


- Accueil
- volume 13 (2009)
- numéro 3
- Criblage et performances agronomiques de 45 génotypes de pois chiche (Cicer arietinum L.) soumis à un régime hydrique limité


Visualisation(s): 9883 (38 ULiège)
Téléchargement(s): 641 (3 ULiège)

Criblage et performances agronomiques de 45 génotypes de pois chiche (Cicer arietinum L.) soumis à un régime hydrique limité

Notes de la rédaction
Reçu le 21 mai 2008, accepté le 2 décembre 2008
Résumé
Le pois chiche (Cicer arietinum L.), l’une des plus importantes légumineuses à graines en Tunisie, est pratiqué en deux types de culture : pois chiche d’hiver et pois chiche de printemps. Le pois chiche de printemps a l’avantage d’occuper le sol pendant une courte durée (février à juin). Néanmoins, la culture est exposée aux stress abiotiques et principalement le stress hydrique. En conséquence, le développement végétatif et le rendement potentiel sont négativement affectés en quantité et en qualité. Dans l’objectif de cribler des génotypes de pois chiche tolérants au stress hydrique, une collection de 45 accessions a été testée, en culture tardive de printemps, sous un régime hydrique stressant. L’essai in situ a été réalisé dans la région de Chott Mariem (Tunisie) appartenant à l’étage bioclimatique du semi-aride supérieur. Le stress hydrique a été imposé au début de la floraison et s’est accentué durant les phases de grossissement et de maturité des graines. L’évaluation de la tolérance à la sècheresse des génotypes de pois chiche a été basée sur des paramètres agronomiques relatifs au rendement en grains et ses composantes, à l'efficience de l'utilisation de l'eau et à la durée thermique. L'analyse statistique a révélé une importante diversité biologique entre les génotypes testés. Les 45 accessions peuvent être réparties en trois groupes. Un premier groupe est composé de génotypes 'Kabuli', sensibles à la sècheresse, ne convenant pas pour une culture de printemps dans les zones semi-arides. Un second groupe de génotypes, parmi lesquels deux sont du type 'Dési', est tolérant au stress hydrique et peut être recommandé pour une culture de printemps dans les zones semi-arides. Le dernier groupe renferme des génotypes moyennement sensibles au stress hydrique. Ils peuvent être conduits en culture de printemps dans les zones du semi-aride supérieur avec des irrigations complémentaires.
Abstract
Screening and agronomic performances of 45 chickpea genotypes (Cicer arietinum L.) submitted to a limited hydrous pattern. In Tunisia, chickpea (Cicer arietinum L.) is one of the most important legume grain crops. It is conducted during two periods of the year either as winter crop or as spring crop. The last type has the advantage to occupy the soil in a short period of time (February to June). However, it exposes plants to water stress. Consequently, vegetative development and grain yield are negatively affected in quantity and quality. In order to study plant reactions to drought conditions, 45 genotypes of chickpea (C. arietinum) were tested in delayed spring culture under a water stressed treatment. A field trial was carried out in the region of Chott Mariem belonging to the semi-arid superior. The drought stress is established at the beginning of flowering stage and was accentuated during the grain filling and the seed maturity phases. To evaluate the chickpea genotypes stress tolerance, parameters related to the vegetative development, the seed yield, the water use efficiency and the thermal duration were measured. The analysis of the grain yield and its components revealed that there is a biological diversity among the genotypes tested. The 45 accessions can be divided into three groups. The first one is composed of 'Kabuli' chickpea genotypes which are sensitive to drought stress. They are not appropriate for spring crop in semi-arid regions. The second group is composed of genotypes, of which two are of 'Dési' type, that are tolerant to drought stress and can be recommended for a spring crop in the superior semi-arid zones. The last group of genotypes is moderately sensitive to drought stress. They can be led as spring crop in the semi-arid superior zones with complementary irrigations.
Table des matières
1. Introduction
1Les légumineuses alimentaires constituent une source très importante de protéines végétales qui peuvent corriger le déficit en protéines animales. En plus de leur importance dans le régime alimentaire des êtres humains, elles ont un intérêt particulier dans le concept d’agriculture durable dans la mesure où leur introduction dans l’assolement instaure la rotation des cultures, la diversification des productions et la protection de l’environnement par l’économie des engrais azotés et des désherbants chimiques. Pour pouvoir introduire ces espèces dans les systèmes de culture, il faudrait impérativement améliorer les performances génétiques des variétés.
2En Tunisie, la culture des légumineuses à graines reste marginale par rapport aux céréales. Elles n’occupent que 6 à 7 % des superficies emblavées en céréales. Le pois chiche (Cicer arietinum L.) et la fève (Vicia faba L.) occupent 86 % des superficies des légumineuses avec une production annuelle de 22 000 tonnes (Kramer, 1983) et un rendement moyen ne dépassant pas 7 qx.ha-1. La production nationale ne couvre pas les besoins internes du pays et l’Etat a recours à des importations (Aouani et al., 2001). Dans nos conditions, deux types de culture de pois chiche sont pratiquées, notamment le pois chiche d’hiver et le pois chiche de printemps. Ils diffèrent entre eux par les dates de semis en novembre-décembre pour le pois chiche d’hiver et en fin février-début mars pour le pois chiche de printemps (Saccardo et al., 1990). Le cycle de culture est de huit mois pour un semis d’hiver et de trois mois seulement pour un semis de printemps. Singh et al. (1994) ont indiqué que le semis précoce permet à la culture de pois chiche de profiter des pluies hivernales engendrant un développement végétatif et un rendement meilleurs. En fait, le pois chiche d’hiver a un rendement potentiel plus élevé que celui du pois chiche de printemps (Saxena, 1980 ; Malhotra, 1998). Néanmoins, le pois chiche d’hiver ne manque pas d’inconvénients. L'occupation du sol est plus longue et les travaux d’entretien, notamment le désherbage, sont multiples. Les conditions climatiques hivernales caractérisées par une hygrométrie relative élevée de l’ordre de 93 % et des températures clémentes, variant de 9 à 18,5 °C (Habaieb et al., 2003), favorisent le développement des maladies très redoutables, en l’occurrence le fusarium (Fusarium oxysporum), l’anthracnose (Ascochyta rabiei) et autres, telles que la pourriture grise (Botrytis cinerea), la pourriture humide des racines (Fusarium solani) et la pourriture sèche des racines (Rhizoctonia bataticola), surtout pour les variétés sensibles.
3Le pois chiche est une espèce rustique par son système radiculaire puissant qui se développe dans les deux sens, latéral et pivotant, et qui peut dépasser un mètre de profondeur (Saxena, 1987). Malgré cela, le pois chiche de printemps craint la sècheresse accompagnée de stress hydrique et thermique (Singh et al., 1994). Le stress hydrique reste la principale contrainte limitante de la production du pois chiche cultivé sous un régime pluvial (Kramer, 1983 ; Blum et al., 1989). Il dépend de plusieurs facteurs, entre autres les fréquences et la répartition des précipitations le long du cycle de la culture, l’évaporation et la capacité de stockage de l'eau dans le sol (Belhassen et al., 1995).
4Comme second stress abiotique lié au stress hydrique, les hautes températures supérieures à 38 °C retardent considérablement la floraison du pois chiche (Ellis et al., 1994). Les températures supérieures à 30 °C, qui se manifestent durant une période de 3 à 4 jours de suite, occasionnent de lourdes pertes du rendement. D’après Singh et al. (1994), 50 % des fleurs des plantes exposées durant l’anthèse à une température supérieure à 30 °C sont presque stériles. Slama (1998) a indiqué que les cultivars dont les gousses ont mûri pendant des jours chauds ont subi des réductions du rendement en grains.
5Singh et al. (1994) ont préconisé qu’un semis retardé d’une culture de pois chiche, conduite en sec ou en irrigué, permet de déterminer son rendement potentiel. Le screening des génotypes de pois chiches tolérants au stress hydrique pourrait être effectué en retardant le semis de trois semaines de la date normale du semis de printemps (Malhotra et al., 1996). En outre, selon Sarrafi et al. (1993), il est difficile de sélectionner directement pour la résistance à la sècheresse car le contrôle génétique de ce caractère quantitatif est très complexe. L’approche actuelle consiste à sélectionner pour plusieurs paramètres liés à la résistance au stress hydrique.
6Sous des conditions de sècheresse, la sélection empirique des génotypes tolérants au stress hydrique est basée sur le rendement en grains et ses composantes. Toutefois, toutes les composantes adoptées pour ce type de criblage devraient être caractérisées par une forte corrélation avec un rendement en grains élevé et stable, un niveau d’héritabilité élevé et une expression de la tolérance facilement mesurable et répétitive (Serraj et al., 2003). D’après Ofori (1996), le rendement en grains le plus élevé est prévisible si toutes les composantes du rendement sont à leurs niveaux maximums.
7Yousaf et al. (1999a ; 1999b) ont indiqué que le rendement en grains est un caractère complexe qui résulte de l’interaction d’une multitude de facteurs hautement sensibles aux variations environnementales. Il pourrait être estimé sur la base de la performance d’un certain nombre de composantes morphophysiologiques et phénologiques telles que la hauteur de la plante, le nombre de branches, de gousses et de graines par plant, la date de floraison et le poids de 100 graines (Singh, 1977). Selon Jain et al. (1991), la combinaison entre ces composantes s’est révélée le meilleur moyen pour l’amélioration du rendement. Silim et al. (1993) ont indiqué que le rendement potentiel élevé et la floraison précoce représentent les deux principaux critères de sélection pour la tolérance au stress hydrique de la lentille et du pois chiche. Singh et al. (1991) ont conclu que, sous des conditions de stress hydrique, 75 % des variations du rendement en grains sont attribués aux dates de floraison et de maturité et au poids de 100 graines. Seulement, ces variations, traduites par des corrélations négatives entre certaines de ces composantes, peuvent être atténuées par des phénomènes de compensation (Ofori, 1996).
8L’indice de récolte, qui indique le degré de reconversion d’une partie de la biomasse aérienne en graines (Bouzerzour et al., 1998), est également considéré comme un critère potentiel pour la sélection indirecte des génotypes à haut rendement et tolérants au stress hydrique (Berger et al., 2006).
9Par ailleurs, Kuruvadi (1989) et Erchidi et al. (2000) ont signalé que le stress hydrique induit une diminution de la densité stomatique qui ne se traduit pas toujours par une réduction des pertes en eau par la plante à cause d’un phénomène de compensation lié à l’augmentation de la taille des stomates (Sapra et al., 1975 ; Kuruvadi, 1989 ; Wang et al., 1993). De même, Huang et al. (2000) ont indiqué que l’efficience de l’utilisation de l’eau est un important facteur pour la détermination de la résistance au déficit hydrique. Sur la base de ce critère, le pois chiche d’hiver a été trouvé le plus efficient (Kamel, 1990 ; Saccardo et al., 1990 ; Soltani et al., 1999).
10En Tunisie, la culture du pois chiche est exposée à deux types de sècheresse. L’une est intermittente et causée par la rupture des précipitations et l’autre est terminale et se produit pendant la phase de floraison et de remplissage des graines. Les dégâts les plus graves se manifestent sur le pois chiche cultivé dans les zones semi-arides où le rendement en grains peut chuter de 40 % à 100 % (Saxena, 1987 ; Slama, 1998). Pour remédier à cette contrainte de production, la solution la plus efficace réside dans l'amélioration de la tolérance des génotypes de pois chiche à la sècheresse et de l'efficience de l'utilisation de l'eau (Boubaker et al., 1995 ; 1997). Dans un programme d'amélioration des légumineuses à graines, le développement des génotypes de pois chiches tolérants au stress hydrique et adaptés à une large gamme d'humidité variant d'une saison à une autre et d'un milieu à un autre est un objectif de premier ordre. Le présent travail est réalisé dans l’objectif de sélectionner les génotypes les plus performants pour une culture de pois chiche de printemps en zones semi-arides qui couvrent les deux tiers du territoire tunisien (MEAT, 2001). Le criblage des génotypes de pois chiche est basé sur la détermination du bilan hydrique de cette culture, la délimitation des stades phénologiques et leur coïncidence avec l’avènement des stress hydrique et thermique et l’évaluation de l’impact de ces stress sur le rendement et ses composantes.
2. Matériel et méthodes
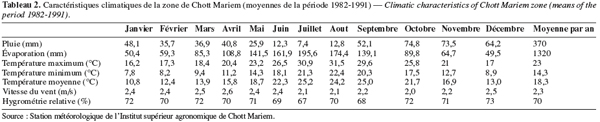
11Le comportement agronomique de 45 génotypes de pois chiche (Tableau 1), dont 43 sont du type 'Kabuli' et 2 du type 'Dési', a été évalué sous les conditions pluviales de la région semi-aride de Chott Mariem qui est localisée au centre est de la Tunisie (latitude : 35°52’ nord, longitude : 10°38’ est et altitude : 6 m). Le microclimat du site expérimental est semi-aride supérieur à hiver doux et humide et été chaud et sec (Tableau 2). Le sol est peu évolué d’origine non climatique, d’apport alluvial, vertique, de texture sablo-limoneuse, de structure massive et favorable aux grandes cultures. Ce sol a un taux d’humidité (Hpr) de 24,4 % à la capacité au champ et de 12,3 % au point de flétrissement permanent.
12Le précédent cultural est une jachère travaillée. Avant l’installation de l’essai, le sol a été préparé par un labour moyen, d’une profondeur de 30 cm, suivi par deux façons superficielles en vue d’obtenir un bon lit de semences. Les niveaux phosphaté et potassique du sol, déterminés par analyse, étaient favorables à une production optimale de légumineuses. Par conséquent, aucun apport de ces deux éléments n’a été réalisé. Le semis a été effectué le 16 avril 2006 (soit un retard de 5 semaines par rapport à la date normale) à la densité de 22,2 plants par mètre carré et avec des écartements de 0,1 x 0,45 m2. La lutte contre les mauvaises herbes envahissant les parcelles expérimentales a été réalisée par un désherbage manuel.
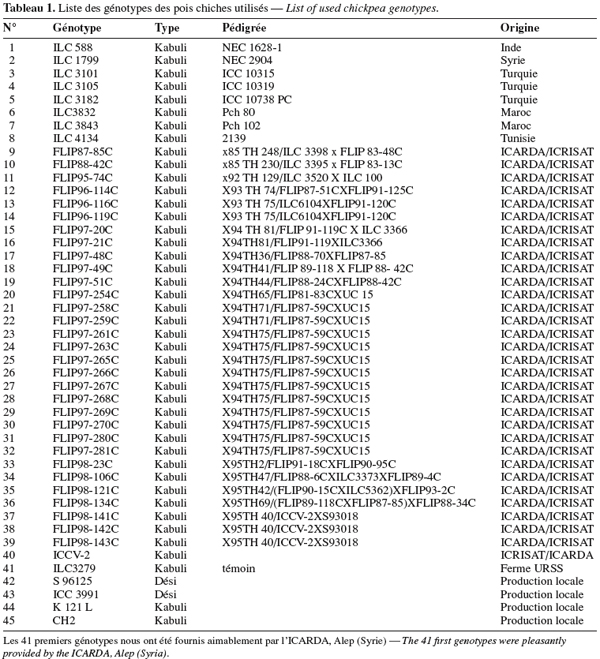
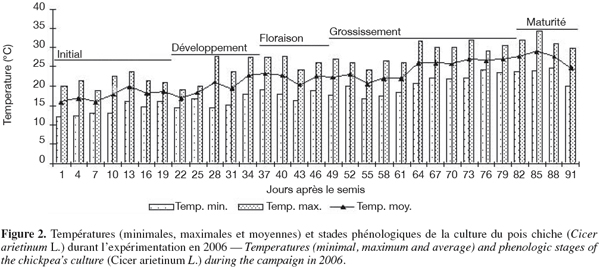
13Des irrigations localisées d’appoint ont été réalisées avec de l’eau du barrage de Nébhana dont le résidu sec est de 1 g.l-1. Les goutteurs intégrés du système d’irrigation sont du type GR, avec un écartement de 0,4 m et ont un débit nominal de 2 l.h-1 chacun. Le test d’uniformité réalisé a montré que le débit réel moyen des goutteurs est de 1,75 l.h-1. La quantité totale d’eau reçue par la culture est de 171,7 mm dont 46,7 mm sont d’origine pluviale (Figure 1). Les températures minimales et maximales varient respectivement de 10,6 à 28,7 °C et de 18,2 à 36,5 °C (Figure 2). L’évapotranspiration de la culture (ETc) a été déterminée dans l’intervalle séparant deux irrigations successives suivant l’équation simplifiée du bilan hydrique du sol (Choisnel, 1984) :
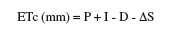
14où P = Précipitations (mm) ; I = Irrigation (mm) ; D = Drainage (mm) et ∆S = variations de l’humidité dans le sol (mm).
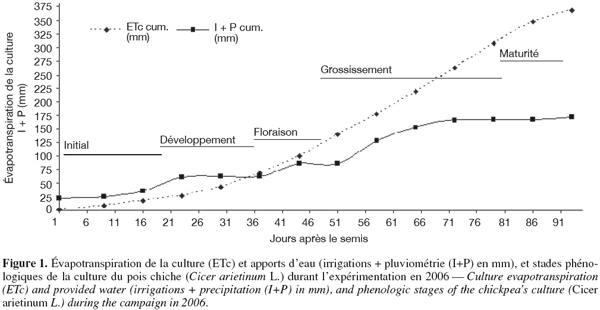
15L’ETc (mm) a été pratiquement calculée sur la base de l’évapotranspiration de référence (ET0) et le coefficient cultural (Kc) (Ben Mechlia, 1998) selon la relation :
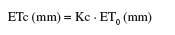
16L’évapotranspiration de référence (ET0) a été calculée à partir de la formule de Blanney-Criddle (Poirée et al., 1966). Le coefficient cultural (Kc) ainsi que les durées des phases physiologiques du pois chiche ont été adoptés conformément aux normes utilisées par la FAO (Allen et al., 1998).
17Les variables agronomiques étudiées sur l'ensemble des génotypes sont les suivantes :
18– la date de floraison (j) : le nombre de jours depuis le semis jusqu'à l’épanouissement de 50 % des fleurs,
19– le poids de 100 graines (g),
20– le rendement en grains par m2 (g.m-2) : le poids total des graines récoltées par m2,
21– la production de biomasse aérienne par m2 (g.m-2) : le poids sec total de la partie aérienne des plantes récoltées par m2,
22– l’indice de récolte (%) : le rapport du rendement en grains par la production de biomasse aérienne par m2,
23– l’efficience de l’utilisation de l’eau (kg.m-3) : le rapport du rendement en grains obtenu par l’évapotranspiration de la culture,
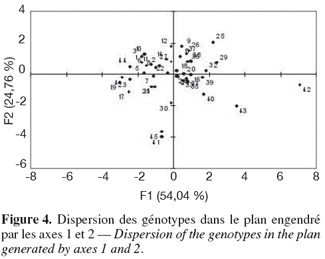
24– la durée thermique (comme déterminée par Gate, 1995) en degré par jour calculée comme suit :
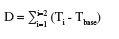
25où Ti = Température moyenne journalière, soit (T max. + T min)/2 et Tbase = Température de base qui représente la température en dessous de laquelle la plante ne fleurit pas. Pour le pois chiche, Tbase est estimée à 15 °C (Slama, 1998).
26Le dispositif expérimental adopté est en blocs aléatoires randomisés avec trois répétitions. Les résultats obtenus lors de ce travail ont été traités par une analyse de variance à un facteur pour tester les différences entre les cultivars. Les moyennes ont été soumises au test des groupes homogènes (Student-Newman et Keuls au seuil de 5 %). Une analyse en composantes principales (ACP) est faite pour identifier les principales variables agronomiques discriminantes des génotypes soumis au régime hydrique limité. Le but est d'identifier les variables qui pourraient servir de critères de base pour la sélection des génotypes tolérants au stress hydrique. Les logiciels utilisés à ces fins sont SPSS 10.0 et XLSTAT 2008.
3. Résultats et discussion
27Le bilan hydrique de la culture du pois chiche a montré que les stades phénologiques au sein de l'essai sont répartis sur deux phases : une première phase sous stress composée des stades initial et développement et une seconde phase durant laquelle le stress hydrique a affecté les stades de grossissement et de maturité (Figure 1). Cette phase de stress a coïncidé avec le début de la floraison des génotypes précoces, soit 36 jours après le semis. En fait, la phase de floraison, qui est particulièrement sensible à la sècheresse, a un effet direct sur la date de maturité et le rendement en grains de la culture du pois chiche (Summerfield et al., 1996).
28Durant le stade floraison, la température maximale a varié de 25,9 à 29 °C avec une moyenne de 26,8 °C. Tandis qu’aux stades remplissage des graines et maturité, elle a varié de 24,7 à 32,2 °C avec une moyenne de 30,1 °C (Figure 2). Roberts et al. (1980) ont remarqué une augmentation linéaire du taux de floraison du pois chiche à des températures variant de 10,8 à 29,3 °C. D'ailleurs, des températures variant de 20 à 30 °C le jour et une température voisine de 20 °C la nuit favorise un bon développement du pois chiche (Skrypetz, 2002). Par contre, Singh et al. (1994) ont indiqué que le pois chiche est sensible aux hautes températures durant la phase reproductive et surtout pendant les phases de remplissage des graines et de maturité. De même, comme déjà précisé, l’exposition d’une culture de pois chiche au cours de l’anthèse à des températures supérieures à 30 °C pendant 3 à 4 jours occasionne la stérilité de 50 % des fleurs et une réduction progressive du rendement en grains. D’après Ellis et al. (1994), les températures élevées, supérieures à 38 °C, retardent considérablement la floraison chez le pois chiche. En fait, il semble que la culture de pois chiche a subi un léger stress thermique durant les stades de remplissage des graines et de maturité.
3.1. Analyse individuelle des variables agronomiques étudiées
29Les résultats de l’analyse de la variance à un facteur ont montré qu’il y a une variabilité génotypique hautement significative pour tous les paramètres étudiés. Les coefficients de variation sont faibles et varient de 2,22 % pour la date de floraison à 8 % pour l’efficience de l’utilisation de l’eau et la durée thermique (Tableau 3). Ces résultats révèlent la précision élevée de l’essai.
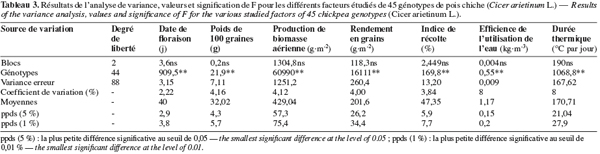
30La date de floraison des génotypes de pois chiche varie de 36 à 47 jours après le semis (Tableau 4). Chez le pois chiche, ce paramètre exerce un effet important sur l’expression du potentiel génétique. En effet, Silim et al. (1993) ont indiqué qu’en présence du stress hydrique, la date de floraison contribue avec 37 à 69 % dans la variation génotypique totale du rendement du pois chiche. Les durées des phases de développement du pois chiche qui est considéré comme une plante de jours longs, particulièrement la date de floraison, dépendent étroitement du génotype, de la température et de la photopériode (Roberts et al., 1980 ; Perez de la Vega, 1996). La comparaison des moyennes a permis de définir 8 groupes homogènes de cultivars présentant beaucoup de chevauchements entre eux. Le premier groupe, ayant la date de floraison la plus précoce, est composé des génotypes 4, 44, 9, 10, 6, 23, 27, 3, 1, 25, 36, 45, 5, 37, 12, 13, 19, 26, 7, 20, 11, 2, 21, 24, 8, 14, 17, 22, 38, 16, 34, 39, 15, 18, 35, 33. Le dernier groupe, qui a la date de floraison la plus tardive, est composé des génotypes 43, 29, 32, 42, 40, 30, 41. Le génotype ILC 3105 parait avoir la floraison la plus précoce (36 jours après le semis), alors que ILC3279 parait avoir la floraison la plus tardive (47 jours après le semis). Le premier groupe a tendance à manifester un mécanisme d’échappement ou d’esquive au déficit hydrique. Abernethy (1987) a indiqué que la floraison précoce engendre une longue période de remplissage des graines et par conséquent, un rendement potentiel élevé. Une floraison tardive induit une courte période reproductive et un faible rendement en grains. Malhotra et al. (2002) ont signalé que la floraison précoce est la principale composante d'évitement du stress hydrique chez le pois chiche. Ce type de résistance traduit la capacité de la plante de terminer son cycle de développement avant l’épuisement des réserves de l’eau dans le sol. La floraison précoce a été largement utilisée comme critère de sélection de génotypes tolérant le stress hydrique (Saxena et al., 1993). L'exploitation de la variabilité génétique de la durée du cycle des cultures est un moyen important dans l'ajustement du cycle de la culture à la saison pluvieuse. La superposition de la saison de croissance à la saison pluvieuse permet de profiter des disponibilités en eau pour la transpiration et fait échapper la culture au stress hydrique durant sa période reproductive. La durée de la saison des pluies est généralement de 4 à 5 mois dans le semi-aride tunisien. L'utilisation de variétés plus ou moins précoces permet une augmentation et une stabilisation des rendements dans ces zones.
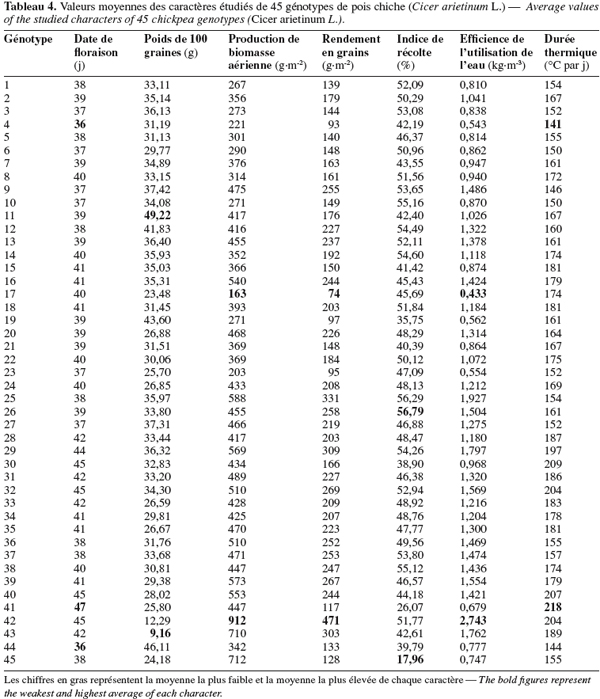
31La température est un facteur déterminant du rendement en grains du pois chiche puisqu’elle règle son rythme de croissance et de développement. En fait, une plante ne peut passer d’un stade phénologique à un autre que si ses durées thermiques sont satisfaites. Les effets de la température varient en fonction du stade de développement végétatif. L’apparition des températures extrêmes entraine des dégâts irréversibles qui se traduisent par la destruction des fleurs, des gousses et parfois même de la plante entière. Les durées thermiques déterminées pour la floraison des génotypes de pois chiche varient de 141 à 218 °C par jour (Tableau 4). La comparaison des moyennes permet de définir 9 groupes homogènes de cultivars. Le premier groupe, caractérisé par les plus faibles durées thermiques, est composé des génotypes 4, 44, 9, 10, 6, 27, 3, 23, 1, 25, 36, 45, 5, 37, 12, 19, 7, 13, 26, 20, 11, 2, 21, 24, 8, 17, 14, 38, 22, 34, 16, 39, 35, 15, 18, 33. Le dernier groupe, caractérisé par les durées thermiques les plus élevées, est composé des génotypes 31, 28, 43, 29, 32, 42, 40, 30, 41. Les génotypes à floraison précoce paraissent caractérisés par de faibles durées thermiques, alors que les génotypes à floraison tardive sont exigeants en durée thermique. Afin de palier aux effets néfastes des hautes températures, les génotypes à floraison tardive devraient avoir un semis précoce.
32L'efficience de l'utilisation de l'eau, définie comme étant le ratio entre la production en graines et la quantité d'eau consommée par la plante, exprime la capacité des feuilles d’échanger l'eau contre le gaz carbonique et traduit le rapport entre la photosynthèse et la transpiration. Huang et al. (2000) ont indiqué qu’elle représente un important critère de sélection pour la tolérance au stress hydrique. Dans nos conditions, ce paramètre varie de 0,43 à 2,74 kg.m-3 d’eau (Tableau 4). La comparaison des moyennes permet de faire apparaitre 18 groupes homogènes. Le génotype 42 s’individualise et se montre le plus efficient. Un groupe assez efficient est composé des génotypes 43, 29, 25. Par contre, le groupe le moins efficient est composé des génotypes 17, 4, 23, 19. Il en découle que, dans les conditions de l’essai, le premier groupe de génotypes de pois chiche tolère le déficit hydrique et peut donc être adopté pour une culture printanière. Tandis que le second groupe, qui chevauche avec celui qui renferme les génotypes 45 et 41 (ILC 3279 ou Chetoui), a manifesté une forte réduction de l’efficience de l’utilisation de l’eau. Il pourrait être conduit en culture d’hiver et ne devrait pas être envisagé pour une culture de printemps. L'efficience de l'utilisation de l'eau peut être améliorée par le choix de génotypes adaptés au déficit hydrique de la région considérée et par l'ajustement des besoins en eau de la culture à l'offre climatique. En Tunisie, la première alternative est la plus efficace car les précipitations sont toujours variables dans le temps et dans l’espace.
33Le poids de 100 graines varie de 9,1 à 49,2 g (Tableau 4). La comparaison des moyennes fait apparaitre 11 groupes homogènes. Le premier groupe est formé des génotypes du type 'Dési' : 42 et 43, qui ont produit des graines du plus petit calibre. Il est suivi par un groupe de génotypes du type 'Kabuli' : 17, 45, 23, 41, 33, 35, 24, 20, 40, 39, 6, 34, 22, 38, dont les graines sont un peu plus grosses, tandis que les génotypes 11 et 44 ont les graines du plus gros calibre. Les génotypes 12 et 19 ont produit des graines de moindre calibre. Meadley et al. (1970) ont suggéré que le poids de 100 graines est un important caractère agronomique pour la sélection de génotypes à haut rendement. De même, Acevedo et al. (1987) ont signalé que, sous contrainte hydrique, les plantes tolérantes au stress hydrique sont caractérisées par des poids de 100 graines élevés. Il parait donc que les deux derniers groupes de génotypes testés tolèrent plus le stress hydrique que les premiers groupes. Miller et al. (2002) ont signalé que le pois chiche du type 'Dési' est caractérisé par des graines de petit calibre, par comparaison à celui du type 'Kabuli'. Ceci pourrait indiquer que, chez le pois chiche, le poids de 100 graines est un caractère génétique. En fait, les deux génotypes, S 96125 et ICC 3991, sont du type 'Dési' et ont les plus faibles poids de 100 graines.
34La production de biomasse aérienne varie de 163 à 912 g.m-2 (Tableau 4). La comparaison des moyennes fait apparaitre 19 groupes. Le groupe de génotypes ayant la plus faible production de biomasse aérienne est composé de 17, 23, 4, alors que le génotype 42 représente, à lui seul, le groupe ayant produit la biomasse la plus élevée. Les génotypes 43 et 45 forment le groupe précédent et ont produit des biomasses aériennes moins élevées. D’après Pacucci et al. (2006), dans des conditions de sècheresse, la vigueur des plants et la production élevée de la matière sèche pourraient être envisagées comme principales caractéristiques pour le maintien et l’amélioration du rendement du pois chiche. Boubaker et al. (1995) ont rapporté que sous hautes conditions de stress hydrique, la vigueur des plantules de céréales peut être utilisée pour l’identification des génotypes tolérants au stress hydrique.
35Le rendement en grains varie de 74 à 471 g.m-2 (Tableau 4). La comparaison des moyennes permet de distinguer 18 groupes homogènes de génotypes. Le groupe ayant le plus faible rendement en grains est composé des génotypes 17, 4, 23, 19. Le génotype 42 représente, à lui seul, le groupe ayant le rendement en grains le plus élevé. Il est précédé par le groupe formé des génotypes 43, 29, 25, 42 qui ont donné des rendements en grains moins élevés. Singh et al. (1994) ont indiqué que les génotypes tolérants au stress hydrique produisent 40 à 50 % de leur rendement en grains potentiels, alors que les génotypes sensibles n’en produisent que 10 %. D’après Sarrafi et al. (1993), la résistance à la sècheresse d’un génotype pourrait être évaluée par sa capacité à maintenir un rendement acceptable en conditions de déficit hydrique.
36L’indice de récolte varie de 18 à 56,8 % (Tableau 4). La comparaison des moyennes fait apparaitre 13 groupes homogènes de génotypes. Le génotype 45 représente le premier groupe et a le plus faible indice de récolte. Les génotypes 16, 17, 31, 5, 39, 27, 23, 35, 24, 20, 28, 34, 33, 36, 22, 2, 6, 8, 42, 18, 1, 13, 32, 3, 9, 37, 29, 12, 14, 38, 10, 25, 26 forment le groupe caractérisé par l'indice de récolte le plus élevé. Selon Rosielle et al. (1975), le rendement en grains et l’indice de récolte sont influencés par les conditions du milieu de culture. D’ailleurs, Silim et al. (1993) ont remarqué qu’un haut indice de récolte est associé à une moindre sensibilité à la sècheresse.
3.2. Analyse en composantes principales (ACP) des variables étudiées
37L’analyse en composantes principales (Frontier, 1981) a montré que le rendement en grains, l’efficience de l’utilisation de l’eau et la production de biomasse aérienne ont les contributions les plus élevées dans l’édification du premier axe avec des valeurs respectives de 28,43, 28,43 et 24,68 %, alors que l’axe 2 est composé principalement de l’indice de récolte (49 %), du poids de 100 graines (19,55 %) et de la date de floraison (18,65 %) (Tableau 5).
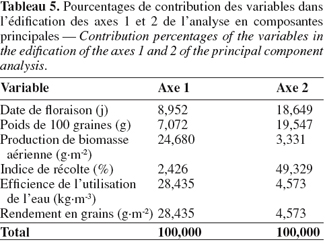
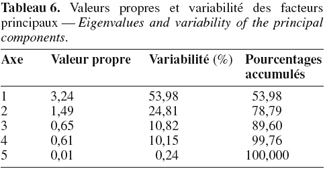
38Les deux premiers axes (1 et 2) ont les valeurs propres les plus élevées. Ils totalisent à eux seuls 78,79 % de la variabilité totale (Tableau 6). Le premier axe absorbe à lui seul 53,98 % de la variabilité observée. C’est un axe de rendement et d’efficience de l’utilisation de l’eau. Il est en corrélation négative avec le poids de 100 graines et positive avec la date de floraison, la production de la biomasse aérienne, le rendement en grains et l’efficience de l’utilisation de l’eau (Figure 3). Il sépare les génotypes principalement selon leur tolérance au stress hydrique et à moindre importance selon leur précocité. Le second axe explique 24,8 % de la variabilité observée. Il est en corrélation négative avec la date de floraison et positive avec l’indice de récolte et le poids de 100 graines. C’est un axe d’architecture du plant et de calibre des graines. Il permet de différencier les génotypes selon le calibre des graines et l’indice de récolte.
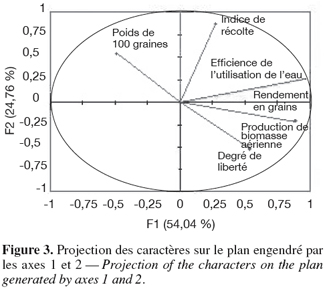
39En considérant l’axe 1, on distingue deux groupes de génotypes (Figure 4). Le premier groupe est formé des génotypes qui ont toléré le stress hydrique. Ils ont produit de hauts rendements en grains et en biomasse aérienne. Ils sont caractérisés par une efficience de l’utilisation de l’eau élevée. Certains génotypes sont précoces, tels que 16, 25, 36, 39 et d’autres sont tardifs, tels que 29, 31, 32, 40, 42 et 43. Le second groupe renferme des génotypes sensibles au stress hydrique. Ils ont donné de faibles rendements en grains et biomasse aérienne et ont présenté une efficience de l’utilisation de l’eau réduite. Certains génotypes sont précoces, tels que 4, 19, 23, 45 et d’autres sont tardifs, tels que 17 et 41.
40En considérant l’axe 2 (Figure 4), deux groupes de génotypes peuvent être distingués. Le premier englobe des génotypes de haut rendement dont les graines sont de gros calibre et ont des indices de récolte élevés, tels que 9, 12, 13, 16, 25 et 29. Le second groupe est caractérisé par un rendement en grains élevé mais dont les graines sont de petit calibre, tels que 20, 38, 39, 40, 42 et 43.
4. Conclusion
41La mise en culture de la collection des 45 génotypes de pois chiche a montré une variabilité génotypique hautement significative pour tous les paramètres agronomiques et phénologiques étudiés. Elle indique une importante diversité biologique entre ces génotypes de pois chiches.
42Un groupe est constitué de génotypes fortement stressés par le manque d’eau et les températures relativement élevées. Certains sont précoces, tels que 4, 19, 23, 45 et d’autres sont tardifs, tels que 17 et 41. Ils ont donné de faibles rendements en grains et en biomasse aérienne et leur efficience d’utilisation de l’eau est réduite. L’aire de culture de ces génotypes n'est certainement pas en milieu semi-aride en culture de printemps. On peut supposer qu'elle serait plutôt limitée aux zones humides et sub-humides du territoire tunisien avec un semis d’hiver pour les génotypes qui ont une floraison tardive et un semis de printemps pour ceux qui ont une floraison précoce.
43Un second groupe est constitué de génotypes ayant toléré les stress hydrique et thermique. Ces génotypes ont produit de hauts rendements en grains et des biomasses aériennes élevées et ont une efficience de l’utilisation de l’eau élevée. Certains sont précoces et caractérisés par des graines de gros calibre, tels que les génotypes 16, 25 et 29. D’autres sont tardifs et caractérisés par des graines de petit calibre, tels que 20, 38, 39, 40, 42 et 43. Ces génotypes pourraient être conduits en culture de printemps dans le semi-aride tunisien.
44Le dernier groupe est composé du reste des génotypes qui sont du type 'Kabuli'. Ils sont moyennement affectés par les stress hydrique et thermique. Ils paraissent plus adaptés pour une culture d’hiver que pour une culture de printemps. Ils peuvent être conduits en culture d’hiver dans les zones du semi-aride supérieur avec des irrigations complémentaires.
45Ainsi, sous des conditions de stress hydrique, parmi les caractères agronomiques étudiés, le rendement en grains, l’efficience de l’utilisation de l’eau, la production de biomasse aérienne, l’indice de récolte, la date de floraison et le poids de 100 graines sont les plus utiles pour la caractérisation des génotypes de pois chiches tolérants au stress hydrique. Néanmoins, le poids de 100 graines parait un caractère génétique relié au type de pois chiche. En fait, le pois chiche du type 'Dési' est caractérisé par des graines de petit calibre et de faible poids de 100 graines, par comparaison au pois chiche du type 'Kabuli'. Afin de valoriser davantage les ressources génétiques et de mieux discerner les génotypes de pois chiche qui répondent aux exigences de l’agriculteur, d’autres techniques de criblage de génotypes tolérants au stress hydrique basées sur les marqueurs chimiques, biochimiques et moléculaires et intégrant d’autres critères agronomiques comme le calibre, la couleur et la qualité organoleptique des graines peuvent être utilisées.
46Remerciements
47Les auteurs remercient l’ICARDA (International Center for Agricultural Research in the Dry Areas) pour la fourniture du matériel végétal et Messaoud Mars, Maitre de Conférence à l’Institut supérieur agronomique de Chott Mariem, pour sa collaboration et sa contribution dans la révision du présent document.
Bibliographie
Abernethy R.H., 1987. Response of Cicer Milkvetch seed to osmoconditioning. Crop Sci., 27, 117-121.
Acevedo E. & Ceccarelli S., 1987. Role of the physiologist-breeder in a breeding program for drought resistance conditions. In: Baker F.W.G., ed. Drought resistance in cereals. Wallingford, UK: CAB International, 117-139.
Allen G., Pereira L., Raes D. & Smith M., 1998. Crop evapotranspiration: guidelines for computing crop water requirements. In: FAO Irrigation and drainage, paper n°24, http:www.fao.org/docrep/X0490E/x0490eob.htm, (08/12/07).
Aouani M.E., Mhamdi R., Jebara M. & Amarger N., 2001. Characterization of rhizobia nodulating chickpea in Tunisia. Agronomie, 21, 577-581.
Belhassen E., This D. & Monneveux P., 1995. L’adaptation génétique face aux contraintes de sècheresse. Cah. Agric., 4, 251-261.
Ben Mechlia N.B., 1998. Manuel de formation : application des données climatiques à la planification et à la gestion efficace de l’irrigation. Projet INAT - CGRE. Mise au point d’un système d’irrigation. In: Séminaire FAO/OMM/PNUE.
Berger J.D. et al., 2006. Genotype by environment studies demonstrate the critical role of phenology in adaptation of chickpea (Cicer arietinum L.) to high and low yielding environments of India. Field Crops Res., 98, 230-244.
Blum A.L, Golan G. & Mayer J., 1989. Yield stability and canopy temperature of wheat genotypes under drought stress. Field Crops Res., 22, 289-296.
Boubaker M. & Yamada T., 1995. Differential genotypic responses of string wheat early seedling growth to limited moisture conditions. Tropicultura, 13(2), 50-53.
Boubaker M. & Ben-Hammouda M., 1997. Screening durum wheat for drought tolerance at the seedling growth stage. Agric. Mediterr., 127(3), 267-274.
Bouzerzour H., Djekoune A., Benmhammed A. & Hassous L., 1998. Contribution de la biomasse aérienne, de l’indice de récolte et de la précocité au rendement en grain de l’orge (H. vulgare L.) en zone semi-aride d’altitude. Cah. Agric., 7, 307-317.
Choisnel E., 1984. Un modèle agrométéorologique opérationnel de bilan hydrique utilisant des données climatiques. In : Les besoins en eau des cultures. Versailles, France : Editions Inra, 116-132.
Ellis R.H. et al., 1994. Towards the reliable prediction of time to flowering in six annual crops. V. Chickpea. Exp. Agric., 30, 271-282.
Erchidi A.E., Benbella M. & Talouizte A., 2000. Relation entre certains paramètres contrôlant les pertes en eau et le rendement grain chez neuf variétés de blé dur soumises au stress hydrique. CIHEAM, Options Méditerranéennes, Séries Séminaires, 40, 279-282.
Frontier S., 1981. Méthode statistique. Applications à la biologie, la médecine et l’écologie. Paris : Masson.
Gate P., 1995. Ecophysiologie du blé de la plante à la culture. Cachan, France : Editions Tech & Doc, Lavoisier.
Habaieb H. & Masmoudi-Charfi C., 2003. Calcul des besoins en eau des principales cultures exploitées au nord de la Tunisie : estimation de l'évapotranspiration de référence par différentes formules empiriques (cas des régions de Tunis, Béja et Bizerte). Sècheresse, 14(4), 257-265.
Huang B. & Goo H., 2000. Root physiological characteristics associated with drought resistance in tall fescue cultivars. Crop Sci., 40, 196-203.
Jain K.S., Sharma H.L., Mehra R.B. & Khare J.P., 1991. Multiple correlation and regression analysis in lentil. Lens Newsl., 18, 11-13.
Kamel M., 1990. Winter chickpea: status and prospects. CIHEAM, Options Méditerranéennes, Séries Séminaires, 9, 145-150.
Kramer P.J., 1983. Water relations of plants. New York, USA: ICRISAT, Academic Press Inc.
Kuruvadi S., 1989. Stomatal frequency in bread wheat under irrigated and rainfed conditions. Rachis, 8, 22-28.
Malhotra M.C., 1998. Germplasm program legumes. Annual report. Aleppo, Syria: ICARDA.
Malhotra R.S. & Johansen C., 1996. Germplasm program legumes. Annual Report. Aleppo, Syria: ICARDA.
Malhotra R.S. & Saxena M.C., 2002. Strategies for overcoming drought stress in chickpea. Caravan ICARDA, 17.
Meadley J. & Milbour J., 1970. The growth of vining peas. II. The effect of density of planting. J. Agric. Sci., 74, 273-278.
MEAT (Ministère de l’Environnement et de l’Aménagement du Territoire), 2001. Communication initiale de la Tunisie à la Convention Cadre des Nations Unies sur les changements climatiques. Tunis : MEAT.
Miller P. et al., 2002. Growing chickpea in the Northern Great Plains. Montguide, 3.
Ofori I., 1996. Correlation and path-coefficient analysis of components of seed yield in Bambara groundnut (Vigna subterranea L.). Euphytica, 91, 103-107.
Pacucci G.C., Troccoli B. & Leoni B., 2006. Effect of supplementary irrigation on yield of chickpea genotypes in a Mediterranean climate. Agric. Eng. Int., 8.
Perez de la Vega M., 1996. Plant genetic adaptedness to climatic and edaphic environment. Euphytica, 92(1-2), 27-38.
Poirée M. & Ollier C., 1966. Irrigation. Les réseaux d’irrigation, théorie, technique et économie des arrosages. 3e éd. Paris : Editions Eyrolles.
Roberts E.H., Summerfiel R.J., Minchin F.R. & Haley P., 1980. Phenology of chickpea (Cicer arietinum L.) in contrasting aerial environments. Exp. Agric., 16, 343-360.
Rosielle A.A. & Frey K.J., 1975. Estimates of selection parameters associated with harvest index in oat lines derived from a bulk population. Euphytica, 24, 121-131.
Saccardo F. & Calcagno F., 1990. Consideration of chickpea plant ideotypes for spring and winter sawing. CIHEAM, Options Méditerranéennes, Séries Séminaires, 9, 35-41.
Sapra V.T., Hughes J.L. & Sharma G.C., 1975. Inheritance and physiological effects of stomatal frequency in Barley. Crop Sci., 11, 780-783.
Sarrafi A., Mentewab A. & Monneveux P., 1993. Variabilité génétique de la fluorescence chlorophyllienne chez les haploïdes doubles d’orge et son utilisation dans la sélection pour la résistance au stress hydrique. In: Monneveux P. & Ben Salem M. Tolérance à la sècheresse des céréales en zone méditerranéenne. Diversité génétique et amélioration variétale. Paris : Editions INRA, 396-402.
Saxena M.C., 1980. Recent advances in chickpea agronomy. In: Proceedings of the international workshop on chickpea improvement, 28 February-2 March 1979, ICRISAT, Hyderabad, India, 96-98.
Saxena M.C., 1987. Agronomy of chickpea. In: Saxena M.C. & Singh K.B., eds. The chickpea. Wallingford, UK: CAB International, 207-232.
Saxena N., Johansen P.C., Saxena M.C. & Silim S.N., 1993. Selection for drought and salinity resistance in cool-season food legumes. In: Singh K.B. & Saxena M.C., eds. Breeding for stress tolerance in cool-season food legumes. Chichester, UK: John Wiley & Sons, 245-270.
Serraj R. et al., 2003. Management of drought in ICRISAT cereal and legume mandate crops. In: Kijne J.W., Barker R. & Molden D., eds. Water productivity in agriculture: limits and opportunities for improvement. Wallingford, UK: CAB International, 127-144.
Silim S.N. & Saxena M.C., 1993. Adaptation of spring-sown chickpea to the Mediterranean basin. I. Response to moisture supply. Field Crops Res., 34, 12l-136.
Singh T.P., 1977. Harvest index in lentil. Euphytica, 26, 833-839.
Singh K.B., Bejiga G., Saxena M.C. & Sinh M., 1991. Transferability of selection indices from drought-free to drought-prone environments in chickpea. Int. Chickpea Newsl., 24, 19-22.
Singh K.B. et al., 1994. Current status and further strategy in breeding chickpea for resistance to biotic and abiotic stresses. Euphytica, 73, 137-149.
Skrypetz S., 2002. Pois chiches : situation et perspectives. Bull. Bimens., 15(16).
Slama F., 1998. Les cultures industrielles et les légumineuses à graines. Tunis : Centre universitaire de diffusion.
Soltani A., Ghassemi-Golezani K., Khooie F.R. & Moghaddem M., 1999. A simple model for chickpea growth and yield. Field Crops Res., 62, 213-224.
Summerfield R.J., Ellis R.H. & Craufurd P.Q., 1996. Phenological adaptation to cropping environment. From evaluation descriptors of times to flowering to the genetic characterisation of flowering responses to photoperiod and temperature. Euphytica, 92(1-2), 281-286.
Wang H. & Clarke J.M., 1993. Genotypic, intraplant and environmental variation in stomatal frequency and size in wheat. Can. J. Plant Sci., 73, 671-678.
Yousaf A., Haq M.A., Tahir G.R. & Ahmed N., 1999a. Effect of various morphological traits on chickpea yield under drought and normal field conditions. Pak. J. Biol. Sci., 2(3), 1071-1073.
Yousaf A. & Tahir G.R., 1999b. Correlation and regression studies in chickpea genotypes. Pak. J. Biol. Sci., 2, 318-319.
Pour citer cet article
A propos de : Kamel Ben Mbarek
Institut supérieur agronomique de Chott Mariem. TU-4042 Sousse (Tunisie). E-mail : benmbarekkamel@yahoo.fr
A propos de : Abdelhamid Boujelben
Institut supérieur agronomique de Chott Mariem. TU-4042 Sousse (Tunisie).
A propos de : Chérif Hannachi
Institut supérieur agronomique de Chott Mariem. TU-4042 Sousse (Tunisie).
A propos de : Mohsen Boubaker
Ecole supérieure d’Agriculture de Mograne. TU-1120 Zaghouan (Tunisie).
