


- Home
- volume 12 (2008)
- Effets de la vitesse et de la durée du remplissage du grain ainsi que de l'accumulation des assimilats de la tige dans l'élaboration du rendement du blé dur (Triticum durum Desf.) dans les conditions de culture des hautes plaines orientales d'Algérie


View(s): 6145 (34 ULiège)
Download(s): 436 (4 ULiège)

Effets de la vitesse et de la durée du remplissage du grain ainsi que de l'accumulation des assimilats de la tige dans l'élaboration du rendement du blé dur (Triticum durum Desf.) dans les conditions de culture des hautes plaines orientales d'Algérie

Editor's Notes
Reçu le 22 juin 2006, accepté le 3 octobre 2007
Résumé
La présente étude a été conduite sur le site expérimental de la station ITGC de Sétif. L'objectif est de déterminer les différences de durée et de vitesse de remplissage et la contribution des assimilats des tiges au rendement de 5 génotypes de blé dur (Triticum durum Desf.). L'effet année est significatif, ce qui explique la fluctuation des conditions de croissance d'une année à une autre. Le rendement en grains est associé à une grande quantité d'assimilats stockés et transférés des tiges pour le remplissage des grains. Le génotype Mbb qui possède une hauteur importante de la tige transfère plus d'assimilats. Les génotypes ADS497 et Deraa présentent des feuilles étendard de grande dimension. Le début de la phase active du remplissage du grain correspond au début de la sénescence foliaire. Mbb présente un rythme de dessèchement foliaire plus lent, la vitesse de dessèchement stationnaire enregistré par ADS497 est de -0,5957 cm2 par jour. La vitesse de remplissage des grains est négativement liée à la durée de remplissage. La participation des assimilats venant de la tige diminue lorsque le milieu permet l'expression d'un meilleur rendement en grains.
Abstract
Effects of speed and the duration of grain filling and the accumulation of the assimilates of the stem in developing the durum wheat yield (Triticum durum Desf.) in the culture conditions of the high plains of eastern Algeria. The present study was led on the experimental site of station ITGC in Setif. The objective is to determine the differences of duration and speed of filling and the contribution of the assimilates of the stems to the yield of 5 durum wheat genotypes (Triticum durum Desf.). The year effect is significant, what explains the fluctuation of the conditions of one growth year to another. The grain yield is associated to the great quantity of assimilates stored and transferred from the stems to the grain filling. The Mbb genotype which has an important stem height transfers more assimilates. The genotypes ADS497 and Deraa present large standard leaves. The beginning of the active phase of the grain filling corresponds to the beginning of the foliar senescence. Mbb presents a slower foliar drying rate, the speed of drying stationary recorded by ADS497 is of -0.5957 cm2 per day. The speed of grain filling is negatively related to the duration of filling. The participation of the assimilates coming from the stem decreases when the environment allows the expression of a better grain yield.
Table of content
1. Introduction
1La culture du blé dans les régions semi-arides est soumise à différentes contraintes abiotiques qui minimisent de façon très significative le rendement en grains. Le transfert des sucres solubles stockés dans le pédoncule de l'épi est un facteur très souvent cité par plusieurs auteurs et qui reste à vérifier dans la présente étude.
2L'accumulation des sucres pendant la pré-anthése contribue à raison de 13 à 70 % au rendement en grains. Sous stress hydrique, le pédoncule de l'épi représente un site de stockage des réserves d'assimilats en vue d'un meilleur remplissage des grains, ces sucres accumulés proviennent de l'hydrolyse des réserves d'amidon des racines.
3La part des assimilats transférés est relativement marginale en bonnes années pour devenir important lors des années défavorables, lorsque le stress hydrique affecte fortement l'appareil photosynthétique.
4Le rendement en grains, sous système de culture pluviale et sous environnement contraignant, est la résultante de la durée, de la vitesse de remplissage et de la capacité de translocation des assimilats stockés dans la tige. Sous de telles conditions de production, la durée de remplissage et par conséquent le poids du grain atteignent rarement leurs valeurs maximales, d'où une baisse du rendement en grains (Abbassenne et al., 1998).
2. Matériel et méthodes
2.1. Mise en place de l'expérimentation
5L'expérimentation a été conduite sur le site expérimental de la station ITGC de Sétif (Algérie), sur cinq variétés de blé dur. Le matériel végétal est semé au cours de la mi-novembre sur trois campagnes agricoles consécutives 2000-2001 à 2002-2003 sur des parcelles élémentaires de 6 rangs de 5 m de long avec un espace inter-rangs de 20 cm, soit une superficie parcellaire de 6 m². Le dispositif expérimental est constitué de blocs complètement randomisés avec quatre répétitions et le précédent cultural est une jachère intégrale.
6Les variétés concernées par l'étude sont Mohammed Ben Bachir (Mbb), Waha, Derraa, Heider et 439/ADS-97 (Tableau 1). Le site expérimental est pratiquement soumis au climat des hautes plaines orientales algériennes, qui se caractérise par des hivers froids, un régime pluviométrique irrégulier, des gelées printanières très fréquentes et des vents chauds et desséchants en fin de cycle de la céréale (Baldy et al., 1993). L'altitude moyenne avoisine 1000 m.

2.2. Conditions climatiques
7La quantité moyenne de pluie enregistrée par campagne agricole est variable ; la valeur la plus élevée était de 521,8 mm durant la campagne 2002-2003 ; la valeur la plus basse, de 215,9 mm au cours de la campagne 2001-2002. Durant les campagnes expérimentales, la quantité de pluie était moyenne la première année, faible la deuxième année et importante la troisième, importante en comparaison de la moyenne de 1981 à 2003 (Tableau 2).
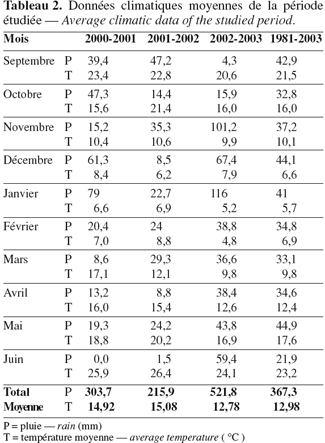
8Une comparaison inter-mensuelle des deux années extrêmes (2001-2002 et 2002-2003) nous révèle des fluctuations de quantité de pluie variables selon les mois, mais en moyenne cette quantité est supérieure lors de la campagne 2002-2003 (Figure 1).
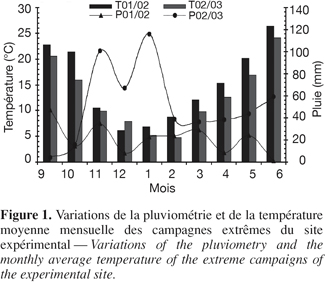
9Concernant la température moyenne, on enregistre une diminution qui débute à partir du mois de septembre (environ 20 °C) jusqu'au mois de février (environ 6 °C), puis la température remonte vers le mois de juin (environ 25 °C), cette augmentation coïncide avec la phase de remplissage des grains (Tableau 2).
10La courbe qui dessine les températures mensuelles moyennes des deux années extrêmes suit une même allure pour les deux années, avec une légère différence en faveur de la campagne 2001-2002 (Figure 1).
2.3. Suivi et notations
11Les observations ont porté sur la date du stade épiaison, la durée de la phase végétative (PVG), la matière sèche obtenue après passage à l'étuve à 85 °C pendant 24 heures. La différence entre le poids de la matière sèche totale de l'échantillon et celle des graines est prise comme étant le poids de la matière sèche accumulée dans les tiges. La vitesse de croissance végétative (VCV), exprimée en g.j-1.m-², est déterminée par le rapport respectif RDT/PRG. A maturité, on détermine le rendement en grains, la biomasse totale produite, l'indice de récolte et la hauteur du chaume.
12La vitesse de remplissage par grain (V, mg.j-1) est estimée par régression linéaire. La durée de remplissage (D, j) est déterminée par le rapport du poids du grain atteint à maturité sur la vitesse de remplissage (V). La vitesse de remplissage est ramenée au nombre de grains par m2 (VRG, g.j-1.m-2). La matière sèche accumulée dans les tiges a été déterminée à l'épiaison, à son maximum et à maturité. La quantité de matière sèche des tiges transférée vers le grain est estimée par la différence entre le poids maximal de la matière sèche accumulée dans les tiges (g.m-2) moins le poids de la matière sèche des tiges mesuré à maturité (g.m-2).
13Elle est ensuite relativisée par rapport au rendement en grains par m2 et au poids moyen d'un grain :
14T (%RDT) = 100 (T/RDT)
15T (%P1G) = 100 [(1000T/NGM2) / P1G].
16La vitesse de la sénescence foliaire (Vsf, cm2.j-1) a été déterminée lors de la campagne 2002-2003. Elle est estimée par régression de la surface verte sur le temps, compté en jours calendrier à partir de la date d'épiaison. La durée de vie (Dsf) de la surface verte de la feuille étendard est calculée par le rapport de la surface verte (SF, cm2) au stade épiaison divisée par la vitesse de dessèchement (Vsf, cm2.j-1) : Dsf (j) = SF / Vsf.
17Les sucres solubles du pédoncule ont été déterminés au stade épiaison selon le protocole suivant : 200 mg de matière sèche du col de l'épi sont fixés dans 10 ml d'éthanol absolu pendant 60 minutes. On ajoute aux différents tubes à essai traités 3,5 ml d'eau distillée. On place les différents tubes bien fermés, pendant 2 heures, dans un bain-marie dont la température a été portée au préalable à 65 °C.
18Après traitement à la température, on laisse les tubes à l'obscurité pendant 48 heures puis on fait réagir l'extrait avec la solution de l'acide sulfurique et l'anthrone pendant 8 minutes à 92 °C. Ensuite on procède à la lecture de l'absorbance au spectromètre à 528 nm, avec l'éthanol comme témoin.
19Les lectures obtenues sont ensuite traduites en μg.ml-1 de solution, en utilisant la courbe d'étalonnage, établie suite à la lecture de l'absorbance des solutions de concentration connue de glucose variant de 5 à 50 μg.ml-1 de solution.
2.4. Analyse des données
20Une analyse de la variance à deux critères de classification, environnement et génotype (Tableau 3) a été faite pour les variables mesurées sur les différentes répétitions. Le modèle adopté dans l'analyse est le modèle additif (Steel et al., 1982) : Yijk = μ + Ei +Gj + GxEij + B(E)jk + eijk où
21Yijk est la valeur mesurée dans l'environnement i, sur le génotype j, du bloc k, pour un caractère donné,
22μ est la moyenne générale du caractère mesuré sur les i environnements, les j génotypes et les k blocs,
23Ei est l'effet de l'environnement I,
24Gj est l'effet du génotype j,
25GxEij est l'interaction entre le génotype j et l'environnement i,
26B(E)jk est l'effet des blocs k, hiérarchisés aux environnements i,
27eijk est la résiduelle du modèle.

3. Résultats et discussion
28L'analyse de la variance des variables mesurées indique un effet année significatif (Tableau 4). Cet effet met en évidence l'influence des fluctuations des conditions de croissance d'une campagne à l'autre et les différences de sensibilité vis-à-vis de ces fluctuations extériorisées par les génotypes évalués.
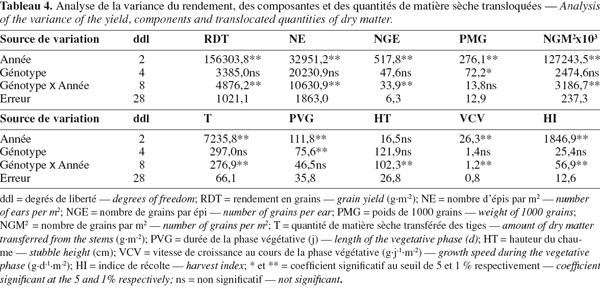
29La campagne 2000-2001 a été plus favorable à l'expression du rendement en grains, du nombre d'épis par m2, des grains par m2 et de la fertilité des épis mais pas du poids de 1000 grains. Il y a eu une plus grande quantité d'assimilats transférés associés à une plus courte durée de la phase végétative, une vitesse de croissance végétative plus réduite et un meilleur indice de récolte. Peu de différences pour la hauteur du chaume apparaissent entre les années (Tableau 5).
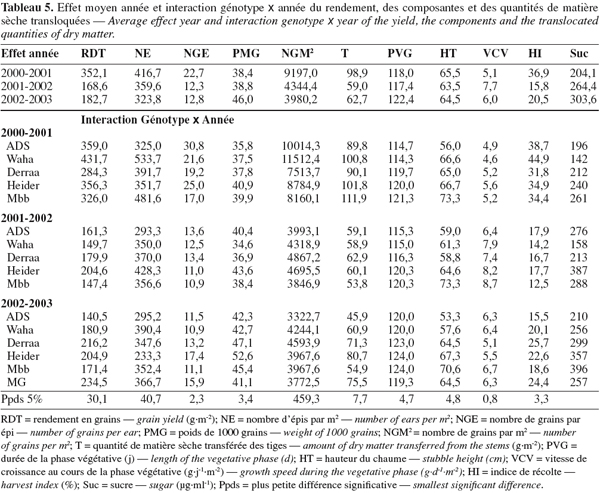
30L'année 2001-2002 a été plus défavorable notamment pour le rendement en grains. La vitesse de croissance végétative a été plus élevée mais la faiblesse de l'indice de récolte indique que la biomasse accumulée au stade épiaison n'a pas bien été valorisée sous forme de grains suite à l'effet des stress hydriques intenses agissant en post-anthèse et qui ont surtout affecté le nombre de grains par épi (Tableau 5).
31La réponse moyenne des génotypes aux conditions de croissance de la campagne 2002-2003 a été proche de celle de la campagne 2001-2002. Les moyennes observées sont très similaires sauf pour la durée de la phase végétative qui a été plus longue de 5 jours (Tableau 5).
32L'effet moyen génotype, testé par rapport à la variance d'interaction, n'est significatif que pour le poids de 1000 grains et la durée de la phase végétative. Pour les autres caractères, l'importance de la variance d'interaction rend non significatif l'effet moyen génotype et donc les différences entre génotypes doivent être approchées par campagne (Tableau 5).
33Waha présente le meilleur rendement en grains en 2000-2001. Cette performance est associée à la montée d'un grand nombre d'épis par m2 conduisant à un nombre de grains par m2 et un indice de récolte élevés. Pour ADS, on observe une bonne fertilité de l'épi ; Heider présente le meilleur PMG et Mbb la plus grande quantité d'assimilats transloquée (Tableau 5).
34Heider conduit au meilleur rendement en grains au cours de la deuxième et troisième année. Ces performances sont associées à une amélioration des trois composantes, épis par m2, grains par épi, poids de 1000 grains et à la capacité de translocation mais avec un indice de récolte similaire à celui des autres génotypes (Tableau 5).
35L'augmentation du rendement en grains de ce génotype est proportionnelle à celle de la biomasse aérienne conduisant à peu de variation de l'indice de récolte. Du point de vue de la durée de la phase végétative et pour les trois campagnes, ADS et Waha sont plus précoces, Derraa est intermédiaire et Heider et Mbb sont plus tardifs.
36La hauteur du chaume est à l'avantage de Mbb et Heider. ADS et Waha sont plus courtes et Derraa est de taille intermédiaire. Les valeurs mesurées par le taux de croissance végétative et la vitesse de remplissage du grain varient en fonction des génotypes et des années (Tableaux 5 et 6).
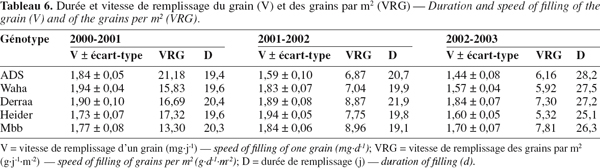
37Waha présente la meilleure vitesse de remplissage du grain en 2000-2001, Heider en 2001-2002 et Derraa en 2002-2003. Mbb remplit moins vite le grain en 2000-2001 et ADS en 2001-2002 et 2002-2003 (Tableau 6).
38Ramenées au nombre de grains produits par unité de surface, les différences du point de vue de la vitesse de remplissage deviennent plus évidentes entre génotypes. Elles sont plus élevées en 2000-2001 et moins élevées au cours des deux années suivantes. La durée de remplissage du grain suit l'évolution inverse, donc lorsque la vitesse de remplissage des grains augmente, la durée de remplissage diminue (Tableau 6). Les différences entre génotypes sont peu marquées pour la durée de remplissage.
39Au stade épiaison, ADS et Derraa développent une feuille étendard de grande surface avec une moyenne de 15,5 et 16,5 cm2, alors que Waha avec 11,5 cm2 présente une feuille de faible surface. MBB et Heider ont des feuilles de dimension proche de celles de Waha (Tableau 7).
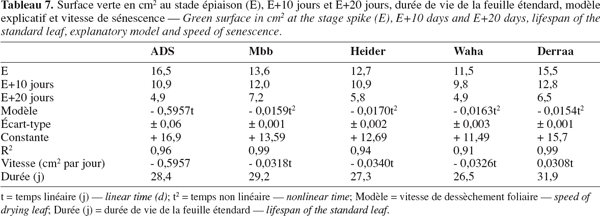
40Dix jours après l'épiaison, dès le début de la phase active de remplissage du grain, la surface verte de la feuille étendard est réduite de 33,9 %, 11,7 %, 14,1 %, 14,7 % et 17,4 % respectivement pour ADS, Mbb, Heider, Waha et Derraa.
41Après 20 jours, la réduction relative est de 70,3 %, 47,0 %, 54,3 %, 57,3 % et 58,0 % pour les mêmes génotypes dans l'ordre cité ci-dessus (Tableau 6). ADS dessèche plus vite la feuille étendard à l'opposé de Mbb qui développe un rythme de dessèchement relativement plus lent (Figure 2).
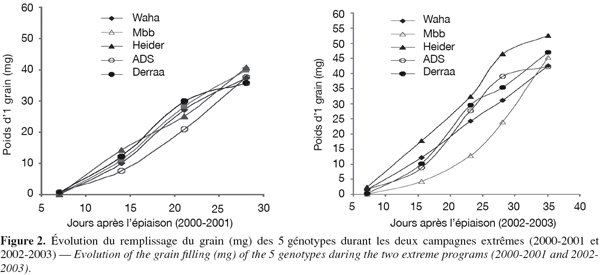
42La vitesse de dessèchement foliaire est de -0,5957 cm2 par jour pour ADS, celle des autres variétés est variable dans le temps (Figure 3, Tableau 7). Elle est modeste du stade épiaison jusqu'à l'épiaison + 15 jours, puis elle devient plus rapide après ce stade (Figure 3). La durée de vie de la feuille étendard varie de 26,5 jours pour Waha à 31,9 jours pour Derraa (Tableau 7).
43L'étude des liaisons entre le rendement en grains, la quantité des assimilats transférée, la vitesse et la durée de remplissage du grain et les différentes variables analysées indiquent que le rendement ne présente pas de liaison significative avec la durée et la vitesse de remplissage du grain, le poids de 1000 grains, la durée de la phase végétative et la hauteur du chaume (Tableau 8).
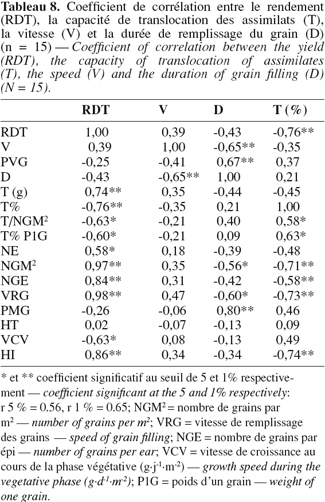
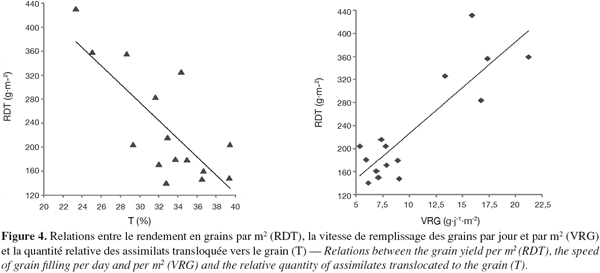
44Il est cependant significativement corrélé aux épis par m2, au nombre de grains par m2, au nombre de grains par épi et à la quantité des assimilats transloquée (Figure 4). Il est négativement corrélé à la quantité relative (T en % RDT) d'assimilats transloquée (Figure 3), à la quantité transférée par grain, à la quantité traduite en % du poids moyen d'un grain et au taux de croissance végétative (Tableau 8).
45La vitesse de remplissage du grain est négativement liée à la durée de remplissage. Cette dernière variable est positivement liée au poids de 1000 grains et à la durée de la phase végétative. Elle est liée négativement au nombre de grains par m2 et à la vitesse de remplissage des grains par m2 (Tableau 8).
46En outre, la quantité d'assimilats transloquée montre des liaisons négatives avec les variables positivement corrélées avec le rendement en grains, dont entre autres les épis par m2, les grains par m2, les grains par épi, la vitesse de remplissage des grains par m2 et l'indice de récolte. Elle est positivement corrélée avec la part des assimilats transférée par grain (Tableau 8).
47Ces résultats indiquent que lorsque les conditions de croissance et la capacité génotypique conduisent à la réalisation d'un nombre de grains par m2 élevé via les épis par m2 ou les grains par épi, elles aboutissent à l'extériorisation d'une vitesse de remplissage des grains par m2 et d'un indice de récolte élevé dont la résultante est l'expression d'un haut rendement en grains (Figure 4).
48Ces mêmes conditions sont associées à un transfert de grandes quantités d'assimilats stockés dans les tiges. La part des assimilats venant des tiges participe, cependant, de moins en moins à mesure que le milieu permet l'expression d'un meilleur rendement en grains.
49La phase de remplissage est la continuité du processus de production mis en place dès la levée et dont la finalité est le rendement en grains. Le rendement est lui-même la résultante de l'exploitation des sites de grains par m2 (épis par m2 et grains par épi) et de leur remplissage (poids moyen d'un grain).
50Au cours du déroulement de la phase de remplissage, la cinétique d'accumulation de la matière sèche du grain est sous la dépendance de deux sources principales d'assimilats qui sont la photosynthèse de la feuille étendard et la part des réserves (produites lors des périodes de pré- et post-anthèse et stockées dans les tiges) qui est transloquée.
51L'importance relative de ces assimilats est dépendante de la quantité de matière sèche accumulée au stade épiaison qui détermine le potentiel des assimilats stockés et celui de sites à remplir. Elle est dépendante aussi des conditions de croissance de la post-anthèse qui favorisent ou non une activité photosynthétique optimale.
52Une longue durée de remplissage est souvent indicatrice d'une activité photosynthétique optimale, par contre une vitesse de remplissage élevée est indicatrice des effets des stress hydriques (Sofield et al., 1977).
53Les quantités des assimilats transloquées dépendent de l'état de fonctionnement des vaisseaux conducteurs, de la capacité du génotype à utiliser les réserves pour le remplissage du grain et de l'écart du poids du grain de son potentiel suite au déficit de l'activité de photosynthèse de la post-anthèse (Wardlaw, 2002).
54Ainsi, lorsque les conditions climatiques sont moins contraignantes lors de la phase de remplissage, la plante fait moins appel aux réserves d'assimilats et arrive à assurer le remplissage du grain avec le produit de l'activité de photosynthèse de la post-anthèse.
55Par contre, lorsque les conditions climatiques durant cette phase sont contraignantes, l'activité photosynthétique de la post-anthèse est réduite ou devient nulle, la plante fait alors appel aux assimilats stockés dans les tiges, réduit la durée de la phase végétative et accroît son taux de remplissage.
56Dans ce cas de figure, le rendement obtenu dépend du degré de réduction de la durée de remplissage, de celui de l'inhibition de la photosynthèse, de l'augmentation de la vitesse de remplissage et de la quantité des assimilats transloquée. Ces caractéristiques sont génotype dépendants.
57Pour un environnement variable, le génotype idéal est celui qui évite le stress hydrique grâce à la modulation de son cycle de développement. Il doit être capable de maintenir l'activité photosynthétique sous stress hydrique le plus longtemps possible, être apte à augmenter son taux de remplissage dans le cas où la durée de remplissage est fortement réduite et être en mesure d'utiliser les assimilats stockés si son potentiel de poids du grain est affecté.
58Des résultats contradictoires sont rapportés dans la littérature en ce qui concerne la contribution de la vitesse et de la durée de remplissage au rendement en grains. Gebeyehou et al. (1982) trouvent que la durée contribue beaucoup plus que la vitesse au rendement en grains alors que Nass et al. (1975) ainsi que Triboi et al. (1985) rapportent par contre que l'effet de la vitesse sur le rendement est de loin plus important que la durée de remplissage. Ce que corroborent les résultats de la présente étude.
59Gebeyehou et al. (1982) rapportent des valeurs pour la vitesse de remplissage variant de 12,4 à 15,5 g.j-1.m-2 pour deux années consécutives et pour 11 génotypes de blé dur différents, alors que la durée de remplissage variait de 33,0 à 40,3 jours. Ces résultats sont assez similaires à ceux de cette étude, sauf que la durée est plus courte dans le cas de la présente étude.
60Gebeyehou et al. (1982) notent une corrélation phénotypique non significative entre la vitesse et la durée et une corrélation environnementale significative et négative qui explique que les conditions de croissance qui favorisent la vitesse de remplissage défavorisent la durée de remplissage.
61Sofield et al. (1977) mentionnent que la vitesse de remplissage est plus élevée sous haute température et que la durée est plus longue sous température modérée. Biddinger et al. (1977) rapportent que la contribution des tiges au rendement variait de 10 à 70 % selon les génotypes et les environnements.
62Ces stress hydriques réduisent des assimilats stockés dans les tiges après l'épiaison et affectent l'appareil photosynthétique au cours de la phase de remplissage. Sous de telles conditions, le rendement est la résultante de l'activité photosynthétique de la post-anthèse. Les résultats de la présente étude montrent que les assimilats participent au remplissage du grain en situation défavorable, mais que cette participation est relativement moindre au cours des années favorables.
63Ceci corrobore ce qui est rapporté par Triboi et al. (1985) et par Blum et al. (1996) qui mentionnent que le flux des assimilats vers le grain dépend d'une part de la quantité d'assimilats stockés dans les tiges et d'autre part de l'assimilation post-anthèse. La quantité est fonction du nombre de tiges produites par m2, du type de variété, de la hauteur du chaume et des conditions climatiques spécifiques à l'année (Blum et al., 1996).
64Les résultats de la présente étude indiquent que dans l'échantillon restreint de génotypes étudiés, la vitesse de remplissage contribue au rendement via le nombre de grains par m2, alors que la durée affecte le poids de 1000 grains. Ces deux variables se compensent mutuellement.
65La contribution relative des assimilats de la pré-anthèse au rendement en grains ne devient importante que lorsque le rendement est faible, car l'activité photosynthétique sera affectée par le stress hydrique, ou bien aussi lorsque les conditions de croissance de la pré-anthèse sont telles que la plante aborde la phase de remplissage fortement affectée par le stress hydrique. La sénescence de la feuille étendard ne montre pas de liaison significative avec les principaux paramètres analysés.
66Dans ce contexte, Fokar et al. (1996) trouvent que les génotypes qui se caractérisent par une sénescence foliaire précoce font plus appel aux hydrates de carbone solubles dans l'eau (WSC). Ceci indique que la capacité d'utilisation des WSC est une caractéristique constitutive.
4. Conclusion
67L'expression d'un rendement élevé est associée positivement à un nombre d'épis par m2, aux nombres de grains par m2 et de la fertilité des épis mais négativement liée au poids de 1000 grains.
68Cette situation est associée aussi à une plus grande quantité d'assimilats transférée, une plus courte durée de la phase végétative, une vitesse de croissance végétative plus réduite et un meilleur indice de récolte.
69La vitesse de dessèchement foliaire est modeste au stade épiaison puis devient plus rapide 10 jours après, elle prend la même allure pour les 5 génotypes testés, avec un rythme plus élevé pour le génotype ADS.
70L'étude des liaisons entre le rendement en grains et les différentes variables analysées indique que la part des assimilats venant des tiges diminue à mesure que le milieu permet l'expression d'un meilleur rendement en grains, car lorsque les conditions climatiques sont moins contraignantes, la plante arrive à assurer le remplissage du grain avec le produit de l'activité de photosynthèse de la post-anthèse.
71Le génotype idéal est celui qui sera apte à maintenir, sous conditions défavorables, l'activité photosynthétique et à augmenter son taux de remplissage grâce à l'utilisation des assimilats stockés dans la tige.
Bibliographie
Abbassenne F., Bouzerzour H. & Hachemi L., 1998. Phénologie et production du blé dur (Triticum durum Desf.) en zone semi-aride. Ann. Agron. INA, 18, 24-36.
Baldy C., Ruelle P. & Fernandes A., 1993. Résistance à la sécheresse du sorgho-grain en climat méditerranéen. Sécheresse, 4, 85-93.
Biddinger F., Muscrave R.B. & Fischer R.A., 1977. Contribution of stored pre-anthesis assimilates to grain yield in wheat and barley. Nature, 20, 431-433.
Blum A. & Pnuel Y., 1996. Physiological attributes associated with drought resistance of wheat cultivars in a mediterranean environment. Aust. J. Agric. Res., 41, 799-810.
Fokar M., Nguyen H.T. & Blum A., 1996. Heat tolerance in spring wheat II. Grain filling. Euphytica, 104, 9-15.
Gebeyehou G., Knott D.R. & Baker R.J., 1982. Relationships among duration of vegetative and grain filling phases, yield components and grain yield in durum wheat cultivars. Crop Sci., 22, 287-290.
McIntosh M.S., 1983. Analysis of combined experiments. Agron. J., 75, 153-156.
Nass H.G. & Resier B., 1975. Grain filling period and grain yield relationships in spring wheat. Can. J. Plant Sci., 55, 673-678.
Sofield T., Evans J., Cook M.G. & Wardlaw I.F., 1977. Factors influencing the rate and duration of grain filling in wheat. Aust. J. Plant Physiol., 4, 785-797.
Steel R.G.D. & Torrie J.H., 1982. Principles and procedures of statistics, a biometrical approach. New York: McGraw-Hill Inc.
Triboi E., Planchon J. & Magne J., 1985. Déterminisme du poids moyen du grain chez le blé. Effet sur la variation du rendement. C. R. Acad. Agric. Fr., 71, 871-886.
Wardlaw I.F., 2002. Interaction between drought and chronic high temperature during kernel filling in wheat in a controlled environment. Ann. Bot., 90, 469-476.
To cite this article
About: Fayçal Bahlouli
Université Mohamed Boudiaf. Faculté des Sciences. 28000 M'sila. Algérie.
E-mail : faycal.bahlouli@caramail.com
About: Hamenna Bouzerzour
Université Ferhat Abbas. Faculté des Sciences. 19000 Sétif. Algérie.
About: Ammar Benmahammed
Université Ferhat Abbas. Faculté des Sciences. 19000 Sétif. Algérie.
