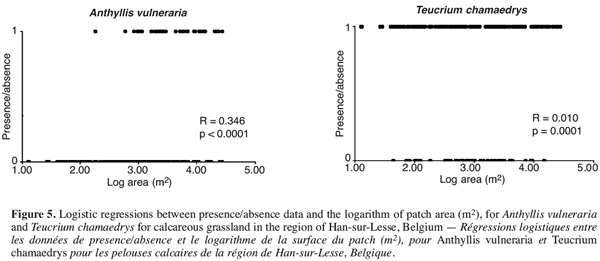
29To examine the influence of landscape structure on the fate of individual species, we tested whether the size of a patch was significantly related to the presence of typical calcareous grassland species in this patch by means of logistic regressions for 39 species (Table 1). Our hypothesis was that larger patches had a higher probability to host individual typical species because of more diverse, and then probably more favourable, environmental conditions, and because of a higher capacity to sustain large populations, less prone to extinction. For 28 out of the 39 species, patch area influenced significantly their probability of presence. Such a significant role of area has been found in other and more detailed studies (Ouborg, 1993). Figure 5 gives two examples of logistic regression output. Besides the general pattern linked to species richness, this preliminary analysis showed that spatial configuration of habitat may also be important for the maintenance of individual plant species. In the future, more refined analysis should examine the relationships between species life-history traits and their response to habitat configuration.

5. Conclusions
30As elsewhere in Europe, Belgian calcareous grasslands suffered from dramatic fragmentation during the last century. Detailed studies on the evolution of those habitats showed that this fragmentation is a rather recent phenomenon, dating from 1950s. Landscape ecology provides a pertinent conceptual framework to study the impacts of fragmentation on biodiversity, as illustrated by the case study presented here. However, examination of field situations showed that the response of biodiversity to landscape modifications may be more complex than expected on the basis of the simple hypotheses generated by the different theoretical frameworks related to landscape ecology. When examining such responses, communities and species biological traits should be taken into account both for the generation of hypotheses and for the interpretation of correlative patterns.
Bibliographie
Baudry J., Tatoni T. (1993). Changes in landscape patterns and vegetation dynamics in Provence, France. Landscape Urban Plann. 24, p. 153–159.
Bobbink R., Willems JH. (1987). Increasing dominance of B. pinnatum (L.) Beauv. in chalk grasslands: a threat to a species rich ecosystem. Biol. Conserv. 40, p. 301–314.
Bruun HH. (2000). Patterns of species richness in dry grassland patches in an agricultural landscape. Ecography 23, p. 641–650.
Decocq O., Delescaille LM., Dewitte T. (1996). Les pelouses calcicoles : une origine agro-pastorale. In Les pelouses calcicoles en Région Wallonne. Vierves-sur-Viroin, Belgique : Entente nationale pour la Protection de la Nature, p. 16–21.
Delescaille LM. (2000). La gestion des pelouses sèches en Belgique. Actes du séminaire de Bourges : Pelouses sèches relictuelles de France : connaissances scientifiques et gestion des sites, p. 11–26.
Dugnoille X. (2003). Etude du maillage écologique en relation à la gestion des pelouses calcaires du parc de la Lesse et de la Lomme. Travail de fin d’études. Bruxelles, ULB école Interfacultaire de Bioingénieurs, 53 p.
Forman RTT., Godron M. (1986). Landscape Ecology. New York: Wiley, 620 p.
Hanski I. (1989). Metapopulation dynamics: does it help to have more of the same? Trends Ecol. Evol. 4, p. 113–114.
Hanski I. (1998). Metapopulation dynamics. Nature 396, p. 41–49.
Krauss J., Klein AM., Steffan-Dewenter I., Tscharntke T. (2004). Effects of habitat area, isolation, and landscape diversity on plant species richness of calcareous grasslands. Biodiversity Conserv. 13, p. 1427–1439.
Lande R. (1988). Genetics and demography in biological conservation. Science 241, p. 1455–1460.
Leduc L., Mahy G. (2004). Evolution d’un paysage écologique de Calestienne de la fin du XVIIe à nos jours. Parcs Réserves 59, p. 41–56.
MacArthur RH., Wilson EO. (1963). An equilibrium theory of insular zoogeography. Evolution 17, p. 373–387.
Moilanen A., Nieminen M. (2002). Simple connectivity measures in spatial ecology. Ecology 83 (4), p. 1131–1145.
Morgan JW. (1999). Effects of population size on seed production and germinability in an endangered, fragmented grassland plant. Conserv. Biol. 13, p. 266–273.
Opdam P., Van Apeldoorn R., Schotman A., Kalkhoven J. (1993). Population responses to landscape fragmentation. In Vos CC., Opdam P. (Eds.). Landscape ecology of a stressed environment. London: Chapman & Hall, p. 147–171.
Ouborg J. (1993). Isolation, population size and extinction: the classical and metapopulation approaches applied to vascular plants along the Dutch Rhine-system. Oikos 66, p. 298–308.
Piessens K., Honnay O., Hermy M. (2005). The role of fragment area and isolation in the conservation of heathland species. Biol. Conserv. 122, p. 61–69.
Poschlod P., WallisDeVries MF. (2002). The historical and socio-economic perspective of calcareous grasslands – lessons from the distant and recent past. Biol. Conserv. 104, p. 361–376.
Vandermotten C., Decroly JM. (1995). Géographie. In Archambeau N., Joris F. (Eds) Wallonie. Atouts et références d’une Région. Gouvernement wallon. Namur. (http://www.wallonie-en-ligne.net/1995_Wallonie_Atouts-References/1995_ch04-1_Vandermotten-Chr_ Decroly-J-M.htm).
Young A., Boyle T., Brown T. (1996). The population genetics consequences of habitat fragmentation for plants. Trends Ecol. Evol. 11, p. 413–418.
