


- Startpagina tijdschrift
- volume 12 (2008)
- numéro 2
- Improvement of xylanase production by Penicillium canescens 10-10c in solid-state fermentation


Weergave(s): 3060 (19 ULiège)
Download(s): 114 (1 ULiège)

Improvement of xylanase production by Penicillium canescens 10-10c in solid-state fermentation

Nota's van de redactie
Received on May 2, 2006, accepted on September 10, 2007
Résumé
Optimisation de la production des xylanases par Penicillium canescens 10-10c en culture solide. L’intérêt que suscitent les hémicellulases, plus particulièrement les xylanases, est de plus en plus important, que ce soit d’un point de vue théorique ou d’un point de vue appliqué. Or, il est paradoxal de constater que le coût élevé de leur production limite leur utilisation à grande échelle. L’emploi de xylane purifié comme substrat de culture augmente le coût de production de l’enzyme. En conséquence, pour des applications commerciales, il convient de développer des procédés à partir de substrats peu coûteux. C’est dans ce cadre que se situe cette étude. Elle vise à optimiser la production de xylanases en milieu solide à partir de résidus agricoles. La souche envisagée est Penicillium canescens 10-10c, sélectionnée dans notre laboratoire pour son aptitude à produire une activité xylanase exempte d’activité cellulase. Les travaux concernent l’optimisation de différents paramètres de culture avec comme optique le développement ultérieur d’un réacteur semi-solide à base de tourteau de soja. Ces paramètres sont : la composition du milieu de culture, la température d’incubation, les phénomènes d’induction et de répression. Le tourteau de soja non broyé (taille > 10 mm) donne une bonne activité enzymatique. L’augmentation de la quantité de milieu de culture entraîne une baisse de la production enzymatique. La présence de lactose, de saccharose ou d’amidon de blé a un effet positif sur la production des xylanases tandis que la présence de xylose, de mannose, de galactose, d’arabinose, de cellobiose, de pectine ou de méthylcellulose réduit la production de xylanase. Les phosphates dipotassique ou disodique améliorent la production. La production enzymatique obtenue en fiole (250 ml) après une incubation de 7 jours à 30 °C est de l’ordre de 14 000 U.g-1 de substrat carboné. La nature de l’inoculum affecte la productivité enzymatique. En effet, les productivités enzymatiques respectives suivantes sont obtenues : 965 U.g-1 par jour avec une pré-culture solide, 473 U.g-1 par jour avec une pré-culture liquide et 383 U.g-1 par jour avec des spores. Les niveaux d’activités enzymatiques obtenus avec cette technique sont supérieurs aux niveaux présentés dans la littérature et démontrent donc toutes les potentialités de cette souche et de cette technique pour la production des xylanases.
Abstract
Among hemicellulases, xylanases are catalysts of considerable interest so as fundamental than applied point of view. However, it is paradoxical to note that the high cost of their production limits their use on a large scale. The use of purified xylan as culture substrate increases the production cost of the enzyme. Consequently, for commercial applications, it is advisable to develop processes starting from inexpensive substrates. The purpose of this study is to optimise xylanases production in solid-state fermentation based on agricultural residues. The strain is Penicillium canescens 10-10c, selected in our laboratory for his ability to produce xylanase activity free of cellulase. Assays concern optimization of different culture parameters in order to develop in the future a solid-state fermentation reactor with soya oil cake. These parameters are: medium composition, temperature incubation, induction and repression mechanisms. Soya oil cake in pellets (size > 10 mm) gave a higher enzymatic activity. Great volume of culture medium reduced the enzymatic production. The presence of lactose, saccharose or starch of corn has a positive effect on the production of xylanase while the presence of xylose, mannose, galactose, arabinose, cellobiose and pectin or methylcellulose reduces the production of xylanase. The sources of phosphorus (di-potassic and di-sodic) enhance xylanase production. The enzymatic production obtained in Erlenmeyer flasks (250 ml) after 7 days incubation at 30°C is about 14 000 U.g-1 of carbon source. The nature of inoculum affects the enzymatic productivity. Indeed, better productivity was obtained with inoculation by solid preculture (956 U.g-1 per day) than liquid preculture (473 U.g-1 per day) and spores suspension (383 U.g-1 per day). These observed enzymatic activity levels are higher than those related in the literature, which shows all the potentialities of this strain and this technique for the production of xylanase and allow to develop our solid-state fermentation bioreactor.
Inhoudstafel
1. Introduction
1Among hemicellulose, xylan is a heteregeneous polysaccharide in which β-1, 4-linked-D-xylopyranose residues are the main constituents. Depending on their origin, xylans may also contain variable amounts of arabinosyl and 4-O-methylglucuronic acid residues and acetyl groups. The most important enzyme in the xylan biodegradation is the endo-1, 4-β-xylanase (EC 3.2.1.8) that releases xylopyranose units. Xylanases offer considerable interest as catalysts in various biotechnological applications, such as bleaching of kraft pulps (Niku-Paavola et al., 1994; Vicuna et al., 1997; Madlala et al., 2001), clarifying juices and wines (Philippe, 1997), improving the nutritional value of animal feed (Buchert et al., 1994; Medel et al., 2002).
2A variety of micro-organisms, bacteria and filamentous fungi have been reported to produce xylanolytic enzymes (San-Lang et al., 2003; Henning et al., 2005; Virupakshi et al., 2005). Among these, Penicillium canescens 10-10c is able to produce xylanase free of cellulase activities (Gaspar, 1999). In submerged culture, xylanase production by this filamentous strain is optimized by reducing hydrodynamic stress and by increasing oxygen transfer (Yasser, 2003). The best production was obtained in medium containing soya meal and wheat straw and reached 535 U.ml-1 after 120 h (Gaspar, 1999).
3The process of enzyme production in solid-state fermentation was previously investigated with this strain (Yasser, 2003). The maximum of activity produced after 12 days at 30°C has reached 9 632 units.g-1 of carbon source.
4The purpose of this study was to improve this enzyme production by testing different culture conditions in order to enhance the productivity (in terms of units.g-1 substrate per day) on agricultural substrate available in quantities and easy to manipulate.
2. Material and methods
2.1. Strain
5P. canescens 10-10c was supplied by G.I Kvesidatse, Institute of Plant Biochemistry, Academy of Sciences, Tbilisi, Georgia.
6Enzyme production by using solid-state fermentation.
7Erlenmeyer flasks (250 ml) containing 5 g of substrate plus 20 ml of distilled water supplemented by casein peptone (0.75% W/V) were autoclaved, inoculated with a suspension of spores to reach 106 CFU.g-1 of carbon source. The flasks were then incubated at 30°C.
2.2. Enzyme extraction
8The enzyme from each flask was extracted with distilled water in a total volume of 100 ml by stirring during 1 h at room temperature. After filtration, the clear supernatant was used as the enzyme source.
2.3. Enzyme assay
9Xylanase activity was measured according to Bailey et al. (1992) using 1% birchwood xylan as substrate. Reducing sugars were assayed by dinitrosaycylic acid method with xylose as standard. One unit of xylanase activity is defined as the amount of enzyme that released 1 µmol of reducing sugar equivalent to xylose per minute.
2.4. Protein assay
10Protein was measured by bicinchoninic acid method (Sigma acid protein assay kit).
2.5. Cultivation in trays
11Enamel metallic trays of sizes 17*11*5 cm3 were used to cultivate the fungus strain after covering with aluminium foil and autoclaving. The wet Penicillium soya oil cakes were extracted from trays and assayed for xylanase.
12A three-stage cultivation technique was employed: spores suspension, liquid-state preculture, solid-state preculture. In the first, trays containing 50 g of soya oil cake plus 200 ml of distilled water containing 1,5 g of casein peptone and 2 g of Na2HPO4.2H2O were autoclaved, inoculated with a suspension of spores to reach 250.105 spores. In the second, 10 ml distilled water containing 300 mg of soya oil cake, 100 mg Na2HPO4.2H2O, 75 mg casein peptone were autoclaved, inoculated with 250.105 spores. Tubes containing the mixture were incubated at 30°C for 1 day in an orbital shaker (150 rpm) for mycelium production. In the third stage, an inoculum of 250.105 spores was added to the enzyme production medium (EPM) in Erlenmeyer flasks (250 ml) after autoclaving and incubation at 30°C for 1 day. The composition of EPM was 5 g soya oil cake (5 mm) plus 20 ml of distilled water containing casein peptone (0.75% W/V).
13Then, the 1 day old preculture was used to inoculate the trays containing soya oil cake (49,70 g in the second and 45 g in the third stage). The carbon sources, supplemented by the mineral solution prior to heat sterilisation (121°C, 20 min) were prepared as follows:
14– in the second stage: 190 ml of distilled water plus 1.425 g of casein peptone, and 1.9 g of Na2HPO4.2H2O.
15– in the third stage: 180 ml of distilled water plus 1.35 g of casein peptone and 1.8 g of Na2HPO4.2H2O.
3. Results and discussion
3.1. Enzyme production on various carbonaceous substrates
16Penicillium strain was grown on medium containing casein peptone (0.75% W/V), 20 ml of distilled water and 5 g of soya oil cake alone or soya oil cake supplemented with various substrate (soya meal, wheat bran, pulp beet) at two ratios (70 : 30, 50 : 50 respectively). Table 1 shows that the soya oil cake at 100% yielded the highest xylanase production. The maximum activity after 7 days reached 12 000 units.g-1 of soya oil cake. The combination of soya oil cake and another agricultural waste gradually decreased the levels of xylanase production. The variations of the production level observed with various lignocellulosic materials are probably related on differences in composition and the accessibility of the substrates (Yasser, 2003). The soya oil cake could constitute a significant source of nitrogen. Thus, it is also probable that the effect of these two sources of nitrogen, soya bean oil cake and yeast extract, on the xylanasic production, is related to the evolution of the neutral pH that they induce (Gaspar, 1999).
17With wheat straw, xylanase production by P. canescens 10-10c attained 9 632 units.g-1 of substrate after 12 days of culture (Yasser, 2003; Yasser et al., 2003).
3.2. Effect of carbon source size
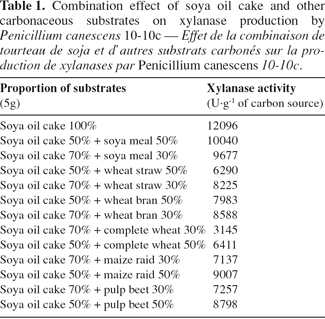
18The particles size of the substrate had an influence on the ratio surface/volume (S/V) and thus on the accessibility of the substrate for the microorganisms, on the possibilities of diffusion of the oxygen, heat, and exo-enzymes. The availability of oxygen is determined by diffusion, which in turn is determined by porosity, particle size and consistency of the substrate (Alvarez-Martinez, 1987; Mitchell et al., 1988). It is clear that particle size affected enzyme production. Smaller particle sizes of wheat straw < 0.5 mm having greater surface to volume ratio (Weiland, 1988) gave the higher xylanase production by Thermoascus aurantiacus in solid-state culture (Kalogeris et al., 1998). In our study, two sizes of soya oil cake were tested. Production of enzyme was carried out on pellets of soya oil cake (size >10 mm) and on pellets crushed in order to obtain after sieving particles of 1 mm. Figure 1 shows that the activity produced by P. canescens 10-10c is more important with the soya oil cake remained in pellets. The enzymatic level of production is higher 26% (14 485 U.g-1) than with the crushed soya oil cake (11 496 U.g-1). Perhaps this negative effect is a result of a reduction of aeration in the medium by settling. These findings confirm relevant data before reported (Gaspar, 1999) on xylanase production by P. canescens 10-10c with micronisation of soya meal.
3.3. Effect of quantity of medium
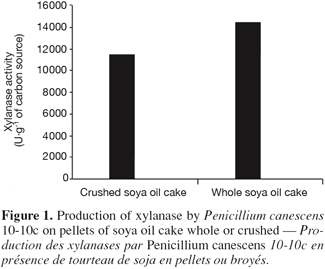
19For the static cultures, an adequate thickness of medium is necessary to allow a good ventilation of the mass in order to maintain aerobic conditions favourable to the cell multiplication and the synthesis of the metabolites (Durand et al., 1989). Generally in solid fermentation, the optimal thicknesses of culture are low. Indeed, great volumes or thicknesses of culture release significant heat in the culture medium. That constitutes difficulties for the large-scale productions. In the tempe production, similar observations have been reported concerning metabolic heat accumulation when increasing the fermentor size (Aidoo et al., 1982). Consequently, the temperature within a bed or package of tempe may rise 10-16°C above that of environment. A steep temperature gradient of 3°C.cm-1 bed thickness during active growth has been reported in a fermentor employing a bed height of 6.5 cm (Rathbun et al., 1983). Production of enzyme was measured by loading the medium in flasks in order to multiplicate it by 2, 3, etc. and these media were respectively named 2X, 4X, etc. The contents of the flasks were extracted and assayed from each set. The highest xylanase production was observed when the bed height of the medium is the lower (Figure 2). The same result is observed during the production of penicillin in solid-state fermentation by Penicillium chrysogenum (Barrios-Gonzalez et al., 1988). The optimal production of penicillin is obtained with increasing the culture medium concentration only by two. Then increasing medium concentration had an adverse effect on penicillin production by P. chrysogenum (Barrios-Gonzalez et al., 1988). Increasing of medium concentration causes a bad aeration and a bad heat transfer in the mass, and then reduces enzymatic productivity. Thus, with a concentration of the medium of 10X (the height of bed is 30 cm), important putrefaction releases itself from the flasks at the end of the 7 days of incubation. This situation causes a weak development of the strain. The culture is almost impossible under these conditions.
3.4. Effect of temperature
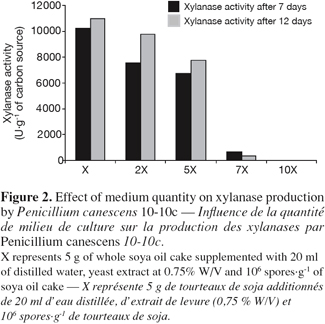
20Variations of a few degrees around the optimal temperature can notably modify the growth and the metabolism of the microorganism. Indeed, the solid substrates, because of their heterogeneity, their nature and their low moisture, involve a bad evacuation of calories (Saucedo-Castaneda et al., 1990; Prior et al., 1992). Great quantities of calories can accumulate in the medium, generating micro gradients, which progressively form more significant gradients. It is thus significant to be able to optimize precisely the temperature of culture. The influence of temperature incubation on the enzymatic production, cellular viability, level of extracellular proteins and lost of fresh weight (initial weight / final weight) were studied by following kinetic evolution of these parameters during fermentation process respectively at 23°C, 30°C and 37°C. Figures 3 and 4 indicate respectively a fall of the enzymatic production, level of extracellular proteins and cellular viability at 23°C and at 37°C. At 37°C, figure 5 indicates also great losses of fresh weight in flasks. The losses would be then mainly constituted by water evaporation. The optimum temperature cultivation for xylanase production by P. canescens 10-10c in solid-state fermentation is 30°C. That temperature allows obtaining a good cellular viability, best enzymatic production and extracellular protein synthesis, low loss of fresh weight. An adequate temperature control was an important factor to reach these productivities in reactor scale.
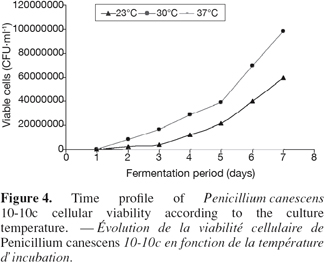
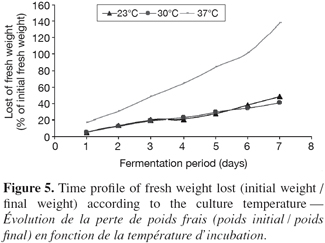
3.5. Effect of different salts
21Mineral salts are usually used in enzymatic production (San-Lang et al., 2003; Yasser, 2003; Yasser et al., 2003; Henning et al., 2005; Virupakshi et al., 2005). Experience was made with incorporating in the 20 ml of water used to moisten the substrate respectively:
22– at a concentration of 0.75% (W/V), different salts (NaCl, KCl, MgSO4, FeSO4, MnCl2, ZnSO4, CaCl2, and CuSO4).
23– at a concentration of 1% (W/V), different sources of inorganics phosphorus (K2HPO4, KH2PO4, Na2HPO4.2H2O, NaH2PO4.2H2O).
24Among them added at 0.75% (W/V) concentration, NaCl and KCl did not have effect on xylanase production (Table 2) whereas the other salts are negative for enzyme excretion: MgSO4, FeSO4, MnCl2, ZnSO4, CaCl2, and CuSO4. Phosphorus (mono-potassic and mono-sodic) induces a fall of enzyme production whereas sources of phosphorus di-potassic and di-sodic enhance xylanase production. This would be due partly to the modification of the initial pH induced by these ions. Sources of phosphorus mono-potassic and mono-sodic involve a fall of initial pH and sources of phosphorus di-potassic and di-sodic at 1% (W/V) increase initial pH towards neutrality. Indeed, our studies have shown us that in solid-state fermentation, initial neutral pH is very favourable to the xylanase synthesis by P. canescens 10-10c. An initial pH different of neutrality involves a fall of the enzymatic production. These results confirm relevant data (Gaspar, 1999; Yasser, 2003).
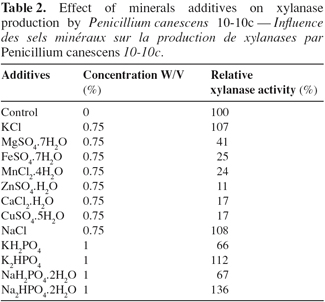
3.6. Effect of carbon sources as additive
25Soluble carbonaceous substrates have already been tested in order to evaluate their effects on xylanases production by other strains (Leathers et al., 1986; Priem et al., 1991; Smith et al., 1991; Haltrich et al., 1996; Abdel-Sater et al., 2001; Souza et al., 2001). Xylose, mannose, galactose, arabinose, lactose, cellobiose, saccharose, wheat starch, methylcellulose and pectine were used at a concentration of 5% (W/W) to supplement soya oil cake in order to evaluate their effect on xylanase production by P. canescens 10-10c in solid-state fermentation. The enzyme was assayed after 7 days incubation at 30°C. Among carbon sources assayed, mono- (xylose, mannose, galactose, arabinose), di- (cellobiose) and polysaccharides (methylcellulose and pectin) reduce xylanase production (Table 3). That can be explained by a catabolic repression of various substrates. On the other hand, lactose, saccharose and wheat starch increased enzyme production by P. canescens 10-10c.
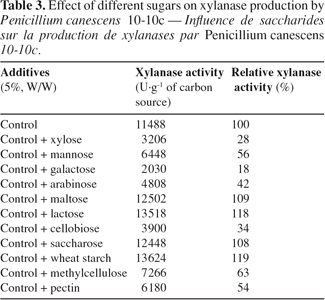
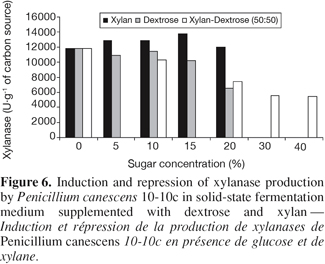
3.7. Effect of addition of dextrose and xylane on xylanase production in solid-state fermentation
26Several works highlighted the respective inducer or repressive character of xylan and dextrose on the production of xylanases (Espinar et al., 1992; Espinar et al., 1994; Singh et al., 1995; Yasser, 2003; Yasser et al., 2003).
27However, this observation depends in certain cases on culture conditions (Archana et al., 1997; Souza et al., 2001). To evaluate these effects, different levels of xylane (5, 10, 15, and 20%), dextrose (5, 10, 15, and 20%) and xylane with dextrose (5, 10, 15, and 20% for each) were incorporated into soya oil cake.
28Our results (Figure 6) confirm these observations. Indeed, we observed that the addition of xylan to soya oil cake incresead slighthy xylanase production. Previously, Yasser et al. (Yasser, 2003; Yasser et al., 2003) have showed similar observations for the same strain cultivated on wheat bran. On the other hand, xylanase production was repressed by dextrose. Addition of dextrose to the medium containing xylan produced also catabolic repression. Negative effect in this case is more important. Enzyme production was reduced of 50%. Perhaps, is it possible that accumulated xylose issued from the hydrolyse of xylan leaded to this important repression (Senior et al., 1989). But no complete repression was observed even at 20% dextrose.
3.8. Culture on trays
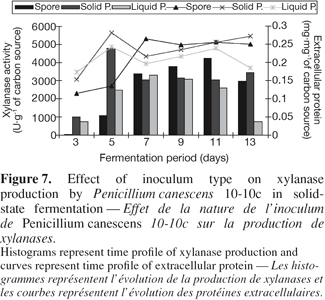
29Alternative ways of inoculation such as spore suspension, mycelium suspension obtained by liquid or solid precultures of spores were compared. The comparison (Figure 7) showed the interest of spores pregermination in solid medium, where phase of latency is reduced and where we quickly obtain optimum production of xylanase. Optimum productivity obtained are 965 U.g-1 per day with inoculation by solid preculture, 473 U.g-1 per day with inoculation by liquid preculture and 383 U.g-1 per day with spores inoculation. In xylanase production by solid-state fermentation with Thermoascus aurantiacus (Kalogeris et al., 1998), better results were obtained with liquid preculture (4 942 U.g-1) than spores suspension (4 714 U.g-1) and solid state culture (4 000 U.g-1) of wheat straw. However, in penicillin production by P. chrysogenum NRRL 1951, medium inoculation by spores gave better results than inoculation with mycelium obtained by preculture of spores (Barrios-Gonzalez et al., 1988).
4. Conclusion
30The hyperproductor character of xylanase by P. canescens 10-10c was already proven by other work (Gaspar, 1999; Yasser, 2003; Yasser et al., 2003). The present study confirms this observation. Indeed, the maximal enzymatic production obtained after 7 days incubation is 14 485 U.g-1 of substrate in Erlenmeyer. This level is higher than levels presented in the literature and shows all the potentialities of this strain and this technique for the production of xylanases. For commercial applications, the use of agricultural by-products for the production of these enzymes to replace purified xylan makes it possible to reduce the cost of the enzyme considerably. In addition, cultivation conditions (composition of the medium, cultivation temperature, induction and repression mechanisms, nature of inoculum) are essential to get good results.
Bibliographie
Abdel-Sater M.A. & El-Said A.H.M., 2001. Xylan-decomposing fungi and xylanolytic activity in agricultural and industrial wastes. Int. Biodeterior. Biodegradation, 47, 15-21.
Aidoo K.E., Hendry R. & Wood B.J.B., 1982. Solid substrate fermentations. Adv. Appl. Microbiol., 28, 201-237.
Alvarez-Martinez L.R., 1987. Modelling fungal Rhizopus oligosporus growth on extruded corn by solid substrate fermentation. Thesis: Colorado State University, Fort Collins (USA).
Archana A. & Satyanarayana T., 1997. Xylanase production by Thermophilic Bacillus Licheniformis A99 in solid-state fermentation. Enzyme Microb. Technol., 21, 12-17.
Bailey M.J., Biely P. & Poutanen K., 1992. Inter-laboratory testing of methods for assay of xylanase activity. J. Biotechnol., 23, 257-270.
Barrios-Gonzalez J., Tomasini A. & Viniegra G.L.G., 1988. Penicillin production by solid-state fermentation. In: Solid-state fermentation in bioconversion of agro-industrial raw materials. Proceedings of the seminar ORSTOM-MONTPELLIER (France), Montpellier, 25-27 July 1988. Montpellier, France: ORSTOM, 39-51.
Buchert J., Tenkanen M., Kantelinen A. & Viikari L., 1994. Application of xylanases in the pulp and paper industry. Bioresource Technol., 50, 65-72.
Durand A. & Chereau D., 1989. A new pilot reactor for solid-state fermentation: application to the protein enrichment of sugar beet pulp. Biotechnol. Bioeng., 31(5), 476-486.
Espinar M.T.F., Ramon D., Pinaga F. & Valles S., 1992. Xylanase production by Aspergillus nidulans. FEMS Microbiol. Lett., 9, 91-96.
Espinar M.T.F., Pena J., Pinaga F. & Valles S., 1994. α-L-arabinofuranosidase production by Aspergillus nidulans. FEMS Microbiol. Lett., 115, 107-112.
Gaspar A., 1999. Etude des interactions entre les paramètres biologiques et les grandeurs physiques liées à l’agitation et à l’aération lors de la production de xylanases par Penicillium canescens. Thèse de doctorat : Faculté Universitaire des Sciences Agronomiques de Gembloux (Belgique).
Haltrich D. et al., 1996. Production of fungal xylanases. Bioresource Technol., 58, 137-161.
Henning J., Astrid M., Kristain B.R.K. & Lisbeth O., 2005. Production of cellulases and hemicellulases by three penicillium species: effect of substrate and evaluation of cellulase adsorption by capillary electrophoresis. Enzyme Microb. Technol., 36, 42-48.
Kalogeris E., Christakopoulos P. & Kekos Macris B.J., 1998. Studies on the solid-state production of thermostable endoxylanases from Thermoascus aurantiacus: characterization of two isozymes. J. Biotechnol., 60, 155-163.
Leathers T.D., Detroy R.W. & Bothast R.J., 1986. Induction and glucose repression of xylanase from a color variant strain of Aureobasidium pullulans. Biotechnol. Lett., 8, 867-872.
Madlala A.M., Bissoon S., Singh S. & Christov L., 2001. Xylanase-induced reduction of chlorine dioxide consumption during elemental chlorine-free bleaching of different pulp types. Biotechnol. Lett., 23, 345-351.
Medel P. et al., 2002. Processing of barley and enzyme supplementation in diets for young pigs. Anim. Feed Sci. Technol., 95, 113-122.
Mitchell D.A., Greenfield P.F. & Doelle H.W., 1988. Development of a model solid-state fermentation system. Biotechnol. Tech., 2(1), 1-6.
Niku-Paavola M.L., Ranua M., Suurnakki A. & Kantelinen A., 1994. Effects of lignin modifying enzymes on pine kraft pulp. Bioresource Technol., 50, 73-77.
Philippe S., 1997. Xylan degradation by the anaerobic bacterium Bacteroides xylanolyticus. Thesis: Wageningen Agricultural University (Netherlands).
Priem B., Dobberstein J. & Emeis C.C., 1991. Production of β-1,4-xylanase in continuous culture by Aureobasidium pullulans CBS 58475. Biotechnol. Lett., 13, 149-154.
Prior B.A., Du Preez J.C. & Rein P.W., 1992. Environmental parameters. In: Solid-substrate cultivation. London; New York (USA): Elsevier Applied Science, 65-86.
Rathbun B.L. & Shuler M.L., 1983. Heat and mass transfer effects in static solid substrate fermentation: design of fermentations chambers. Biotechnol. Bioeng., 25, 929-938.
San-Lang W. et al., 2003. Production of xylanases from rice bran by Streptomyces actuosus A-151. Enzyme Microb. Technol., 33, 917-925.
Saucedo-Castaneda G. et al., 1990. Heat transfer simulation in solid substrate fermentation. Biotechnol. Bioeng., 35, 802-808.
Senior D.J., Mayers P.R. & Saddler J.N., 1989. Xylanase production by Trichoderma harzianum E58. Appl. Microbiol. Biotechnol., 32, 137-142.
Singh A., Kuhard R.C. & Kumar M., 1995. Xylanase production by a hyperxylanolytic mutant of Fusarium oxysporum. Enzyme Microb. Technol., 17, 551-553.
Smith D.C. & Wood T.M., 1991. Xylanase production by Aspergillus awamori. Development of a medium and optimization of the fermentation parameters for the production of extracellular xylanase and β-xylosidase while maintaining low protesase production. Biotechnol. Bioeng., 38, 883-890.
Souza D.F., Souza C.G.M. & Peralta R.M., 2001. Effect of easily metabolizable sugars in the production of xylanase by Aspergillus tamarii in solid-state fermentation. Process Biochem., 36, 835-838.
Vicuna R., Escobar F., Osses M. & Jara A., 1997. Bleaching of eucalyptus kraft pulp with commercial xylanases. Biotechnol. Lett., 19, 575-578.
Virupakshi S., Gireesh B.K., Satish R.G. & Naik G.R., 2005. Production of xylanolytic enzyme by a thermoalkaliphilic Bacillus sp. JB-99 in solid-state fermentation. Process Biochem., 40, 431-435.
Weiland P., 1988. Principles of solid-state fermentation. In: Zadrazil F. & Reiniger P., eds. Treatment of lignocellulosics with white rot fungi. London: Elsevier, 64-76.
Yasser B., 2003. Optimisation de la production de xylanases par Penicillium canescens 10-10c. Thèse de doctorat : Faculté Universitaire des Sciences Agronomiques de Gembloux (Belgique).
Yasser B., Philippe J. & Thonart P., 2003. Xylanase production by Penicillium canescens 10-10c in solid-state fermentation. Appl. Biochem. Biotechnol., 108, 737-748.
Om dit artikel te citeren:
Over : Allah Antoine Assamoi
Gembloux Agricultural University – FUSAGx. Centre wallon de Biologie industrielle. Unité de Bio-industries. Passage des Déportés, 2. B-5030 Gembloux (Belgium). E-mail: bioindus@fsagx.ac.be
Over : Franck Delvigne
Gembloux Agricultural University – FUSAGx. Centre wallon de Biologie industrielle. Unité de Bio-industries. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Over : Jean-Marc Aldric
Gembloux Agricultural University – FUSAGx. Centre wallon de Biologie industrielle. Unité de Bio-industries. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Over : Jacqueline Destain
Gembloux Agricultural University – FUSAGx. Centre wallon de Biologie industrielle. Unité de Bio-industries. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Over : Philippe Thonart
Gembloux Agricultural University – FUSAGx. Centre wallon de Biologie industrielle. Unité de Bio-industries. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
