


- Accueil
- volume 13 (2009)
- numéro 2
- Mutualisme pucerons – fourmis : étude des bénéfices retirés par les colonies d'Aphis fabae en milieu extérieur


Visualisation(s): 8335 (69 ULiège)
Téléchargement(s): 304 (6 ULiège)

Mutualisme pucerons – fourmis : étude des bénéfices retirés par les colonies d'Aphis fabae en milieu extérieur

Notes de la rédaction
Reçu le 18 septembre 2007, accepté le 6 mai 2008
Résumé
La relation de coopération entre pucerons – fourmis est un bel exemple de mutualisme dans le règne animal, les premiers cherchant protection et hygiène, les seconds une source de sucres nécessaires à la survie de la colonie. La présente étude s'est intéressée à recenser les bénéfices retirés par Aphis fabae Scopoli (Homoptera, Aphididae) de ses relations de mutualisme avec Lasius niger L. (Hymenoptera, Formicidae). Plusieurs paramètres ont été observés en milieu extérieur sur des plants de fèves des marais infestés initialement par 100 individus en présence ou non d'une colonie de L. niger. En présence de fourmis, les plantes étaient constamment infestées par un nombre de pucerons plus important et la proportion d'individus ailés y était également similaire ou plus grande, selon la date d'observation. Un nombre moins important de prédateurs aphidiphages sur les plantes en présence de fourmis a permis d'expliquer en partie ces observations. Les nombres moyens de pucerons parasités ne différaient pas, que les plantes soient explorées ou non par les fourmis, suggérant que L. niger est peu efficace face aux attaques de parasitoïdes. Par contre, très peu de pucerons appartenant à des espèces différentes d'A. fabae ont été observés sur les plants mis en présence des fourmis. Ces observations suggèrent que L. niger adopte un comportement de prédation sur les pucerons avec lesquels elle n'entretient aucune relation mutualiste. Les observations menées n'ont pas permis de mettre en évidence un quelconque effet des fourmis sur la vigueur des plantes hôtes des pucerons, bien que sensiblement moins d'exuvies et de taches de miellat étaient présentes sur les plantes dont les colonies de pucerons étaient visitées par L. niger. L'ensemble de ces résultats confirme que L. niger améliore les conditions de vie des colonies de pucerons d'A. fabae dont elle exploite le miellat, principalement grâce à la protection qu'elle apporte contre les prédateurs et la réduction de la pression de compétition exercée par les autres espèces non myrmécophiles de pucerons.
Abstract
Aphid – ant mutualism: an outdoor study of the benefits for Aphis fabae. Aphid – ant relationships are common examples of mutualism. Aphids are indeed submitted to predation and therefore require protection, while ants are continuously looking for new sugar sources. The present work aimed to study the benefits that a mutualistic relationship with Lasius niger L. (Hymenoptera, Formicidae) could bring to the black bean aphid Aphis fabae Scopoli (Homoptera, Aphididae). Several parameters were observed in the field, on broad bean plants infested with an initial amount of 100 A. fabae and in presence or not of a L. niger colony. More aphids were observed on plants being visited by ants as well as a higher proportion of winged individuals. One explanation is that fewer predators were observed on plants being visited by ants, demonstrating their protective role. However, the number of parasitized aphids was not reduced in presence of L. niger. On the other hand, fewer different aphid species were present on plants foraged by ants, what suggests that they could exert a predation on unattended aphids. Our observations do not allow to conclude on any impact of L. niger on the fitness of the aphid host plant, although fewer exuvia and honeydew spots were observed when they were present. All these results confirm that L. niger increase the fitness of A. fabae colonies mainly by decreasing the number of predators and by reducing competition from aphid species unattended by ants.
Table des matières
1. Introduction
1Les interactions entre espèces sont généralement classées par les spécialistes en catégories telles que la compétition, la prédation, le parasitisme et le mutualisme (Cushman et al., 1991). Les termes servant à parler de mutualisme sont divers : on parle de symbiose, de commensalisme, de coopération, d'aide mutuelle, de facilitation, d'altruisme réciproque et d'entraide (Boucher et al., 1982). Le mutualisme peut être défini comme une association à bénéfices réciproques pour deux organismes (Boucher et al., 1982). La mise en place d'une telle relation dépend des avantages que chacun peut apporter à l'autre, mais aussi des couts que le mutualisme entraine (Way, 1963). Des études théoriques suggèrent que le mutualisme s'applique uniquement à des situations où le cout de maintien de ce système est faible pour chacun des participants et où les bénéfices sont relativement importants (Bristow, 1991).
2La coopération entre les groupes des pucerons et des fourmis est un exemple bien connu de mutualisme puisqu'il s'apparente à la relation homme – bétail : un apport nutritionnel est échangé contre protection et entretien (Dixon, 1985 ; Stadler et al., 2005). En Europe, on estime qu'un tiers des espèces d'aphidés ne sont pas soignées par les fourmis, un tiers a une relation de mutualisme facultatif et un tiers a un mutualisme obligatoire (Stadler et al., 1998). La balance entre couts et bénéfices a fait pencher maintes fois la relation dans l'une ou l'autre direction et la diversité actuelle des espèces dans ces groupes et de leurs relations illustre bien la multitude des facteurs qui entrent en compte (Stadler et al., 2005).
3Alors que les fourmis y trouvent une source de sucres nécessaires à la survie de leur colonie, les pucerons retirent de nombreux bénéfices d'une telle relation et ont ainsi tout intérêt à la faire perdurer (Banks, 1962 ; Dixon, 1985). C'est ainsi qu'en présence de l'autre, chaque espèce manifeste des changements de comportement, voire de morphologie (Buckley, 1987). Certaines espèces de pucerons augmentent la quantité de phloème ingérée et adaptent alors la quantité et la qualité de leur miellat afin de satisfaire les demandes des fourmis (Völkl et al., 1999 ; Fischer et al., 2001 ; Yao et al., 2001). Suite à la palpation que les fourmis pratiquent avec leurs antennes sur le corps des pucerons, ceux-ci excrètent des gouttes de miellat qu'ils évitent alors d'éjecter, afin de faciliter leur récolte par les fourmis (Sudd, 1967). En échange, les fourmis changent leur comportement initial de prédateurs pour devenir éleveuses de pucerons (Huber, 1810) et tendre à améliorer leur santé et leur durée de vie. Ainsi, elles les protègent contre leurs nombreux prédateurs (Pontin, 1959 ; Yao et al., 2000) parmi lesquels les syrphes ou les coccinelles (Verheggen et al., 2007 ; 2008) et participent activement à l’hygiène de la colonie (Way, 1963).
4Les fourmis augmentent généralement la durée de vie des colonies de pucerons qu'elles soignent (Bristow, 1984 ; Mahdi et al., 1993 ; Sloggett et al., 2000). Cependant, les études dévouées à la mise en évidence des bénéfices retirés par les colonies de pucerons souffrent d'être réalisées en conditions de laboratoire. L'objectif de la présente étude est d'évaluer en milieu extérieur l'impact de la présence de Lasius niger L. (Hymenoptera, Formicidae) sur le bien-être de colonies d'Aphis fabae Scopoli 1763 (Homoptera, Aphididae), par le biais d'observations de terrain en conditions semi-contrôlées.
2. Matériel et méthodes
2.1. Matériel biologique
5Les plants de fève des marais (Vicia faba L.) proviennent de semences mises à germer dans des pots de 9 x 9 x 10 cm à raison de 8 graines par pot. Après germination, les plantes sont repiquées individuellement dans des pots de taille identique. Le substrat est composé de perlite et de vermiculite dans le rapport 1:1, permettant à la fois l'aération du milieu et la conservation de l'humidité apportée par un arrosage pratiqué trois fois par semaine. L'élevage des pucerons (A. fabae) est pratiqué par infestation naturelle à partir d'une plante âgée infestée vers des plantes saines disposées dans la même cage. La culture des fèves et l'élevage des pucerons sont réalisés dans des pièces séparées et climatisées, à une température de 23 ± 3 °C et une humidité de 70 ± 5 %.
6La récolte de la fourmilière de L. niger a été réalisée sur le Campus de la Plaine de l'ULB à Bruxelles le 18 avril 2007 par temps ensoleillé. Une motte de terre de 20 cm de diamètre environ avec herbe et racines est prélevée et désagrégée pour découvrir certaines chambres de la fourmilière. Les fourmis et la terre sont placées dans un bac dont les bords sont fluonés (polytetrafluoroéthylène) afin d'empêcher toute fuite des ouvrières. L'excavation est ensuite examinée afin de localiser les chambres de la fourmilière où est logé le couvain, qui est alors placé dans le même bac. Afin de conserver la fourmilière jusqu'au début de l'expérimentation, des tubes d'eau et de sucre sont ajoutés pour compléter les besoins nutritionnels. L'apport en protéines est constitué d'insectes frais ou congelés : deux fois par semaine, drosophiles, blattes ou vers de farine sont déposés dans le bac.
2.2. Dispositif expérimental
7Sur un terrain fraichement labouré, un traitement herbicide a été pratiqué 15 jours avant le début de l'expérimentation. Seize plants de V. faba sont disposés en deux cercles concentriques, à 0,5 et 1,50 m de rayon, à raison de 8 plants sur chaque cercle (Figure 1). Un dispositif de 16 plants est mis en présence d'une fourmilière de L. niger, un autre dispositif de 16 plants est placé en l'absence de fourmilière en son centre. Les plantes sont placées dans des pots de 13 x 13 x 13 cm contenant 3 à 4 cm de billes d'argex dans le fond et de la terre prise dans la parcelle expérimentale. Chaque pot placé autour de la fourmilière de L. niger est disposé dans un bac en plastique. Celui-ci est rempli de cailloux et de billes d'argex afin de permettre le passage facile des fourmis du sol vers les pots. L'infestation par A. fabae est réalisée manuellement sur des plantes âgées d'une semaine. Sur chaque plante, 100 adultes aptères ont été introduits au début de l'expérience. Les pots témoins sont placés dans des bacs similaires remplis d'eau pour éviter le passage de fourmis vers les plantes.
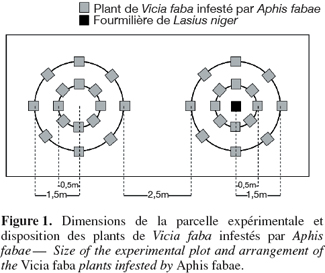
2.3. Observations réalisées
8Afin de mettre en évidence l'effet de la présence de fourmis sur l'hygiène et le développement de colonies d'A. fabae, différentes observations (présentées ci-dessous) ont été réalisées sur les acteurs biologiques suivants :
9– A. fabae,
10– les espèces d'autres pucerons,
11– les prédateurs et parasitoïdes présents,
12– les plants de V. faba,
13– les fourmis elles-mêmes.
14Les observations ont été réalisées une fois par semaine du 9 mai au 20 juin 2007. Les hauteurs des plants de V. faba ont été mesurées et une évaluation de leur vigueur a été établie sur base d'observations visuelles portant sur la présence de rouilles, nécroses, chloroses et autres dégâts dus à la présence de phytophages. Les colonies d'A. fabae ont été caractérisées sur base :
15– du nombre de pucerons,
16– de la proportion de pucerons ailés,
17– de la proportion de pucerons momifiés,
18– de la présence d'autres espèces d'aphidés.
19Les prédateurs et parasitoïdes présents sur les plants ont également été comptés et identifiés. Enfin, le nombre de fourmis présentes sur la plante et se nourrissant des nectaires extrafloraux a également été noté.
2.4. Analyse statistique
20L'analyse des données a été réalisée à l'aide du logiciel MINITAB v.14 par analyse de la variance à un facteur fixe (présence de fourmis), séparément pour chaque date d'observation. Les tests de corrélation de Pearson ont également été réalisés.
3. Résultats et discussion
3.1. La présence de fourmis influence-t-elle le développement des colonies de pucerons ?
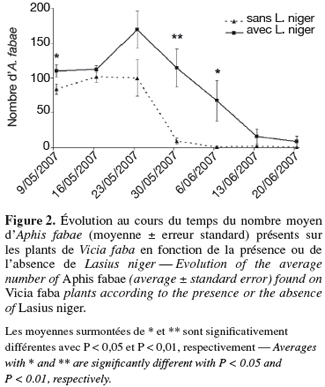
21Taille des colonies de pucerons. L'observation de la figure 2 montre l'évolution au cours du temps du nombre moyen d'A. fabae présents sur les plants de V. faba en fonction de la présence ou de l'absence de L. niger. Le nombre d'A. fabae est supérieur ou égal sur les plantes avec fourmis tout au long des 7 semaines d'observation. L'analyse de la variance montre en effet que les pucerons soignés par les fourmis sont plus nombreux le 9 mai (ANOVA, F1,31 = 4,97 ; P = 0,030), le 30 mai (ANOVA, F1,31 = 14,25 ; P = 0,001) et le 6 juin (ANOVA, F1,31 = 5,15 ; P = 0,031). Les fourmis semblent donc favoriser la croissance des colonies de pucerons. Cette conclusion est en accord avec l'étude de El-Ziady et al. (1956) qui avaient déjà mis en évidence que les colonies d'A. fabae étaient de plus grande taille en présence de fourmis en conditions naturelles. Par contre, Stadler et al. (2002) ont obtenu des résultats opposés. Cependant, cette dernière étude a été réalisée en laboratoire où les pucerons soignés n'ont donc jamais été attaqués par leurs ennemis naturels. La reproduction d'A. fabae est favorisée par la présence de fourmis si les bénéfices qu'elle entraine sont supérieurs aux couts engendrés. Dans le cas présent, comme les pucerons qui ont été soignés par les fourmis sont plus nombreux, on peut supposer que les bénéfices retirés par ces dernières ont été supérieurs aux couts occasionnés par la protection et l'entretien d'une telle colonie de pucerons. Aucune différence de taille de colonie n'a été observée entre les deux cercles concentriques autour de la fourmilière.
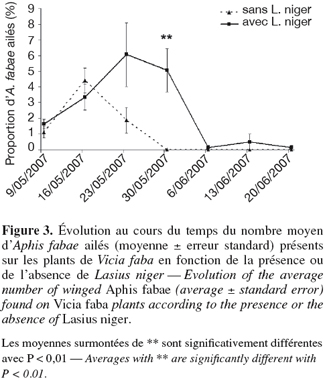
22Proportion de pucerons ailés. L'évolution de la proportion de pucerons ailés au cours des semaines d'observation est présentée à la figure 3. La proportion de pucerons ailés est globalement supérieure dans le cas des plantes avec fourmis mais la différence n'est significative qu'à la date du 30 mai (ANOVA, F1,31 = 13,79 ; P = 0,001). Selon de nombreuses études antérieures (El-Ziady et al., 1956 ; Johnson, 1959 ; Hölldobler et al., 1990), la production d'ailés est retardée dans la saison par la présence de fourmis. En effet, on observe sur le graphique un décalage d'une semaine de la proportion maximale de pucerons ailés en présence de fourmis sur les plantes infestées. Les pucerons ailés étant généralement produits en plus grandes proportions en conditions défavorables, comme la présence répétée de prédateurs ou la baisse de la qualité ou de la quantité de nourriture, il aurait été normal de constater également un nombre d'ailés moins important en présence de fourmis. Cependant, la densité des populations d'A. fabae étant plus importante en présence de fourmis et la production de formes ailées étant positivement corrélée à la densité des populations (Kunert et al., 2005), il n'est pas anormal de retrouver une proportion d'ailés supérieure en présence de fourmis. Nos résultats confirment ceux obtenus par El-Ziady et al. (1956) qui expliquent la stabilisation de la proportion d'individus ailés comme le résultat engendré par l'ensemble des bénéfices accordés par la présence de fourmis, qui tendent également à augmenter la taille et la vigueur de la colonie de pucerons.
3.2. Comment les fourmis influencent-elles le développement des colonies de pucerons ?
23Prédateurs. Les résultats précédents montrent le rôle des fourmis sur le nombre de pucerons composant les colonies et sur la proportion d'individus ailés. Il serait intéressant de mettre en évidence les facteurs sur lesquels les fourmis ont une influence.
24Les espèces de prédateurs aphidiphages ont été recensées et dénombrées. Trois espèces de syrphes ont été observées : Syrphus ribesii L., Metasyrphus corollae Fabricus et Episyrphus balteatus De Geer. Les espèces Propylea quatuordecimpunctata L., Calvia decemguttata L., Coccinella septempunctata L. (Coleoptera, Coccinellidae) ont été également observées. Le site expérimental étant placé à proximité (< 10 m) de l'arboretum de la Faculté universitaire des Sciences agronomiques de Gembloux, il n'est pas étonnant de retrouver certaines espèces plus forestières comme C. decemguttata.
25Le nombre moyen d'espèces de prédateurs observées sur une plante infestée d'A. fabae en présence ou non de fourmis est présenté à la figure 4. On peut observer que le nombre de prédateurs présents sur les plantes visitées par les fourmis est resté, durant toute la durée des observations, égal ou inférieur au nombre de prédateurs sur les plants sans fourmis. La différence entre le nombre de prédateurs observés sur les plantes avec et sans fourmis se révèle significative à la date du 16 mai (ANOVA, F1,31 = 6,97 ; P = 0,013). Le nombre étonnamment élevé de prédateurs sur les plantes sans fourmi du 6 juin ne peut être interprété, le nombre de pucerons sur les plantes à cette date étant très bas. Néanmoins, la présence de miellat sur feuilles et tiges pourrait expliquer l'attraction de ces prédateurs aphidiphages.
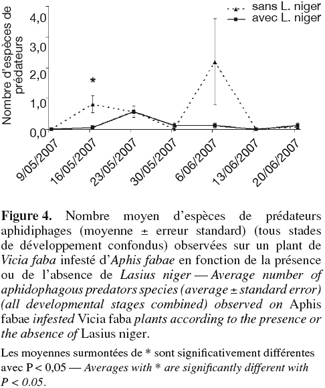
26Ces résultats accordent d'autant plus d'importance au rôle des fourmis que le nombre de pucerons était toujours supérieur sur les plantes visitées par celles-ci. En effet, on s'attendrait à observer un nombre plus important de prédateurs là où le nombre de proies est plus important. Or, ce ne fut pas le cas. Ces résultats confirment d'autres études telles que celles de El-Ziady et al. (1956) et de El-Ziady (1960) qui concluent que les fourmis ont tendance à faire diminuer le nombre de prédateurs sur les plantes où elles sont présentes.
27Parasitoïdes. Les espèces de parasitoïdes ont été recensées et la proportion de pucerons parasités a été calculée lors de chaque semaine d'observation (Figure 5). Les espèces de parasitoïdes identifiées sont Aphidius ervi Haliday, Praon volucre Haliday, Lysiphlebus fabarum Marshall, Lysiphlebus testaceipes Cresson et Adialytus ambiguus Haliday. La proportion de pucerons parasités est restée inférieure à 10 %, sauf en date du 30 mai où cette même proportion s'est accrue tant au niveau des plantes non-visitées que de celles visitées par les fourmis. L'analyse de la variance ne montre pas de différence significative pour aucune des 7 dates. Les fourmis ne procureraient donc pas de protection des pucerons vis-à-vis des parasitoïdes.
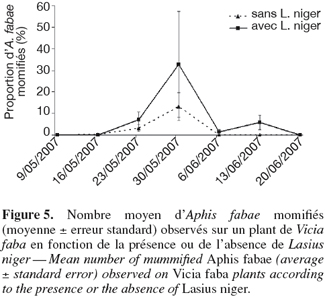
28Malgré l'absence de différence significative, on constate que le nombre de pucerons momifiés est supérieur en présence de fourmis à la date du 30 mai. Les parasitoïdes ont en effet développé des comportements de défense qui leur permet de pratiquer l'oviposition malgré tout (Kaneko, 2003). De plus, la même étude a démontré que la présence de fourmis, notamment L. niger, favorisait l'émergence de parasitoïdes adultes en repoussant notamment les prédateurs intraguildes, en protégeant les pucerons parasités encore vivants des prédateurs et en empêchant l'oviposition d'hyperparasitoïdes (Kaneko, 2003).
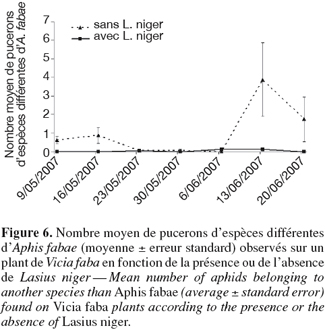
29Autres espèces de pucerons. D'autres espèces de pucerons ont été dénombrées et déterminées. Quatre espèces supplémentaires ont ainsi été identifiées : Acyrthosiphon pisum (Harris), Megoura viciae (Buckton), Macrosiphum euphorbiae (Thomas) et Aulacorthum solani (Kaltenbach). Certains pucerons n'ont pas pu être identifiés en raison de leur stade de développement précoce au moment des observations.
30On remarque à la figure 6 que presque aucun puceron d'espèces différentes d'A. fabae n'a été observé sur les plantes visitées par les fourmis. De petites colonies de pucerons d'espèces précitées ont été décelées sur plusieurs plantes sans fourmis alors qu'aucune colonie (à l'exception de deux pucerons ailés isolés) n'a été observée sur les plantes visitées par les fourmis. L'explication qui peut être donnée à ces différences est celle de la prédation des fourmis sur les pucerons d'espèces autres qu'A. fabae. Les espèces observées durant l'expérience ne sont pas myrmécophiles (Engel et al., 2001 ; Almehdi, communication personnelle), les fourmis présentes ont donc probablement supprimé ces espèces au profit d'A. fabae. Une autre hypothèse consiste à expliquer l'absence de pucerons d'espèces différentes comme le résultat de compétitions interspécifiques entre pucerons, où la prospérité des colonies myrmécophiles prendrait le dessus sur l'installation de nouvelles colonies.
3.3. Les fourmis améliorent-elles la vigueur des plantes infestées ?
31Les observations réalisées sur les plantes infestées ont pour objectif de décrire brièvement leur vigueur afin de déterminer si la présence de fourmis est bénéfique pour celles-ci. Les variables concernant la plante qui ont été observées sont la hauteur et les symptômes apparents de maladies. On observe que la hauteur moyenne des plantes est inférieure à 20 cm jusqu'au 30 mai, c'est-à-dire jusqu'à ce que le nombre de pucerons chute. Les plantes ont grandi à partir de l'absence de pucerons : ces deux variables sont en effet négativement corrélées (Pearson = -0,303 ; P < 0,001).
32Les hauteurs moyennes des plantes visitées et non visitées par les fourmis sont restées identiques durant toute la durée de l'observation, à l'exception du 16 mai où les plantes visitées par les fourmis étaient significativement plus grandes (ANOVA, F 1,31 = 5,8 ; P = 0,022).
33L'observation régulière des plantes a conduit à la constatation d'une meilleure « hygiène » des plantes visitées par les fourmis, surtout du point de vue du nombre d'exuvies et de taches de miellat, même si toutes présentaient régulièrement des nécroses, chloroses, rouilles, feuilles boursouflées ou des traces d'attaques de phytophages.
3.4. Comment a évolué la fréquentation des plantes par les fourmis ?
34La présence des fourmis sur les plantes du dispositif a été observée durant toute l'expérience. Le nombre total de fourmis sur chaque plante augmente jusqu'au 30 mai puis diminue. Ce nombre est corrélé avec le nombre de pucerons sur les plants de fèves de manière hautement significative (Pearson = 0,291 ; P = 0,002). Par contre, aucune corrélation significative n'a été observée entre la température journalière moyenne (Pearson = 0,068 ; P =0,477) ou les précipitations (Pearson = - 0,116 ; P = 0,222).
35Le nombre de fourmis qui se trouvent sur les colonies de pucerons a aussi été observé. Celui-ci évolue parallèlement au nombre total de fourmis sur la plante ; la corrélation entre ces deux variables se révèle très hautement significative (Pearson = 0,916 ; P < 0,001). Cela suggère que les fourmis se déplacent jusqu'aux plantes afin d'établir des relations de mutualisme avec les pucerons.
36Le nombre de fourmis présentes sur les nectaires extrafloraux a enfin été compté. Il est nul jusqu'au 23 mai puis augmente légèrement. Cette évolution est due à deux facteurs principaux :
37– la décroissance du nombre de pucerons sur les plantes,
38– la croissance de la taille des plantes et du nombre de nectaires extrafloraux.
39En effet, si le nombre de pucerons diminue, les fourmis recherchent d'autres sources de sucres dont le nombre augmente alors que la plante grandit.
40Construction d'une structure protectrice. De nombreuses espèces de fourmis construisent des structures physiques externes à leur colonie, telles que des ponts, des tunnels, des abris et des avant-postes (Anderson et al., 2001). Lorsque ces structures sont destinées à protéger les espèces d'insectes avec lesquelles les fourmis entretiennent des relations de mutualisme, elles peuvent porter les noms que Linné leur avait initialement prêtés : « étable » ou « bergerie », mais le terme d'« abris » est plus communément utilisé (Anderson et al., 2001). Lors de la présente étude, la colonie de L. niger initialement placée au centre du dispositif expérimental a installé un avant-poste dans le pot d'un plant infesté de V. faba. En date du 8 mai 2007, une ébauche de construction de terre a été observée à la base de la plante. Cette construction s'est ensuite agrandie pour couvrir l'entièreté de la tige, enfermant les pucerons présents. En date du 24 mai, celle-ci mesurait plus de 5 cm de haut. De nombreuses fourmis étaient présentes continuellement sur la plante. Ce nombre dépassait fréquemment la vingtaine alors que 2,9 ± 0,6 fourmis en moyenne étaient présentes sur l'ensemble des plantes visitées par les fourmis. Aucun prédateur, parasitoïde ou espèce de pucerons autre qu' A. fabae n'a été observé (Figure 7).

4. Conclusion
41L'observation du mutualisme pucerons – fourmis en conditions semi-naturelles a permis d'évaluer les bénéfices retirés par les pucerons. L'évolution différente des colonies de pucerons se nourrissant sur les plantes auxquelles les fourmis avaient accès par rapport à celles dont elles étaient exclues montre que les pucerons retirent un intérêt certain à être soignés par les fourmis. En effet, les pucerons des colonies soignées se sont mieux reproduits que les pucerons des colonies qui ne l'étaient pas. De plus, les fourmis excluent les prédateurs et les pucerons non-myrmécophiles des plantes où A. fabae est présent, diminuant la prédation et la compétition interspécifique pour le phloème. Les soins apportés par les fourmis aux colonies et à la plante (notamment le nettoyage du miellat et des exuvies) bénéficient aux pucerons en ralentissant le développement de champignons. Ces bénéfices pour les pucerons rejoignent directement l'intérêt des fourmis : elles maintiennent leur source de nourriture dans les meilleures conditions possibles.
42Remerciements
43Nous adressons nos remerciements au professeur Charles Gaspar pour la relecture du présent document.
Bibliographie
Anderson C. & McShea D.W., 2001. Intermediate-level parts in insect societies: adaptive structures that ants build away from the nest. Insectes Sociaux, 48, 291-301.
Banks C.J., 1962. Effects of the ant Lasius niger on insects preying on small populations of Aphis fabae on bean plants. Ann. Appl. Biol., 50, 669-679.
Boucher D.H., James S. & Keeler K., 1982. The ecology of mutualism. Annu. Rev. Ecol. Syst., 13, 315-347.
Bristow C.M., 1984. Differential benefits from ant attendance to two species of Homoptera on New York ironweed. J. Anim. Ecol., 53, 715-726.
Bristow C.M., 1991. Why are so few aphids ant-tended? In: Huxley C.R. & Cutler D.F., eds. Ant-plant interactions. Oxford, UK: Oxford University Press.
Buckley R., 1987. Interactions involving plants, Homoptera and ants. Annu. Rev. Ecol. Syst., 18, 111-135.
Cushman J.H. & Addicott J.F., 1991. Conditional interactions in ant-plant-herbivore mutualisms. In: Huxley C.R. & Cutler D.F., eds. Ant-plant interactions. Oxford, UK: Oxford University Press.
Dixon A.F.G., 1985. Aphid ecology. Glasgow, London, UK: Blackie & Son Ltd., 92-103
El-Ziady S. & Kennedy J.S., 1956. Beneficial effects of the common garden ant, Lasius niger, on the black bean aphid, Aphis fabae. Proc. Roy. Soc., 31, 61-65.
El-Ziady S., 1960. Further effects of Lasius niger L. on Aphis fabae Scopoli. Proc. Roy. Soc., 35, 33-38.
Engel V., Fischer M., Wäckers F. & Völkl W., 2001. Interactions between extrafloral nectaries, aphids and ants: are there competition effects between plant and Homopteran sugar sources? Oecologia, 129, 577-584.
Fischer M. & Shingleton A., 2001. Host plant and ants influence the honeydew sugar composition of aphids. Funct. Ecol., 15, 544-550.
Hölldobler B. & Wilson E.O., 1990. The ants. Cambridge, UK: Harvard University Press.
Huber P., 1810. Recherches sur les mœurs des fourmis indigènes. Paris ; Genève, Suisse : J.J. Paschoud.
Johnson B., 1959. Ants and form reversal in aphids. Nature, 184, 740-741.
Kaneko S., 2003. Impacts of two ants, Lasius niger and Pristomyrmex pungens (Hymenoptera: Formicidae), attending the brown citrus aphid, Toxoptera citricidus (Homoptera: Aphididae), on the parasitism of the aphid by the primary parasitoid, Lysiphlebus japonicus (Hymenoptera: Aphidiidae), and its larval survival. Appl. Entomol. Zool., 38(3), 347-357.
Kunert G. et al., 2005. Alarm pheromone mediates production of winged dispersal morphs in aphids. Ecol. Lett., 8(6), 596-603.
Mahdi T. & Whittaker J.B., 1993. Do birch trees (Betula pendula) grow better if foraged by wood ants? J. Anim. Ecol., 62, 101-116.
Pontin A.J., 1959. Some records of predators and parasites adapted to attack aphids attended by ants. Entomol. Mon. Mag., 95, 154-155.
Sloggett J.J. & Majerus M.E.N., 2000. Aphid-mediated coexistence of ladybirds (Coleoptera: Coccinellidae) and the wood ant Formica rufa: seasonal effects, interspecific variability and the evolution of a coccinellid myrmecophile. Oikos, 89, 345-359.
Stadler B. & Dixon A.F.G., 1998. Costs of ant attendance for aphids. J. Anim. Ecol., 67, 454-459.
Stadler B., Dixon A.F.G. & Kindlmann P., 2002. Relative fitness of aphids: effects of plant quality and ants. Ecol. Lett., 5, 216-222.
Stadler B. & Dixon A.F.G., 2005. Ecology and evolution of aphid-ant interactions. Ann. Rev. Ecol. Evol. System., 36, 345-372.
Sudd J., 1967. An introduction to the behaviour of ants. London: Edward Arnold.
Verheggen F.J. et al., 2007. Electrophysiological and behavioral responses of the multicolored Asian Lady Beetle, Harmonia axyridis Pallas, to sesquiterpene semiochemicals. J. Chem. Ecol., 33, 2148-2155.
Verheggen F.J. et al., 2008. Aphid and plant secondary metabolites induce oviposition in an aphidophagous hoverfly. J. Chem. Ecol., 34(3), 301-307.
Völkl W. et al., 1999. Ant-aphid mutualisms: the impact of honeydew production and honeydew sugar composition on ant preferences. Oecologia, 118, 483-491.
Way M.J., 1963. Mutualism between ants and honeydew-producing Homoptera. Ann. Rev. Ecol., 8, 307-344.
Yao I., Shibao H. & Akimoto S.I., 2000. Costs and benefits of ant attendance to the drepanosiphid aphid Tuberculatus quercicola. Oikos, 89, 3-10.
Yao I. & Akimoto S.I., 2001. Ant attendance changes the sugar composition of the honeydew of the drepanosiphid aphid Tuberculatus quercicola. Oecologia, 128, 36-43.
Pour citer cet article
A propos de : François Verheggen
Gembloux Agricultural University – FUSAGx. Unité d'Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : entomologie@fsagx.ac.be
A propos de : Lise Diez
Gembloux Agricultural University – FUSAGx. Unité d'Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
A propos de : Claire Detrain
Université libre de Bruxelles. Unité d'Ecologie sociale. CP 231. Boulevard du Triomphe. B-1050 Bruxelles (Belgique).
A propos de : Éric Haubruge
Gembloux Agricultural University – FUSAGx. Unité d'Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
