


- Accueil
- Volume 27 (2023)
- Numéro 4
- Morphological traits and sustainability of plus shea trees (Vitellaria paradoxa C.F.Gaertn.) in Côte d'Ivoire


Visualisation(s): 1759 (78 ULiège)
Téléchargement(s): 109 (0 ULiège)

Morphological traits and sustainability of plus shea trees (Vitellaria paradoxa C.F.Gaertn.) in Côte d'Ivoire

Document(s) associé(s)
Version PDF originaleRésumé
Caractères morphologiques et durabilité des arbres plus de karité (Vitellaria paradoxa C.F.Gaertn.) en Côte d’Ivoire
Description du sujet. Le karité (Vitellaria paradoxa C.F.Gaertn.) est une plante essentielle des savanes naturelles/semi-naturelles et des systèmes agroforestiers d'Afrique, contribuant aux revenus des ménages ruraux et représentant une importante culture de rente pour l'exportation. Il est donc important d'améliorer nos connaissances sur la viabilité et la diversité génétique des génotypes des « Arbres Plus de Karité » (APK : arbres à karité dont la supériorité génétique n'a pas encore été prouvée par un test de descendance approprié) afin de conserver la diversité génétique et d’améliorer le potentiel de production de cette espèce.
Objectifs. Nous avons caractérisé la diversité de la collection in situ de APK en Côte d'Ivoire et évalué sa durabilité à long terme.
Méthode. Douze traits morphologiques qualitatifs ont été étudiés sur 220 APK échantillonnés au hasard parmi 405 APK également sélectionnés aléatoirement dans les collections in situ des districts de la Bagoué et du Tchologo dans le nord de la Côte d'Ivoire, afin d'évaluer la durabilité de la conservation in situ.
Résultats. La plupart des traits morphologiques qualitatifs étaient très diversifiés d'après l'indice de diversité de Shannon (varie de 0,55 à 0,98) avec des différences significatives entre les districts. La classification ascendante hiérarchique a permis de classer les spécimens APK en trois groupes. Il est important de noter que nous avons confirmé que la collecte in situ n'était pas une option réalisable pour la conservation à long terme de cette espèce car elle est menacée par un taux de mortalité élevé (8,15 % après trois ans) et des niveaux élevés d'infestation par les Loranthaceae (83,68 %).
Conclusions. Ainsi, nous recommandons l'établissement d'une collection ex situ d'arbres à karité, gérée par une organisation de recherche, afin de sauvegarder la durabilité et la préservation de cet important patrimoine génétique. Pour maximiser la diversité génétique de la future collection ex situ recommandée, nous suggérons d'établir une collection noyau basée sur la diversité moléculaire caractérisée par des marqueurs moléculaires, tels que le polymorphisme d’un seul nucléotide (SNP).
Abstract
Description of the subject. Shea tree (Vitellaria paradoxa C.F.Gaertn.) is an essential component of natural/semi-natural savannas and agroforestry systems of Africa. It contributes to local household incomes and represents an important cash crop for export. It is important to improve our knowledge about the viability and genetic diversity of “plus shea tree” (PST: shea trees whose genetic superiority has not yet been proven by an appropriate progeny test) genotypes to conserve the genetic diversity of the species and the potential for producing improved species.
Objectives. We characterized the diversity of an in situ PST collection in Côte d’Ivoire and evaluated its long-term sustainability.
Method. Twelve qualitative morphological traits were studied on 220 PSTs randomly sampled among 405 PSTs also randomly selected from in situ collections in the Bagoué and Tchologo districts of northern Côte d’Ivoire to assess in situ conservation sustainability.
Results. Most qualitative morphological traits were highly diverse based on the Shannon diversity index (0.55-0.98 range) with significant differences between districts. Hierarchical ascending classification gathered the PSTs into three groups. The lack of sustainability of the in situ collection for long-term conservation of the species was confirmed because it is endangered by a high mortality rate (8.15% after three years) and high levels of infestation by Loranthaceae (83.68%).
Conclusions. We recommend the establishment of an ex situ collection of shea trees managed by a research organization to preserve this important genetic heritage. To maximize the genetic diversity of the future ex situ collection, we suggest establishing a core collection based on molecular diversity characterized from molecular markers, such as single nucleotide polymorphism (SNP).
Table des matières
Received 26 April 2023, accepted 29 August 2023, available online 25 September 2023.
This article is distributed under the terms and conditions of the CC-BY License (http://creativecommons.org/licenses/by/4.0)
1. Introduction
1Shea butter tree or shea tree (Vitellaria paradoxa C.F.Gaertn.) is an indigenous African tree species belonging to the Sapotaceae family. It grows naturally within a wide belt of more than 3.4 million km2 across 21 countries between western Senegal and eastern Uganda, and it supports an estimated 16.2 millions of shea nut collectors (Sallé et al., 1993; Hall et al., 1996; Glew & Lovett, 2014; Naughton et al., 2015). The species is taxonomically subdivided into two subspecies, namely V. paradoxa subsp. paradoxa found in West and Central Africa, and V. paradoxa subsp. nilotica, located in East Africa. The tree grows up to 20 m height and 1 m diameter at breast height, and has a lifespan of more than 200 years. In Côte d'Ivoire, shea tree populations grow in the semi-arid zone of the savanna region (extreme north of the country), similar to other sub-Saharan African countries; however, some populations also occupy the pre-forest transition zone thanks to their acquired high level of adaptation (Diarrassouba et al., 2007). In theory, these adaptation processes should generate higher genetic richness in Côte d'Ivoire compared to other countries where this plant is restricted to the semi-arid zone of the savanna regions.
2The locations from which shea tree germplasm was introduced to other countries in Africa are not well known because of changes in vegetation. The species probably spread out from refugia. Furthermore, human management indicates ongoing semi-domestication (Allal et al., 2011). The current diversity and distribution of shea trees reflects a long history of evolution and responses to climate change and other environmental factors that determine the morphology of its organs (stem, leaf, flower, fruit, and seed) (Allal et al., 2011). The large distribution of shea was doubled by practicing allogamy as a mode of reproduction, but this practice might have led to high intra-specific diversity (Diarrassouba et al., 2007).
3Shea tree is a socio-economically important species for many traditional communities in Africa. It provides substantial income to rural households in Western and Central Africa. Because women predominantly collect shea fruit for transformation, it is considered as a “female crop” in all production areas. Several tree parts are used in rituals, pharmaceutics, cosmetics, construction, and food products (Diarrassouba et al., 2009). The fruit and associated main derived product – shea butter (“beurre de karité” in French) – are used for local consumption and for trade on local and international markets. The concept of exporting shea kernels to Europe on a large scale was first introduced by French colonial administrations in the early 20th century. However, at the end of the 1950s, low shea production led shea collectors and processors to use the kernels and butter for their own domestic consumption and/or for sale on local periodic and cross-border markets (Wardell et al., 2021a). It was only after independence in 1960 that the new international demand for edible cocoa butter equivalents (CBEs) led to the introduction of several efforts to regulate and control the shea trade through stabilization funds and parastatal marketing boards (Wardell et al., 2021b).
4These products provide enormous benefits and contribute to national economies. The export of raw shea kernel and shea butter to international markets in Europe, Asia, and the USA has risen in recent years (by 25% between 1994 and 2004) and is expected to keep rising (GEPA, 2014; Bello-Bravo et al., 2015; Elias, 2015; Rousseau et al., 2015; Jasaw et al., 2017). Shea kernels are processed to manufacture a wide range of food products (including chocolate) and cosmetics (Schreckenberg et al., 2006). However, shea trees face various threats, including climate change, emerging bioagressors, natural regeneration issues, demographic pressure, bushfires, overexploitation, changing land uses, and cutting for firewood and charcoal (Boffa et al., 1996). Consequently, shea tree was listed as a “vulnerable species” by the International Union for Conservation of Nature (IUCN) in 1998 and is expected to be upgraded to “endangered species” in the future. The function of genetic variation studies in assisting plant conservation approaches and improving cultivars has long been recognized (Ramanatha Rao & Hodgkin, 2002). High genetic diversity is key to the adaptation potential to current and future biotic and abiotic constraints in the context of global change (Lassois et al., 2016).
5Despite the socio-economic importance of shea butter as a multi-purpose product and the risk of shea tree becoming an endangered species, key knowledge gaps remain regarding its diversity, optimal conservation approaches, and ways of improving its resilience. In Côte d'Ivoire, as in many producing countries, the genetic resources of shea trees are conserved and maintained in the field by farmers. This diversity is an important underexploited heritage, but it is increasingly under threat. Therefore, it is essential to protect and conserve shea trees in a sustainable way. Genetic and morphological trait studies are required to manage the genetic resources of shea trees effectively. In particular, the development of national and regional plans to conserve their genetic resources should be promoted (Boffa, 2015). Following this author’s recommendations and based on a participatory survey, the shea breeding program of Côte d'Ivoire identified 639 “plus shea trees” (PSTs) (shea trees whose genetic superiority has not yet been proven by an appropriate progeny test) in the Bagoué and Tchologo districts (Diarrassouba et al., 2017). These PSTs constitute part of the in situ collection of shea trees in Côte d’Ivoire, in place since 2017.
6We characterized the diversity of this collection based on the variation of qualitative morphological traits and evaluated its durability. Our results are expected to provide a basis for relevantly involving the identified PSTs in subsequent hybridization programs, generating high-performance plant material and restoring the shea tree sector in Côte d'Ivoire. In particular, our results are expected to help reinforce and adapt the conservation process of this endangered species.
2. Materials and methods
2.1. Study area
7Côte d'Ivoire is divided into 31 districts with varying cropping patterns, climates, livelihoods, soil types, and natural resources. The country covers 322,462 km² of terrestrial area, and is located in West Africa (4°30' to 10°30' N latitude and 2°30' to 8°30' W longitude). It belongs to the tropical zone. Based on climatic factors and variation in vegetation, the savannas of the northern part of Côte d’Ivoire – where shea trees grow – are subdivided in two main zones called the Sudanian and sub-Sudanian savannas. The Sudanian savanna corresponds to the main shea tree production zone, whereas the sub-Sudanian savanna corresponds to a transition production zone. Our study was carried out in the Sudanian savannas of the Tchologo and Bagoué districts in the northern region of Côte d’Ivoire (Figure 1), where shea trees are a significant component of the farming system. Tchologo district covers 17,382 km², with 467,958 inhabitants (density 26.92 inhabitants·km-²). This district includes three departments: Ferkessédougou, Kong, and Ouangolodougou (Figure 1). In contrast, Bagoué district covers 10,678 km², with 375,687 inhabitants (35.2 inhabitants·km-²). Bagoué district includes three departments: Boundiali, Kouto, and Tengréla (Figure 1).
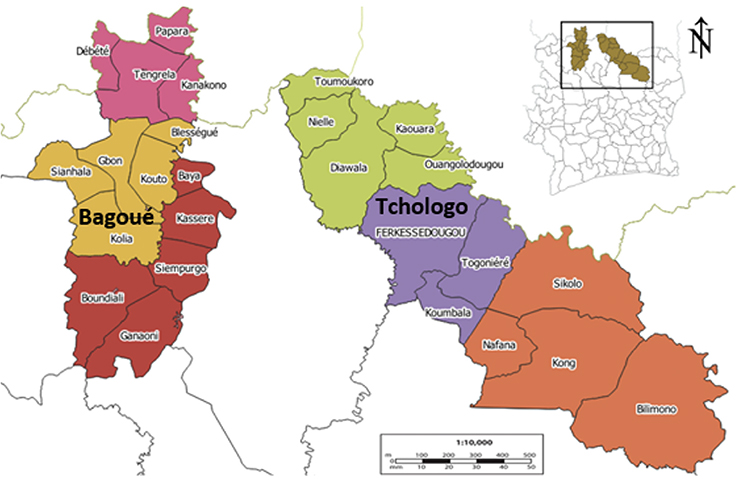
Figure 1. Localization of the two study areas in Côte d’Ivoire. Enlarged image: Bagoué District (left) including Tengrela (pink), Kouto (yellow) and Boundiali (red) departments; Tchologo District (right) including Ouangolodougou (green) Ferkéssédougou (purple) and Kong (orange) departments — Localisation des deux zones d’étude en Côte d’Ivoire. Image agrandie : Région de la Bagoué (à gauche) comprenant les départements de Tengrela (rose), Kouto (jaune) et Boundiali (rouge) ; Région du Tchologo (à droite) comprenant les départements de Ouangolodougou (vert), Ferkéssédougou (violet) et Kong (orange).
8The climate in these two districts is Sudanese, with two main seasons (dry and rainy). The dry season extends from November to April and overlaps with the Harmatan (dry and dusty) season extending between December and February, with temperatures peaking in March and April. The rainy season extends from May to October, and peaks in August and September. Annual rainfall is around 1,200 mm (Brou, 2005). The annual average temperature is 27 °C. The vegetation is Sudano-Guinean, dominated by wooded and grassy savannas (N’Guessan et al., 2015).
2.2. Plant material
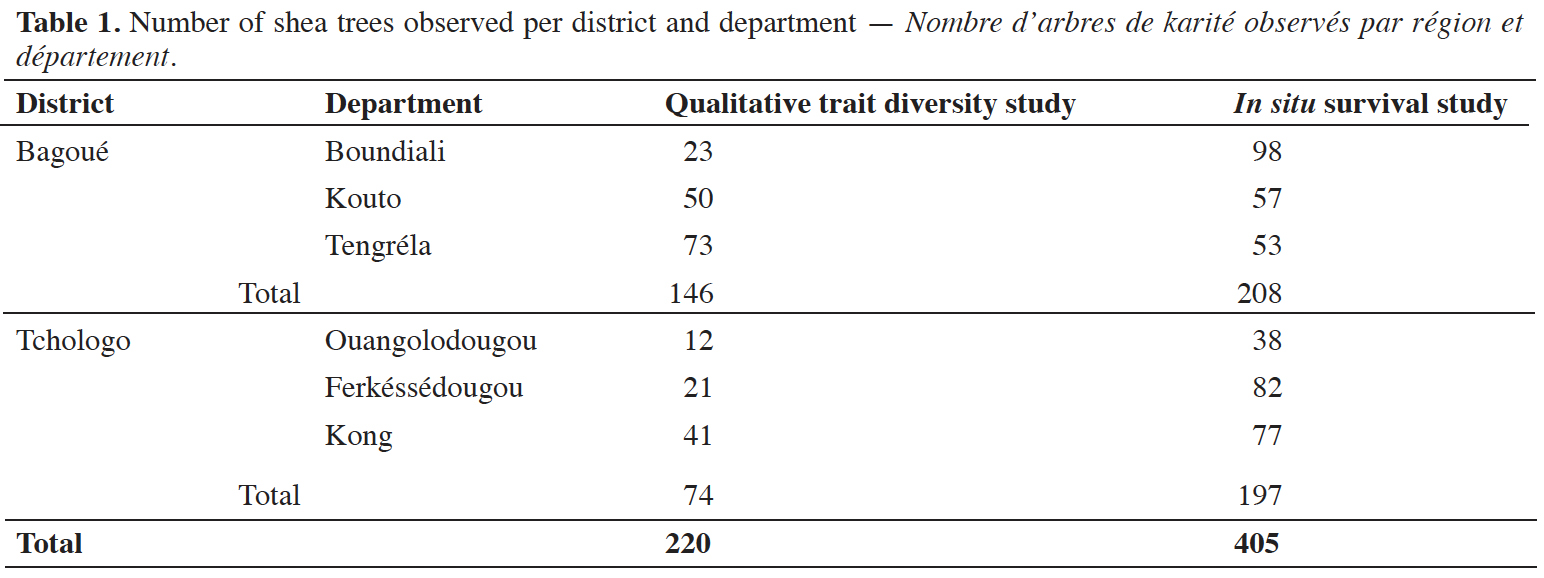
9The shea trees considered in this study were mature trees regenerated/protected by farmers, probably for centuries (Wardell et al., 2021a) and belonged to the subspecies paradoxa. They had been previously designated as “plus shea trees” (PSTs), i.e. identified by the Shea breeding program of the University of Peleforo Gon Coulibaly (Côte d’Ivoire) in collaboration with the Agence Nationale d'Appui au Développement Rural (ANADER, Côte d’Ivoire), following an extensive survey between February and June 2017. This program consisted of a participatory survey with farmers allowed to select PSTs based on specific criteria, including high fruit yield, sweet taste of the fruit pulp, large fruit size, early flowering every year, and periodicity of fruit production. Six hundred and thirty-nine shea trees – considered as “plus” trees – were selected, geo-referenced for future identification and conserved in situ on the land plots of 400 farmers in the Bagoué and Tchologo districts. Four hundred and five PSTs were randomly sampled in the two districts to evaluate their capacity for in situ survival. Then, 200 PSTs were randomly sampled among these 405 PSTs, and 20 more PSTs from Tengréla Department were selected for diversity analyses of qualitative traits (Table 1).
2.3. Analysis of qualitative traits diversity
10Twelve qualitative morphological variables were selected based on the minimum descriptors of shea trees as defined by the International Plant Genetic Resources Institute (IPGRI, INIA, 2006). These traits were evaluated on 220 randomly selected PSTs (Table 1). The descriptors were observed on trees, branches, healthy adult and young fresh leaves, and seeds, as listed in table 2. Five descriptors were observed from 10 leaves randomly selected from the crown. The two seed-related descriptors were observed from 10 seeds of each individual tree. Inflorescence density was determined by looking at the presence and density of flowers on the terminal branches. The remaining four descriptors were based on the crown shape, the branching pattern, the tree growth habit and the branch density.

11The data were analyzed using Rstudio version 4.2.2. The normalized Shannon-Weaver diversity index (H') (range: 0 to 1) was used to assess the diversity of each qualitative trait (Yao et al., 2015). It was calculated as follows:
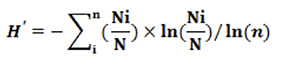
12where Ni is the number of individuals observed for a given modality and a given descriptor, N is the total number of individuals, and n is the number of modalities for a given characteristic. When H' is close to 1, the diversity of a given qualitative trait is high in all populations (Yao et al., 2015). The equal probability of the frequencies (pi = Ni/N) of appearance of the modalities of a given qualitative trait was tested using the χ2 test at the 5% threshold. It was also used to test the homogeneity of the proportion of descriptor modalities between the Bagoué and Tchologo districts. A significant result indicated that at least two of the observed frequencies were different. Multiple correspondence analysis (MCA) and hierarchical ascending classification (HAC) structured the PSTs based on the qualitative descriptors. MCA is a multivariate descriptive method that groups variables according to the variability that they explain in a given population. This method allowed us to generate a table of the contribution to the inertia showing the degree of linkage between variables and components. The choice of axes was made on the basis of the elbow method. HAC was carried out using the unweighted paired groups method of analysis (UPGMA) to cluster shea trees based on their morphological diversity.
2.4. Evaluation of the sustainability of the in situ PST collection
13The sustainability of the in situ collection was evaluated based on two parameters of the randomly selected 405 PSTs (Table 1). The mortality rate was monitored over the three years following initial PST identification and selection by the Shea breeding program of the University of Peleforo Gon Coulibaly (Côte d’Ivoire) in collaboration with the Agence Nationale d'Appui au Développement Rural (ANADER, Côte d’Ivoire) in 2017. The parasitic plant Loranthaceae was the main biotic threat to the survival of the in situ collection. Its occurrence was quantified for each PST.
3. Results
3.1. Diversity of qualitative traits
14Diversity of growth descriptors
15Crown shape (CRS). The six crown shape phenotypes were distributed unevenly. The broadly pyramidal phenotype was most common (26.36%; Figure 2). The other phenotypes (elliptical, oblong, spherical, semicircular, and pyramidal) represented 18.18%, 17.27%, 15.45%, 13.18%, and 9.55% of total phenotypes, respectively. The χ2 test was highly significant (χ2 = 21.26; p = 0.001). The Shannon-Weaver index was high (H' = 0.97; Table 3). The proportions of CRS modalities differed significantly in the Bagoué and Tchologo districts (χ ² = 12.05; p = 0.034).
 Figure 2. Crown shape diversity of plus-shea trees from the Bagoué and Tchologo districts in northern Côte d’Ivoire — Diversité des formes de la cime des arbres plus de karité des régions de la Bagoué et du Tchologo dans le nord de la Côte d’Ivoire.
Figure 2. Crown shape diversity of plus-shea trees from the Bagoué and Tchologo districts in northern Côte d’Ivoire — Diversité des formes de la cime des arbres plus de karité des régions de la Bagoué et du Tchologo dans le nord de la Côte d’Ivoire.
Left to right: broadly pyramidal (26.36%), elliptical (18.18%), oblong (17.27%), spherical (15.45%), semicircular (13.18%) and pyramidal (9.55%) phenotypes — De gauche à droite : phénotypes largement pyramidal (26,36 %), elliptique (18,18 %), oblong (17,27 %), sphérique (15,45 %), semi-circulaire (13,18 %) et pyramidal (9,55 %).
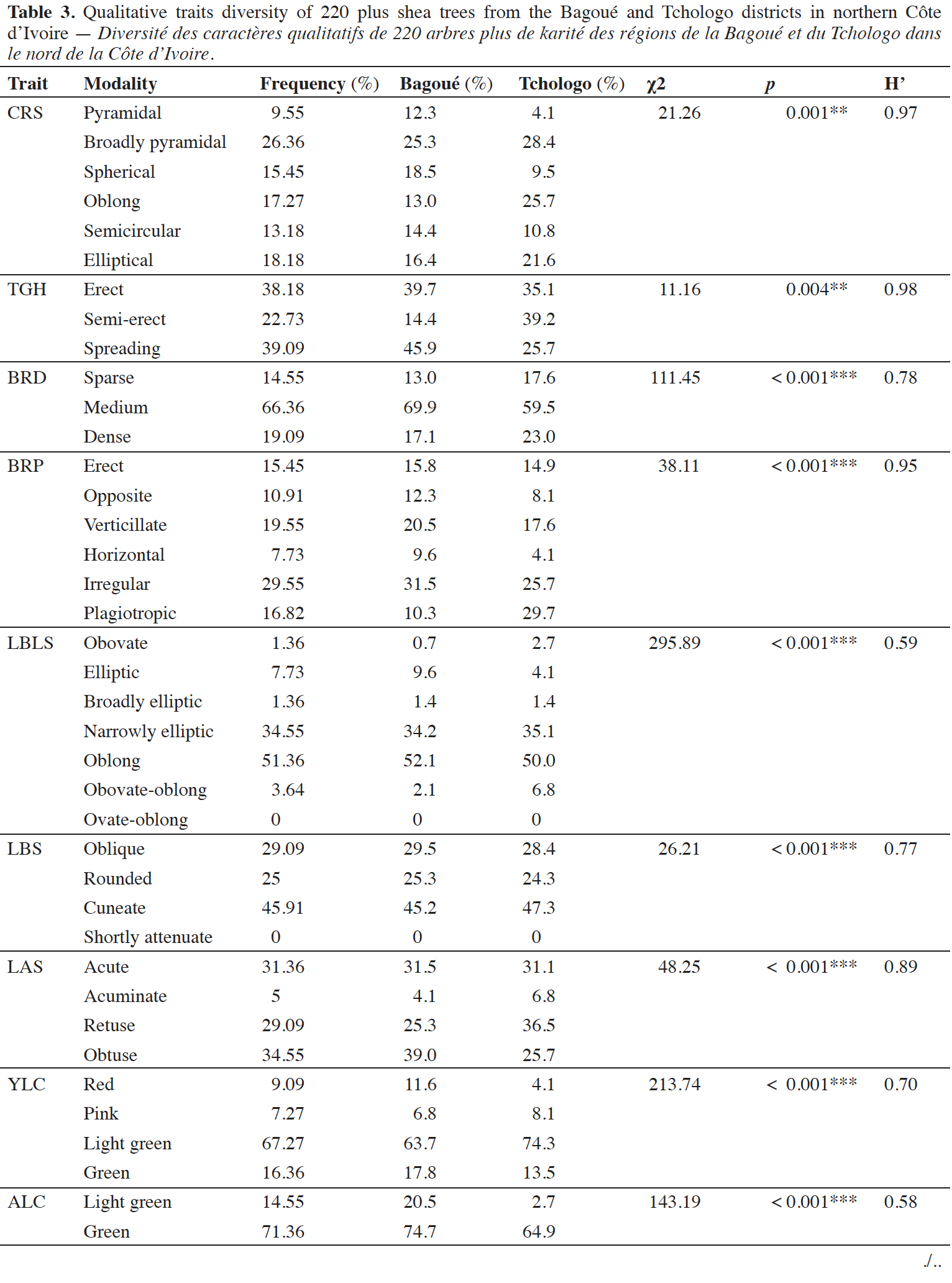
16Tree growth habit (TGH). The χ2 test was significant (χ2 = 11.16; p = 0.004) for the three phenotypes describing tree bearing. Extended and erect phenotypes were most common (38.18% and 39.09% of the population, respectively). In comparison, the semi-erect phenotype represented 22.73% (Table 3) of the total trees. The Shannon-Weaver index was H' = 0.98. The proportions of TGH modalities differed significantly in the Bagoué and Tchologo districts (χ ² = 18.7; p < 0.001).
17Branching density (BRD). Three types of branch density (sparse, medium, dense) were identified, and occurred at unequal frequencies (χ2 = 111.45; p ˂ 0.001). The medium-density phenotype was prevalent (69.97%; Table 3). The Shannon-Weaver index was H’ = 0.78. The proportions of all BRP modalities were statistically similar in the Bagoué and Tchologo districts (χ ² = 2.38; p = 0.3).
18Branching pattern (BRP). The irregular phenotype was most common (29.55%) among the branching pattern phenotypes. The verticillate, plagiotropic, erect, opposite and horizontal phenotypes represented 19.55%, 16.82%, 15.54%, 10.91% and 7.73% of the total, respectively. The Shannon-Weaver index was H' = 0.95. The proportions of the BRP modalities differed significantly in the Bagoué and Tchologo districts (χ2 = 14.62; p = 0.01). Plagiotropism of the BRP was higher in Tchologo district (29.7%) than in Bagoué district (10.3%). The others modalities were statistically similar.
19Diversity of leaf descriptors
20Leaf blade shape (LBLS). Five leaf blade shapes were observed (obovate, elliptic, broadly elliptic, narrowly elliptic, and oblong), while the obovate-oblong shape was not observed. Oblong and narrowly elliptic leaves were most frequent (51.36% and 34.55%, respectively; Table 3). The Shannon-Weaver index was H' = 0.59. The LBLS modalities were statistically similar in the Bagoué and Tchologo districts (χ2 = 6.45; p = 0.26).
21Leaf base shape (LBS). Three out of four types of leaf base shape were observed (oblique, rounded, and cuneate) in varying proportions (χ2 = 26.21; p ˂ 0.001). The short attenuated type was not observed. The wedge-shaped phenotype was most common (45.91%). The other two phenotypes (rounded and oblique) represented 25% and 29.09% of total phenotypes, respectively (Table 3). The Shannon-Weaver index was H' = 0.77. The proportions of all LBS modalities were statistically similar in the Bagoué and Tchologo districts (χ2 = 0.09; p = 0.96).
22Leaf apex shape (LAS). The four phenotypes of apex shape (obtuse, acute, rounded, acuminate) were present at varying frequencies (χ2 = 48.25; p ˂ 0.001), with a Shannon-Weaver index H' = 0.89. The acute, rounded, and obtuse shapes were most common (31.36%, 29.09%, and 34.55% of the total, respectively). The acuminate phenotype was least common (5%). The proportions of all LAS modalities were statistically similar in the Bagoué and Tchologo districts (χ2 = 5.33; p = 0.17).
23Young leaf color (YLC). Four colors (green, light green, pink, red) of young leaves were observed at varying proportions (χ2 = 213.74; p ˂ 0.001), with a Shannon-Weaver index H' = 0.70. The predominantly light green phenotype was most prevalent (67.27%), followed by the green phenotype (16.36%), and the red (9.09%) and pink (7.27%) phenotypes (Table 3). The proportions of all YLC modalities were statistically similar in the Bagoué and Tchologo districts (χ2 = 4.6; p = 0.2).
24Adult leaf color (ALC). Three colors (light green, green, dark green) were observed at varying proportions on adult leaves (χ2 = 143.19; p ˂ 0.001), with a Shannon-Weaver index H' = 0.58. Pinkish green leaves were not observed. The green, light green and dark green phenotypes represented 71.36%, 14.55%, and 14.09% of total phenotypes, respectively (Table 3). The light green and dark green modalities differed significantly in Bagoué district vs Tchologo district. Light green was higher in Bagoué (20.5%) than in Tchologo (2.7%), whereas dark green was higher in Tchologo (32.4%) than in Bagoué (4.8%).
25Diversity of inflorescence descriptors
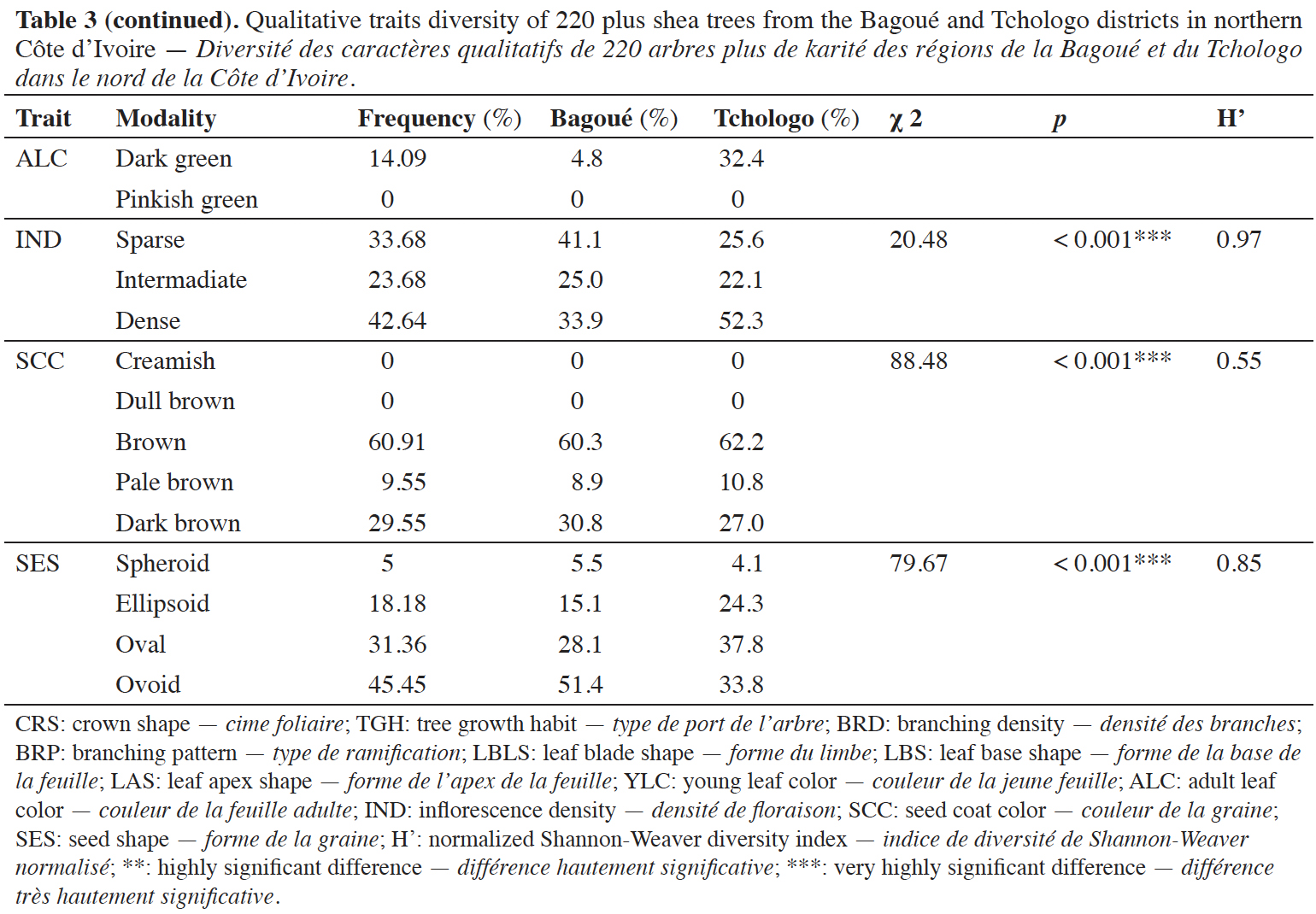
26Inflorescence density (IND). Dense, intermediate, and sparse inflorescence densities represented 42.64%, 23.68%, and 33.68% of the phenotypes, respectively (χ2 = 13.71; p = 0.0001) and the Shannon-Weaver index was H' = 0.97 (Table 3). The proportion of IND modalities differed significantly in the Bagoué and Tchologo districts (χ2 = 19.59; p < 0.001).
27Dense and sparse flowering types dominated in Tchologo (55.55%) and Tengréla (56.25%) departments, respectively (Figure 3).
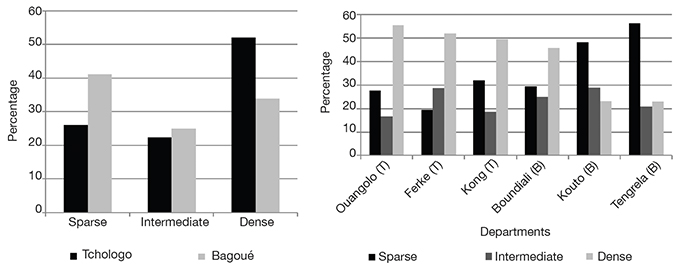 Figure 3. Diversity of the flowering characteristics in plus shea trees — Diversité des caractères de floraison des arbres plus de karité.
Figure 3. Diversity of the flowering characteristics in plus shea trees — Diversité des caractères de floraison des arbres plus de karité.
Ouangolo: Ouangolodougou; Ferké: Ferkéssédougou; T: Tchologo; B: Bagoué.
28Diversity of seed descriptors
29Seed coat color (SCC). Three of the five possible phenotypes (brown, pale brown, dark brown) were present in both districts (χ2 = 88.48; p ˂ 0.001). Creamish and dull brown seed coat colors were not observed. The brown phenotype was most common (60.91%), followed by dark brown (29.54%) and pale brown (9.55%) (Figure 4; Table 3). The Shannon-Weaver index was H' = 0.55. The proportions of all SCC modalities were statistically similar in the Bagoué and Tchologo districts (χ2 = 0.46; p = 0.8).
 Figure 4. Seed color — Couleur des graines.
Figure 4. Seed color — Couleur des graines.
Left, brown; middle, dark brown; right, pale brown— à gauche, marron ; au milieu, marron foncé ; à droite, marron clair.
30Seed shape (SES). Three out of four seed shape phenotypes were observed at varying proportions (χ2 = 79.67; p ˂ 0.001). The ovoid and oval forms were most common (49.54% and 31.82%, respectively), followed by the ellipsoid phenotype (18.64%). The spheroid shape was not observed. The Shannon-Weaver index was H' = 0.85. The proportions of all SES modalities were statistically similar in the Bagoué and Tchologo districts (χ2 = 7.34; p = 0.06).
3.2. Structuring and clustering based on PST qualitative traits
31The multiple correspondence analysis (MCA) of the qualitative descriptors showed that the total inertia was 3.1, with 44 active modalities. The elbow method was applied to the 34 eigenvalues obtained in the MCA to select the first three factorial axes for the analysis. A significant jump was detected just after the third eigenvalue; therefore, the three factorial planes formed by the first three factorial axes of the MCA were used. These three axes represented 18.4% of total inertia in the plot, while axes 1, 2 and 3 represented 6.5%, 6.4%, and 5.5% of the total, respectively. The representation of variables in the factorial planes showed that axis 1 crossed with axis 2 and axis 3, and axis 2 crossed with axis 3, representing 12.35%, 11.7%, and 11.33% of reality, respectively. For each axis, the average theoretical percentage of inertia explained by each modality was 2.27% (100%/44). However, the relative contribution of each modality to each axis varied considerably (axis 1: 0-17%; axis 2: 0-11.25%; axis 3: 0-13.25%). Axis 1 was mostly explained by CRS and certain leaf variables (LBLS, LAS, YLC, and ALC), axis 2 by LAS, LBLS, and certain growth descriptors (CRS, TGH, BRD, and BRP), and axis 3 by certain growth (BRP, TGH, and CRS) and leaf (ALC and YLC) descriptors. The variables contributing to the formation of the different axes are showed in figure 5. Following this analysis, we used the nine descriptors listed in table 4 to perform a hierarchical ascending classification (HAC).
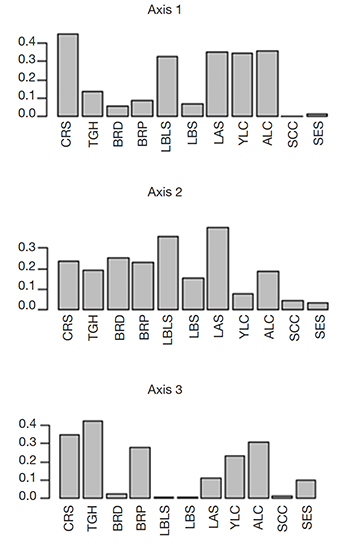 Figure 5. Contribution of the qualitative traits to axes 1, 2 and 3 of the multiple correspondence analysis (MCA) — Contribution des caractères qualitatifs aux axes 1, 2 et 3 de l’analyse des correspondances multiples.
Figure 5. Contribution of the qualitative traits to axes 1, 2 and 3 of the multiple correspondence analysis (MCA) — Contribution des caractères qualitatifs aux axes 1, 2 et 3 de l’analyse des correspondances multiples.
CRS, TGH, BRD, BRP, LBLS, LBS, LAS, YLC, ALC, SCC, SES: see table 3 — voir tableau 3.
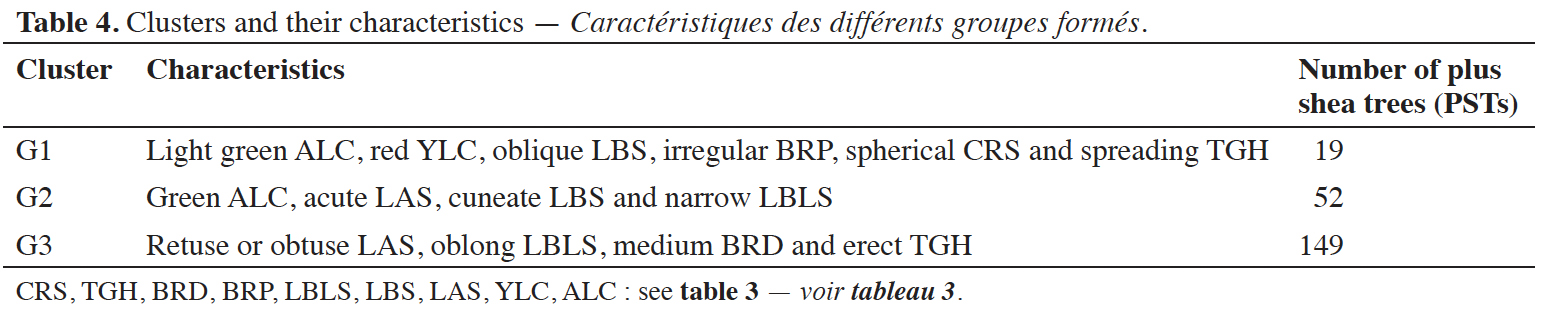
32HAC of 220 PSTs separated them into different classes according to their degree of similarity based on morphological qualitative traits. The dendrogram obtained from Gower’s distance and the unweighted paired groups method of analysis (UPGMA) delineated three clusters (Figure 6). These three phenotypic groups represented the morphological characteristics of the PST population:
33– Group 1 was characterized by ‘Light Green Adult Leaf’, ‘Red Young Leaf’, ‘Oblique Leaf Blade’, ‘Irregular Branching Patterns’, ‘Spherical Crown’ and ‘Spreading Tree Growth Habit’;
34– Group 2 was characterized by ‘Green Adult Leaf’, ‘Acute Leaf Apex’, ‘Cuneate Leaf Blade’ and ‘Narrow Leaf Blade Shape’;
35– Group 3 included ‘Retuse or Obtuse Leaf Apex’, ‘Oblong Leaf Blade’, ‘Medium Branch Density’ and ‘Erect Tree Growth Habit’.
36The Wilks test was significant (alpha = 0.01, p < 0.0001), rejecting the null hypothesis of equality of the three vector groups, whose characteristics are presented in table 4.
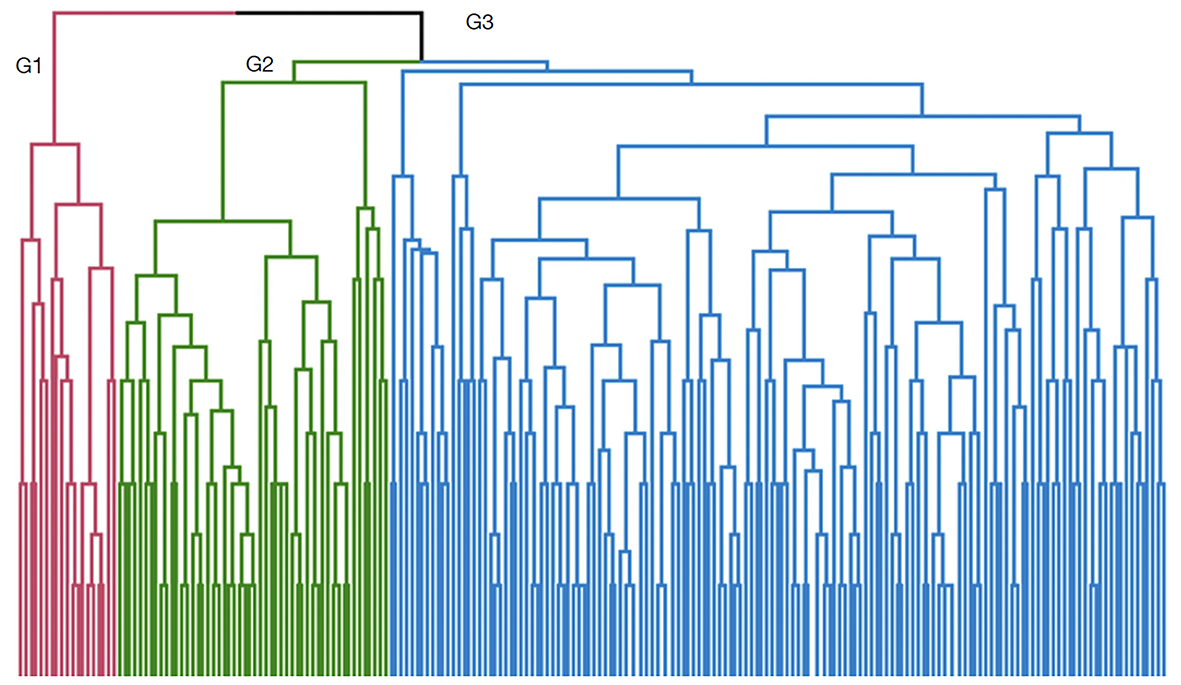
Figure 6. Clustering of the plus-shea trees (PSTs) sample based on morphological traits using the UPGMA method — Classification des arbres plus de karité (APK) sur la base des caractères morphologiques avec la méthode UPGMA.
37Figure 7 shows the dispersion of individuals in the three groups in factorial plans 1 and 2. Distinct groups were not obtained when the Bagoué and Tchologo districts were used as grouping variables, based on the dispersion of individuals in the first two factorial planes (Figure 8).
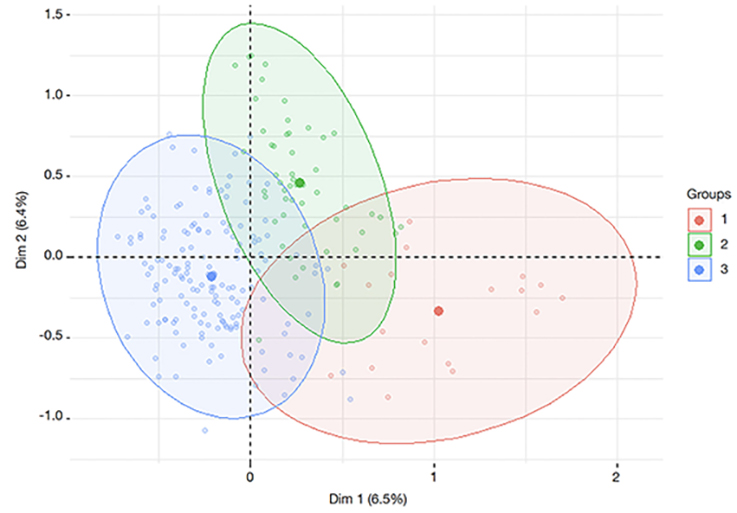
Figure 7. Distribution of individuals from the three groups of plus shea trees (PSTs) from the Bagoué and Tchologo districts — Répartition des individus des trois groupes d’Arbres Plus de Karité (APK) des régions de la Bagoué et du Tchologo.
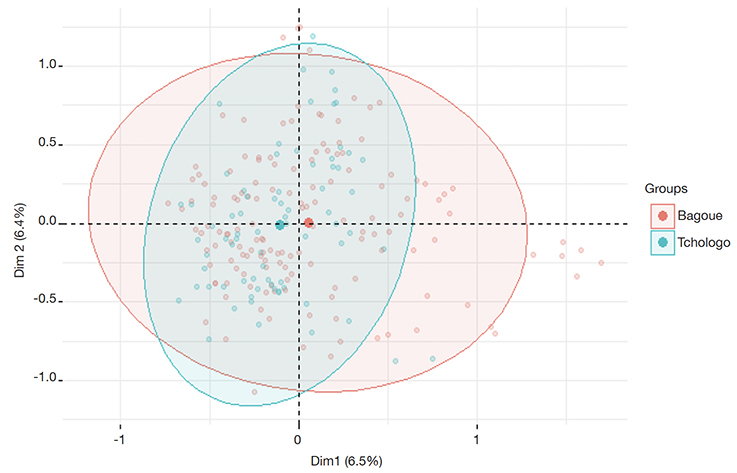
Figure 8. Distribution of individuals of plus shea trees (PSTs) using Bagoué and Tchologo districts as groups — Répartition des arbres Plus de karité (APK) en utilisant les régions de la Bagoué et du Tchologo comme groupes.
3.3. Evaluation of the sustainability of the in situ PST collection
38Trees surviving three years after the setting up of in situ conservation. Three years after PSTs were identified and in situ conservation was established, 33 out of 405 trees had died (8.15% of the collection). A higher mortality rate was recorded in Bagoué district (11.54%) compared to Tchologo district (4.57%). Kouto department (12.07%; Bagoué) had the highest mortality rate, and Kong department (2.60%; Tchologo) had the lowest (Figure 9).
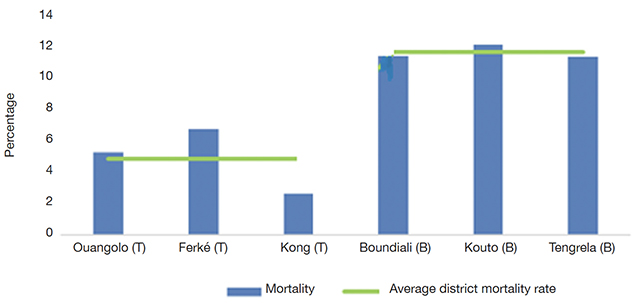
Figure 9. Mortality rate of plus shea trees three years after an in situ collection was set up — Taux de mortalité des arbres plus de karité, trois ans après la mise en place d’une collection in situ.
Ouangolo: Ouangolodougo; Ferké: Ferkéssédougo; T: Tchologo; B: Bagoué.
39Presence of the parasitic plant Loranthaceae. Overall, 83.64% of the PSTs had at least one clump of mistletoe. The PSTs of Tengréla Department were most impacted, with at least one clump per tree. Kong was the least affected department (62.67% infestation rate), followed by Boundiali (73.91%). Overall, the trees of Bagoué district were more infested than those of Tchologo district, but no significant difference was evidenced (85.34% vs 81.91% infestation rates, respectively; Figure 10).
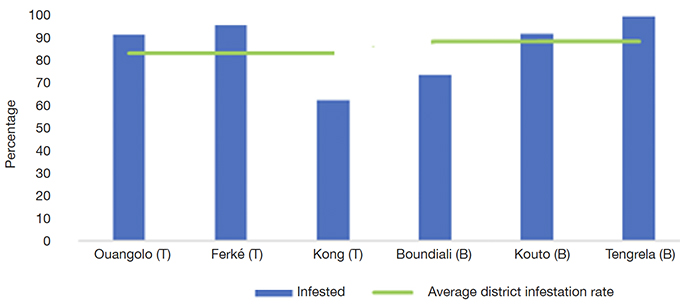
Figure 10. Rates of plus shea trees infestation by Loranthaceae — Taux d’infestation des arbres plus de karité par les Loranthacées.
Ouangolo: Ouangolodougo; Ferké: Ferkéssédougo; T: Tchologo; B: Bagoué.
4. Discussion
40This study demonstrates that the morphological diversity of the qualitative traits of the in situ PST collection of Côte d'Ivoire is high. Most shea tree descriptors (IPGRI, INIA, 2006) displayed high diversity (Shannon diversity index – a reliable indicator of trait diversity – range: 0.55-0.98). However, in view of the descriptors studied in the present work, the Shannon index is a mixture of the number of modalities, the frequencies in each of them and especially the easiness to distinguish the different modalities of each descriptor. For example, distinguishing the 'dark green', 'green' and 'light green' ALCs was difficult because of borderline situations with a qualitative modality as opposed to a quantitative one. Conversely, scoring the 4 YLC modalities (green, light green, pink, and red) was much easier, especially when differentiating between red or pink from (light) green.
41The whole range of diversity of the growth descriptors and inflorescence density was assessed, even though frequencies were variable and unequal. The PST collection was primarily characterized by medium branch density (67%) and intermediate to dense inflorescence density (66%). However, inflorescence density was higher in Tchologo district than in Bagoué district (52.1% vs 33.9% dense flowering, respectively). Lower percentages in Bagoué district might be explained by the low rate of dense flowering in Kouto and Tengréla departments (23% each). The high percentage of medium branch density might be explained by farmers preferentially keeping trees with medium shade to limit potential negative effects like reduced speculative yields in these agro-forestry systems. Shading by shea trees negatively impacted maize grain and straw yields (46.64% and 32.8% reductions, respectively) compared to exposed areas (Saidou, 2013). Four types of crown shape have been identified in shea parkland located at Tengréla (Diarrassouba et al., 2009; Soro et al., 2011). In contrast, we identified six types of crown shape, likely due to our survey area being larger. Crown shape can be modified by agricultural practices, but competition with other trees may also have an effect; however, this explanation is less likely because our study was conducted on isolated trees.
42Certain leaf characteristics were not observed in our study, e.g., ovate-oblong LBLS, short-attenuate LBS, and pinkish-green ALC. Furthermore, even though young leaves exhibited a variety of colors, green dominated (83%) over red/pink (17%). Mature leaves were entirely green, without any pinkish-green hue. LBLS was primarily characterized by narrow elliptic and oblong shapes (86%), while the remaining five types represented only 14%. This study highlights higher trait diversity compared to another study (Diarrassouba, 2008), possibly due to differences in scale, as our study evaluated PST distributions across two major districts as opposed to one park. Only three of the five potential seed colors delineated in the IPGRI were found, with brown seeds dominating (61%). In contrast, all the four seed shape categories were found, with ovoid and oval-shaped seeds dominating (45.45% and 31.36%, respectively). Anyomi et al. (2023) reported variations in shea nut shape and recorded two ellipsoid seed shapes in 86.32% of the samples. This might be explained by farmers preferentially selecting certain characteristics, hence a negative selection pressure on the others. For instance, local communities can define shea varieties according to certain criteria such as fruit size, pulp taste, fruiting period, and seed color (Diarrassouba, 2008).
43Seven of the twelve qualitative descriptors were statistically similar in the Bagoué and Tchologo districts. This could mean that these descriptors are common to the Bagoué and Tchologo districts and could be excluded from a potential future similar study comparing shea trees in these districts. Other descriptors (CRS, TGH, BRP, ALC and IND) were significantly different in the Bagoué and Tchologo districts. These differences were mainly linked to tree growth descriptors and could reveal that shea trees from the two districts differed in their vegetative growth. This type of differentiation is often linked to the farmers' choice to keep the trees on their land, in a crop-dependent manner. These differences could also be explained by the difference in the number of individuals studied in each district (74 individuals in Tchologo; 146 in Bagoué): the small number of modalities increased the differences in their proportions (Table 3).
44Hierarchical ascending classification based on descriptive morphological characteristics was used to structure the trees and generated three clusters. The low proportion of variance explained by the first three axes is a characteristic of the MCA, which generally gives pessimistic measures of extracted information. The results of this analysis should be treated with caution; however, it identified a profile of shea trees according to their characteristics that could be used to identify and classify PSTs using qualitative descriptors. Diarrassouba (2008) also concluded that the qualitative morphological traits of shea trees could be used to discern classes. Our results also showed no genetic difference in the qualitative traits of Bagoué district trees vs Tchologo district trees.
45Our study also confirmed that the in situ collection is endangered because of a high mortality rate and high Loranthaceae infestation (83.64% overall; 85.34% and 81.91% in the Bagoué and Tchologo districts, respectively). Our results corroborate those of Ahamidé et al. (2017), who recorded 87.25% of shea tree infestation by Loranthaceae under field conditions. The PSTs of the Kong and Boundiali departments had the lowest infestation rates (63% and 74%, respectively); however, infestation rates above 90% were measured in other departments. Different levels of shea tree infestation by Loranthaceae have been found among departments (Yao et al., 2020). Furthermore, the shea parklands in northern Côte d’Ivoire are infested by two genera of Loranthaceae, namely, Tapinanthus bangwensis (Engl. & K.Krause) Danser and Agelanthus dodoneifolius (DC.) Polhill & Wiens (Soro et al., 2004; Yao et al., 2020). Loranthaceae seeds are mainly spread by birds that consume their fruit and excrete them in their feces (Yao et al., 2020). Because shea trees are the major tree species of the West African savanna, they serve as resting refuges for many birds; this might explain their high rate of infestation by Loranthaceae. Therefore, Loranthaceae represent a major threat to PSTs; yet, the spread of this plant species cannot be controlled, although it is causing significant damage to cultivated and wild trees and shrubs (Boussim et al., 2012; Azo’O et al., 2013). To reduce damage, annual control actions must be implemented, during which Loranthaceae are manually removed from the in situ PST collection. However, the long distances separating individual trees in the collection would require significant human and financial resources to control this biotic stressor. Another solution would be to group individual trees of interest in an ex situ collection that could be managed and controlled near a research center in Côte d'Ivoire. This approach would also ensure the sustainability of the collection by protecting it from other constraints that negatively affect survival. Thirty-three PSTs were lost in Bagoué and Tchologo over a 3-year period. Consequently, keeping plant genetic resources in situ on farming land poses clear management issues. In five shea parklands in Benin, the number of trees per hectare decreased by 0, 9, 12, 16 and 37 trees in Bohicon, Savé, Parakou, Bembéréké and Kandi, respectively, from 2005 to 2017 (Gnanglè, 2017 cited in Lovett & Phillips, 2018). This represents mortality rates of 0%, 34.62%, 44.44%, 39.02% and 75.51%, respectively.
46Mortality differed between the Bagoué and Tchologo districts (12% and 5%, respectively) over the 3-year period. Farmers stated different reasons for tree mortality in the two districts. In Tchologo district, mortality was attributed to the uprooting of trees by strong winds. In Bagoué district, multiple reasons were given, including uprooting by wind, uprooting by machines during the installation of the new electrification network in the district, and cutting down of trees by some farmers who considered them to hinder crop growth. Therefore, human actions clearly have a strong impact on the loss of shea tree diversity in Côte d’Ivoire. Cubero (1997) previously highlighted that in situ conservation sensu stricto is vulnerable. Because farmers have the right to grow what they want, this preference will always be driven by profitable agriculture (Cubero, 1997). Consequently, the collection in its in situ form is threatened. Therefore, alternative, complementary forms of safeguarding must be implemented that will preserve diversity over the long term. An ideal solution would be to create a core collection in a protected and monitored environment. To accomplish this, an analysis of the genetic diversity of the in situ collection is required using single nucleotide polymorphism (SNP) markers to optimize its genetic diversity (Nguyen et al., 2020). This genetic diversity analysis will be important for long-term improvement of shea trees and future studies using genomic selection like marker-assisted selection (MAS) and genome-wide association study (GWAS) (Hale et al., 2021).
5. Conclusions
47This study demonstrates that the qualitative morphological traits of PSTs display high genetic diversity across two districts in Côte d'Ivoire. This important diversity constitutes a heritage that must be preserved for future generations. However, current in situ conservation efforts are not sufficient to ensure sustainable conservation because farmers are under pressure to produce crops. Biotic constraints (i.e., Loranthaceae infestation on most trees) and abiotic constraints (i.e., wind and human actions) endanger the PST population both in the short and medium term. Monitoring is required to ensure long-term conservation. Based on these results, we recommend the creation of an ex situ collection that could be managed by a research center to guarantee sustainability. Furthermore, we recommend selecting a core collection based on the molecular diversity of PSTs to maximize the genetic diversity of this collection.
Bibliographie
Ahamidé I.D.Y. et al., 2017. Diversité des Loranthaceae et leur impact sur Vitellaria Paradoxa C.F.Gaertn. : un fruitier à grande valeur socio-économique au Nord-Bénin. Eur. Sci. J., 13(24), 217-217, doi.org/10.19044/esj.2017.v13n24p217
Allal F. et al., 2011. Past climate changes explain the phylogeography of Vitellaria paradoxa over Africa. Heredity, 107(2), 174-186, doi.org/10.1038/hdy.2011.5
Anyomi W.E. et al., 2023. Heritability and genetic advance estimates of key shea fruit traits. Agronomy, 13(3), 640, doi.org/10.3390/agronomy13030640
Azo’O J.R.N., Tchatat M., Mony R. & Dibong S.D., 2013. Parasitisme et ethnobotanique des Loranthaceae à Lokomo (Est-Cameroun). J. Anim. Plant Sci., 19, 2923-2932.
Bello-Bravo J., Lovett P.N. & Pittendrigh B.R., 2015. The evolution of shea butter’s “Paradox of paradoxa” and the potential opportunity for Information and Communication Technology (ICT) to improve quality, market access and women’s livelihoods across rural Africa. Sustainability, 7(5), 5752-5772, doi.org/10.3390/su7055752
Boffa J.-M., 2015. Opportunities and challenges in the improvement of the shea (Vitellaria paradoxa) resource and its management. Occasional Paper 24. Nairobi: World Agroforestry Centre.
Boffa J.-M., Knudson D.M., Yameogo G. & Nikiema P., 1996. Shea nut (Vitellaria paradoxa) production and collection in agroforestry parklands of Burkina Faso. In: Leakey R.R.B., Temu A.B., Melnyk M. & Vantomme P., eds. Domestication and commercialization of non-timber forest products in agroforestry systems. Non-Wood Forest Products 9. Roma: FAO, 110-122.
Boussim I.J. et al., 2012. Prospects for an integrated control of Loranthaceae species parasitizing Vitellaria paradoxa C.F.Gaertn in Burkina Faso. Int. J. Biol. Chem. Sci., 6(1), 355-364, doi.org/10.4314/ijbcs.v6i1.31
Brou Y.T., 2005. Climat, mutations socio-économiques et paysages en Côte d’Ivoire. Mémoire de synthèse des activités scientifiques présenté en vue de l’obtention de l’Habilitation à Diriger des Recherches : Université des Sciences et Techniques de Lille (France).
Cubero J.I., 1997. Conservation through plant breeding and in situ conservation. Bocconea, 7, 207-212.
Diarrassouba N., 2008. Caractérisation morphologique chez le karité : Vitellaria paradoxa C.F.Gaertn. (Sapotaceae) et analyse de certains facteurs anthropiques affectant la structure de la diversité génétique des peuplements en Côte d’Ivoire. Thèse de doctorat : Université d'Abidjan-Cocody (Côte d'Ivoire).
Diarrassouba N., N’Guessan A., Kouablan K. & Abdourahamane S., 2007. Évaluation des performances de quelques descripteurs quantitatifs et leur utilisation dans la structuration de la population d’un parc naturel de karité en Côte d’Ivoire. Plant Genet. Ressour. Newsl., 152, 65-72.
Diarrassouba N. et al., 2009. Typology of shea trees (Vitellaria paradoxa C.F.Gaertn.) using morphological traits in Côte d’Ivoire. Gene Conserve, 33, 752-780.
Diarrassouba N., Yao S.D.M. & Traoré B., 2017. Identification participative et caractérisation des arbres élites de karité dans la zone de production en Côte d’Ivoire. (Rapport d’étape N°2 No. N° 069/2016.), projet FIRCA/Karité. Éditeur ?
Elias M., 2015. Gender, knowledge-sharing and management of shea (Vitellaria paradoxa) parklands in central-west Burkina Faso. J. Rural Stud., 38, 27-38, doi.org/10.1016/j.jrurstud.2015.01.006
GEPA (Ghana Export Promotion Authority), 2014. GEPA Export. Portal. Compléter la réf.
Glew D. & Lovett P.N., 2014. Life cycle analysis of shea butter use in cosmetics: from parklands to product, low carbon opportunities. J. Clean. Prod., 68, 73-80, doi.org/10.1016/j.jclepro.2013.12.085
Hale I. et al., 2021. Genomic resources to guide improvement of the shea tree. Front. Plant Sci., 12, 720670.
Hall J.B. et al., 1996. Vitellaria paradoxa: a monograph. Bangor, UK: School of Agricultural and Forest Sciences, University of Wales.
IPGRI, INIA, 2006. Descriptors for shea tree (Vitellaria paradoxa). Roma: International Plant Resource Institute.
Jasaw G.S. et al., 2017. Ecosystem services trade-offs from high fuelwood use for traditional shea butter processing in semi-arid Ghana. Ecosyst. Serv., 27, 127-138, doi.org/10.1016/j.ecoser.2017.09.003
Lassois L. et al., 2016. Genetic diversity, population structure, parentage analysis, and construction of core collections in the French apple germplasm based on SSR Markers. Plant Mol. Biol. Rep., 34(4), 827-844, doi.org/10.1007/s11105-015-0966-7
Lovett P. & Phillips L., 2018. Agroforestry shea parklands of Sub-Saharan Africa: threats and solutions. Washington, DC: The Word Bank.
Naughton C.C., Lovett P.N. & Mihelcic J.R., 2015. Land suitability modeling of shea (Vitellaria paradoxa) distribution across sub-Saharan Africa. Appl. Geogr., 58, 217-227, doi.org/10.1016/j.apgeog.2015.02.007
N’Guessan K.A. et al., 2015. Indicateurs de dégradation physique des sols dans le Nord de la Côte d’Ivoire : cas de Boundiali et Ferkessédougou. Afr. Sci. Rev. Int. Sci. Technol., 11(3), 115-128, doi.org/10.4314/afsci.v11i3
Nguyen N.N. et al., 2020. Genome-wide SNP discovery and core marker sets for assessment of genetic variations in cultivated pumpkin (Cucurbita spp.). Hortic. Res., 7(1), 1-10, doi.org/10.1038/s41438-020-00342-9
Ramanatha Rao V. & Hodgkin T., 2002. Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Organ Cult., 68(1), 1-19, doi.org/10.1023/A:1013359015812
Rousseau K., Gautier D. & Wardell D.A., 2015. Coping with the upheavals of globalization in the shea value chain: the maintenance and relevance of upstream shea nut supply chain organization in western Burkina Faso. World Dev., 66, 413-427, doi.org/10.1016/j.worlddev.2014.09.004
Saidou A., 2013. Effet d’un système agroforestier à karité (Vitellaria paradoxa C.F.Gaertn.) sur le sol et le potentiel de production du maïs (Zea maize) en zone soudanienne du Bénin. Int. J. Biol. Chem. Sci., 6(5), doi.org/10.4314/ijbcs.v6i5.16
Sallé G., Boussim J.I., Raynal-Roques A. & Brunck F., 1993. Le karité : état de nos connaissances et perspectives de recherche. In : Riedacker A. et al., eds. Physiologie des arbres et arbustes en zones arides et semi-arides. Montrouge, France : John Libbey Eurotext, 427-439.
Schreckenberg K. et al., 2006. Domesticating indigenous fruit trees as a contribution to poverty reduction. For. Trees Livelihoods, 16(1), 35-51, doi.org/10.1080/14728028.2006.9752544
Soro D., N’Da-Adopo A., Da K.P. & Traore D., 2004. Lutte contre les parasites chez le karité. Agron. Afr., 16(3), 21-28, doi.org/10.4314/aga.v16i3.1647
Soro D., Traore K. & Kassi N.J., 2011. Variabilité des caractères morphologiques chez le karité (Vitellaria paradoxa), dans le Nord de la Côte d’Ivoire. Int. J. Biol. Chem. Sci., 5(3), doi.org/10.4314/ijbcs.v5i3.72263
Wardell D.A. et al., 2021a. Shea (Vitellaria paradoxa C.F.Gaertn.) – a peripheral empire commodity in French West Africa, 1894-1960. Int. For. Rev., 23(4), 511-533, https://www.cifor.org/knowledge/publication/8402
Wardell D.A. et al., 2021b. Shea (Vitellaria paradoxa C.F.Gaertn.) – the emergence of global production networks in Burkina Faso, 1960-2021. Int. For. Rev., 23(4), 534-561, doi.org/10.1505/146554821834777189
Yao S.D.M. et al., 2015. Fiabilité d’une liste minimale de descripteurs agromorphologiques recommandée par le COGENT dans l’étude de la diversité génétique du cocotier (Cocos nucifera L.). J. Anim. Plant Sci., 26, 4006-4022.
Yao S.D.M. et al., 2020. Loranthaceae species infesting shea trees (Vitellaria paradoxa C.F. Gaertn.) and factors involving attacks in Northern Côte d’Ivoire. Ecologia, 10(3), 101-109, doi.org/10.3923/ecologia.2020.101.109
