


- Home
- Volume 14 (2010)
- numéro 3
- Revue bibliographique sur la restauration des pelouses calcicoles en Europe : contraintes rencontrées et solutions proposées


View(s): 4620 (51 ULiège)
Download(s): 296 (6 ULiège)

Revue bibliographique sur la restauration des pelouses calcicoles en Europe : contraintes rencontrées et solutions proposées

Editor's Notes
Reçu le 14 novembre 2008, accepté le 16 octobre 2009
Résumé
Les pelouses calcicoles, qui sont considérées comme des hotspots de biodiversité en Europe, ont subi un fort déclin durant le siècle dernier. Dès lors, il est apparu que la restauration de ces habitats s'imposait en tant que stratégie de conservation. La restauration consiste à recréer les conditions biotiques et abiotiques permettant aux espèces typiques de s'installer. En fonction du type de dégradation du milieu, plusieurs contraintes peuvent survenir lors de la restauration, comme l'enrichissement du sol, l'envahissement par des espèces compétitives ou des conditions de croissance difficiles pour les plantes. Afin de surmonter ces contraintes, la communauté scientifique a mis au point des techniques qui se sont révélées plus ou moins efficaces. Cet article a pour but de décrire les contraintes qui peuvent survenir lors de la restauration de pelouses calcicoles ainsi que d'exposer les techniques proposées dans la littérature scientifique afin de les surmonter. Il décrit aussi les objectifs spécifiques à la restauration des pelouses calcicoles en tenant compte de l'histoire de ces milieux et dans le contexte des changements globaux. Différents paramètres peuvent être pris en compte pour estimer le succès des campagnes de restauration. Là aussi, une analyse de la littérature scientifique était nécessaire afin d'aborder au mieux le monitoring des pelouses restaurées.
Abstract
The restoration of calcareous grasslands in Europe: a review of constraints and solutions. Calcareous grasslands that are considered as biodiversity hotspots in Europe, have suffered an important decline during the last century. In this context, restoration of suitable habitats has been promoted as a conservation strategy. Restoration consists in the recreation of favorable abiotic and biotic conditions that permit the recolonization by typical species. Depending on the degradations, several constraints can be encountered during restoration, such as soil enrichment, the presence of competitive species, or harsh conditions for plant growth. In order to overcome these constraints, scientists have proposed techniques that were demonstrated to be more or less efficient. In this article, the different constraints that can occur while restoring calcareous grasslands are described as well as the techniques proposed in the scientific literature. We also describe restoration objectives specific to calcareous grasslands, taking the history of these habitats into account and considering global change issue. Different parameters can be considered to value restoration success. Here again, an analysis of the scientific literature is needed in order to best assess restoration monitoring.
Table of content
1. Introduction
1Les pelouses calcicoles sont des écosystèmes qui sont parmi les plus riches d'Europe occidentale pour de nombreux groupes biologiques (Prendergast et al., 1993 ; Willems, 2001 ; Poschold et al., 2002 ; WallisDeVries et al., 2002 ; Piqueray et al., 2007). On y retrouve une grande richesse en espèces végétales et en insectes notamment (Delescaille et al., 1991). Il n'est pas rare de retrouver sur ces pelouses de 30 à 40 espèces de plantes supérieures par m2 et jusqu'à 80 espèces si l'on inclut les bryophytes (Bobbink et al., 1987a ; Dutoit et al., 1996b ; Hutchings et al., 2002). Toutefois, cette richesse exceptionnelle est actuellement fortement menacée. Le reboisement artificiel ou spontané, ainsi que l'amendement des sols, l'urbanisation et l'exploitation de carrières ont mené à une régression intense des surfaces de pelouses calcicoles dans toute l'Europe (Lee et al., 2001 ; Dutoit et al., 2004 ; Adriaens et al., 2006). Il en résulte que ces pelouses sont à présent réduites à de petits îlots isolés les uns des autres et qui ne garantissent plus la survie à long terme des espèces qui leur sont inféodées (Saunders et al., 1991). Dans ce contexte, il est apparu clairement que la restauration d'un réseau écologique, c'est-à-dire d'un agencement d'éléments écologiques favorables à la survie à long terme des populations d'espèces, s'imposait comme une stratégie importante pour la conservation de ces espèces typiques (Jongman et al., 2004). Cela demande de restaurer des surfaces d'habitat dégradées ou détruites, c'est-à-dire de diriger leur développement vers un écosystème cible en accélérant ou en outrepassant les stades intermédiaires de la succession (Jordan et al., 1987 ; Bakker et al., 1999). Deux conditions sont nécessaires au succès d'une restauration. Premièrement, il faut recréer et maintenir les conditions environnementales nécessaires au développement d'une pelouse calcicole. Deuxièmement, il faut que les espèces typiques de l'habitat s'installent sur les surfaces restaurées. Ces deux objectifs se heurtent tous les deux à un certain nombre de contraintes et de difficultés.
2L'objectif de cette synthèse est d'exposer ces contraintes et d'identifier les solutions que la recherche scientifique nous propose actuellement pour les surmonter. Bien que ce sujet ait déjà fait l'objet de plusieurs revues bibliographiques (Muller et al., 1998 ; Hutchings et al., 2002 ; Walker et al., 2004), les nombreuses publications récentes sur le sujet justifient de faire de nouveau le point sur nos connaissances en restauration de ces milieux. Par ailleurs, certains concepts récents comme la dette d'extinction ou les mécanismes de facilitation ont permis l'émergence de nouvelles techniques ou ont remis en question notamment les objectifs et les priorités liés à la restauration des pelouses calcicoles. C'est donc dans le but d'intégrer ces données récentes en écologie de la restauration que nous synthétisons dans cet article les connaissances actuelles sur la manière de restaurer les pelouses calcicoles.
2. Objectifs et planification de la restauration
3Avant de se lancer dans les actions de restauration à proprement parler, le gestionnaire doit se poser la question de l'objectif à atteindre. Idéalement, la restauration devrait aboutir à l'écosystème qui était présent avant la dégradation. Toutefois, il est rare que les données existent sur ce qui était présent avant cette dégradation, c'est donc un site ou un ensemble de sites voisins jugés en bon état qui serviront d'écosystème de référence (Perrow et al., 2002). Cela ne pose aucun problème si l'on considère que cet écosystème est le reflet de la gestion actuelle et qu'il est en équilibre avec celle-ci. Cependant, il a été montré que la composition en espèces d'un site dépend largement de l'histoire de ce site. Premièrement, la présence de certaines espèces peut être le reflet de l'utilisation ancienne du site, cet effet pouvant perdurer durant plus d'un siècle (Dutoit et al., 2004 ; Alard et al., 2005). Par ailleurs, suite à la fragmentation, la composition en espèces des pelouses et leur richesse se trouvera modifiée selon la théorie de la biogéographie insulaire (MacArthur et al., 1967). Toutefois, ces changements n'interviennent pas du jour au lendemain et un délai existe entre le moment où la structure spatiale de l'habitat est modifiée et celui où les espèces qui ne rencontrent plus les conditions propices à leur survie à long terme vont disparaitre, ce qui crée une dette d'extinction dans la communauté (Tilman et al., 1994). Une telle dette a déjà été mise en évidence au sein de pelouses semi-naturelles (Lindborg et al., 2004b ; Helm et al., 2006), mais pas dans tous les cas (Adriaens et al., 2006 ; Lindborg, 2007). La considération d'un habitat qui est un reflet du passé comme écosystème de référence est évidemment problématique, puisque l'on sait alors dès le départ que l'objectif qu'on se fixe n'est pas atteignable.
4En plus de considérer les effets de l'histoire du site, les gestionnaires du futur auront à prendre en compte les changements globaux dans la définition des objectifs de la restauration. Ils devront tout d'abord considérer les potentiels changements climatiques, au risque de restaurer un habitat qui n'est plus en équilibre avec son environnement climatique. Cet aspect est particulièrement délicat puisqu'il est difficile de prédire les changements climatiques à l'échelle locale (Lavendel, 2003). Ensuite, les dépôts d'azote atmosphérique, importants dans certaines régions d'Europe, sont eux aussi susceptibles d'entrainer des modifications dans les écosystèmes de référence et en voie de restauration (Stevens et al., 2004 ; Horswill et al., 2008). Il convient donc de ne pas les négliger tant au niveau de la définition des objectifs que de la gestion elle-même.
5Par ailleurs, la restauration ne doit pas s'appréhender uniquement au niveau de la tache d'habitat, mais aussi au niveau du paysage écologique. Bien que le choix des zones à restaurer soit souvent dépendant de contraintes socio-économiques, certains critères d'ordre écologique doivent entrer en compte quand cela est possible. Le but étant d'une part d'optimiser les résultats au niveau des taches d'habitat restaurées et d'autre part, que la restauration permette de réduire au mieux les effets de la fragmentation de l'habitat (Hobbs, 2002). En raison des capacités de dispersion limitées de nombreuses espèces des pelouses calcicoles, il convient de considérer dans le choix des parcelles à restaurer la présence de ces espèces soit sur la parcelle elle-même, soit dans le voisinage proche. Plusieurs auteurs ont souligné l'importance de la présence de sources de graines proches pour le succès de la restauration (Bakker et al., 1996 ; Kiefer et al., 1996 ; Pärtel et al., 1998 ; Willems et al., 1998). Dans le cas de taches d'habitat isolées, la connexion avec d'autres taches peut être améliorée par la création ou l'entretien de corridors écologiques (Hobbs, 2002). Ces corridors peuvent prendre la forme de différents éléments linéaires du paysage (bords de routes, lisières, etc.) où des populations peuvent se développer au moins transitoirement (Kiviniemi et al., 1999). Finalement, l'histoire des taches d'habitat et l'éventuelle dette d'extinction qui pèse sur ceux-ci est à prendre en considération, notamment pour la définition des priorités en termes de restauration.
3. Habitats restaurables
6La restauration des pelouses calcicoles peut être réalisée à partir des habitats issus de la dégradation de celles-ci, c'est-à-dire :
7– les forêts secondaires ou les plantations de résineux (Kiefer et al., 1996 ; Baba, 2003 ; Bisteau et al., 2005),
8– les habitats issus de l'agriculture intensive (Pywell et al., 2002 ; Walker et al., 2004 ; Kiehl et al., 2006a),
9– les anciennes carrières (Wheater et al., 1997 ; Cullen et al., 1998).
10Dans tous les cas, l'environnement de départ ne permet pas la réinstallation directe des espèces typiques des pelouses calcicoles. Il convient donc dans un premier temps de recréer les conditions abiotiques (principalement le niveau de fertilité) et biotiques (structure de végétation) nécessaires à l'établissement d'une pelouse riche en espèces.
4. Restauration des conditions abiotiques (niveau de fertilité)
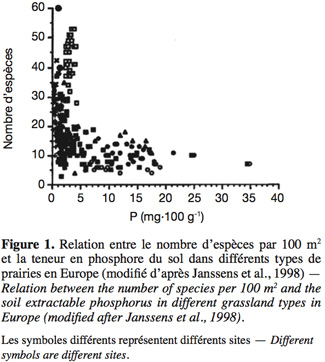
11La relation qui existe entre la productivité d'un milieu (ou les facteurs qui l'influencent comme la teneur en éléments minéraux du sol) et sa richesse en espèces est un concept d'intérêt majeur pour la conservation de la biodiversité, qui a déjà été largement étudié (Al-Mufti et al., 1977 ; Wheeler et al., 1991 ; Janssens et al., 1998 ; Loreau, 2000 ; Mittelbach et al., 2001 ; Jacquemyn et al., 2003). Bien que cette relation puisse varier selon le milieu considéré (Mittelbach et al., 2001), la teneur en éléments minéraux du sol est, dans de nombreux cas, un facteur limitant la richesse en espèces (Figure 1). Par ailleurs, il est généralement admis que l'enrichissement en azote et autres éléments minéraux est l'une des principales causes de perte de diversité pour de nombreux écosystèmes (Marrs, 1993 ; Janssens et al., 1998 ; Maron et al., 2001). Cet enrichissement entraine un changement dans les relations de compétition entre espèces, qui mène le plus souvent à la dominance de quelques espèces compétitives au détriment d'un grand nombre d'espèces qui le sont moins (Bakker, 1987 ; Marrs, 1993 ; Bobbink et al., 1998 ; Pywell et al., 2002) et participe ainsi à un phénomène plus global d'homogénéisation de la flore causé par les pratiques humaines (McKinney et al., 1999). Les pelouses calcicoles n'échappent pas à la règle. Historiquement, ces écosystèmes n'ont jamais été amendés et de grandes quantités de matière en étaient exportées par le pâturage itinérant. Il en résulte que ces milieux sont, par nature, pauvres en éléments nutritifs et de faible productivité (Grime, 1979 ; Critchley et al., 2002) et leur enrichissement entraine une perte importante de richesse spécifique (Willems et al., 1993 ; Jacquemyn et al., 2003). La restauration d'un niveau de fertilité adéquat est donc une condition importante pour l'établissement d'une pelouse riche en espèces.
12Il est évident que c'est lors de la restauration d'anciennes terres cultivées ou de prairies amendées que cette contrainte est la plus forte. Toutefois, Hurst et al. (1999a) ont montré qu'un enrichissement en azote du sol pouvait être enregistré durant les premiers stades d'enfrichement des pelouses par les graminées sociales. Toutefois, l'influence majeure de l'afforestation des pelouses calcicoles est plutôt un ralentissement de la minéralisation de la matière organique qui se résorbe rapidement suite à la réouverture du milieu (Piqueray et al., 2010). L'enrichissement en azote peut aussi se présenter comme un phénomène récurrent, sous la forme de dépôts atmosphériques d'oxydes d'azote ou de composés ammoniacaux (Bobbink et al., 1998).
13Au cours du temps, les sols ont tendance à s'appauvrir naturellement par lessivage des éléments nutritifs. Toutefois, ce phénomène est très lent (Marrs et al., 1991). Dès lors, différentes techniques permettant d'accélérer la réduction de la fertilité du sol ont été mises au point (Marrs, 2002 ; Walker et al., 2004).
14Une première technique consiste à faucher le site à restaurer et à en exporter la biomasse produite. Le principe sous-jacent étant d'enlever, par l'exportation, une quantité d'éléments plus grande que celle qui entre dans la parcelle (Marrs et al., 1998). Bien que l'idée semble cohérente du point de vue théorique, ses résultats sont le plus souvent modestes, que ce soit pour les pelouses calcicoles (Rizand et al., 1989 ; Kiehl et al., 2003) ou pour d'autres types de pelouses (Marrs et al., 1998 ; McCrea et al., 2001).
15Différents traitements du sol permettent de réduire les teneurs en éléments minéraux ou leur disponibilité. Parmi celles-ci, le traitement du sol par des composés adsorbants, tels que les oxydes et hydroxydes de fer et d'aluminium, a été proposé comme technique pour réduire la disponibilité du phosphore notamment. Les résultats liés à ces techniques sont relativement intéressants en termes de réduction de la concentration en phosphore dans les plantes (Walker et al., 2004) ou de phosphore soluble dans le sol (Janssens, 1998). Toutefois, leur utilisation peut poser un certain risque du fait de la toxicité des composés utilisés. Ces techniques sont donc à proscrire à proximité des zones sensibles. D'autres traitements moins risqués ont été proposés. L'ajout de matériel inerte (gravier, boues de carrières) permet de diluer les concentrations en éléments minéraux (Marrs, 1993). L'ajout de matériel riche en carbone (écorce, paille, sciure) permet, lui, d'augmenter l'activité microbienne et d'immobiliser ainsi une partie de l'azote contenu dans le sol (Blumenthal et al., 2003 ; Spiegelberger, 2006). Ces techniques, bien que potentiellement intéressantes, montrent toutefois des résultats contrastés (Blumenthal et al., 2003 ; Walker et al., 2004).
16Une autre technique, qui est aussi probablement la plus efficace à ce jour, consiste à enlever la couche superficielle du sol où la concentration en éléments minéraux est la plus forte. Dans de nombreux cas, cette technique a permis de réduire fortement la concentration d'azote et de phosphore dans le foin récolté sur les parcelles restaurées, ainsi que de diminuer la productivité du site (Hopkins et al., 1999 ; Walker et al., 2004). En conséquence, on peut observer, suite à ce traitement, une meilleure installation des espèces typiques des pelouses, bien que la richesse spécifique n'augmente pas forcément à court terme (Hopkins et al., 1999 ; Kiehl et al., 2007). À défaut d'enlever cette couche superficielle, un labour profond permet de la diluer dans les couches moins riches situées en profondeur (Pywell et al., 2002).
5. Restauration des conditions biotiques (structure de végétation)
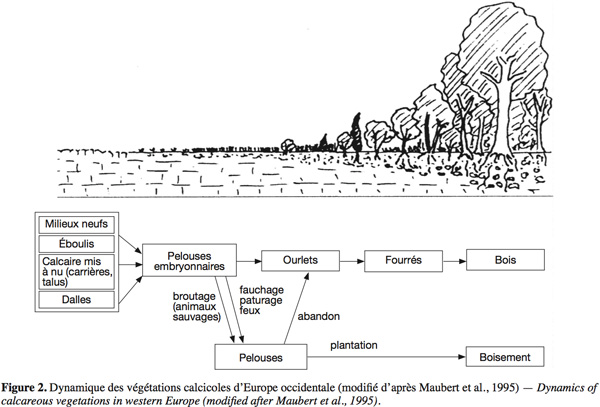
17Les pelouses calcicoles ne constituent pas, en Europe occidentale, un habitat climacique. Elles font partie d'une série dynamique de la végétation qui va de la roche calcaire mise à nu jusqu'à la forêt climacique, qui est le plus souvent la hêtraie calcicole dans nos régions, les pelouses calcicoles formant une voie détournée de la succession normale par le pâturage ou la fauche, autrement dit un plagioclimax (Green, 1972 ; Maubert et al., 1995 ; Dutoit et al., 1996a) (Figure 2). Il résulte de cela que les pelouses calcicoles peuvent être restaurées à partir des différents stades de cette série évolutive. Selon que l'état de départ correspond à un stade antérieur (dalle rocheuse, pelouse embryonnaire, par exemple dans les anciennes carrières) ou postérieur (fourré, forêt), il s'agira d'accélérer ou de faire régresser la succession pour obtenir l'habitat désiré (Prach et al., 2007). Ces manipulations de la succession font appel d'une part aux relations entre les plantes et le sol, qui ont déjà été abordées dans la section précédente, mais surtout aux relations plantes-plantes qu'elles soient intra- ou interspécifiques. Pendant de nombreuses années, l'étude de ces interactions s'est limitée à celle de la compétition pour les ressources. Ce n'est que récemment que la facilitation a été considérée comme une composante essentielle dans le fonctionnement des écosystèmes et de leur développement (Brooker et al., 2008). Ces deux pendants des interactions plantes-plantes ont des applications directes dans le cas de la restauration des pelouses calcicoles. La limitation de la compétition est primordiale pour restaurer des milieux envahis par les espèces ligneuses ou les graminées sociales. Par ailleurs, le renforcement des effets de facilitation peut être un outil très intéressant lors de la restauration de pelouses sur des sols nus.
18En effet, certaines activités humaines telles que l'exploitation de carrière ou la construction de voies de communication laissent des zones rocheuses totalement dépourvues de végétation. La recolonisation naturelle de ces zones dénudées mène souvent, dans un premier temps, à la formation de milieux semblables aux milieux semi-naturels issus des pratiques agropastorales anciennes, servant ainsi de refuge à des espèces et des communautés devenues rares (Cullen et al., 1998). Toutefois, ces milieux sont fortement hostiles (absence de sol, sècheresse, pauvreté en éléments nutritifs) pour l'installation et pour le développement des espèces végétales, ce qui rend le processus extrêmement lent et donc souvent incompatible avec les délais imposés pour la réhabilitation de ces sites (Wheater et al., 1997 ; Cullen et al., 1998). Dès lors, l'objectif principal de nombreux projets de réhabilitation écologique de carrières a été d'obtenir rapidement un couvert végétal indigène, mais pas forcément de recréer des communautés végétales riches en espèces (Dixon et al., 1984 ; Davis et al., 1985 ; Clemente et al., 2004). Pourtant, certaines techniques montrent un intérêt pour la restauration de pelouses calcicoles ou de communautés affines à partir d'anciennes carrières et ce, dans un délai raisonnable. Le remodelage du profil topographique à l'aide d'explosifs suivi d'un ensemencement permettent d'accélérer la restauration de communautés végétales des dalles rocheuses calcaires (Cullen et al., 1998). Davis et al. (1993) ont montré que l'application de faibles quantités d'engrais pouvait favoriser le développement des espèces cibles. Néanmoins, il s'agit de rester prudent car même dans des conditions extrêmement difficiles, l'application trop importante d'engrais peut empêcher le développement de pelouses riches en espèces (Mitchley et al., 1996). Enfin, les avancées récentes dans la connaissance des interactions plantes-plantes a permis la mise en place d'essais de nouvelles techniques de restauration (Brooker et al., 2008). L'utilisation d'espèces-nurses destinées à stabiliser le sol et/ou à créer des conditions micro-climatiques favorables pourrait se révéler comme un outil intéressant pour la création de pelouses à partir de la roche nue. Cette technique a déjà été utilisée avec plus ou moins de succès pour la restauration d'autres types de milieux (Castro et al., 2002 ; Gómez-Aparicio et al., 2004 ; Huber-Sannwald et al., 2005), mais rarement pour la restauration de pelouses calcicoles (Mitchley et al., 1996). Pourtant, il a été montré dans plusieurs études que la régénération d'un certain nombre d'espèces typiques des pelouses calcicoles est favorisée par la présence d'un couvert végétal et ce, d'autant plus que le milieu est xérique et peu productif (Hillier, 1990 ; Ryser, 1993). Dès lors, cette technique pourrait s'avérer intéressante dans le cadre de la restauration de pelouses calcicoles à partir de roches nues. Toutefois, leur utilisation n'est pas toujours simple et leurs effets demandent souvent plusieurs années avant de s'exprimer pleinement (Brooker et al., 2008).
19D'un autre côté, l'abandon progressif des pratiques agro-pastorales depuis la fin du 19e siècle a entrainé l'évolution de grandes surfaces de pelouses calcicoles vers des forêts secondaires. Trois phases peuvent être reconnues au cours de cette évolution : phase de colonisation par les graminées sociales (Brachypodium pinnatum, Bromus erectus) ; phase pré-forestière (colonisation par les arbustes Prunus spinosa, Crataegus monogyna, Corylus avellana, etc.) ; phase forestière dominée par des espèces arborées (Wells, 1969 ; Dutoit et al., 1996a ; Delescaille et al., 2004). La plantation artificielle de conifères (principalement Pinus nigra et Pinus sylvestris) dès le milieu du 19e siècle a contribué à amplifier ce phénomène (Vandermotten et al., 1995). Ces forêts naturelles et artificielles issues d'anciennes pelouses calcicoles présentent un certain potentiel pour la restauration de cet habitat. Dans ce cas, les techniques de restauration et de gestion post-restauration visent principalement à maitriser la compétition par les espèces ligneuses et les graminées sociales.
20La première phase de restauration de milieux forestiers consiste à déboiser ou débroussailler la parcelle à restaurer. Cette pratique n'a lieu qu'une fois, mais elle ne suffit pas à assurer la restauration d'une pelouse calcicole (Delescaille et al., 1990 ; Zobel et al., 1996 ; Dzwonko et al., 1998). En effet, les espèces ligneuses qui composent les formations à restaurer ont pour la plupart la capacité de rejeter vigoureusement de souche. Afin de réduire cette vigueur, une technique de brûlage des souches de buis (Buxus sempervirens) au chalumeau a été utilisée avec succès dans la vallée du Viroin (Belgique), mais le travail est assez contraignant, l'utilisation de ce matériel étant malaisée sur les terrains fort accidentés. Une autre possibilité consiste à réaliser un gyrobroyage des souches après la coupe, qui va réduire leur capacité de rejet. Toutefois, cette technique occasionne des dégâts à la végétation en place et ne peut donc être utilisée que si celle-ci ne présente pas d'intérêt conservatoire particulier (Delescaille, comm. pers.).
21En plus des rejets de ligneux, il importe de limiter ou de réduire l'envahissement par les graminées sociales. Différentes techniques existent, qui permettent de gérer les rejets ligneux et les graminées sociales conjointement. La fauche a été testée comme technique de gestion dans de nombreuses études (e.a. Wells, 1969 ; Green, 1980 ; Burger, 1984 ; Bobbink, 1991 ; Bobbink et al., 1993 ; Maron et al., 2001). Elle a principalement été utilisée pour le contrôle des graminées sociales, mais peut aussi servir pour éliminer les rejets de souches ligneux (Delescaille et al., 1991). Il ressort de ces études que les meilleurs résultats sont obtenus en fauchant plusieurs fois par an (Wells, 1969 ; Bobbink et al., 1993). Si cela n'est pas possible, il est préférable de réaliser la fauche au début de l'été (juin-juillet), au moment où les parties aériennes des plantes contiennent leur maximum de ressources, plutôt qu'en fin de saison (Green, 1980 ; Krüsi, 1981 ; Bobbink et al., 1987b ; 1991 ; Köhler et al., 2005). Dans la plupart des cas, le foin issu de la fauche est exporté (Bobbink et al., 1987b ; 1991 ; Köhler et al., 2005), afin de contribuer à l'appauvrissement du sol. Toutefois, Kahmen et al. (2002) n'ont pas pu montrer de différences notables entre des parcelles gérées avec ou sans exportation du foin et ce, sur une période de 25 ans. Lorsque la situation le permet (terrain peu accidenté et facile d'accès), ces actions de débroussaillage et de fauche peuvent être réalisées mécaniquement à l'aide d'engins tractés. Cela permet de réduire fortement les couts de main-d'œuvre liés à la gestion (Delescaille et al., 1995).
22Bien que les pelouses calcicoles de certaines régions soient issues de la fauche traditionnelle (Köhler et al., 2005), la plupart d'entre elles se sont formées et maintenues grâce au pâturage itinérant (Wells, 1969 ; Delescaille, 2002). C'est donc assez naturellement que des techniques de restauration et de gestion par pâturage ont été testées à travers toute l'Europe. La mise en place d'un système de gestion par pâturage se révèle être un exercice complexe.
23Premièrement, se pose la question du choix de l'espèce animale et de la race à utiliser pour le pâturage. Il est généralement admis que les moutons de race rustique ainsi que les chèvres sont les animaux les mieux adaptés pour la gestion des pelouses calcicoles (Dutoit et al., 1996a ; Delescaille, 2002). Les deux espèces sont très sélectives quant aux végétaux consommés. Si les moutons sont très efficaces pour le contrôle des graminées sociales, les chèvres montrent quant à elles une nette préférence pour les éléments ligneux. Il existe donc une certaine complémentarité entre les deux espèces en ce qui concerne la restauration des pelouses (Haumann, 1998 ; Colas et al., 2002 ; Delescaille, 2002 ; Tischew et al., 2008). Deuxièmement, il faut s'assurer que la charge de pâturage imposée permette de contrôler les ligneux et les graminées sociales sans pour autant en arriver à un surpâturage qui serait préjudiciable pour les espèces cibles. Cela ne laisse parfois qu'une marge de manœuvre très étroite (Dutoit et al., 1996b). Il est généralement préconisé d'appliquer une charge de l'ordre de cinq moutons par hectare et par an sur les pelouses nouvellement restaurées. Cette charge devra ensuite être ramenée progressivement à un ou deux moutons par hectare et par an, qui est la charge conseillée pour les pelouses à l'entretien (Dutoit et al., 1996b ; Verbeke et al., 1996 ; Delescaille, 2000 ; Colas et al., 2002). Toutefois, ce ne sont que des valeurs moyennes, la charge étant fonction de la productivité du milieu qui peut fortement varier d'une station à l'autre et d'une année à l'autre dans la même station (Colas et al., 2002). Troisièmement, il existe plusieurs modalités de gestion par pâturage. Idéalement, la gestion devrait se faire par pâturage itinérant, tel qu'il était pratiqué anciennement (Dutoit et al., 1996b). Cependant, les conditions socio-économiques actuelles ne permettent pas le plus souvent de réinstaurer le système de pâturage traditionnel. C'est donc la plupart du temps un système de pâturage en enclos qui est mis en place. Dans le cas des pelouses sèches, la meilleure alternative est le pâturage tournant, où une forte charge en bétail pâture la pelouse durant une durée assez courte. Cette technique permet en outre d'avoir une bonne maitrise de la pression de pâturage imposée (Verbeke et al., 1996).
24Bien que la fauche et le pâturage soient les techniques les plus couramment utilisées pour contrôler les rejets ligneux et les graminées sociales, d'autres techniques ont été envisagées telles que le feu (Dutoit et al., 1996a ; Verbeke et al., 1996 ; Köhler et al., 2005) ou l'utilisation d'herbicides (Hurst et al., 1999b). Globalement, ces techniques donnent des résultats au mieux médiocres et posent des problèmes éthiques, pratiques et de sécurité. Dès lors, elles sont peu recommandables, bien que le feu puisse s'avérer intéressant pour réouvrir les milieux fortement embroussaillés (Dutoit et al., 1996a).
25Comme cela a déjà été sous-entendu, il faut noter que, quel que soit le milieu de départ, les techniques de fauche ou de pâturage doivent être appliquées de manière récurrente sur les pelouses nouvellement créées afin d'assurer le maintien à long terme de celles-ci. Toutefois, dans le cas de restaurations à partir de sols rocheux, tel est le cas des anciennes carrières, il est connu que le pâturage peut freiner l'installation d'un couvert végétal (Davis et al., 1985 ; 1993). Dès lors, et bien que l'effet de la fauche sur ces milieux ne soit pas connu, il est préférable d'attendre que le couvert végétal soit comparable à celui des pelouses de référence avant d'y appliquer une gestion régulière.
6. Restauration de la communauté végétale
6.1. Introduction
26Une fois les conditions biotiques et abiotiques rétablies, il importe que le cortège d'espèces typiques puisse se reformer. Il arrive qu'un certain nombre d'espèces ait pu subsister dans l'habitat dégradé, parfois de manière peu visible. Pour les autres espèces, trois mécanismes peuvent intervenir dans ce processus de colonisation : l'expression de la banque de graines, la dispersion spatiale de propagules par voies naturelles et la dispersion spatiale, plus ou moins dirigée, de propagules par des actions anthropiques.
6.2. Persistance des espèces végétales dans les habitats dégradés
27Un certain nombre d'espèces végétales sont capables de survivre dans les habitats issus de la dégradation des pelouses calcicoles, en particulier dans les formations forestières ou pré-forestières qui se développent suite à l'abandon de celles-ci. Si une grande partie des espèces typiques des pelouses est capable de survivre dans des pelouses récemment abandonnées, très peu d'entre elles sont par contre capables de survivre sous un couvert forestier dense, citons toutefois Brachypodium pinnatum, Carex flacca et Viola hirta (Poschlod et al., 1998 ; Bisteau et al., 2005). Dans les premières années suivant la restauration, le développement végétatif de ces espèces persistantes va être un des mécanismes les plus actifs de recolonisation de l'espace restauré (Hofmans et al., 1990). Il résulte dès lors que l'état initial de la parcelle restaurée va avoir un impact important sur le succès de la restauration (Dzwonko et al., 1998).
6.3. Expression de la banque de graines
28La banque de graines est l'ensemble des graines et des fruits présents sur ou dans le sol (Leck et al., 1989). Sa composition en un endroit donné est globalement fonction de la composition historique de la végétation sus-jacente, de la longévité des graines, des stratégies de régénération de chaque espèce et des facteurs environnementaux influençant la persistance des graines (Harper, 1977 ; Thompson et al., 1979 ; Cavers et al., 1989). Pour que cette banque de graines soit utile à la reconstitution du cortège d'espèces typiques, il est donc nécessaire qu'une pelouse calcicole ait été présente historiquement à l'endroit ou à proximité du site restauré et que les graines des espèces cibles aient pu persister dans les conditions environnementales rencontrées avant la restauration de conditions favorables à leur germination. Certaines espèces caractéristiques des pelouses calcicoles sont connues pour répondre à ces conditions. C'est le cas par exemple de Carex flacca, Origanum vulgare, Euphorbia cyparissias et quelques Fabaceae. Toutefois, il a été montré à plusieurs reprises que la banque de graines des pelouses calcicoles était majoritairement transitoire (i.e. les graines ne survivent qu'un an dans le sol) et ne jouait donc qu'un rôle limité dans la reconstitution du cortège floristique typique (Dutoit et al., 1995 ; Hendry et al., 1995 ; Kiefer et al., 1996 ; Pärtel et al., 1998 ; Bakker et al., 1999 ; Bisteau et al., 2005 ; Bossuyt et al., 2006).
6.4. Dispersion spontanée des espèces cibles
29La pluie de graines, c'est-à-dire l'ensemble des graines atteignant un endroit donné suite à une dispersion spatiale, est considérée comme une contrainte importante pour la restauration des pelouses calcicoles et des milieux semi-naturels en général (Bakker et al., 1996 ; 1999 ; Pärtel et al., 1998). En effet, plusieurs études ont montré que la plupart des espèces typiques de ces milieux possédaient de faibles capacités de dispersion spontanée. Une étude menée par Verkaar et al. (1983) sur quatre espèces typiques des pelouses calcicoles a donné des capacités de dispersion spontanée de 0,3 à 3,5 m pour ces espèces. D'après Stampfli et al. (1999), peu d'espèces de ces pelouses sont capables de se disséminer à plus de 25 m et les échanges à longue distance ont parfois été considérés comme insignifiants (Maurer et al., 2003). Toutefois, ces études ne mettent en évidence que le caractère improbable des phénomènes de dispersion à longue distance ; si l'on considère un temps suffisamment long, de tels phénomènes peuvent se produire. Par exemple, Willems (2001) a pu observer l'apparition de Gentianella germanica sur une pelouse, 25 ans après sa restauration. Cette espèce, connue pour ses faibles capacités de dispersion (Verkaar et al., 1983), provenait probablement d'un site situé à 300 m de là et aurait été dispersée par des blaireaux. Récemment, des modèles ont été mis au point afin de permettre une approche plus quantitative et multi-espèces de la dispersion. De tels modèles existent pour la dispersion par le vent (Tackenberg et al., 2003) et par épizoochorie (Römermann et al., 2005). Une application récente de ce dernier modèle a permis de montrer que la dispersion par les mammifères joue un rôle tout à fait significatif dans la distribution des espèces des pelouses calcaires (Adriaens et al., 2007). Une telle application manque encore concernant la dispersion par le vent, mais il est certain que ces modèles pourraient remettre en cause une partie des conclusions sur le potentiel de dispersion de certaines espèces des pelouses calcicoles. Quoiqu'il en soit, il est un fait que beaucoup d'espèces typiques des pelouses calcicoles présentent des capacités de dispersion relativement faibles. Dès lors, la présence de sources de graines dans les environs immédiats du site restauré sera d'une importance considérable pour la reconstitution d'une communauté végétale typique à partir des pools d'espèces et de gènes locaux (Bakker et al., 1996 ; Kiefer et al., 1996 ; Pärtel et al., 1998 ; Willems et al., 1998).
6.5. Techniques permettant d'améliorer la disponibilité en graines
30Dans beaucoup de cas, les capacités de dispersion temporelle (banque de graines) et spatiale (pluie de graines) ne suffisent pas à restaurer complètement le cortège floristique après plusieurs années. Ce problème est d'autant plus important que les sites sont isolés et que les espèces typiques ne sont plus présentes dans leur voisinage immédiat (Pärtel et al., 1998 ; Lindborg et al., 2004a). En outre, il apparait souvent qu'un certain nombre d'espèces, notamment les espèces rares typiques des vieilles pelouses, ne parviennent pas à recoloniser les milieux réouverts rapidement (Kiefer et al., 1996 ; Zobel et al., 1996 ; Dzwonko et al., 1998 ; Lindborg et al., 2004a). Il arrive d'ailleurs que certaines espèces ne soient pas présentes dans la pluie de graines, malgré la présence de populations-sources à proximité (Bakker et al., 1996). Il en résulte que, dans le cas particulier de ces espèces ou dans le cas de sites isolés, il convient de mettre en place des mesures visant à améliorer la disponibilité en graines, voire à introduire volontairement des espèces.
31Une première manière d'améliorer la disponibilité en graines sur un site restauré est d'adopter un système de gestion par pâturage itinérant ou tournant. Les animaux, les moutons en particulier, sont capables de transporter un nombre très important de graines dans leur pelage (exozoochorie) ou leur tube digestif (endozoochorie) (Fischer et al., 1996 ; Mitlacher et al., 2002 ; Couvreur et al., 2004 ; Cosyns et al., 2005). Dès lors, dans un système de pâturage où les animaux passent alternativement sur des anciennes pelouses puis des pelouses restaurées, un grand nombre de graines d'espèces caractéristiques peut être transporté en provenance de populations-sources relativement éloignées (Mouissie et al., 2005). Les moutons sont particulièrement efficaces en ce qui concerne l'exozoochorie, puisque leur laine dense, bouclée et grasse permet même le transport de diaspores dépourvues de dispositif d'adhésion (Shmida et al., 1983 ; Fischer et al., 1996). Pour ces raisons, le pâturage tournant a souvent été considéré comme un outil important pour la restauration du cortège floristique des pelouses calcicoles (Gibson et al., 1987 ; Barbaro et al., 2001 ; Hellström et al., 2003). Dans un ordre d'idée similaire, quoique bien moins étudié, les activités humaines de gestion ou de loisirs sur les pelouses calcicoles peuvent être responsables de la dispersion d'espèces typiques de ces milieux, par fixation sur les vêtements ou les machines servant à la gestion (Clifford, 1956 ; Bakker et al., 1996 ; Strykstra et al., 1997).
32Un second type de méthode, plus ciblé que le précédent, consiste à transporter un pool de graines provenant d'une pelouse en bon état (site donneur) vers un site restauré. Différentes méthodes ont déjà été proposées. Premièrement, la transplantation peut se faire directement sous forme de graines. Ces graines peuvent soit provenir du commerce, soit être récoltées sur site à la main ou mécaniquement (machine à brosses, pompe à vide) afin de réduire les couts de main-d'œuvre, puis ressemées sur le site restauré (Stevenson et al., 1997 ; Riley et al., 2004 ; Edwards et al., 2007). Une seconde méthode, consistant à transplanter des mottes d'un site vers un autre, a été proposée par Pärtel et al. (1998). Toutefois, le succès de cette technique était assez limité. Enfin, une technique qui a été utilisée avec des résultats prometteurs est le transfert de foin. Elle consiste à faucher un site donneur le matin quand les végétaux sont mouillés et que les graines y restent colées, puis à répandre le foin fraichement coupé sur le site restauré (Jones et al., 1995 ; Kiehl et al., 2006b ; 2007 ; Edwards et al., 2007). Quelle que soit la technique utilisée, il est largement conseillé de transplanter du matériel végétal provenant de sites proches, afin de préserver l'intégrité du pool génétique régional des espèces et de profiter des adaptations locales (Stevenson et al., 1997 ; Jones et al., 1999 ; Smith et al., 2005). Cette remarque est surtout valable dans le cas de l'utilisation de graines provenant du commerce et dont l'origine est souvent incertaine.
33Dans le cas d'espèces particulièrement rares, ou présentant des difficultés à se réinstaller dans les milieux restaurés, des mesures particulières pourraient être prises afin d'introduire volontairement l'espèce dans un site donné (semis, transplantation de plantes, plantules, rhizomes, etc.). De telles méthodes nécessitent évidemment une connaissance approfondie de l'écologie de l'espèce cible. Ces méthodes ont déjà été testées sur plusieurs espèces (Gilfedder et al., 1997 ; Morgan, 1999). Une expérience de ce type a été tentée en Suisse sur Pulsatilla vulgaris, une espèce typique des pelouses calcicoles, par Pfeifer et al. (2002) dans le but de renforcer les populations de l'espèce. Plusieurs espèces ont pu être ainsi réintroduites sur des pelouses restaurées au Royaume-Uni (Hutchings et al., 2002).
7. Évaluation du succès et de la pérennité de la restauration
34L'évaluation est une étape délicate du processus de restauration, mais néanmoins tout à fait indispensable. La question se pose quant aux paramètres à prendre en compte lors de cette évaluation. Dans l'abcédaire sur l'écologie de la restauration (Society for Ecological Restoration International Science & Policy Working Group, 2004), les auteurs donnent toutefois les grands principes à intégrer pour une évaluation complète du succès de la restauration. D'après ce guide, neuf attributs de l'écosystème restauré doivent être pris en compte :
35– même diversité et même structure de communauté que les sites de référence,
36– présence d'espèces indigènes,
37– présence des groupes fonctionnels nécessaires pour une stabilité à long terme,
38– capacité de l'environnement physique de maintenir des populations reproductrices des espèces à sa stabilité,
39– fonctionnement normal de l'écosystème,
40– intégration dans le paysage écologique,
41– menaces potentielles éliminées ou réduites,
42– résilience aux perturbations naturelles,
43– maintien de l'écosystème de lui-même au même degré que son écosystème de référence.
44La plupart du temps, les limitations de temps et de budget ne permettent pas un monitoring aussi complet du succès de la restauration. Dans leur méta-analyse, Ruiz-Jean et al. (2005) ont mis en évidence que le succès de la restauration était actuellement basé sur des mesures de diversité (principalement plantes et arthropodes), de structure de végétation et de processus écologiques. Ils concèdent toutefois que la prise en compte de deux mesures relevant de chacun de ces trois paramètres constitue déjà une évaluation valable du succès de la restauration.
45Dans le cas de la restauration des milieux semi-naturels sans apport de graines dirigé, il serait cependant intéressant d'évaluer le fonctionnement des populations d'espèces nouvellement apparues sur les sites restaurés. En effet, en raison du caractère exceptionnel des phénomènes de dispersion à longue distance, il est probable que ces populations se sont formées à partir d'un nombre de graines très limité au départ. Dès lors, il résulte que ces populations peuvent présenter des effets dits « de fondation » diminuant leur résilience aux perturbations naturelles et aux changements globaux, ainsi qu'un risque accru de dépression de consanguinité pouvant entrainer une diminution du fitness de la population (Ridley, 1997 ; Society for Ecological Restoration International Science & Policy Working Group, 2004).
46Enfin, les pelouses calcicoles nécessitant un entretien récurrent, le succès de la restauration ne sera total que si un système d'entretien permanent est assuré suite à la campagne de restauration.
8. Conclusion et perspectives
47La restauration des milieux semi-naturels en général et des pelouses calcicoles en particulier est un problème complexe, mettant en œuvre les compétences issues de nombreuses disciplines scientifiques (écologie, pédologie, ingénieries biologique et civile, etc.). Les processus nécessaires à sa complète réussite sont encore méconnus, bien que ce ne soit peut-être qu'une question de temps (Zobel et al., 1996 ; Pärtel et al., 1998).
48Actuellement, la restauration des pelouses calcicoles a pour but principal de recréer, en partie, les paysages écologiques qui existaient par le passé. Cela implique de restaurer des pelouses dans les sites où elles existaient jadis. Cependant, certains auteurs ont insisté récemment sur l'opportunité que représentaient les milieux fortement anthropisés (bords de route, parcs des villes, etc.) pour y installer des espèces indigènes et sur l'intérêt biologique et sociologique de cette pratique (Simmons et al., 2007). La réflexion pourrait être poussée plus loin en proposant que, non seulement les espèces utilisées soient indigènes, mais qu'elles forment en plus des communautés végétales indigènes d'intérêt biologique. Les travaux réalisés jusqu'à présent en restauration de milieux semi-naturels nous ont donné les bases théoriques et pratiques qui permettraient de mener à bien un tel projet.
Bibliographie
Adriaens D., Honnay O. & Hermy M., 2006. No evidence of a plant extinction debt in highly fragmented calcareous grasslands in Belgium. Biol. Conserv., 133, 212-224.
Adriaens D., Honnay O. & Hermy M., 2007. Does retention potential affect the distribution of plant species in highly fragmented calcareous grasslands? Ecography, 30, 505-514.
Al-Mufti M.M. et al., 1977. A quantitative analysis of shoot phenology and dominance in herbaceous vegetation. J. Ecol., 65, 759-791.
Alard D. et al., 2005. Patterns of secondary succession in calcareous grasslands: can we distinguish the influence of former land uses from present vegetation data? Basic Appl. Ecol., 6, 161-173.
Baba W., 2003. Changes in the structure and floristic composition of the limestone grasslands after cutting trees and shrubs and mowing. Acta Soc. Bot. Pol., 72, 61-69.
Bakker J.P., 1987. Restoration of species-rich grassland after a period of fertilizer application. In: van Andel J., Bakker J.P. & Snaydon R.W., eds. Disturbance in grasslands: causes, effects and processes. Dordrecht, The Netherlands: Dr W. Junk Publishers, 185-200.
Bakker J.P. et al., 1996. Seed bank and seed dispersal: important topics in restoration ecology. Acta Bot. Neerl., 45, 461-490.
Bakker J.P. & Berendse F., 1999. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends Ecol. Evol., 14, 63-68.
Barbaro L., Dutoit T. & Cozic P., 2001. A six-year experimental restoration of biodiversity by shrub-clearing and grazing in calcareous grasslands of the French prealps. Biodivers. Conserv., 10, 119-135.
Bisteau E. & Mahy G., 2005. Vegetation and seed bank in a calcareous grassland restored from a Pinus forest. Appl. Veg. Sci., 8, 167-174.
Blumenthal D.M., Jordan N.R. & Russelle M.P., 2003. Soil carbon addition controls weeds and facilitates prairie restoration. Ecol. Appl., 13, 605-615.
Bobbink R., 1991. Effects of nutrient enrichment in Dutch chalk grassland. J. Appl. Ecol., 28, 28-41.
Bobbink R. et al., 1987a. Effects of selective clipping and mowing time on species diversity in chalk grassland. Folia Geobotanica Phytotaxonomica, 22, 363-376.
Bobbink R. & Willems J.H., 1987b. Increasing dominance of Brachypodium pinnatum (L.) Beauv. in chalk grasslands: a threat to a species-rich ecosystem. Biol. Conserv., 40, 301-314.
Bobbink R. & Willems J.H., 1991. Impact of different cutting regimes on the performance of Brachypodium pinnatum in Dutch chalk grassland. Biol. Conserv., 56, 1-21.
Bobbink R. & Willems J.H., 1993. Restoration management of abandoned chalk grassland in the Netherlands. Biodivers. Conserv., 2, 616-626.
Bobbink R., Hornung M. & Roelefs J.G.M., 1998. The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation. J. Ecol., 86, 717-738.
Bossuyt B., Butaye J. & Honnay O., 2006. Seed bank composition of open and overgrown calcareous grassland soils: a case study from Southern Belgium. J. Environ. Manage., 79, 364-371.
Brooker R.W. et al., 2008. Facilitation in plant communities: the past, the present, and the future. J. Ecol., 96, 18-34.
Burger R., 1984. Successional limestone grassland communities of the Kaiserstuhl, with regard to their conservation management. In : La végétation des pelouses calcicoles, 1982, Strasbourg. Colloques Phytosociologiques, 11. Vaduz : J. Cramer, 405-420.
Castro J., Zamora R., Hódar J.A. & Gómez J.M., 2002. Use of shrubs as nurse plants: a new technique for reforestation in Mediterranean mountains. Restor. Ecol., 10, 297-305.
Cavers P.B. & Benoit D.L., 1989. Seeds in arable land. In: Leck M.A., Parker V.T. & Simpson R.L., eds. Ecology of soil seed bank. San Diego: Academic Press, 309-328.
Clemente A.S. et al., 2004. Restoration of a limestone quarry: effects of soil amendments on the establishment of native Mediterranean sclerophyllous shrubs. Restor. Ecol., 12, 20-28.
Clifford H.T., 1956. Seed dispersal on footwear. Proc. Bot. Soc. Br. Isles, 2, 129-131.
Colas S., Muller F., Meuret M. & Agreil C., 2002. Pâturage sur pelouses sèches : un guide d'aide à la mise en oeuvre. Orléans, France : Espaces naturels de France, Fédération des Conservatoires d'Espaces Naturels.
Cosyns E., Claerbout S., Lamoot I. & Hoffmann M., 2005. Endozoochorous seed dispersal by cattle and horse in a spatially heterogeneous landscape. Plant Ecol., 178, 149-162.
Couvreur M., Christiaen B., Verheyen K. & Hermy M., 2004. Large herbivores as mobile links between isolated nature reserves through adhesive seed dispersal. Appl. Veg. Sci., 7, 229-236.
Critchley C.N.R. et al., 2002. Plant species richness, functional type and soil properties of grasslands and allied vegetation in English environmentally sensitive areas. Grass Forage Sci., 57, 82-92.
Cullen W.R., Wheater C.P. & Dunleavy P.J., 1998. Establishment of species-rich vegetation on reclaimed limestone quarry faces in Derbyshire, UK. Biol. Conserv., 84, 25-33.
Davis B.N.K., Lakhani K.H., Brown M.C. & Park D.G., 1985. Early seral communities in a limestone quarry: an experimental study of treatment effects on cover and richness of vegetation. J. Appl. Ecol., 22, 473-490.
Davis B.N.K., Lakhani K.H. & Brown M.C., 1993. Experiments effects of fertilizer and rabbit grazing treatments upon the vegetation of a limestone quarry floor. J. Appl. Ecol., 30, 615-628.
Delescaille L.M., 2000. La gestion conservatoire de la pelouse calcicole du Chamousia à Vierves-sur-Viroin (Province de Namur, Belgique) : comparaison de la fauche hivernale et du pâturage ovin en saison de végétation sur la structure et la composition du tapis végétal. Parcs Réserves, 55, 2-9.
Delescaille L.M., 2002. Nature conservation and pastoralism in Wallonia. In: Redecker B. et al., eds. Pasture landscapes and nature conservation. Berlin, Deutschland: Springer-Verlag, 39-52.
Delescaille L.M., Hofmans K. & Maernoudt A., 1990. La pelouse de la Roche Trouée à Nismes (Province de Namur, Belgique) : premiers résultats de la gestion. Nat. Belg., 71, 157-175.
Delescaille L.M., Hofmans K. & Woué L., 1991. Les réserves naturelles du Viroin : trente années d'action d'Ardenne et Gaume dans la vallée du Viroin. Parcs Nat., 46, 4-68.
Delescaille L.M., Dewitte T. & Hofmans K., 1995. La gestion mécanisée des pelouses sèches. Parcs Nat., 50, 13-19.
Delescaille L.M., Henry A. & Hofmans K., 2004. Les pelouses calcicoles : un écosystème en mutation constante. In : Colmant L. et al. Les pelouses calcicoles en région wallonne. Vierves-sur-Viroin, Belgique : Entente nationale pour la Protection de la Nature, 44-52.
Dixon J.M. & Hambler D.J., 1984. An experimental approach to the reclamation of a limestone quarry floor; the first three years. Environ. Conserv., 11, 19-28.
Dutoit T. & Alard D., 1995. Permanent seed banks in chalk grassland under various management regimes: their role in the restoration of species-rich plant communities. Biodivers. Conserv., 4, 939-950.
Dutoit T. & Alard D., 1996a. Les pelouses calcicoles du nord-ouest de l'Europe (Brometalia erecti Br.Bl. 1936) : analyse bibliographique. Écologie, 27, 5-34.
Dutoit T. & Alard D., 1996b. Restauration d'un système de parcours sur les pelouses calcicoles de la vallée de la Seine (Haute-Normandie, France). In: Actes du Colloque international La gestion des pelouses calcicoles, 28-31 mai 1996, Cercle des Naturalistes de Belgique, Vierves-sur-Viroin, Belgique. Vierves-sur-Viroin, Belgique : Cercle des Naturalistes de Belgique, 47-54.
Dutoit T., Buisson E., Roche P. & Alard D., 2004. Land use history and botanical changes in the calcareous hillsides of Upper-Normandy (north-western France): new implications for their conservation management. Biol. Conserv., 115, 1-19.
Dzwonko Z. & Loster S., 1998. Dynamics of species richness and composition in a limestone grassland restored after tree cutting. J. Veg. Sci., 9, 387-394.
Edwards A.R. et al., 2007. Hay strewing, brush harvesting of seed and soil disturbance as tools for the enhancement of botanical diversity in grasslands. Biol. Conserv., 134, 372-382.
Fischer S.F., Poschlod P. & Beinlich B., 1996. Experimental studies on the dispersal of plants and animals on sheep in calcareous grasslands. J. Appl. Ecol., 33, 1206-1222.
Gibson C.W.D., Watt T.A. & Brown V.K., 1987. The use of sheep grazing to recreate species-rich grassland from abandoned arable land. Biol. Conserv., 42, 165-183.
Gilfedder L., Kirkpatrick J.B. & Wells S., 1997. The endangered Tunbridge buttercup (Ranunculus prasinus): ecology, conservation status and introduction to the Township Lagoon Nature Reserve, Tasmania. Aust. J. Ecol., 22, 347-351.
Gómez-Aparicio L. et al., 2004. Applying plant facilitation to forest restoration: a meta-analysis of the use of shrubs as nurse plants. Ecol. Appl., 14, 1128-1138.
Green B.H., 1972. The relevance of seral eutrophication and plant competition to the management of successional communities. Biol. Conserv., 4, 378-384.
Green B.H., 1980. Management of extensive amenity grasslands by mowing. In: Rorison I.H. & Hunt R., eds. Amenity grassland: an ecological perspective. Chichester, UK: Wiley, 155-161.
Grime J.P., 1979. Plant strategies and vegetation processes. Chichester, UK: Wiley.
Harper J.L., 1977. Population biology of plants. New York, NY, USA: Academic Press.
Haumann P., 1998. Biotope conservation with ruminants in Germany: the example of goats on shrub-infested slopes. In: Proceedings of the 2nd International Conference of the LSIRD network, 3-5 December 1998, Bray, Dublin.
Hellström K. et al., 2003. Use of sheep grazing in the restoration of semi-natural meadows in northern Finland. Appl. Veg. Sci., 6, 45-52.
Helm A., Hanski I. & Pärtel M., 2006. Slow response of plant species richness to habitat loss and fragmentation. Ecol. Lett., 9, 72-77.
Hendry G.A.F., Thompson K. & Band S.R., 1995. Seed survival and persistence on a calcareous land surface after a 32-years burial. J. Veg. Sci., 6, 153-156.
Hillier S.H., 1990. Gaps, seed bank and plant species diversity in calcareous grasslands. In: Hillier S.H., Walton D.W.H. & Wells D.A., eds. Calcareous grasslands: ecology and management. Huntington, UK: Bluntisham.
Hobbs R.J., 2002. The ecological context: a landscape perspective. In: Perrow M.R. & Davy A.J., eds. Handbook of ecological restoration. Principles of restoration. Cambridge, UK: Cambridge University Press, 24-45.
Hofmans K. & Delescaille L.M., 1990. La gestion des buxaies thermophiles (Helleboro-Buxetum) en Belgique : l'exemple de la Montagne-aux-Buis à Nismes-Dourbes (Province de Namur). In : Actes du colloque Gérer la nature ?, 17-20 octobre 1989, Anseremme, Belgique. Anseremme, Belgique : Région wallonne, DGRNE, 529-568.
Hopkins A. et al., 1999. Enhancement of botanical diversity of permanent grassland and impact on hay production in environmentally sensitive areas in the UK. Grass Forage Sci., 54, 163-173.
Horswill P. et al., 2008. Base cation depletion, eutrophication and acidification of species-rich grasslands in response to long-term simulated nitrogen deposition. Environ. Pollut., 155, 336-349.
Huber-Sannwald E. & Pyke D.A., 2005. Establishing native grasses in a big sagebrush-dominated site: an intermediate restoration step. Restor. Ecol., 13, 292-301.
Hurst A. & John E., 1999a. The biotic and abiotic changes associated with Brachypodium pinnatum dominance in chalk grassland in south-east England. Biol. Conserv., 88, 75-84.
Hurst A. & John E., 1999b. The effectiveness of glyphosate for controlling Brachypodium pinnatum in chalk grassland. Biol. Conserv., 89, 261-265.
Hutchings M.J. & Stewart A.J.A., 2002. Calcareous grasslands. In: Perrow M.R. & Davy A.J., eds. Handbook of ecological restoration. Restoration in practice. Cambridge, UK: Cambridge University Press, 419-442.
Jacquemyn H., Brys R. & Hermy M., 2003. Short-term effects of different management regimes on the response of calcareous grassland vegetation to increased nitrogen. Biol. Conserv., 111, 137-147.
Janssens F., 1998. Restauration des couverts herbacés riches en espèces. Thèse de doctorat : Laboratoire d'Écologie des Prairies, Université catholique de Louvain (Belgique).
Janssens F. et al., 1998. Relationship between soil chemical factors and grassland diversity. Plant Soil, 202, 69-78.
Jones A.T. & Hayes M.J., 1999. Increasing floristic diversity in grassland: the effects of management regime and provenance on species introduction. Biol. Conserv., 87, 381-390.
Jones G.H., Trueman I.C. & Millett P., 1995. The use of hay strewing to create species-rich grasslands (i) general principles and hay strewing versus seed mixes. Land Contam. Recl., 3, 104-107.
Jongman R.H.G. & Pungetti G., eds., 2004. Ecological networks and greenways: concept, design, implementation. Cambridge, UK: Cambridge University Press.
Jordan W.R.I., Gilpin M.E. & Aber J.D., eds., 1987. Restoration ecology. A synthetic approach to ecological research. Cambridge, UK: Cambridge University Press.
Kahmen S., Poschlod P. & Schreiber K.F., 2002. Conservation management of calcareous grasslands. Changes in plant species composition and response of functional traits during 25 years. Biol. Conserv., 104, 319-328.
Kiefer S. & Poschlod P., 1996. Restoration of fallow or afforested calcareous grassland by clear-cutting. In: Settele J., Margules C.R., Poschlod P. & Henle K., eds. Species survival in fragmented landscapes. Dordrecht, The Netherlands: Kluwer Academic Publishers, 209-218.
Kiehl K., Thormann A. & Pfadenhauer J., 2003. Nährstoffdynamik und Phytomasseproduktion in neu angelegten Kalkmagerrasen auf ehemaligen Ackerflächen. In: Pfadenhauer J. & Kiehl K., eds. Renaturierung von Kalkmagerrasen. Angewandte Landschaftsökologie, 55, 39-71.
Kiehl K., Thormann A. & Pfadenhauer J., 2006a. Evaluation of initial restoration measures during the restoration of calcareous grasslands on former arable fields. Restor. Ecol., 14, 148-156.
Kiehl K. & Wagner C., 2006b. Effect of hay transfer on long-term establishment of vegetation and grasshoppers on former arable fields. Restor. Ecol., 14, 157-166.
Kiehl K. & Pfadenhauer J., 2007. Establishment and persistence of target species in newly created calcareous grasslands on former arable fields. Plant Ecol., 189, 31-48.
Kiviniemi K. & Eriksson O., 1999. Dispersal recruitment and site occupancy of grassland plants in fragmented habitats. Oikos, 86, 241-253.
Köhler B. et al., 2005. Changes in species composition and conservation value of limestone grasslands in Northern Switzerland after 22 years of contrasting managements. Evol. Syst., 7, 51-67.
Krüsi B., 1981. Phenological methods in permanent plot research. Veröffentlichungen Geobotanisches Institut Rübel, 75, 1-116.
Lavendel B., 2003. Ecological restoration in the face of global climate change: obstacles and initiatives. Ecol. Rest., 21, 199-203.
Leck M.A., Parker V.T. & Simpson R.L., 1989. Ecology of soil seed bank. San Diego: Academic Press.
Lee J.T., Woddy S.J. & Thompson S., 2001. Targeting sites for conservation: using a patch-based ranking scheme to assess conservation potential. J. Environ. Manage., 61, 367-380.
Lindborg R., 2007. Evaluating the distribution of plant life-history traits in relation to current and historical landscape configurations. J. Ecol., 95, 555-564.
Lindborg R. & Eriksson O., 2004a. Effects of restoration on plant species richness and composition in Scandinavian semi-natural grasslands. Restor. Ecol., 12, 318-326.
Lindborg R. & Eriksson O., 2004b. Historical landscape connectivity affects present plant species diversity. Ecology, 85, 1840-1845.
Loreau M., 2000. Biodiversity and ecosystem functioning: recent theorical advances. Oikos, 91, 3-17.
MacArthur R.H. & Wilson E.O., 1967. The theory of Island biogeography. Princeton, NJ, USA: Princeton University Press.
Maron J.L. & Jefferies R.L., 2001. Restoring enriched grasslands: effects of mowing on species richness, productivity and nitrogen retention. Ecol. Appl., 11, 1088-1100.
Marrs R.H., 1993. Soil fertility and nature conservation in Europe: theorical considerations and practical management solutions. In: Begon M. & Fitter A.H., eds. Advances in ecological research 24. London, UK: Harcourt Brace & Company, 241-300.
Marrs R.H., 2002. Manipulating the chemical environment of the soil. In: Perrow M.R. & Davy A.J., eds. Handbook of ecological restoration. Principles of restoration. Cambridge, UK: Cambridge University Press, 155-183.
Marrs R.H., Gough M.W. & Griffiths M., 1991. Soil chemistry and leaching losses of nutrients from semi-natural grassland and arable soils on three contrasting parent materials. Biol. Conserv., 57, 257-271.
Marrs R.H., Snow C.S.R., Owen K.M. & Evans C.E., 1998. Heathland and acid grassland creation on arable soils at Minsmere: identification of potential problems and a test of cropping to impoverish soils. Biol. Conserv., 85, 69-82.
Maubert P. & Dutoit T., 1995. Connaître et gérer les pelouses calcicoles. Cahier ATEN. Montpellier, France : Ministère de l'Environnement.
Maurer K., Durka W. & Stöcklin J., 2003. Frequency of plant species in remnants of calcareous grassland and their dispersal and persistence characteristics. Basic Appl. Ecol., 4, 307-316.
McCrea A.R., Trueman I.C. & Fullen M.A., 2001. A comparison of the effects of four arable crops on the fertility depletion of a sandy silt loam destined for grassland habitat creation. Biol. Conserv., 97, 181-187.
McKinney M.L. & Lockwood J.L., 1999. Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol. Evol., 14, 450-453.
Mitchley J., Buckley G.P. & Helliwell D.R., 1996. Vegetation establishment on chalk marl spoil: the role of nurse grass species and fertiliser application. J. Veg. Sci., 7, 543-548.
Mitlacher K., Poschlod P., Rosén E. & Bakker J.P., 2002. Restoration of wooded meadows - a comparative analysis along a chronosequence on Öland (Sweden). Appl. Veg. Sci., 5, 63-73.
Mittelbach G.G. et al., 2001. What is the relationship between species richness and productivity? Ecology, 82, 2381-2396.
Morgan J.W., 1999. Have tubestock plantings successfully established populations of rare grassland species into reintroduction sites in western Victoria? Biol. Conserv., 89, 235-243.
Mouissie A.M., Lengkeek W. & van Diggelen R., 2005. Estimating adhesive seed-dispersal distances: field experiments and correlated random walks. Funct. Ecol., 19, 478-486.
Muller S., Dutoit T., Alard D. & Grévilliot F., 1998. Restoration and rehabilitation of species-rich grassland ecosystems in France: a review. Restor. Ecol., 6, 94-101.
Pärtel M., Kalamees R., Zobel M. & Rosén E., 1998. Restoration of species-rich limestone grassland communities from overgrown land: the importance of propagule availability. Ecol. Eng., 10, 275-286.
Perrow M.R. & Davy A.J., 2002. Handbook of ecological restoration. Cambridge, UK: Cambridge University Press.
Pfeifer E., Holderegger R., Matthies D. & Rutishauser R., 2002. Populations biologische Untersuchungen an einer Flaggschiff-Art der Magerrasen: Pulsatilla vulgaris Mill. in der Nordostschweiz. Bot. Helv., 112, 153-171.
Piqueray J., Bisteau E., Bottin G. & Mahy G., 2007. Plant communities and species richness of the calcareous grasslands in southeast Belgium. Belg. J. Bot., 140, 157-173.
Piqueray J. et al., 2010. Rapid restoration of a species-rich ecosystem assessed from soil and vegetation indicators: the case of calcareous grasslands restored from forest stands. Ecol. Indic., sous presse.
Poschlod P. et al., 1998. Plant species richness in calcareous grasslands as affected by dispersability in space and time. Appl. Veg. Sci., 1, 75-90.
Poschlod P. & WallisDeVries M.F., 2002. The historical and socioeconomic perspective of calcareous grasslands: lessons from the distant and recent past. Biol. Conserv., 104, 361-376.
Prach K., Marrs R.H., Pysek P. & van Diggelen R., 2007. Manipulation of succession. In: Walker L.R., Walker J. & Hobbs R.J., eds. Linking restoration and ecological succession. New York, NY, USA: Springer, 121-149.
Prendergast J.R. et al., 1993. Rare species, the coincidence of diversity hotspots and conservation strategies. Nature, 365, 335-337.
Pywell R.F. et al., 2002. Restoration of species-rich grassland on arable land: assessing the limiting processes using a multi-site experiment. J. Appl. Ecol., 39, 294-309.
Ridley M., 1997. Évolution génétique. Paris : DeBoeck Université.
Riley J.D., Craft I.W., Rimmer D.L. & Smith R.S., 2004. Restoration of magnesian limestone grassland: optimizing the time for seed collection by vacuum harvesting. Restor. Ecol., 12, 311-317.
Rizand A., Marrs R.H., Gough M.W. & Wells T.C.E., 1989. Long-term effects of various conservation management treatments on selected soil properties of chalk grassland. Biol. Conserv., 49, 105-112.
Römermann C., Tackenberg O. & Poschlod P., 2005. How to predict attachment potential of seeds to sheep and cattle coat from simple morphological seed traits. Oikos, 110, 219-230.
Ruiz-Jean M.C. & Aide T.M., 2005. Restoration success: how is it being measured. Restor. Ecol., 13, 569-577.
Ryser P., 1993. Influences of neighbouring plants on seedling establishment in limestone grassland. J. Veg. Sci., 4, 195-202.
Saunders D.A., Hobbs J.A. & Margules C.R., 1991. Biological consequences of ecosystem fragmentation: a review. Conserv. Biol., 5, 18-32.
Shmida A. & Ellner S., 1983. Seed dispersal on pastoral grazers in open Mediterranean chaparral. Isr. J. Bot., 32, 147-159.
Simmons M.T., Venhaus H.C. & Windhager S., 2007. Exploiting the attributes of regional ecosystems for landscape design: the role of ecological restoration in ecological engineering. Ecol. Eng., 30, 201-205.
Smith B.M., Diaz A., Winder L. & Daniels R., 2005. The effect of provenance on the establishment and performance of Lotus corniculatus L. in a re-creation environment. Biol. Conserv., 125, 37-46.
Society for Ecological Restoration International Science & Policy Working Group, 2004. The SER International Primer on Ecological Restoration. SER & Tucson, http://www.ser.org, (06.03.08).
Spiegelberger T., 2006. Land use, soil nutrient availability and conservation of biodiversity on mountain grasslands. PhD Thesis: Mathematisch-Naturwissenschaftlichen Fakultät, Universität Freiburg (Deutschland).
Stampfli A. & Zeiter M., 1999. Plant species decline due to abandonment of meadows cannot easily be reverse by mowing: a case study from the southern Alps. J. Veg. Sci., 10, 151-164.
Stevens C.J., Dise N.B., Mountford J.O. & Gowing D.J.G., 2004. Impact of nitrogen deposition on the species richness of grasslands. Science, 303, 1876-1879.
Stevenson M.J., Ward L.K. & Pywell R.F., 1997. Re-creating semi-natural communities: vacuum harvesting and hand collection of seed on calcareous grasslands. Restor. Ecol., 5, 66-76.
Strykstra R.J., Verweij G.L. & Bakker J.P., 1997. Seed dispersal by mowing machinery in a Dutch brook valley system. Acta Bot. Neerl., 46, 387-401.
Tackenberg O., Poschlod P. & Bonn S., 2003. Assessment of wind dispersal potential in plant species. Ecol. Monogr., 73, 191-205.
Thompson K. & Grime J.P., 1979. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. J. Ecol., 67, 893-921.
Tilman D., May R.M., Lehman C.L. & Nowak M.A., 1994. Habitat destruction and the extinction debt. Nature, 371, 65-66.
Tischew S., Mann S., Schecher K. & Jäger U., 2008. Restoration of fallow dry grasslands in cooperation with local farmers. In: 6th European Conference on Ecological Restoration, 8-12 September 2008, Ghent, Belgium.
Vandermotten C. & Decroly J.M., 1995. Géographie. In: Joris F., ed. Wallonie. Atouts et références d'une région. Namur, Belgique : Gouvernement wallon, 463.
Verbeke W. & Lejeune M., 1996. Pâturage ou fauchage, la meilleure gestion pour la flore des pelouses calcicoles ? In : Actes du Colloque international La gestion des pelouses calcicoles, 28-31 mai 1996, Cercle des Naturalistes de Belgique, Vierves-sur-Viroin, Belgique. Vierves-sur-Viroin, Belgique : Cercle des Naturalistes de Belgique, 55-59.
Verkaar H.J., Schenkeveld A.J. & Van de Klashorst M.P., 1983. The ecology of short-lived forbs in chalk grasslands: dispersal of seeds. New Phytol., 95, 335-344.
Walker K.J. et al., 2004. The restoration and re-creation of species-rich lowland grassland on land formerly managed for intensive agriculture in the UK. Biol. Conserv., 119, 1-18.
WallisDeVries M.F., Poschlod P. & Willems J.H., 2002. Challenges for the conservation of calcareous grasslands in northwestern Europe: integrating the requirements of flora and fauna. Biol. Conserv., 104, 265-273.
Wells T.C.E., 1969. Botanical aspects of conservation management of chalk grassland. Biol. Conserv., 2, 36-44.
Wheater C.P. & Cullen W.R., 1997. The flora and invertebrate fauna of abandoned limestone quarries in Derbyshire, United Kingdom. Restor. Ecol., 5, 77-84.
Wheeler B.D. & Shaw S.C., 1991. Above-ground crop mass and species richness of the principal types of herbaceous rich-fen vegetation of lowland England and Wales. J. Ecol., 79, 285-301.
Willems J.H., 2001. Problems, approaches, and results in restoration of Dutch calcareous grassland during the last 30 years. Restor. Ecol., 9, 147-154.
Willems J.H., Peet R.K. & Bik L.P.M., 1993. Changes in chalk-grassland structure and species richness resulting from selective nutrient additions. J. Veg. Sci., 4, 203-212.
Willems J.H. & Bik L.P.M., 1998. Restoration of high species density in calcareous grassland: the role of seed rain and soil seed bank. Appl. Veg. Sci., 1, 91-100.
Zobel M., Suurkask M., Rosén E. & Pärtel M., 1996. The dynamics of species richness in an experimentally restored calcareous grassland. J. Veg. Sci., 7, 203-210.
To cite this article
About: Julien Piqueray
Univ. Liège - Gembloux Agro-Bio Tech. Unité Biodiversité et Paysage. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : Julien.Piqueray@ulg.ac.be
About: Grégory Mahy
Univ. Liège - Gembloux Agro-Bio Tech. Unité Biodiversité et Paysage. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
