


- Accueil
- Volume 28 (2024)
- Numéro 2
- Fall armyworm in West Africa: Earwig predation potential and insecticide selectivity


Visualisation(s): 1377 (26 ULiège)
Téléchargement(s): 94 (0 ULiège)

Fall armyworm in West Africa: Earwig predation potential and insecticide selectivity

Document(s) associé(s)
Version PDF originaleRésumé
La chenille légionnaire d’automne en Afrique de l’Ouest : potentiel de prédation des perce-oreilles et sélectivité des insecticides
Description du sujet. La chenille légionnaire d’automne Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera : Noctuidae) est un ravageur majeur des cultures céréalières de base établi en Afrique de l’Ouest depuis 2016. Récemment, des perce-oreilles ont été observés consommant des stades immatures de S. frugiperda dans des champs de maïs.
Objectifs. Dans cette étude, nous avons évalué le potentiel du perce-oreille Diaperasticus erythrocephalus (Olivier) (Dermaptera : Forficulidae) à consommer les œufs et les larves de la chenille légionnaire d’automne au laboratoire. En outre, nous avons comparé la sélectivité de dix bio- et insecticides les plus utilisés sur cette espèce prédatrice ainsi que sur sa proie afin d’identifier les produits compatibles dans une stratégie de lutte intégrée.
Méthode. La prédation a été évaluée en nourrissant des perce-oreilles ad libitum et les tests de susceptibilité en suivant le protocole adapté IRAC 020.
Résultats. Nos résultats suggèrent que les perce-oreilles jouent un rôle important dans la régulation de la chenille légionnaire d’automne : un individu consomme en moyenne 90,3 ± 16,5 œufs ou 36,4 ± 8,7 larves par jour. Nous avons constaté que l’émamectine benzoate et le spinétorame sont efficaces pour lutter contre la chenille légionnaire d’automne sans affecter la survie de ce prédateur. En revanche, le chlorpyrifos-éthyl, le méthomyl et le spinosad sont non seulement efficaces contre le ravageur, mais aussi très toxiques pour le prédateur.
Conclusions. Nous discutons du potentiel de ces résultats dans le développement d'un programme de lutte intégrée contre ce ravageur envahissant. En particulier, nous recommandons de former les agriculteurs à la reconnaissance des ennemis naturels et aux pratiques de leur conservation.
Abstract
Description of the subject. The fall armyworm, Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae) is a major pest of staple cereal crops established in West Africa since 2016. Recently, earwigs have been observed consuming S. frugiperda immature stages in maize fields.
Objectives. In this study, we evaluated the potential of earwigs, Diaperasticus erythrocephalus (Olivier) (Dermaptera: Forficulidae) to consume fall armyworm eggs and larvae in a laboratory setting. In addition, we compared the selectivity of the ten most used bio- and insecticides on this predatory species as well as on its prey for identifying products compatible with an integrated pest management strategy.
Method. Predation was assessed by feeding earwigs ad libitum and susceptibility tests following the adapted IRAC 020 protocol.
Results. Our results suggest that earwigs play an important role in the regulation of the fall armyworm: one individual consumes a mean of 90.3 ± 16.5 eggs or 36.4 ± 8.7 larvae per day. We found emamectin benzoate and spinetoram to be effective for controlling the fall armyworm, while not affecting the survival of this predator. On the other hand, chlorpyrifos-ethyl, methomyl and spinosad are not only effective against this pest but also highly toxic to the predator.
Conclusions. We discuss the potential of these results in the development of an integrated pest management program against this invasive pest. In particular, we recommend training the farmers on natural enemy recognition and their conservation practices.
Table des matières
Received 2 October 2023, accepted 6 May 2024, available online 21 May 2024.
This article is distributed under the terms and conditions of the CC-BY License (http://creativecommons.org/licenses/by/4.0)
1. Introduction
1Over the last decade, the fall armyworm (FAW), Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae) has become one of the most important damaging invasive pests worldwide. It is native to the tropical and subtropical regions of the Americas, then its global distribution has expanded widely across Africa and Asia (Goergen et al., 2016; Maino et al., 2021; Kenis et al., 2023). This pest poses a significant threat to the production of staple cereal crops, consequently nutritional food security, and livelihoods of millions of farmers (Goergen et al., 2016). In maize, FAW is mainly managed by the application of chemical insecticides (Houngbo et al., 2020; Tambo et al., 2020; Palli et al., 2023), despite repeated demonstrations of resistance (Ahissou et al., 2021a; Kenis et al., 2023).
2A diversity of FAW natural enemies have been identified in Senegal (Tendeng et al., 2019), Benin, Côte d’Ivoire, Ghana (Kenis et al., 2019; Agboyi et al., 2020; Koffi et al., 2020; Dassou et al., 2021), Niger (Laminou et al., 2020) and Burkina Faso (Ahissou et al., 2021b). While, natural enemies vary by country, some species are repeatedly reported in many countries, including Coccygidium spp., Chelonus spp. (Hymenoptera: Braconidae), Trichogramma spp. (Hymenoptera: Trichogrammatidae), Telenomus sp. (Hymenoptera: Platygastridae), earwigs (Dermaptera: Forficulidae), and various ant species. In Burkina Faso, the predatory guild is dominated by two Forficulidae species, Diaperasticus erythrocephalus (Olivier) and Forficula senegalensis (Serville) (Ahissou et al., 2021b). However, their potential as FAW biological control agents has never been evaluated in Africa, despite field observations suggesting their importance.
3Studies have shown that certain earwigs are one of the most important natural enemies for controlling pests in maize crops in Brazil (Figueiredo et al., 2006; Cruz, 2007). Management decisions that focus exclusively on reducing insect pests such as chemical insecticides may also eliminate predators, and thus lead to pest control failure (Guedes et al., 2016). Broad-spectrum insecticides like carbamates, organophosphates, and pyrethroids are highly toxic to earwigs (Zotti et al., 2010; Campos et al., 2011). An alternative to keep these predators in the field is to use selective insecticides only. The choice of more selective insecticides is therefore warranted to allow compatibility and complementarity of chemical and biological control methods, which is vital for the successful of integrated pest management programs against FAW.
4This research was conducted during the Ph.D. thesis of Besmer Régis Ahissou (Ahissou, 2022a) and aimed to investigate the predation potential of an earwig species commonly observed in Burkina Faso maize fields, D. erythrocephalus. Besides, the selectivity of the most used chemical and biological insecticides on adult earwigs and their prey to identify products compatible with an integrated pest management strategy against FAW were evaluated. This study will provide authorities and farmers with detailed information to choose the most appropriate insecticides to incorporate into the biological control of FAW in recently invaded areas.
2. Materials and methods
2.1. Insect culture
5Earwig and FAW colonies were collected from different maize fields located in province of Houet (11°20'N, 4°15'W), and shipped to the laboratory at the University Nazi Boni (UNB) in Bobo-Dioulasso, Burkina Faso. Approximately 60 earwigs were reared in transparent plastic boxes (20 × 10 × 10 cm) with openings covered with a thin white cloth for ventilation. They fed on a diet made of maize powder (10 g) and insect remains consisting of dead pupae and adults of the FAW (50 g) from the laboratory stock rearing. The diet was replaced every two days. Wet cotton in a Petri dish was maintained in each plastic box as a water source for the insects. They were acclimated under laboratory conditions for one month before testing.
6FAW larvae were fed daily on fresh maize leaves until pupation. Pupae were daily collected and placed in a cage (60 × 40 × 40 cm); providing with a white paper for female oviposition. FAW adults were fed on a sugar water solution (100 g·l-1). After oviposition, the white paper was removed and cut to individualize each egg mass in separate boxes (Ahissou et al., 2021a). The F1 generation of FAW was used for all bioassays. Both insects were reared under standard laboratory conditions of 25 ± 2 °C, 60 ± 15% relative humidity, and a 12L: 12D photoperiod.
2.2. Insecticides
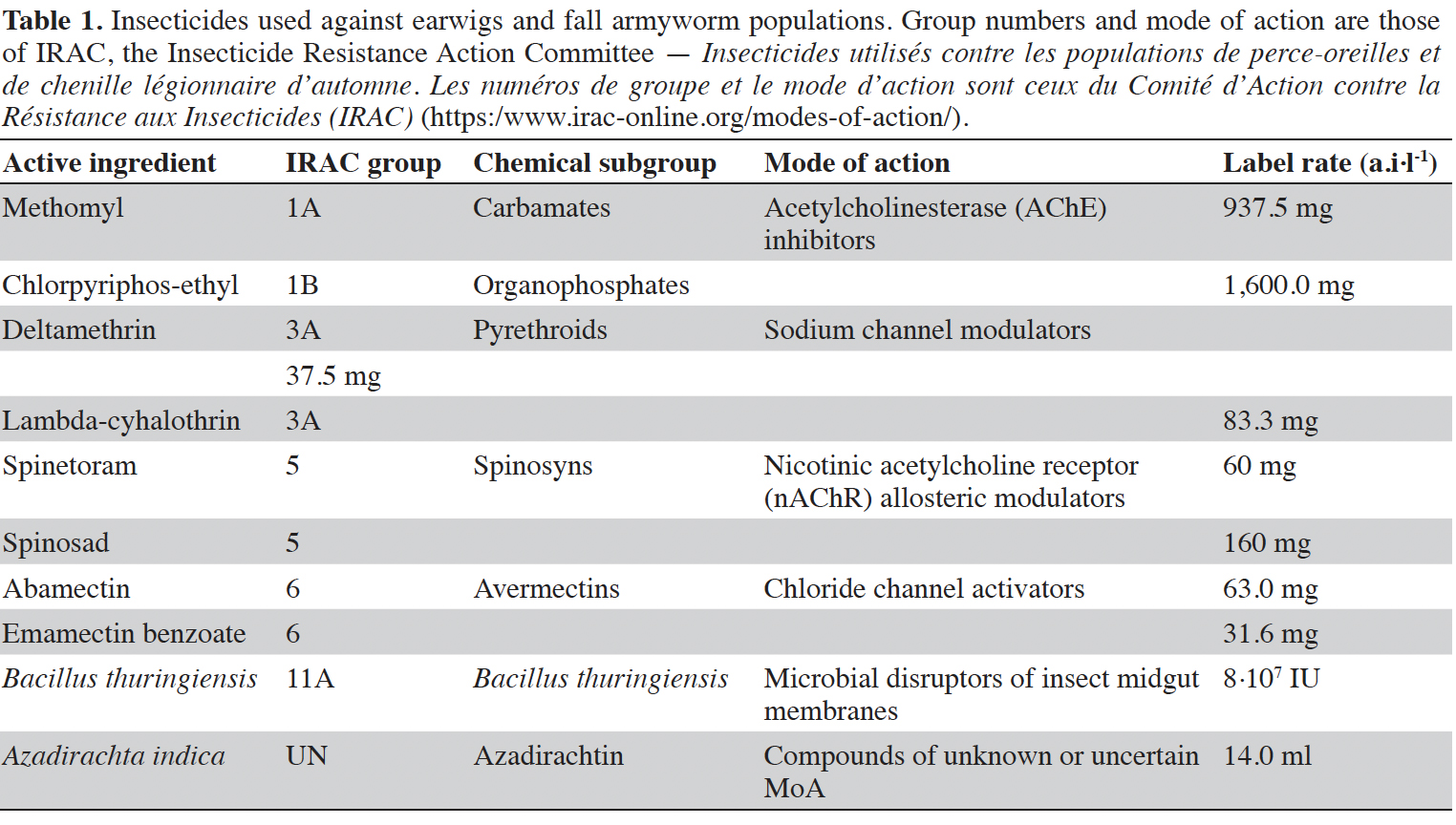
7We evaluated the survival of earwigs after exposure to ten commercial insecticide formulations used in maize fields against FAW (NAR = Normal Application Rate): abamectin 18 g a.i.·l-1 (Acarius® EC, Savana, France, NAR = 1 l·ha-1), chlorpyriphos-ethyl 480 g a.i.·l-1 (Pyrical™ 480 EC, Arysta Lifescience, France, NAR = 1 l·ha-1), deltamethrin 25 g a.i.·l-1 (Tamega® EC, Savana, France, NAR = 0.5 l·ha-1), emamectin benzoate 19 g a.i.·l-1 (Emacot® 019 EC, Savana, France, NAR = 0.5 l·ha-1), lambda-cyhalothrin 25 g a.i.·l-1 (Sunhalothrin® 2.5% EC, Wynca Sunshine, Mali, NAR = 0.5 l·ha-1), methomyl 250 g a.i.·kg-1 (Savahaler® WP, Savana, France, NAR = 1.5 kg·ha-1). Some biopesticides were also included in the assay as spinetoram 120 g a.i.·l-1 (Radiant™ 120 SC, Dow AgroSciences, NAR = 100 ml·ha-1), pinosad 480 g a.i.·l-1 (Laser™ 480 SC, Dow AgroSciences, NAR = 100 ml·ha-1), Bacillus thuringiensis 16,000 IU·mg-1 (Bio K® 16 WP, Savana, France, NAR = 1.5 kg·ha-1), and Azadirachta indica extracts (HN, Bioprotect, Burkina Faso, NAR = 5 l·ha-1). The ten active ingredients, their IRAC group and modes of action are listed in table 1.
2.3. Predation on eggs and larvae of fall armyworm
8FAW eggs (≤ 24 h old) were placed with a brush in each Petri dish (9 cm diameter). Preliminary tests showed that egg viability was not affected by this manipulation. Predation rate was assessed individually by placing D. erythrocephalus male or female (previously starved for 24 h) in Petri dish containing prey and sealed with parafilm. Egg predation rate was determined using 130-150 eggs. To evaluate the predation capacity on the FAW larval instars, the same protocol was used by introducing 60 L1, 30 L2 and 10 L3 larvae in each dish containing fresh maize leaves where a male or female earwig was subsequently added. The controls consisted of dishes containing fresh maize leaves and either eggs or larvae only. Five replicates of (10) earwigs were used for each sex and prey stage. The boxes were maintained under the same aforementioned conditions. After 24 h of earwig exposure, each dish was carefully examined with a binocular microscope to determine prey consumption (eggs and larvae). The eggs were totally consumed and sometimes only a thin transparent film remained. Consumed larvae were easily identified because only the cephalic capsule usually remained.
2.4. Insecticide susceptibility of earwigs and fall armyworms
9Bioassays were conducted according to the IRAC standard leaf bioassay protocol (http://www.irac-online.org/methods/) using adult earwig, D. erythrocephalus and FAW third instar F1. A minimum of five concentrations was used for each insecticide diluted with distilled water containing Triton (0.2 g·l-1) for FAW. Regarding the survival of earwigs, only the recommended dose of different insecticides was tested. Non-treated maize leaves were collected, washed with tap water and dried. Then, they were immersed for 10 seconds in the insecticide solution and dried for 1 h. Control leaves were treated only with a solution of Triton in water. Leaves were placed in Petri dishes (9 cm in diameter) containing blotting paper. A total of 40 insects were exposed individually to each concentration, four replicates were performed for earwigs and five to six for FAW stages (Ahissou et al., 2021a). All bioassays were conducted under laboratory conditions of 25 ± 2 °C, 60 ± 15% relative humidity, and a photoperiod 12L: 12D. Earwig survival was assessed for 24, 48, and 72 h, and the diet (20 mg) was added daily to each Petri dish. FAW larval mortality was assessed after 72 h for all tested insecticides. Insects were considered dead if they failed to move when touched with a small brush.
2.5. Data analysis
10Data recorded on the number of FAW eggs and larvae consumed by adult earwigs, as well as earwig survival to insecticides, were subjected to a generalized linear models (GLMs) procedure, binomial distribution respectively. Multiple comparisons for these data were performed with Tukey’s post hoc test using the 'ghlt' function in the 'multcomp' package for R. The concentration-mortality of the FAW population were corrected for control mortality (Abbott, 1925), and were subjected to probit analysis (Finney, 1971), to calculate values of slope, lethal concentration (LC50 and LC80), and fiducial limits (95%). Insects responses were considered equal when the confidence limits overlapped (Robertson & Preisler, 1992). Control failure likelihood (CFL) was calculated using Guedes (2017) formula by multiplying the percentage of mortality achieved by 100, dividing the product by the minimum efficiency required and subtracting the result from 100. CFL is the probability of treatment failure and the achieved mortality is the percentage of individuals that die when the recommended dose is applied. The required efficacy was set at 80%, as the minimum efficacy threshold required to allow registration of insecticides (Guedes, 2017). If the achieved mortality was higher than the required efficacy of the commercial formulation, CFL values below or near 0% suggest a negligible risk of control failure. Except for the probit analysis performed in SPSS 25.0 statistical package for Windows, all other analysis were performed in R statistical software (R Core Team, 2021) using the RStudio-2021.09.2 interface.
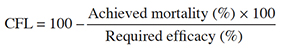
3. Results
3.1. Predation on eggs and larvae of fall armyworm
11The consumption capacity of earwigs varied significantly with the developmental stage of the tested prey (FAW eggs or larvae), and the sex of the predator. Daily egg consumption by female earwigs (98.2 ± 17.6) was significantly higher than that of males (82.4 ± 10.3) (Df = 1; X2 = 197.68; p < 0.001) (Figure 1). Moreover, adult females also consumed significantly more larvae (L1) (38.6 ± 8.2) than males (34.3 ± 8.7) (Df = 5; X2 = 596.9; p = 0.03). In contrast, non-significant difference was observed in consumption between female and male earwigs when fed on FAW L2 (p = 0.8) and L3 (p = 1.0) larvae.
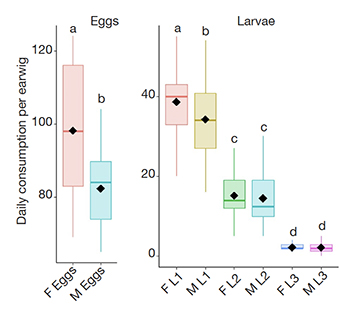
Figure 1. Predation of earwig females (F) and males (M) on fall armyworm eggs and larval stages (L1, L2 and L3) — Capacité de prédation des femelles (F) et des mâles (M) de perce-oreilles sur les œufs et les stades larvaires (L1, L2 et L3) de la chenille légionnaire d’automne.
Different letters indicate significant differences — les lettres différentes indiquent des différences significatives (GLM; Tukey test, p < 0.05); Figure shows the means (black diamonds), SE (boxes) and SD (whiskers) — la figure montre les moyennes (losanges noirs), les erreurs standards (boîtes) et écarts-types (moustaches).
3.2. Survival rate of earwigs to insecticides and control failure likelihood (CFL)
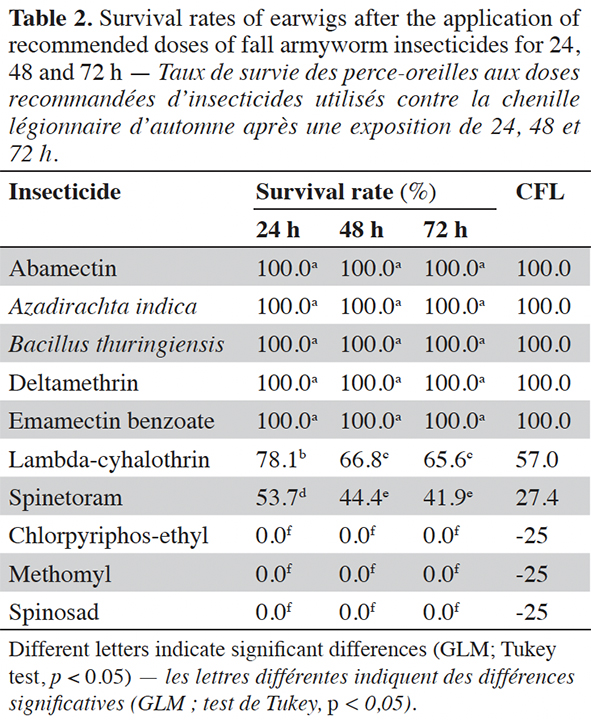
12No mortality was recorded on the control treatments. Survival of earwigs after 24, 48 and 72 h of exposure to insecticides varied significantly according to the type of insecticides (Table 2):
13– chlorpyriphos-ethyl, methomyl, and spinosad insecticides were very toxic for earwigs that all died within the first 24 h of exposure (Table 2). With a zero CFL (-25), these products would eliminate earwigs from maize fields at the recommended dose;
14– lambda-cyhalothrin and spinetoram insecticides were moderately toxic to earwigs at all tested exposure times with a survival rate between 41.9-65.6%. The CFL values of 57.0 and 27.4 respectively; suggesting that at the manufacturer’s recommended rates, these products could provide moderate suppression of earwigs in maize fields;
15– the insecticides abamectin, emamectin benzoate, deltamethrin, A. indica and B. thuringiensis did not affect earwig survival (100%) at all tested exposure times. With an CFL of 100, all earwigs could probably survive in maize fields after the applications of these products at the manufacturer’s recommended rates.
3.3. Susceptibility of fall armyworm to insecticides
16The natural mortality observed in the control treatments was below 5% and was used to correct the insecticide mortality. For all tested insecticides, the Probit model was considered appropriate (Table 3).
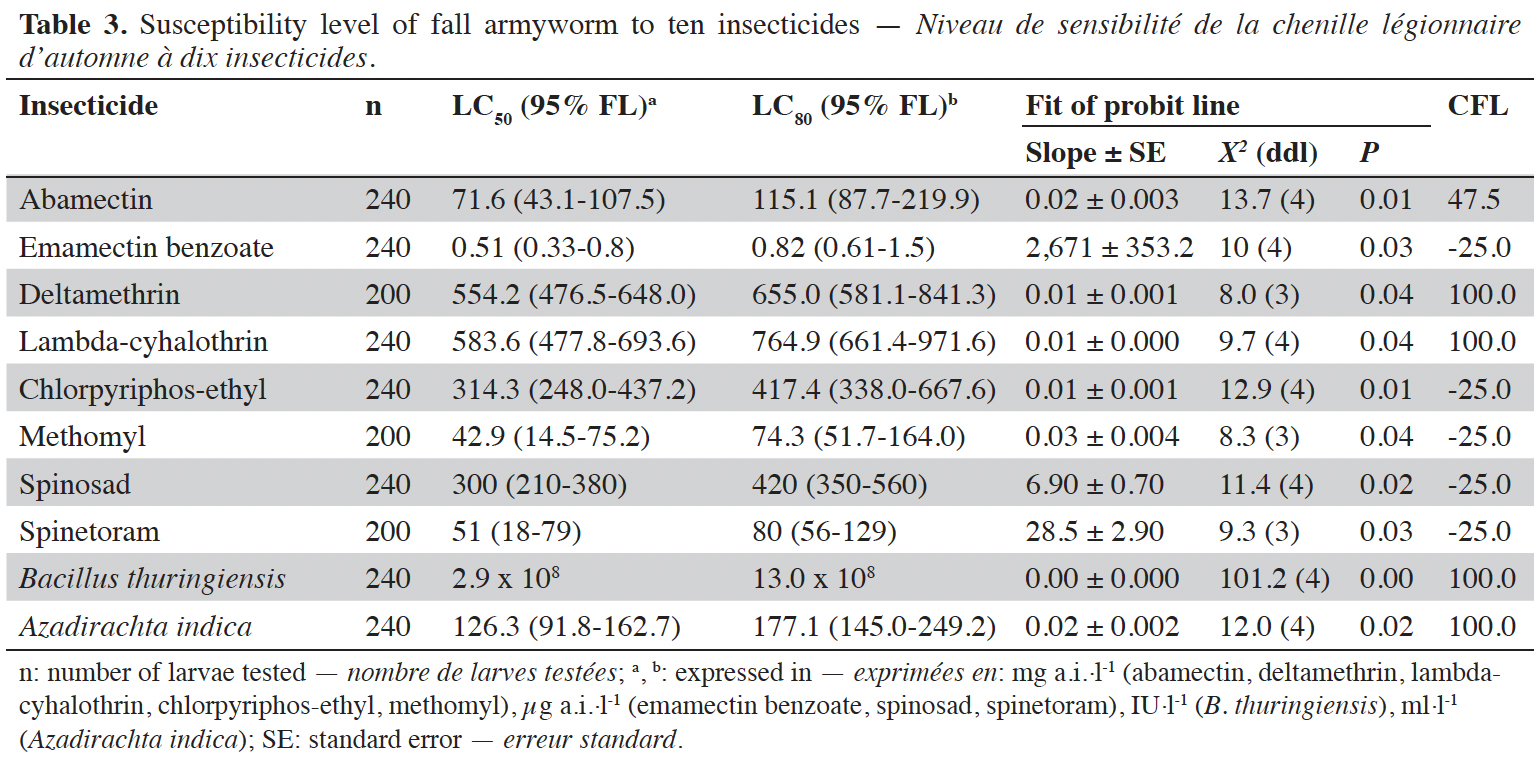
17Spinosyns. Spinetoram and spinosad were highly toxic insecticides to FAW with LC80 values of 80.0 µg·l-1 and 420.0 µg·l-1, respectively (Table 3). These values are 99% lower than those recommended by the manufacturer. The high slope values (6.9 to 28.5) mean that a small increase in insecticide concentration is sufficient to significantly increase larval mortality; suggesting that the FAW population is very sensitive to both molecules. At the dose recommended, very limited failure treatment should be observed.
18Organophosphates and carbamates. The LC80 values for chlorpyrifos-ethyl (417.4 mg·l-1) and methomyl (74.3 mg·l-1) were lower than the recommended doses by 74% and 92%, respectively. Their confidence limits do not overlap, methomyl has a higher toxicity than chlorpyrifos-ethyl. With a CFL of -25 for both molecules, they would be 100% effective against the FAW at the recommended rates.
19Avermectins. The LC80 values of emamectin benzoate and abamectin were 0.82 µg·l-1 and 115.1 mg·l-1, respectively. Since their confidence limits do not overlap, abamectin has a lower toxicity than emamectin benzoate. This value for emamectin benzoate is 99% lower than that recommended by the manufacturer. At the doses recommended by the manufacturers, the CFL is 47.5 for abamectin and -25 for emamectin benzoate.
20Pyrethroids. Deltamethrin and lambda-cyhalothrin were less toxic to FAW with similar LC80 values (confidence limits overlap) of 655.0 mg·l-1 and 764.9 mg·l-1, respectively. These values were 9 to 17 higher than the manufacturer’s recommended rates, at which the CFL is very high (100).
21The LC80 values of A. indica (177.1 ml·l-1) and B. thuringiensis (13.0 × 108 IU·l-1) were 12 to 16 times higher than the manufacturer’s recommended rates, respectively. Their slope is low (< 0.001); suggesting that a large increase in concentration is required to obtain a small increase in mortality. The probabilities that A. indica and B. thuringiensis would not be effective to eradicate the FAW from fields are high (100%).
4. Discussion
22Our study was conducted in the context of associations of several natural enemies with the FAW in West Africa after its invasion (Agboyi et al., 2020; Koffi et al., 2020; Ahissou et al., 2021b). This study assessed the predation potential of earwigs on FAW eggs and larvae in the laboratory. Also, the insecticide's effectiveness against the FAW as well as less toxic to the earwigs that were abundant in untreated maize fields in Burkina Faso were studied (Ahissou et al., 2021b). Most of them are generalist predators that are considered to be major components of biological control agents for insect pests (Cruz, 2007; Prasanna et al., 2018). Their conservation and promotion in the field is an important strategy to promote biological control of arthropod pests.
23Earwig, D. erythrocephalus consumed an average of 90 eggs or 36 larvae of the FAW per day in the present study. Similar results were recorded in Brazil and Argentina for Euborellia annulipes (Lucas) on FAW eggs (da Silva et al., 2009a, 2009b), and Doru luteipes (Scudder) on larvae (Reis et al., 1988; Romero Sueldo et al., 2010). They are recommended for augmentative releases for biological control of FAW in Brazil (Cruz, 2007). These results show that earwigs could play a crucial role in the regulation of the FAW in maize fields in Africa, particularly by consuming their egg masses and early larval stages that are gregarious. We recommend additional studies to test the effectiveness of certain agricultural practices and plants in attracting earwigs in the field for biological control. For example, spraying a sugar solution on maize led to the increase of some species of earwigs in Brazil (Bortolotto et al., 2014). In East Africa, the abundance of some predators is increased in maize fields cultivated in push-pull with intercrops Melinis minutiflora P.Beauv. (Poaceae), Desmodium uncinatum (Jacq.) DC., Desmodium intortum (Mill.) Urb. (both Fabaceae) and trap crops Pennisetum purpureum Schumach., and Sorghum vulgare Pers. (both Poaceae) (Cook et al., 2007; Kebede et al., 2018).
24Insecticides with three modes of action: chloride channel activators (emamectin benzoate), acetylcholinesterase (AChE) inhibitors (chlorpyrifos-ethyl, methomyl), and nicotinic acetylcholine receptor (nAChR) allosteric modulators (spinetoram, spinosad) were found effective on FAW; confirming previous works (Campos et al., 2011; Ríos-Díez & Saldamando-Benjumea, 2011; Gutiérrez-Moreno et al., 2019; Dassou et al., 2021) including those performed in Burkina Faso (Ahissou et al., 2021a; Ahissou et al., 2022b). Based on our results, control failure for FAW would be very low, as the doses required are lower than those authorized by the manufacturers, representing a safe environmental profile. In contrast, insecticides lambda-cyhalothrin, deltamethrin, and abamectin were ineffective against FAW populations collected in Burkina Faso, with a high risk of failure in the field. The ineffectiveness of some pyrethroid insecticides was documented worldwide, e.g. in Puerto Rico and Brazil (Carvalho et al., 2013; Gutiérrez-Moreno et al., 2019). The widespread treatment failures mentioned by producers in West Africa could be related to the use of ineffective molecules such as abamectin, deltamethrin, and lambda-cyhalothrin (Ahissou et al., 2021a), which are widely used against this pest in West Africa (Kansiime et al., 2019; Ahissou et al., 2022c), or to pest resistance to these molecules.
25Genetic studies on the Spodoptera genus have shown that emamectin benzoate inheritance was autosomal, incompletely dominant, and polygenic in S. frugiperda (Muraro et al., 2021) and S. exigua Hübner (Che et al., 2015). Consequently, intensive and continuous application of this active ingredient has contributed to the rapid evolution of high resistance in field populations of S. frugiperda in Brazil (Muraro et al., 2022) and S. exigua in China (Che et al., 2015). However, a low (or absence) cross-resistance between emamectin benzoate and methomyl, spinetoram and chlorpyrifos has been demonstrated in FAW (Muraro et al., 2021). Given that high levels of FAW resistance to effective molecules identified in the present study have already been reported in the Americas (Gutiérrez-Moreno et al., 2019; Muraro et al., 2022), it would be judicious to develop alternative means of control and resistance management systems (e.g., rotation of molecules with different modes of action) to delay this development (Sparks et al., 2021).
26Obtained results categorized three groups of insecticides based on the toxicity values on earwigs and FAW:
27– Emamectin benzoate and spinetoram are effective on FAW and have no (or very little) effect on the earwigs. Both insecticides were found to be selective and may be used in FAW control program in combination with earwigs or without affecting those available in the field. As in the present study, insecticides that target the chloride channel activators (emamectin benzoate, abamectin) and nAChR allosteric modulators (spinetoram) had a safety profile on earwigs, Forficula auricularia L. (Shaw & Wallis, 2010) and E. annulipes (Potin et al., 2022), respectively;
28– Chlorpyrifos-ethyl, methomyl and spinosad are effective on FAW and their use in maize fields may eliminate the pest and the earwigs present. These results confirm previous studies that reported organophosphates and carbamates are generally broad-spectrum compounds with low selectivity for earwigs (Zotti et al., 2010; Campos et al., 2011) as in the case of spinosad, which was harmful to F. auricularia earwigs (Shaw & Wallis, 2010; Fountain & Harris, 2015). Since earwigs are nocturnal predators more likely to be exposed to insecticide residues on plants when feeding at night (rather than by direct contact), we recommend field studies to confirm these results, especially for highly toxic molecules;
29– Deltamethrin, lambda-cyhalothrin, abamectin, A. indica and B. thuringiensis are not effective against either FAW or the predator. Generally, pyrethroid insecticides are considered highly toxic to natural enemies (Talebi et al., 2011), but they were not toxic to earwigs and did not cause mortality, as bioinsecticides in our study.
5. Conclusions
30Earwigs could be promising biological control agents against FAW if they are present early before egg laying and/or hatching; alternatively, we can preserve their regulatory potential by using selective insecticides (like emamectin benzoate and spinetoram). These insecticides are effective against FAW larvae while posing minimal harm to earwigs and other beneficial insects, allowing them as natural predators and keeping the pest populations under the economic threshold. This approach makes chemical control compatible with preserving natural biological control, promoting a more sustainable and effective pest management strategy.
Acknowledgements
31This research was supported by the Academy of Research and Higher Education-Commission Development Cooperation (ARES-CDD) as part of the PRD AGRO-ECO project. The authors would like to thank the project coordinators, Prof. Marie-Paule Kestemont (UCLouvain, Belgium) and Prof. Enoch G. Achigan-Dako (University of Abomey-Calavi, Benin). We also wish to express our thanks and gratitude to the farmers who made this study a success by allowing us access to their fields.
Bibliographie
Abbott W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18, 265-267, doi.org/10.1093/jee/18.2.265a
Agboyi L.K. et al., 2020. Parasitoid complex of fall armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects, 11(68), 1-15, doi.org/10.3390/insects11020068
Ahissou B.R. et al., 2021a. Baseline toxicity data of different insecticides against the fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) and control failure likelihood estimation in Burkina Faso. Afr. Entomol., 29(2), 435-444, doi.org/10.4001/003.029.0435
Ahissou B.R. et al., 2021b. Natural enemies of the fall armyworm Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) in Burkina Faso. Tropicultura, 39(3), 1-21, doi.org/10.25518/2295-8010.1881
Ahissou B.R., 2022a. La chenille légionnaire d’automne en Afrique de l’Ouest : états des lieux, recherche et propositions de stratégies de gestion. Thèse de doctorat : Université de Liège - Gembloux Agro-Bio Tech, Gembloux (Belgique).
Ahissou B.R. et al., 2022b. Susceptibility of fall armyworm Spodoptera frugiperda (JE Smith) to microbial and botanical bioinsecticides and control failure likelihood estimation. Biotechnol. Agron. Soc. Environ., 26(3), 136-140, doi.org/10.25518/1780-4507.19793
Ahissou B.R. et al., 2022c. Farmers’ knowledge and management practices of the fall armyworm (Spodoptera frugiperda Smith) in Burkina Faso. Biotechnol. Agron. Soc. Environ., 26(4), 191-197, doi.org/10.25518/1780-4507.19909
Bortolotto O.C. et al., 2014. Sugar solution treatment to attract natural enemies and its impact on fall armyworm Spodoptera frugiperda in maize fields. Interciencia, 39(6), 416-421.
Campos M.R. et al., 2011. Insecticide selectivity and behavioral response of the earwig Doru luteipes. Crop Prot., 30, 1535-1540, doi.org/10.1016/j.cropro.2011.08.013
Carvalho R.A. et al., 2013. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda. PLoS One, 8(4), e62268, doi.org/10.1371/journal.pone.0062268
Che W. et al., 2015. Cross-resistance and inheritance of resistance to emamectin benzoate in Spodoptera exigua (Lepidoptera: Noctuidae). J. Econ. Entomol., 108(4), 2015-2020, doi.org/10.1093/jee/tov168
Cook S.M., Khan Z.R. & Pickett J.A., 2007. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol., 52, 375-400, doi.org/10.1146/annurev.ento.52.110405.091407
Cruz I., 2007. Controle biológico de Pragas na cultura de milho para produção de conservas (Minimilho), por meio de parasitóides e predadores. Circ. Técnico, 91. Sete Lagoas, Brazil : Embrapa Milho e Sorgo, 1-16.
da Silva A.B., Batista J. de L. & de Brito C.H., 2009a. Capacidade predatória de Euborellia annulipes (Lucas, 1847) sobre Spodoptera frugiperda (Smith, 1797). Acta Sci. - Agron., 31(1), 7-11, doi.org/10.4025/actasciagron.v31i1.6602
da Silva A.B., de Luna Batista J. & de Brito C.H., 2009b. Aspectos biológicos de Euborellia annulipes sobre ovos de Spodoptera frugiperda. Eng. Ambient. – Espírito St. do Pinhal., 6(3), 482-495.
Dassou A.G. et al., 2021. Fall armyworm, Spodoptera frugiperda (J.E. Smith) in maize cropping systems in Benin: abundance, damage, predatory ants and potential control. Int. J. Trop. Insect Sci., 41(4), 2627-2636, doi.org/10.1007/s42690-021-00443-5
Figueiredo M. de L.C., Martins-Dias A.M.P. & Cruz I., 2006. Relationship between fall armyworm and their natural biological control agents in the maize crop. Pesqui. Agropecuária Bras., 41(12), 1693-1698, doi.org/10.1590/S0100-204X2006001200002
Finney D.J., 1971. Probit analysis. 3rd ed. Cambridge, UK: Cambridge University Press.
Fountain M.T. & Harris A.L., 2015. Non-target consequences of insecticides used in apple and pear orchards on Forficula auricularia L. (Dermaptera: Forficulidae). Biol. Control, 91, 27-33, doi.org/10.1016/j.biocontrol.2015.07.007
Goergen G. et al., 2016. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS One, 11(10), e0165632, doi.org/10.1371/journal.pone.0165632
Guedes R.N.C., Smagghe G., Stark J.D. & Desneux N., 2016. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. Annu. Rev. Entomol., 61, 43-62, doi.org/10.1146/annurev-ento-010715-023646
Guedes R.N.C., 2017. Insecticide resistance, control failure likelihood and the first law of geography. Pest Manage. Sci., 73, 479-484, doi.org/10.1002/ps.4452
Gutiérrez-Moreno R. et al., 2019. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol., 112(2), 792-802, doi.org/10.1093/jee/toy372
Houngbo S. et al., 2020. Farmers’ knowledge and management practices of fall armyworm, Spodoptera frugiperda (J.E. Smith) in Benin, West Africa. Agriculture, 10(430), 1-15, doi.org/10.3390/agriculture10100430
Kansiime M.K. et al., 2019. Farmer perception of fall armyworm (Spodoptera frugiperda J.E. Smith) and farm-level management practices in Zambia. Pest Manage. Sci., 75, 2840-2850, doi.org/10.1002/ps.5504
Kebede Y., Baudron F., Bianchi F. & Tittonell P., 2018. Unpacking the push-pull system: assessing the contribution of companion crops along a gradient of landscape complexity. Agric. Ecosyst. Environ., 268, 115-123, doi.org/10.1016/j.agee.2018.09.012
Kenis M. et al., 2019. Telenomus remus, a candidate parasitoid for the biological control of Spodoptera frugiperda in Africa, is already present on the continent. Insects, 10(92), 1-10, doi.org/10.3390/insects10040092
Kenis M. et al., 2023. Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomol. Gen., 43(2), 187-241, doi.org/10.1127/entomologia/2022/1659
Koffi D. et al., 2020. Natural enemies of the fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in Ghana. Florida Entomol., 103(1), 85-90, doi.org/10.1653/024.103.0414
Laminou S. et al., 2020. Parasitism of locally recruited egg parasitoids of the fall armyworm in Africa. Insects, 11(430), 1-13, doi.org/10.3390/insects11070430
Maino J.L. et al., 2021. Regional and seasonal activity predictions for fall armyworm in Australia. Curr. Res. Insect Sci., 1, 100010, doi.org/10.1016/j.cris.2021.100010
Muraro D.S. et al., 2021. Inheritance patterns, cross-resistance and synergism in Spodoptera frugiperda (Lepidoptera: Noctuidae) resistant to emamectin benzoate. Pest Manage. Sci., 77, 5049-5057, doi.org/10.1002/ps.6545
Muraro D.S. et al., 2022. Evidence of field-evolved resistance in Spodoptera frugiperda (Lepidoptera: Noctuidae) to emamectin benzoate in Brazil. Crop Prot., 162, 106071, doi.org/10.1016/j.cropro.2022.106071
Palli S.R. et al., 2023. The fall armyworm: recent advances in biology and management. J. Pest Sci., 96(4), 1341-1343, doi.org/10.1007/s10340-023-01688-4
Potin D.M., Machado A.V.A., Barbosa P.R.R. & Torres J.B., 2022. Multiple factors mediate insecticide toxicity to a key predator for cotton insect pest management. Ecotoxicology, 31, 490-502, doi.org/10.1007/s10646-022-02526-6
Prasanna B.M., Huesing J.E., Eddy R. & Peschke V.M., eds., 2018. Fall armyworm in Africa: a guide for integrated pest management. 1st ed. Mexico: CIMMYT.
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing.
Reis L.L., Oliveira L.J. & Cruz I., 1988. Biologia e potencial de Doru luteipes no controle de Spodoptera frugiperda. Pesqui. Agropecuária Bras., 23(4), 333-342.
Ríos-Díez J.D. & Saldamando-Benjumea C.I., 2011. Susceptibility of Spodoptera frugiperda (Lepidoptera: Noctuidae) strains from Central Colombia to two insecticides, methomyl and lambda-cyhalothrin: a study of the genetic basis of resistance. J. Econ. Entomol., 104(5), 1698-1705, doi.org/10.1603/ec11079
Robertson J.L. & Preisler H.K., 1992. Pesticide bioassays with arthropods. Boca Raton, FL, USA: CRC.
Romero Sueldo G.M., Bruzzone O.A. & Virla E.G., 2010. Characterization of the earwig, Doru lineare, as a predator of larvae of the fall armyworm, Spodoptera frugiperda: a functional response study. J. Insect Sci., 10(38), 1-10, doi.org/10.1673/031.010.3801
Shaw P.W. & Wallis D.R., 2010. Susceptibility of the European earwig, Forficula auricularia, to insecticide residues on apple leaves. New Zeal. Plant Prot., 59, 55-59, doi.org/10.30843/nzpp.2010.63.6568
Sparks T.C. et al., 2021. Insecticide resistance management and industry: the origins and evolution of the Insecticide Resistance Action Committee (IRAC) and the mode of action classification scheme. Pest Manage. Sci., 77(6), 2609-2619, doi.org/10.1002/ps.6254
Talebi K., Hosseininaveh V. & Ghadamyari M., 2011. Ecological impacts of pesticides in agricultural ecosystem. In: Stoytcheva M., ed. Pesticides in the modern world-risks and benefits. Rijeka, Croatia: In Tech Open Access Publisher, 143-169.
Tambo J.A. et al., 2020. Understanding smallholders’ responses to fall armyworm (Spodoptera frugiperda) invasion: evidence from five African countries. Sci. Total Environ., 740, 140015, doi.org/10.1016/j.scitotenv.2020.140015
Tendeng E. et al., 2019. The fall armyworm Spodoptera frugiperda (J. E. Smith), a new pest of maize in Africa: biology and first native natural enemies detected. Int. J. Biol. Chem. Sci., 13(2), 1011-1026, doi.org/10.4314/ijbcs.v13i2.35
Zotti M.J. et al., 2010. Seletividade de inseticidas usados na cultura do milho para ovos e ninfas do predador Doru lineare (Eschscholtz, 1822) (Dermaptera: Forficulidae). Arq. Inst. Biol. (Sao Paulo), 77(1), 111-118, doi.org/10.1590/1808-1657v77p1112010
