


- Home
- Volume 30 (2026)
- Numéro 3
- Promotion of carob (Ceratonia siliqua L.) seedling growth by Streptomyces spp. and Pseudomonas fluorescens in an alkaline and infertile soil


View(s): 39 (0 ULiège)
Download(s): 6 (0 ULiège)

Promotion of carob (Ceratonia siliqua L.) seedling growth by Streptomyces spp. and Pseudomonas fluorescens in an alkaline and infertile soil

Attached document(s)
original pdf fileRésumé
Promotion de la croissance des plantules du caroubier (Ceratonia siliqua L.) par Streptomyces spp. et Pseudomonas fluorescens dans un sol alcalin et infertile
Description du sujet. La réhabilitation durable des sols dégradés constitue un enjeu majeur. Cette étude vise à évaluer le potentiel de couples symbiotiques plantes-bactéries indigènes pour la phytoremédiation d'un site dégradé.
Objectifs. Isoler et sélectionner des souches bactériennes promotrices de la croissance des plantes (PGP) adaptées aux conditions locales afin d'améliorer l'établissement du caroubier (Ceratonia siliqua), une espèce d'intérêt socio-économique.
Méthode. Soixante-quatre isolats d'actinobactéries ou de Pseudomonas fluorescens ont été isolés de la rhizosphère de légumineuses pionnières (Lotus creticus, Retama monosperma) du site. Un criblage phénotypique a évalué leurs traits PGP (production d'acide indole-acétique - AIA, solubilisation du phosphate, antagonisme fongique). Les isolats les plus performants ont été utilisés pour inoculer des plants de caroubier cultivés pendant deux mois en pépinière sur un substrat constitué du sol alcalin et infertile du site, préalablement stérilisé. Les paramètres de croissance ont été mesurés et analysés par Analyse en Composantes Principales (ACP).
Résultats. Le criblage a identifié des isolats aux profils complémentaires : RP5 (production élevée d'AIA : 36,09 µg·mL-1), RP1 (solubilisation du phosphate : halo de 11 mm) et CAARL30 (activité antagoniste contre Alternaria alternata, Aspergillus sp., Cladosporium sp. et Fusarium sp.). L'isolat CAARL35 (Streptomyces sp.) a montré un équilibre entre la production d'AIA et la solubilisation du phosphate. Les tests en pépinière ont confirmé l'efficacité des inoculations. L'ACP a désigné les traitements les plus performants : l'isolat CAARL35, les isolats RP1, RP2, RP5 (P. fluorescens) et la combinaison synergique GAL4+RP1. L'inoculation avec CAARL35 a induit les améliorations les plus significatives, augmentant jusqu'à 41 % la biomasse sèche, 35 % la longueur racinaire et 77 % le nombre de folioles par rapport au témoin non inoculé.
Conclusions. Cette étude démontre le potentiel de souches bactériennes rhizosphériques indigènes, en particulier la souche Streptomyces sp. CAARL35, comme bio-inoculant efficace pour améliorer la croissance du caroubier dans des conditions de sol stressantes. La sélection de couples performants plante-bactérie, adaptés aux contraintes édaphiques locales, offre une stratégie prometteuse et écologique pour la réhabilitation des écosystèmes dégradés.
Abstract
Description of the subject. The sustainable rehabilitation of degraded soils is a major challenge. This study aims to evaluate the potential of indigenous plant-bacteria symbiotic associations for the phytoremediation of a degraded site.
Objectives. To isolate and select Plant Growth-Promoting (PGP) bacterial strains adapted to local conditions to enhance the establishment of carob trees (Ceratonia siliqua), a species of significant socio-economic interest.
Method. Sixty-four isolates consisting of actinobacteria or Pseudomonas fluorescens were obtained from the rhizosphere of pioneer legumes (Lotus creticus, Retama monosperma) from the site. A phenotypic screening assessed their PGP traits (indole-3-acetic acid [IAA] production, phosphate solubilization, fungal antagonism). The most promising isolates were used to inoculate carob seedlings grown for two months in a nursery on a sterilized substrate composed of the site's native alkaline and infertile soil. Growth parameters were measured and analyzed by Principal Component Analysis (PCA).
Results. The screening identified isolates with complementary profiles: RP5 (high IAA production: 36.09 µg·mL-1), RP1 (phosphate solubilization: halo of 11 mm), and CAARL30 (antagonistic activity against Alternaria alternata, Aspergillus sp., Cladosporium sp., and Fusarium sp.). Isolate CAARL35 (Streptomyces sp.) showed a balanced profile of both IAA production and phosphate solubilization. Nursery trials confirmed the effectiveness of the inoculations. PCA identified the most effective treatments: isolate CAARL35, isolates RP1, RP2, RP5 (P. fluorescens), and the synergistic combination GAL4+RP1. Inoculation with CAARL35 induced the most significant improvements, increasing dry biomass by up to 41%, root length by 35%, and the number of leaflets by 77% compared to the non-inoculated control.
Conclusions. This study demonstrates the potential of indigenous rhizobacterial strains, particularly Streptomyces sp. CAARL35, as an effective bioinoculant for enhancing carob tree growth under stressful soil conditions. The selection of high-performing plant-bacteria partnerships, adapted to local edaphic constraints, provides a promising and eco-friendly strategy for the rehabilitation of degraded ecosystems.
Table of content
Received 4 October 2024, accepted 26 May 2026, available online 17 June 2026.
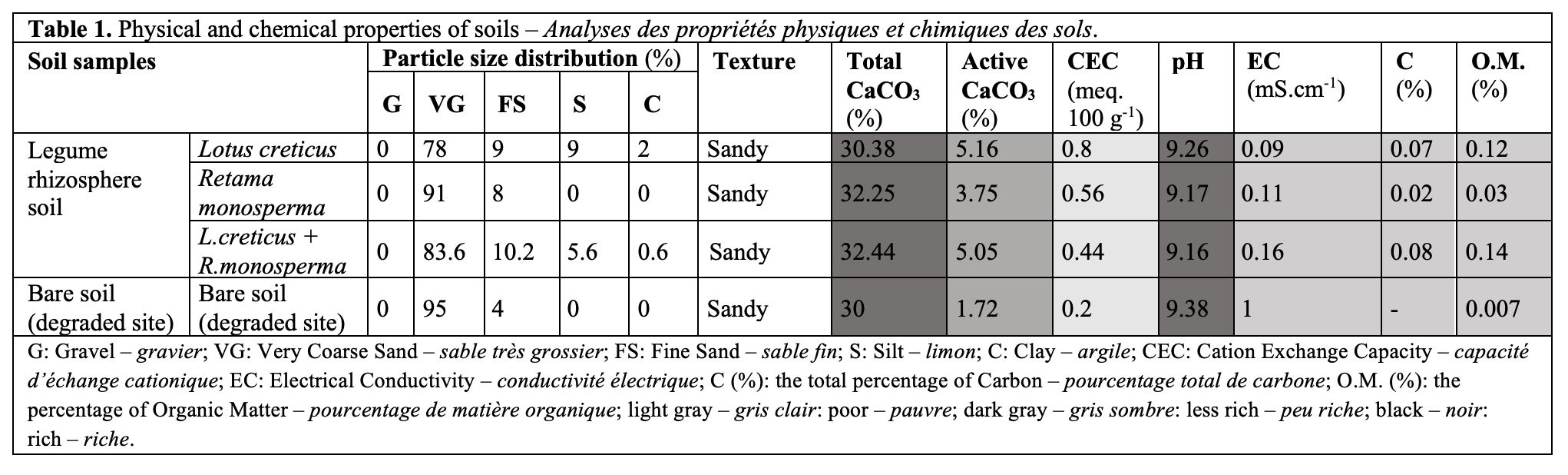
This article is distributed under the terms and conditions of the CC-BY License (https://creativecommons.org/licenses/by/4.0)
1. INTRODUCTION
1The carob tree (Ceratonia siliqua L.) is a dioecious species of the legume family, native to the Mediterranean region (Ortiz et al., 1995). It is renowned for its ability to thrive in arid conditions and nutrient-deficient soils (Thomas et al., 2024). Today, it is cultivated primarily for its socio-economic and ecological importance (Haddarah, 2013). All parts of the tree are used in a wide range of applications in the food, pharmaceutical, and cosmetic industries. Its seeds are subject to commercial transactions whose value far exceeds that of most forest plants (Ait Chitt et al., 2007). Consequently, the carob tree can be considered an excellent candidate for rehabilitating degraded soils (Martins-Loucao et al., 2024).
2Its rhizosphere harbors a diverse microbial community (Dalli et al., 2020). Many of these microorganisms, known as plant growth-promoting rhizobacteria (PGPR), colonize the rhizosphere and closely associate with plant roots (Vacheron et al., 2013). Through direct and indirect mechanisms, PGPR enable plants to thrive in challenging environmental conditions (Lavudi et al., 2023; Hyder et al., 2024; Zhang et al., 2024). They enhance plant nutrition by biologically fixing nitrogen, solubilizing phosphorus, and producing phytohormones such as auxins and gibberellins. They also activate the plant immune system and suppress phytopathogens by producing various metabolites, including hydrogen cyanide (HCN), antibiotics, and lytic enzymes (Khan et al., 2021; Meena et al., 2023). Consequently, PGPR are regarded as a promising tool for sustainable agriculture and a viable substitute for chemical fertilizers (Backer et al., 2018).
3The inoculation of effective microbes, such as PGPR, into plants can increase biomass by more than 64% (Pereira et al., 2020; Azizi et al., 2022), while also enhancing biodiversity, improving soil health, and reducing the need for chemical inputs. However, this approach faces several challenges, particularly regarding the selection of effective strains (Gufwan et al., 2025). In the context of carob cultivation, effective PGPR inoculants can enhance growth by protecting the plant against various fungal pathogens. For instance, Alternaria alternata is responsible for "Ceratonia Blight", a disease characterized by the appearance of black spots on the leaves (Parlascino et al., 2024). Similarly, Fusarium oxysporum infects the root system, inducing wilting that can ultimately lead to the death of the tree in severe cases (Mounir, 2024).
4Although the carob tree shows great potential for reforestation projects, programs that use selected PGPR combined with carob plants constitute a significant research gap (Aeron et al., 2011). Promising candidates for such applications include bacteria from genera like Streptomyces spp. and Pseudomonas spp., whose use in promoting carob tree growth is a notable example (Aeron et al., 2011).
5Reforestation success in sandy quarries within hostile, arid environments is frequently constrained by characteristic challenges: calcareous soils, high alkalinity, and poor fertility. This research investigates a potential biological solution by examining whether the synergistic partnership between the carob tree (Ceratonia siliqua) and selected indigenous rhizobacteria can enhance plant stress tolerance, thereby transforming a key restoration challenge into a viable ecological strategy.
2. MATERIALS AND METHODS
2.1. Soil samples
6Soil samples were collected in autumn 2018 at the Terga sand quarry, located about 85 km from Oran in northwestern Algeria (35°26'35.1"N 1°13'25.9"W). Rhizospheric soil was taken from three different zones: under Retama monosperma, under Lotus creticus, and from areas where both plants grew together. In each case, soil was collected at a depth of 20 cm in the root zone of plants. Bare soil samples were also taken from several locations across the degraded site. All samples were collected in triplicate following the method described by Pochon & Tardieux (1962) and were stored in plastic bags at 4 °C until analysis. Each sample was then divided into two parts: one for isolating PGPR, and the other for physicochemical analysis. The bare soil was later used as a substrate for greenhouse tests. All tests were carried out within the days following sampling.
2.2. PGPR isolation
7Actinobacteria. In order to isolate actinobacteria, serial soil dilutions ranging from 10-1 to 10-5 were prepared in 9 mL of 0.85% NaCl solution and then introduced and cultivated on isolation media namely casein starch agar (CAA) (Sharma et al., 2011) and glucose asparagine agar (GAA) (El-Nakeeb & Lechevalier, 1963) supplemented with 50 µg·mL-1 of fluconazole. The cultures were then incubated at 30 °C for 15 days. Characteristic actinobacteria colonies were then separated and purified using Bennett medium, as described by Jones (1949) and Bouznada et al. (2017) for morphological and biochemical analysis.
8Fluorescent Pseudomonas spp. Isolation of these strains was conducted after preparing serial dilutions 10-1 to 10-5 of 1 g of rhizospheric soils. King B agar was used for isolation (Bultreys & Gheysen, 2000). The incubating conditions were at 28 °C for 72 h. Fluorescence due to pyoverdine production was observed at a wavelength of 360 nm and then purified using the same agar medium. The morphological and biochemical approaches were conducted afterwards (Joffin & Leyral, 2006; Sebihi, 2016).
2.3. Catalase test
9Catalase activity was evaluated using a standard qualitative method. A 24-hour-old bacterial colony was aseptically picked with a sterile inoculating loop and placed on a clean glass slide. A drop of hydrogen peroxide solution (H₂O₂, 3%) was then added. Rapid effervescence and bubble production were interpreted as a positive result, indicating the breakdown of hydrogen peroxide into water and oxygen. The absence of bubble formation was considered a negative result (Taylor & Achanzar, 1972).
2.4. Oxidase test
10Oxidase activity was determined using a commercial oxidase test disc method. A small amount of a 24-hour-old bacterial colony was transferred onto a disc impregnated with freshly prepared reagent (tetramethyl-p-phenylenediamine dihydrochloride). A color change to dark purple within a few seconds was considered a positive result, indicating the presence of cytochrome c oxidase. No color change or a delayed reaction was recorded as a negative result (Steel, 1961).
2.5. Analysis of PGP effects
11Phosphate solubilization. In order to assess the isolates' capacity to dissolve phosphate, spot inoculation was carried out on Pikovskaya (PVK) medium, as described by Pikovskaya (1948). Its composition consists exclusively of Ca3(PO4)2 as the phosphate source and bromophenol blue as the pH indicator. The culture was thereafter incubated at 28 °C for 7 days. Clear zones around the bacterial colonies were taken as evidence for phosphate solubilization. The phosphate solubilization diameter is determined as described by Laabas et al. (2017).
12Production of Indole-3-Acetic Acid (IAA). Bacterial suspensions 1.5 x 108 cfu (colony forming unit·mL-1) of actinobacteria and fluorescent Pseudomonas spp., along with 0.1% L-tryptophan (a precursor to indole-3-acetic acid), were subjected to centrifugation at 10,000 rpm for a duration of 5 min (Malik & Sindhu, 2011). Next, 2 mL of the liquid remaining after centrifugation was combined with 4 mL of Salkowski reagent. The latter consists of 1 mL of a solution containing 0.5 M FeCl3 in 50 mL of 35% HClO4. The pink coloration serves as an indicator of IAA synthesis (Gang et al., 2019). The optical density (O.D.) was measured at 530 nm using a Hitachi U-5100 spectrophotometer. The concentration of IAA was then quantified in µg·mL-1 by reference to a standard curve (De Barros Silva Leite et al., 2018) following the colorimetric method of Gordon & Weber (1951).
13Antifungal activity. Actinobacteria's antifungal activity was tested using the agar cylinder method against phytopathogenic filamentous fungi (Alternaria alternata, Aspergillus sp., Cladosporium sp., Fusarium sp.) obtained from the University of Oran1 applied microbiology laboratory (Thilagam & Hemalatha, 2019) using Potato dextrose agar plate. After a 4-hour pre-diffusion process at 4 °C, the fungus was incubated for 48 to 72 h. Resulting inhibitory zone indicates antifungal activity (Thibodeau et al., 2002).
2.6. Calculation of the Plant Growth Promotion Index (PGPI)
14The PGPI was estimated by the methodology proposed by Agaras et al. (2015; 2020), which incorporates both antagonistic effects and direct growth promotion.
2.7. pH tolerance
15In order to evaluate the influence of pH on the growth of actinobacteria and fluorescent Pseudomonas spp., a 0.1 ml sample of a newly prepared liquid culture was injected with the chosen strains into liquid Bennett medium and liquid King B medium, respectively. The cultures were exposed to a pH range of 4, 5, 5.5, 6.5, 7, 7.5, 8, 9, 10, and 11 by modifying the pH using 1N HCl and 1N KOH buffer. The initial absorbance at a wavelength of 600 nm was measured, and then the cultures were incubated at 30 °C. Each pH was tested using triplicate tubes. The optical density of the various strains was evaluated after a period of 48 h. The final value was derived by subtracting the initial O.D. 600 nm as described in the modified study by El Idrissi et al. (1996).
2.8. Salinity tolerance
16Triplicate samples of liquid King B medium (for fluorescent Pseudomonas spp. strains) and liquid Bennett medium (for actinobacterial strains) were each inoculated with 0.1 mL of a fresh bacterial culture at varied NaCl concentrations ranging from 0.50% to 5.00%. The initial optical density I(O.D.) 600 nm was determined, and thereafter the bacteria were cultured at 30 °C for 48 h, the optical density F(O.D.) 600 nm was measured. The increase of optical density indicates the level of bacterial growth: (O.D.) 600 nm = F(O.D.) 600 nm - I(O.D.) 600 nm (El Idrissi et al., 1996).
2.9. Impact of temperature
17The bacterial suspensions were subsequently incubated at several temperatures; 4 °C, 30 °C, 40 °C, and 50 °C, with three replicates for each temperature.
18The initial optical density I(O.D.) 600 nm of the bacterial suspension and after incubation F(O.D.) 600 nm were determined. The optical density of growth is calculated by (O.D.) 600 nm = F(O.D.) 600 nm - I(O.D.) 600 nm, as modified by El Idrissi et al. (1996).
2.10. Plant material preparation and experimental design
19Seeds of carob tree (Ceratonia siliqua L.) were surface-sterilized by immersion in 95% ethanol for 1 min, and in 10% (w/v) NaClO for 10 min, washed five times with sterile distilled water (Tabli et al., 2018). The scarification was done mechanically with preheated punch utilized to create a hole in the seed coat. These seeds are placed on sterile filter paper moistened with sterile distilled water in Petri dishes and incubated in the dark at 28 °C for 72 h. Germinated seeds with 3 cm radicles length were relocated into 200-ml pots, each containing three seeds. The pots held degraded infertile sandy soil (Table 1) that was sterilized twice.
20Plant inoculation. Each plant was inoculated with 1 ml of the preselected bacterial suspension. Those strains were preselected according to their Plant Growth Promoting Index (PGPI). The co-inoculation process was carried out by introducing fluorescent Pseudomonas spp. and actinobacteria isolates simultaneously. In contrast control was injected with 1 ml of a sterile bacterial culture medium. Immature carob plants were cultivated under a randomized complete block design with four replications. A secondary inoculation was administered to all experimental groups 30 days post-establishment. Irrigation was supplied three times per week with sterile distilled water, with one weekly application consisting of Hoagland & Arnon nutrient solution (Hernandez & Dzakovich, 2026).
21Plant growth measurement. For each treatment, growth parameters were measured on four replicate plants. The shoot length (SL) and the root length (RL) were measured via a precision ruler. The leaf number (LN) was determined immediately after harvest. The fresh weight (FW) was determined via an analytical balance immediately after harvest, whereas the dry weight (DW) was recorded after drying the samples at 60 °C until they reached a stable weight.
22Statistical analysis. Plant growth parameters were subjected to a one-way ANOVA. Post-hoc comparisons were carried out using Duncan's test (p ≤ 0.05), and PCA using Statistica 6 software to examine the effect of the isolates on each growth parameter.
3. RESULTS
3.1. Physicochemical analysis of soils
23Soil texture was consistently sandy across all samples (Table 1). The presence of plants significantly increased soil organic matter (OM) and organic carbon (C%) content compared with bare soil. The degraded soil was severely depleted in OM (0.007%), while the rhizosphere soils showed a 10 to 20-fold increase. The highest OM content (0.14%) was recorded in the co-planted treatment. While total CaCO₃ was uniformly high (~30-32.5%), pH was strongly alkaline across all treatments, ranging from 9.16 to 9.38, with no consistent trend related to plant cover.
3.2. Abundance of culturable PGPR
24Culturable analysis revealed a total of 64 distinct microbial isolates. Their abundance varied significantly depending on the leguminous plant's rhizosphere (Figure 1). Retama monosperma appears to selectively enrich for fluorescent Pseudomonas in its rhizosphere, while the co-plantation of both leguminous species creates a synergistic effect that most effectively promotes the growth of actinobacteria.
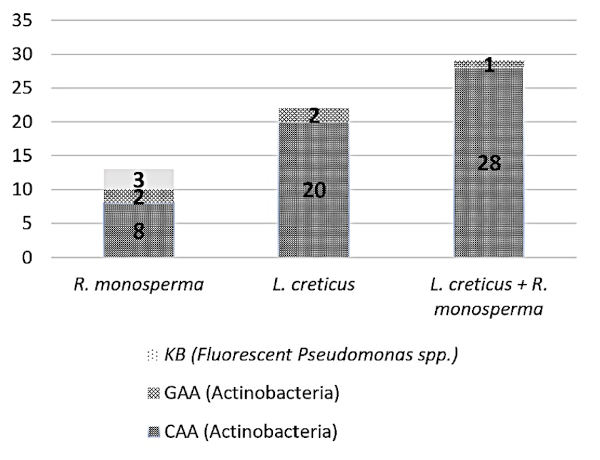
Figure 1. Number of culturable bacterial isolates in the rhizosphere of leguminous plants at the Terga site when using the following culture media: casein starch agar (CAA), glucose asparagin agar (GAA), King B (KB) – Nombre d’isolats bactériens cultivables dans la rhizosphère des plantes légumineuses du site de Terga lors de l’utilisation des milieux de culture suivants : gélose amidon-caséine (CAA), gélose glucose-asparagine (GAA), milieu King B (KB).
3.3. Screening for plant growth-promoting traits and antagonistic potential
25The evaluation of bacterial isolates for plant growth-promoting (PGP) traits revealed a significant diversity in their functional capacities (Table 2), (Figure 2 and 3).
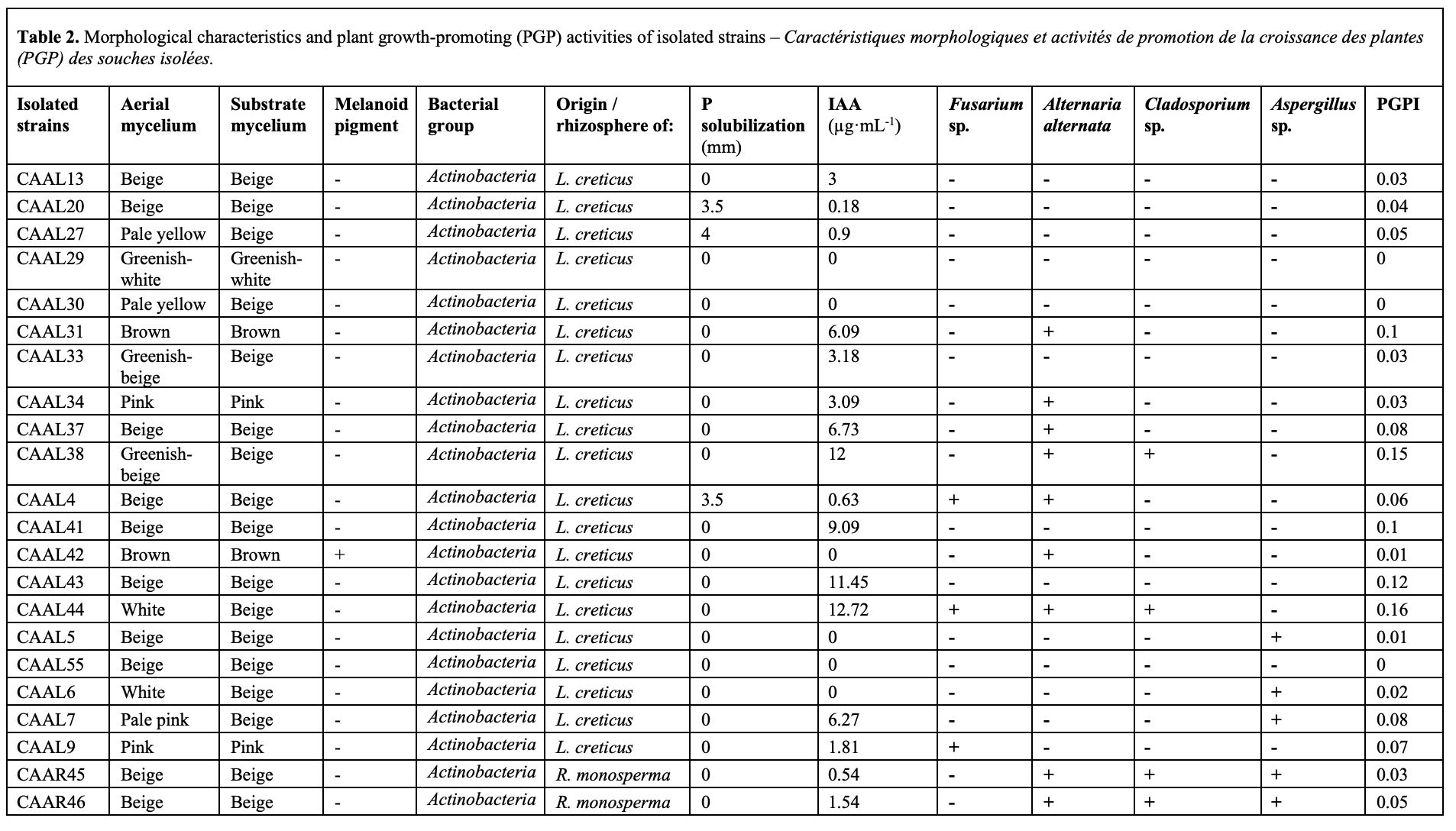
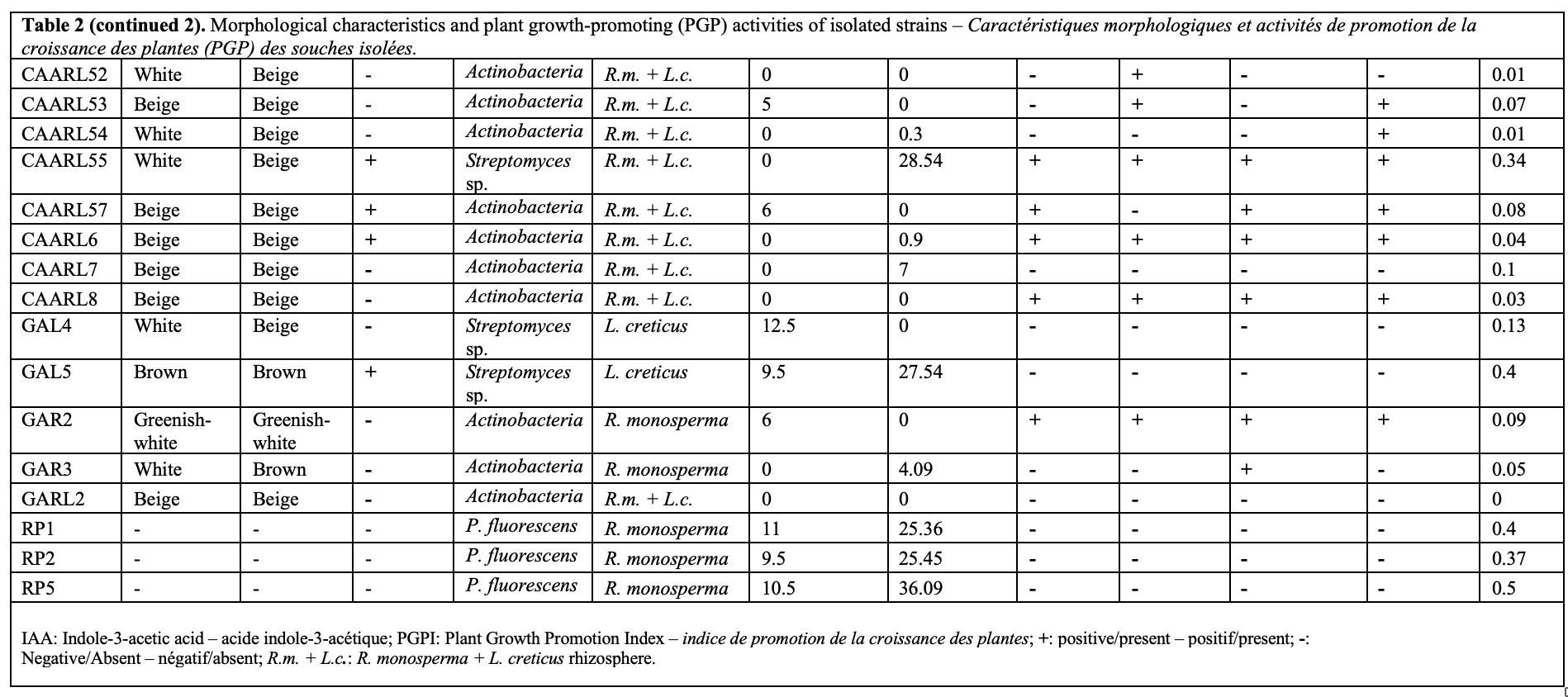
Figure 2. Functional characterization of PGPR isolates: frequency of plant-beneficial traits for the 64 isolates – Caractérisation fonctionnelle des isolats de PGPR : fréquence des traits bénéfiques pour les plantes chez les 64 isolats.
A.E: antagonistic effect – effet antagoniste.
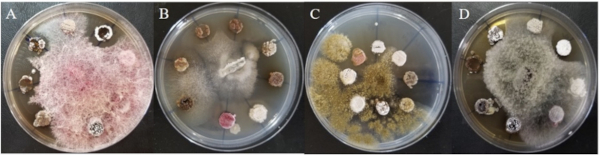 Figure 3. Antifungal effect on phytopathogenic fungi – Effet antifongique sur des champignons phytopathogènes.
Figure 3. Antifungal effect on phytopathogenic fungi – Effet antifongique sur des champignons phytopathogènes.
A: Fusarium sp. ; B: Cladosporium sp. ; C: Aspergillus sp. ; D: Alternaria alternata.
26Indole-3-Acetic Acid (IAA) production. Isolate RP5 (Pseudomonas fluorescens) demonstrated the highest auxin production yield, synthesizing 36.09 µg·mL-1 of IAA. Other notable high-producing strains included GAL5 (27.54 µg·mL-1), CAARL55 (28.54 µg·mL-1), RP1 (25.36 µg·mL-1), and RP2 (25.45 µg·mL-1). This indicates that these strains possess a strong potential to directly stimulate plant root development and growth through the production of this essential auxin.
27Phosphate solubilization. In contrast, the most efficient phosphate solubilizers were distinct from the top IAA producers. The Streptomyces sp. isolate GAL4 exhibited the largest solubilization halo with a diameter of 12.5 mm, followed by Pseudomonas fluorescens strains RP1 (11.0 mm) and RP5 (10.5 mm).
28Antagonistic activity against fungal pathogens. The Streptomyces spp. isolates CAARL55 and CAARL30 displayed the broadest-spectrum antagonism, effectively inhibiting the growth of all four tested fungal pathogens: Fusarium sp., Alternaria alternata, Cladosporium sp., and Aspergillus sp. (Figure 3). This potent antifungal activity positions them as prime candidates for the biocontrol of soil-borne diseases. Furthermore, isolates like GAR2 and CAAL44 also showed strong and broad-spectrum inhibition. The presence of melanoid pigment in some antagonistic strains (e.g., CAARL55, CAARL30') may be linked to their survival and competitive fitness in the rhizosphere.
3.4. Integrated Plant Growth-Promoting Index (PGPI)
29The PGPI, which integrates multiple PGP activities into a single value, identified the isolates exhibiting the most balanced and multifaceted PGP potential. The highest PGPI values were recorded for RP5 (0.50), GAL5 (0.40), and RP1 (0.40). These results highlight that, despite the occurrence of functional specialization among isolates, strains like RP5, which combine high IAA production, significant phosphate solubilization, and the highest overall PGPI score, represent the most promising broad-spectrum PGPR candidates. Interestingly, the strong antifungal isolates CAARL55 and CAARL30' achieved respectable PGPI values of 0.34 and 0.16, respectively, indicating that their strength in biocontrol contributes significantly to their overall PGP potential, even in the absence of phosphate solubilization.
3.5. Morphological and biochemical identification of selected strains
30Three pyoverdine-producing isolates (RP1, RP2, RP5) were obtained on King's B medium. These Gram-negative bacilli tested positive for catalase and oxidase. They were psychrotolerant (growth at 4 °C) but failed to grow at 42 °C. This phenotypic profile is consistent with classification within the Pseudomonas fluorescens species complex as described by Bergey’s systematic manual (Brenner et al., 2005).
31Four Gram-positive, filamentous bacterial isolates (CAARL35, CAARL55, GAL4, GAL5) were obtained from the rhizosphere. All isolates developed a substrate mycelium and a distinct, pigmented aerial mycelium with colors varying from white and beige to brown (Table 3). Melanin production was observed for isolates CAARL55 and GAL5. This morphological differentiation is consistent with typical characteristics for the genus Streptomyces spp. (Goodfellow et al., 2012).
3.6. Physiological characterization: effect of temperature, pH, and salinity on growth
32The growth of the bacterial isolates (RP1, RP2, RP5, and CAARL35), assessed by optical density (O.D.) measurements, was investigated under varying physicochemical conditions. With respect to temperature (Figure 4), all strains exhibited peak growth at 30 °C, followed by a sharp decline at temperatures exceeding 40 °C, indicating that these isolates are mesophilic in nature. The effect of pH on bacterial growth (Figure 5) demonstrated an optimal proliferation at near-neutral pH values (6.5–7.0), with a marked reduction in O.D. recorded under both acidic (pH < 5.5) and alkaline (pH > 9.0) conditions, reflecting a neutrophilic physiological profile. Regarding salinity tolerance (Figure 6), growth was highest at low NaCl concentrations (0.50–0.75%) and declined progressively with increasing salt concentrations, suggesting that the tested isolates are non-halophilic bacteria with limited tolerance to osmotic stress.
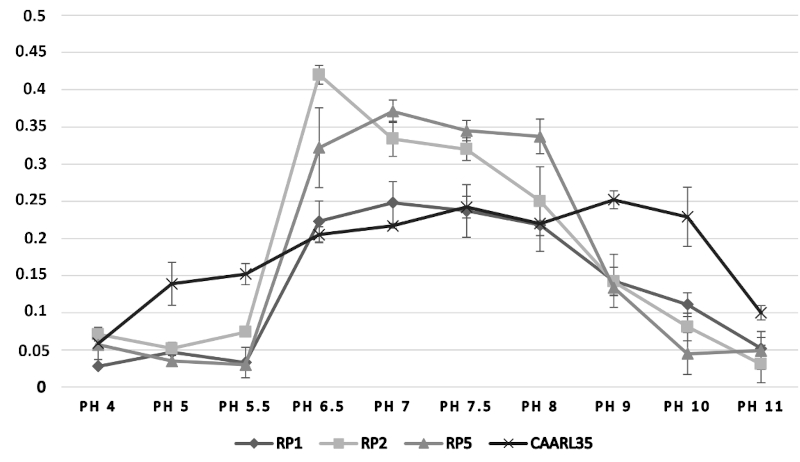
Figure 4. Effect of pH on the growth of selected strains, expressed as the change in optical density at 600 nm (O.D. 600 = Final O.D. – Initial O.D.) – Effet du pH sur la croissance des souches sélectionnées, exprimé par la variation de la densité optique à 600 nm (D.O. 600 = D.O. finale – D.O. initiale).
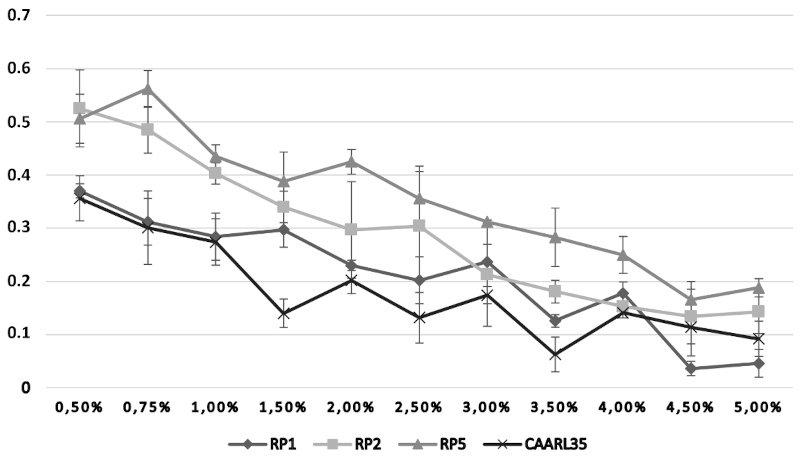
Figure 5. Effect of salinity on growth of selected strains, expressed as the change in optical density at 600 nm (O.D. 600 = Final O.D. – Initial O.D.) – Effet de la salinité sur la croissance des souches sélectionnées, exprimé par la variation de la densité optique à 600 nm (D.O. 600 = D.O. finale – D.O. initiale).
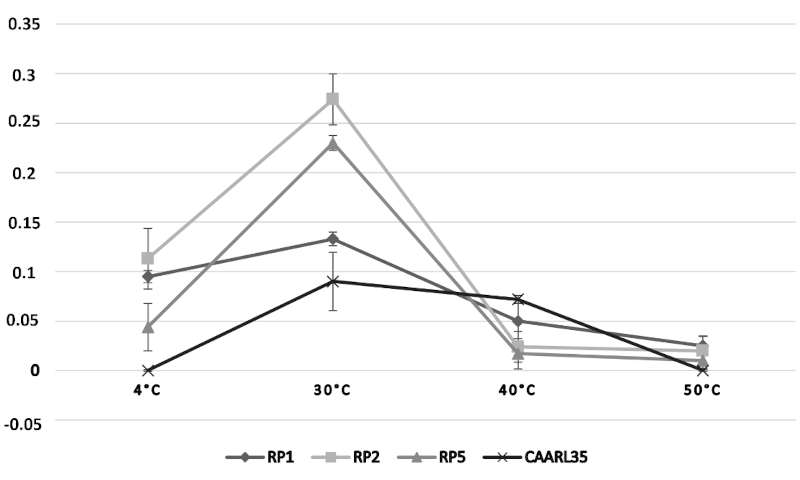 Figure 6. Effect of temperature on growth of selected strains, expressed as the change in optical density at 600 nm (O.D. 600 = Final O.D. – Initial O.D.) – Effet de la température sur la croissance des souches sélectionnées, exprimé par la variation de la densité optique à 600 nm (D.O. 600 = D.O. finale – D.O. initiale).
Figure 6. Effect of temperature on growth of selected strains, expressed as the change in optical density at 600 nm (O.D. 600 = Final O.D. – Initial O.D.) – Effet de la température sur la croissance des souches sélectionnées, exprimé par la variation de la densité optique à 600 nm (D.O. 600 = D.O. finale – D.O. initiale).
3.7. Effects on plant growth
33The results, presented in figure 7 and table 4, show significant variations among the treatments compared to the non-inoculated control.

Figure 7. Growth comparison of carob tree (Ceratonia siliqua L.) inoculated (right) versus control (left) non inoculated – Comparaison de la croissance du caroubier (Ceratonia siliqua L.) inoculé (à droite) par rapport au témoin non inoculé (à gauche).
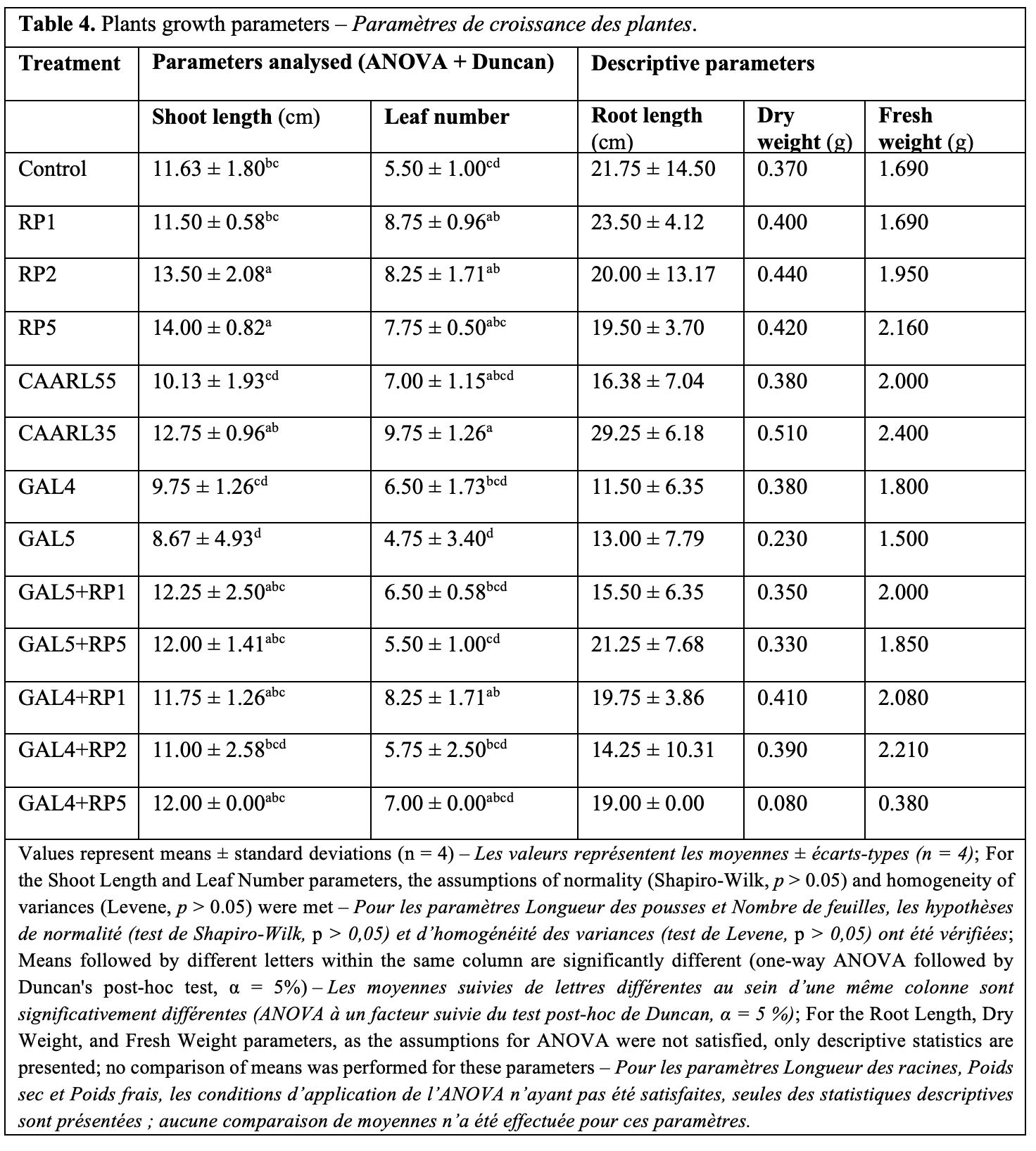
34Isolate CAARL35 induced the greatest mean root length (29.25 cm), although this value was not statistically different from the control due to high variability. Regarding root length, the GAL4 and GAL5 treatments, as well as the GAL5+RP1 combination, resulted in a significant reduction of this parameter.
35Contrasting effects were observed on plant growth parameters. Dry weight was significantly increased by several bacterial treatments, most notably Pseudomonas fluorescens isolates (RP1, RP2, RP5) and the Streptomyces sp. isolate CAARL35, as well as the combination GAL4+RP1. The CAARL35 treatment had the most pronounced effect, yielding a dry weight of 0.51 g and outperforming all other treatments. In contrast, GAL5 and GAL4+RP5 treatments drastically reduced dry weight, with GAL4+RP5 reaching a minimum of 0.08 g. A similar overall trend was noted for fresh weight.
36Regarding leaf development, the number of leaves was significantly enhanced by isolates RP1, RP5, and CAARL35, and by the combination GAL4+RP1. Consistent with its effect on biomass, the Streptomyces isolate CAARL35 again demonstrated the highest performance in promoting foliar growth.
3.8. Principal Component Analysis (PCA)
37PCA was employed to visually depict the variation in the growth characteristics and the positioning of individuals based on distinct isolates and their various combinations (Figure 8). The two axes account for 80.13% of the overall variation. The initial axis explains the largest proportion of variation, specifically 60.01%. It exhibits a negative correlation with growth indices such as aerial and root lengths, fresh weight, dried weight and leaf number. The second axis accounts for 20.12% of the information. It has a positive correlation with fresh and dry weight as well as a negative correlation with aerial and root lengths.
38The PCA analysis revealed that the Pseudomonas fluorescens RP1, RP2, RP5, and Streptomyces sp. CAARL35, along with the combination GAL4+RP1, exhibit high efficiency in promoting carob tree growth. These isolates can be considered promising candidates for developing a microbial inoculum to improve carob tree growth.
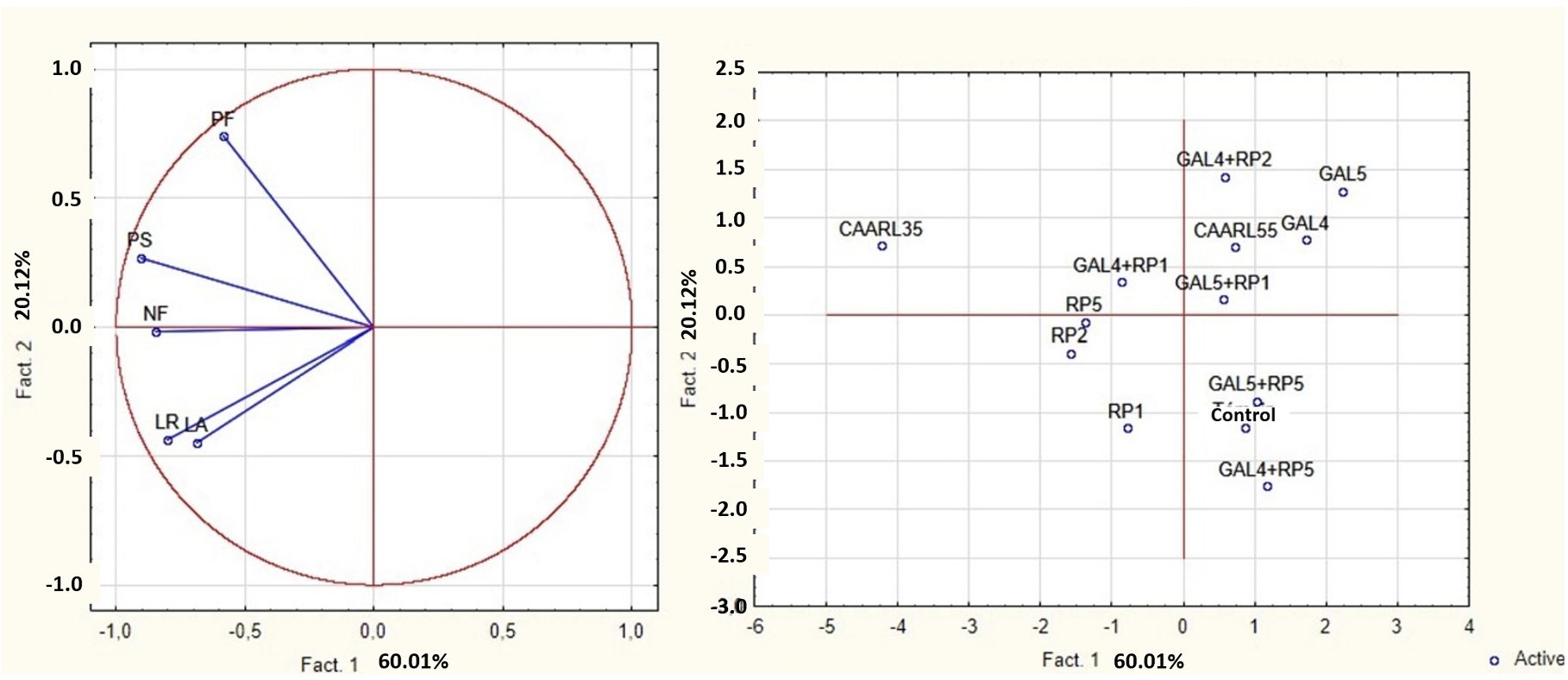
Figure 8. Principal Component Analysis (PCA) of the isolates effect on the growth parameters of the carob plants – Analyse en composantes principales (ACP) de l’effet des isolats sur les paramètres de croissance des plants de caroubier.
PF: fresh weight – poids frais ; PS: dry weight – poids sec ; NF: leaf number – nombre de foliole; LR: root length – longueur des racines ; LA: length of the aerial parts – longueur des parties aériennes.
4. DISCUSSION
39These results demonstrate a positive relationship between soil organic matter content and microbial abundance (based on culturable isolates). This finding can be explained by the fact that actinomycetes of the genus Streptomyces play a key role in degrading organic matter, thereby actively participating in the biogeochemical carbon cycle (Alvarez et al., 2017). According to our results, legumes improve both the biological fertility of the soil (by increasing organic matter) and its chemical fertility (by altering the dynamics of calcium carbonate and salts) (Kumar et al., 2020). Thanks to their root exudates, which are rich in organic acids, vitamins, and sugars, they attract a significant diversity of microorganisms to their rhizosphere, thereby creating a micro-environment favorable to microbial growth and interactions (Hu et al., 2018).
40The results of IAA production indicate that the top producers were isolated from the rhizosphere of different host plants (L. creticus and mixed R. monosperma + L. creticus). This suggests that the rhizosphere of legumes can be an excellent source for isolating IAA-producing bacteria, which aligns with the findings of Megu et al. (2024).
41Furthermore, this production varies for each bacterial type. It has been demonstrated that Pseudomonas fluorescens are massive producers of IAA (Laabas et al., 2017), and they improve root health, promoting proper plant growth.
42Similarly, production of IAA by Streptomyces spp. stimulates root growth and the formation of root hairs. This allows the plant to have better access to water and nutrients (Zhang et al., 2018).
43Regarding phosphorus, as it is a central pillar of energy metabolism (ATP), DNA/RNA synthesis, and membrane structure, improved phosphorus nutrition directly explains the observed increases in dry weight, fresh weight, and foliar development (Essahibi et al., 2019). This nutrient is of critical importance for carob trees (Ceratonia siliqua) (Batlle & Tous, 1997), which respond positively to phosphorus availability, especially during the initial growth phases (Correia & Martins-Loucao, 1997) which is in accordance with these results. A well-developed root system, established early on, allows the young tree to access more water and nutrients, ensuring better survival and faster growth in its first few years.
44It is important to note that phosphorus availability is highly dependent on soil pH, being most available in soils with a pH between 6.0 and 7.0 (Hinsinger, 2001; Shen et al., 2011). In the typically alkaline (high pH) calcareous soils of the Mediterranean region, such as the soil used in this experiment (30% total CaCO₃ and a pH of 9.38), phosphorus can become fixed (locked up) and unavailable to plants (Brady & Weil, 2016). This is why the use of phosphate-solubilizing bacteria, such as Pseudomonas fluorescens and Streptomyces sp. can be a sustainable and ecological solution for carob tree growth.
45It is noteworthy that several isolates with high IAA production, such as CAARL55, showed no detectable phosphate solubilization activity under the tested conditions. This functional specialization highlights the importance of consortium-based approaches, where different bacteria with complementary plant growth-promoting (PGP) traits can be combined to provide multiple benefits to the plant (Kumar & Dubey, 2020).
46Streptomyces species are among the most effective fungal antagonists found in soil ecosystems. Their long-term coexistence and interactions with soil fungi have driven the evolution of a wide range of antagonistic strategies, including the production of antibiotics and cellulolytic enzymes (Le et al., 2022). They also possess the ability to induce the plant's immune system, promoting healthy growth under various conditions (Khan et al., 2023). Consequently, Streptomyces sp. isolate CAARL35 can be a powerful biological solution to counter phytopathogenic fungi as described by Boukaya et al. (2018).
47The results on co-inoculations corroborate literature data reporting unpredictable effects (Saidi, 2022; Guigard, 2023). The GAL4+RP1 association proved beneficial, suggesting a synergistic interaction, while GAL4+RP5 was strongly inhibitory, which may reflect incompatibility or competitive interactions that is detrimental to the plant. These isolates could produce inhibitory compounds that, at certain concentrations, become phytotoxic or disrupt the beneficial root microbiome, explaining their negative effects (Yegorenkova et al., 2016).
48This finding demonstrates a clear observed dissociation: while length growth was only slightly affected, biomass accumulation was strongly impacted. This suggests that PGPR do not simply promote growth in a general sense but rather modulate plant physiological and metabolic processes. For instance, they may enhance resource-use efficiency or carbon partitioning, resulting in plants that are denser rather than simply taller, as described by Saadaoui (2023).
49Although the P. fluorescens isolates (RP1, RP2, RP5) were effective, they were consistently slightly less efficient than the Streptomyces sp. isolate CAARL35 across most measured parameters. CAARL35 appears to function as an exceptional "all-in-one" beneficial strain. In contrast, the P. fluorescens isolates may provide a more targeted metabolic "boost". Their primary mechanism likely involves relieving phosphorus limitation, thereby enabling the plant to fully express its inherent growth potential without causing major alterations to root architecture or shoot elongation.
5. CONCLUSIONS
50This study validates the potential of selected PGPR (Streptomyces spp. and Pseudomonas fluorescens) to enhance the early-stage growth of carob trees in alkaline, calcareous, and infertile soils, offering a strategy for their sustainable rehabilitation. While the results highlight the inherent complexity of these interactions, the CAARL35 strain and the synergistic GAL4+RP1 combination emerged as particularly promising treatments. Conversely, the inhibitory effects observed with GAL4+RP5 underscore that rhizosphere dynamics are intricate and that oversimplifying PGPR as mere growth stimulants is inaccurate. Future research should focus on characterizing the molecular mechanisms underlying these synergies and antagonisms to design rational and effective microbial consortia for sustainable agriculture. It is also important to note the issue of strain incompatibility, which underscores the need for rigorous selection protocols. It is therefore essential to evaluate strains not only for their individual efficacy but also for their compatibility within microbial consortia.
Bibliographie
Aeron A., Kumar S., Pandey P. & Maheshwari D.K., 2011. Emerging role of plant growth promoting rhizobacteria in agrobiology. In: Maheshwari D.K., ed. Bacteria in agrobiology: crop ecosystems. Berlin/Heidelberg, Germany: Springer, 1-36, doi.org/10.1007/978-3-642-18357-7_1
Agaras B.C. et al., 2015. Quantification of the potential biocontrol and direct plant growth promotion abilities based on multiple biological traits distinguish different groups of Pseudomonas spp. isolates. Biol. Control, 90, 173-186, doi.org/10.1016/j.biocontrol.2015.07.003
Agaras B.C. et al., 2020. Biocontrol potential index of Pseudomonas, instead of their direct-growth promotion traits, is a predictor of seed inoculation effect on crop productivity under field conditions. Biol. Control, 143, 104209, doi.org/10.1016/j.biocontrol.2020.104209
Ait Chitt M., Belmir H. & Lazrak A., 2007. Production de plants sélectionnés et greffés de caroubier. Transfert Technol. Agric., 153, 1-4.
Alvarez A. et al., 2017. Actinobacteria: current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere, 166, 41-62, doi.org/10.1016/j.chemosphere.2016.09.070
Azizi S. et al., 2022. Soil inoculation with beneficial microbes buffers negative drought effects on biomass, nutrients, and water relations of common myrtle. Front. Plant Sci., 13, 892826.
Backer R. et al., 2018. Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci., 9, 1473, doi.org/10.3389/fpls.2018.01473
Batlle I. & Tous J., 1997. Carob tree: Ceratonia siliqua L. Promoting the conservation and use of underutilized and neglected crops, 17. Seeland, Germany: Leibniz Institute of Plant Genetics and Crop Plant Research.
Boukaya N. et al., 2018. Biocontrol and plant-growth-promoting capacities of actinobacterial strains from the Algerian Sahara and characterisation of Streptosporangium becharense SG1 as a promising biocontrol agent. Biocontrol Sci. Technol., 28(9), 858-873, doi.org/10.1080/09583157.2018.1501466
Bouznada K. et al., 2017. Saccharothrix ghardaiensis sp. nov., an actinobacterium isolated from Saharan soil. Antonie van Leeuwenhoek, 110(3), 399-405, doi.org/10.1007/s10482-016-0812-4
Brady N.C. & Weil R.R., 2016. Phosphorus and sulfur. In: The nature and properties of soils. Upper Saddle River, NJ, USA: Pearson Press.
Brenner D.J., Krieg N.R. & Staley J.T., 2005. Bergey‘s manual of systematic bacteriology. Vol. 2. The proteobacteria. 2nd ed. Springer.
Bultreys A. & Gheysen I., 2000. Production and comparison of peptide siderophores from strains of distantly related pathovars of Pseudomonas syringae and Pseudomonas viridiflava LMG 2352. Appl. Environ. Microbiol., 66(1), 325-331, doi.org/10.1128/AEM.66.1.325-331.2000
Correia P.J. & Martins-Loucao M.A., 1997. Leaf nutrient variation in mature carob (Ceratonia siliqua) trees in response to irrigation and fertilization. Tree Physiol., 17(12), 813-819, doi.org/10.1093/treephys/17.12.813
Dalli Y., Yahia N. & Bekki A., 2020. Diversity of arbuscular mycorrhizal fungi associated with carob trees (Ceratonia siliqua L.) in western Algeria. Plant Cell Biotechnol. Mol. Biol., 21(17-18), 180-193.
De Barros Silva Leite M.C. et al., 2018. Bioprospection and genetic diversity of endophytic bacteria associated with cassava plant. Rev. Caatinga, 31(2), 315-325, doi.org/10.1590/1983-21252018v31n207rc
El Idrissi M.M. et al., 1996. Characterization of rhizobia isolated from carob tree (Ceratonia siliqua). J. Appl. Bacteriol., 80(2), 165-173.
El-Nakeeb M.A. & Lechevalier H.A., 1963. Selective isolation of aerobic actinomycetes. Appl. Microbiol., 11(2), 75-77, doi.org/10.1128/am.11.2.75-77.1963
Essahibi A. et al., 2019. Responsiveness of carob (Ceratonia siliqua L.) plants to arbuscular mycorrhizal symbiosis under different phosphate fertilization levels. J. Plant Growth Regul., 38(4), 1243-1254.
Gang S. et al., 2019. Analysis of Indole-3-Acetic Acid (IAA) production in Klebsiellaby LC-MS/MS and the Salkowski method. Bio-Protocol, 9(9), e3230, doi.org/10.21769/BioProtoc.3230
Goodfellow M. et al., 2012. Bergey’s manual of systemic bacteriology. Vol. 5. The actinobacteria. Springer.
Gordon S.A. & Weber R.P., 1951. Colorimetric estimation of indole acetic acid. Plant Physiol., 26(1), 192-195, doi.org/10.1104/pp.26.1.192
Gufwan L.A., Gufwan N.M., Abdulraheem M.I. & Jaafar A.A.K., 2025. Biocrust ecology and functionality for regenerative agriculture: a review of the opportunities and challenges facing microbial agroecosystem restoration. Arch. Agron. Soil Sci., 71(1), 1-28, doi.org/10.1080/03650340.2025.2508763
Guigard L., 2023. Réponses phénotypiques et moléculaires du riz au cours d’interactions avec des microorganismes du sol. Thèse de doctorat : Université de Montpellier (France).
Haddarah A., 2013. L’influence des cultivars sur les propriétés fonctionnelles de la caroube libanaise. Thèse de doctorat : Université Libanaise (Liban) et Université de Lorraine (France).
Hernandez A. & Dzakovich M., 2026. Making Hoagland's complete nutrient solution for hydroponics, protocols.io, doi.org/10.17504/protocols.io.36wgqpxp5vk5/v2
Hinsinger P., 2001. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil, 237(2), 173-195, doi.org/10.1023/A:1013351617532
Hu L. et al., 2018. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun., 9(1), 2738, doi.org/10.1038/s41467-018-05122-7
Hyder S. et al., 2024. Plant growth promoting rhizobacteria (PGPR): a green approach to manage soil-borne fungal pathogens and plant growth promotion. In: Kumar V. & Iram S., eds. Microbial technology for agro-ecosystems. Elsevier, 153-176, doi.org/10.1016/b978-0-443-18446-8.00014-0
Joffin J.N. & Leyral G., 2006. Microbiologie technique. Bordeaux, France : CRDP d’Aquitaine.
Jones K.L., 1949. Fresh isolates of actinomycetes in which the presence of sporogenous aerial mycelia is a fluctuating characteristic. J. Bacteriol., 57(2), 141-145, doi.org/10.1128/jb.57.2.141-145.1949
Khan H., Islam M.R., Zaman N.R. & Chowdhury F.T., 2021. Anti-fungal secondary metabolites and hydrolytic enzymes from rhizospheric bacteria in crop protection: a review. J. Bangladesh Acad. Sci., 44(2), 69-84, doi.org/10.3329/jbas.v44i2.51452
Khan S., Srivastava S., Karnwal A. & Malik T., 2023. Streptomyces as a promising biological control agents for plant pathogens. Front. Microbiol., 14, 1285543, doi.org/10.3389/fmicb.2023.1285543
Kumar A. & Dubey A., 2020. Rhizosphere microbiome: engineering bacterial competitiveness for enhancing crop production. J. Adv. Res., 24, 337-352, doi.org/10.1016/j.jare.2020.04.014
Kumar S. et al., 2020. Legumes for carbon and nitrogen cycling: an organic approach. In: Datta R., Meena R.S., Pathan S.I. & Ceccherini M.T., eds. Carbon and nitrogen cycling in soil. Singapore: Springer, 337-375, doi.org/10.1007/978-981-13-7264-3_10
Laabas S. et al., 2017. Impact of single and co-inoculations with rhizobial and PGPR isolates on chickpea (Cicer arietinum) in cereal-growing zone soil. J. Plant Nutr., 40(11), 1616-1626, doi.org/10.1080/01904167.2016.1270317
Lavudi H.N. et al., 2023. Plant growth promoting rhizobacteria (PGPR): an overview for sustainable agriculture and development. In: Parray J.A., Shameem N., Egamberdieva D. & Sayyed R.Z., eds. Rhizobiome. Elsevier, 95-125, doi.org/10.1016/b978-0-443-16030-1.00017-1
Le K.D. et al., 2022. Streptomyces sp. AN090126 as a biocontrol agent against bacterial and fungal plant diseases. Microorganisms, 10(4), 791, doi.org/10.3390/microorganisms10040791
Malik D.K. & Sindhu S.S., 2011. Production of indole acetic acid by Pseudomonas sp.: effect of coinoculation with Mesorhizobium sp. Cicer on nodulation and plant growth of chickpea (Cicer arietinum). Physiol. Mol. Biol. Plants, 17(1), 25-32, doi.org/10.1007/s12298-010-0041-7
Martins-Loucao M.A., Correia P.J. & Romano A., 2024. Carob: a Mediterranean resource for the future. Plants (Basel), 13(9), 1188, doi.org/10.3390/plants13091188
Meena M. et al., 2023. PGPR-mediated synthesis and alteration of different secondary metabolites during plant-microbe interactions. In: Swapnil P. et al., eds. Plant-microbe interaction - recent advances in molecular and biochemical approaches. Elsevier, 229-255, doi.org/10.1016/b978-0-323-91875-6.00002-5
Megu M., Paul A. & Deb C.R., 2024. Isolation and screening of stress tolerant and plant growth promoting root nodulating rhizobial bacteria from some wild legumes of Nagaland, India. S. Afr. J. Bot., 168, 260-269, doi.org/10.1016/j.sajb.2024.03.021
Mounir A.F., 2024. 24 Karat agricultural GEM - part 2. Agrotica, https://agrotica.debbaneagri.com/24-karat-agricultural-gem-pt2/, (27/05/26).
Ortiz P.L., Arista M. & Talavera S., 1995. Germination ecology of Ceratonia siliqua L. (Caesalpiniaceae), a Mediterranean tree. Flora, 190(1), 89-95, doi.org/10.1016/s0367-2530(17)30630-8
Parlascino R., Riolo M. & Cacciola S.O., 2024. First report of Alternaria alternata causing black spots in carob (Ceratonia siliqua) leaves in Europe. J. Plant Pathol., 107(1), 737, doi.org/10.1007/s42161-024-01741-w
Pereira S. et al., 2020. Plant growth-promoting rhizobacteria (PGPR) improve the growth and nutrient use efficiency in maize (Zea mays L.) under water deficit conditions. Heliyon, 6(10), e05106, doi.org/10.1016/j.heliyon.2020.e05106
Pikovskaya R.E., 1948. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya, 17, 362-370.
Pochon J. & Tardieux P., 1962. Technique d'analyse en microbiologie du sol. St-Mandé, France : Éditions de la Tourelle.
Saadaoui N., 2023. Impact stimulateur des PGPR sur la croissance végétale et sur les communautés bactériennes bénéfiques cultivables du microbiote racinaire du blé dur. Thèse de doctorat : Université Ferhat Abbas Sétif 1 (Algérie).
Saidi S., 2022. Amélioration de la symbiose Sinorhizobium meliloti–Medicago sativa après co-inoculation par des Actinobacteria. Thèse de doctorat : Université Ferhat Abbas Sétif 1 (Algérie).
Sebihi F.Z., 2016. Effet PGPR des souches de Pseudomonas fluorescens isolées de la rhizosphère du blé cultivé dans la région de Constantine. Thèse de doctorat : Université des Frères Mentouri-Constantine (Algérie).
Sharma D., Kaur T., Chadha B.S. & Manhas R.K., 2011. Antimicrobial activity of actinomycetes against multidrug resistant Staphylococcus aureus, E. coli and various other pathogens. Trop. J. Pharm. Res., 10, 801-808, doi.org/10.4314/tjpr.v10i6.14
Shen J. et al., 2011. Phosphorus dynamics: from soil to plant. Plant Physiol., 156(3), 997-1005, doi.org/10.1104/pp.111.175232
Steel K., 1961. The oxidase reaction as a taxonomic tool. Microbiology, 25(2), 297-306, doi.org/10.1099/00221287-25-2-297
Tabli N. et al., 2018. Plant growth promoting and inducible antifungal activities of irrigation well water-bacteria. Biol. Control, 117, 78-86, doi.org/10.1016/j.biocontrol.2017.10.010
Taylor W.I. & Achanzar D., 1972. Catalase test as an aid to the identification of Enterobacteriaceae. Appl. Microbiol., 24(1), 58-61, doi.org/10.1128/am.24.1.58-61.1972
Thibodeau P.O., Twagirayesa P. & Charrier F., 2002. Évaluation de 15 fongicides contre la moisissure chevelue (Rhizopus stolonifer et Mucor sp.) de la fraise et de la framboise. Agrosol, 13(1), 75-80.
Thilagam R. & Hemalatha N., 2019. Plant growth promotion and chilli anthracnose disease suppression ability of rhizosphere soil actinobacteria. J. Appl. Microbiol., 126(6), 1835-1849, doi.org/10.1111/jam.14259
Thomas P.A., Garcia‐Martí X., Mukassabi T.A. & Tous J., 2024. International biological flora: Ceratonia siliqua. J. Ecol., 112(8), 1885-1922, doi.org/10.1111/1365-2745.14325
Vacheron J. et al., 2013. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant. Sci., 4, article 356, doi.org/10.3389/fpls.2013.00356
Yegorenkova I.V. et al., 2016. Assessing the efficacy of co-inoculation of wheat seedlings with the associative bacteria Paenibacillus polymyxa 1465 and Azospirillum brasilense Sp245. Can. J. Microbiol., 62(3), 279-285, doi.org/10.1139/cjm-2015-0647
Zhang D.-J. et al., 2018. Auxin modulates root-hair growth through its signaling pathway in citrus. Sci. Hortic., 236, 73-78, doi.org/10.1016/j.scienta.2018.03.038
Zhang T. et al., 2024. Plant growth-promoting rhizobacteria (PGPR) improve the growth and quality of several crops. Heliyon, 10(10), e31553, doi.org/10.1016/j.heliyon.2024.e31553
