


- Portada
- Volume 15 (2011)
- numéro 1
- Lutte contre les ravageurs des stocks de céréales et de légumineuses au Sénégal et en Afrique occidentale : synthèse bibliographique


Vista(s): 32424 (194 ULiège)
Descargar(s): 1654 (37 ULiège)

Lutte contre les ravageurs des stocks de céréales et de légumineuses au Sénégal et en Afrique occidentale : synthèse bibliographique

Notes de la rédaction
reçu le 29 janvier 2010, accepté le 1 juin 2010
Résumé
Les pertes post-récolte des céréales et légumineuses sont encore au Sénégal et en Afrique occidentale un problème majeur. Les réponses apportées pour combattre les insectes, principaux déprédateurs des stocks, ont été essentiellement chimiques. Cependant, compte tenu des nuisances associées à l'utilisation des pesticides, sélection de souches résistantes, pollution de l'environnement, intoxications, la recherche d'alternatives s'impose. Il est fait état des différentes méthodes de protection des stocks pratiquées en alternative ou combinées avec les pesticides. Les principales espèces de ravageurs rencontrées, en particulier Prostephanus truncatus (Horn), insecte émergeant au Sénégal, peuvent faire l'objet d'une lutte à l'aide de méthodes alternatives utilisant en particulier des plantes insecticides. Divers aspects de cette problématique sont passés en revue.
Abstract
Controlling pests of cereals and legumes in Senegal and West Africa: a review. Post-harvest losses of cereals and legumes are a major problem in Senegal and West Africa. The solutions to eliminate insects, major pests of stored products were mainly chemical. However, due to pollution associated with pesticides use, selection of resistant strains, environmental pollution, poisoning, the search for alternatives is needed. It is reported on different methods of protecting stocks performed alternatively or in combination with pesticides. The major pest species encountered, particularly Prostephanus truncatus (Horn), insect emerging in Senegal, could be controlled by alternative methods including specially the use of insecticide plants. Different aspects related to this alternative way to chemical pesticides are reviewed herein.
Tabla de contenidos
1. Contexte
1Dans le Sahel, les céréales et les légumineuses constituent la base de la nourriture des populations. Au Sénégal, le mil et le maïs sont les deux céréales majeures, tandis que l'arachide, produit de rente, jadis base de l'économie sénégalaise, avec le niébé, sont les principales légumineuses cultivées dans le pays. Pour toutes ces spéculations, les rendements actuels sont largement en deçà des attentes de la couverture nationale en nourriture. La sécurité alimentaire continue d'en pâtir. Le bilan de la recherche agricole au Sénégal a énuméré les différentes contraintes quasi communes à toutes ces cultures : sècheresse, déficits hydriques récurrents, maladies, mauvaises herbes, épuisement progressif des sols, manque d'encadrement, absence d'intrants, désengagement de l'État et désorganisation des filières, qualité et performances des semences, etc. (Ba et al., 2005 ; Ndiaye et al., 2005).
2Le manque de ressources alimentaires est comblé depuis toujours par des importations massives, notamment de céréales. Si, au niveau des céréales sèches, la dépendance extérieure est faible, il en est autrement pour le riz et le blé pour lesquels la moyenne annuelle d'importation est de l'ordre de 600 000 tonnes pour une enveloppe estimée à 110 milliards de francs CFA (Fall et al., 2008). Selon le PAM (2008), les produits céréaliers représentent 56 % des importations des produits alimentaires, soit 1,5 millions de tonnes. Le déficit vivrier chronique a été exacerbé par la crise alimentaire mondiale de 2008. En effet, la précarité de l'offre, spécialement pour le riz et le blé, ainsi que le renchérissement des prix d'achat, sont venus aggraver le problème. Pourtant, à la faveur des programmes spéciaux du gouvernement sénégalais lancés depuis 2003, la production céréalière connait ces dernières années une progression régulière. L'émergence de nouveaux paradigmes tels que la réduction de la pauvreté, l'amélioration de la sécurité alimentaire ainsi que les innovations dans les exploitations ont fini par inverser la tendance en matière de recherche. Les cultures vivrières autrefois délaissées au profit des produits d'exportation se voient de plus en plus considérées dans les programmes de recherche des institutions nationales et sous-régionales (Coly, 2007). Au Niger, des chercheurs de l'Université de Niamey et de l'Institut de Recherche pour le Développement ont identifié le gène PHYC jouant un rôle dans la perception de la lumière chez les plantes et responsable de l'adaptation du mil aux conditions de sècheresse, aux sols pauvres et à la grande variabilité des conditions environnementales (IRD, 2009). Selon cette étude, le mil, base alimentaire de 50 millions d'habitants, reste la seule culture correspondant aux conditions du Sahel et aux habitudes alimentaires traditionnelles.
3Malgré les initiatives prises de part et d'autre pour accroître la production, le contexte d'insécurité alimentaire est toujours marqué par des pertes post-récolte non négligeables. La valeur réelle des déperditions notées lors du stockage est depuis fort longtemps sujette à polémique. Devant la gravité des pertes post-récolte du fait des insectes, Labeyrie (1992) disait qu'en Afrique, le paysan travaille pour les insectes. Face à la menace que constituent les insectes, principaux ravageurs des stocks, les moyens de lutte sont essentiellement articulés autour de l'utilisation de pesticides de synthèse. Dans des conditions optimales, leur efficacité à contrôler les nuisibles des stocks est certaine. Toutefois, beaucoup de griefs leur sont portés, parmi lesquels l'accoutumance des insectes et la sélection de souches résistantes (Benhalima et al., 2004), intoxications, pollution de l'environnement et désordres écologiques (Regnault-Roger, 2002). Compte tenu de la réticence des consommateurs à consommer des produits traités aux pesticides, les limites tolérables tendent vers des valeurs infinitésimales. Afin de faire face à la problématique de synthétiser de façon permanente de nouvelles molécules insecticides et puisque les pesticides ne peuvent pas être abandonnés, c'est la gestion intégrée qui est encouragée (Adda et al., 2002 ; Ketoh et al., 2005). Ceci rend obligatoire la découverte d'alternatives moins polluantes. Dans cette optique, la valorisation des plantes à effet insecticide semble séduire partout dans le monde. Ainsi, plusieurs organes et parties des plantes sont exploités aux fins de limiter les pertes post-récolte. Cette nouvelle approche prend de l'ampleur au niveau des programmes de recherches en Afrique. Glitho (2002), sur la base de publications et communications de chercheurs du Réseau des Bruches (REARB), a recensé 43 espèces de plantes appartenant à 33 genres de 15 familles et faisant l'objet de tests biologiques sur les insectes des denrées stockées.
4Nous tenterons dans les parties qui suivent de mettre l'accent sur les alternatives aux pesticides organiques et en particulier, sur l'utilisation des plantes insecticides dans la préservation des récoltes au Sénégal et en Afrique de l'Ouest.
2. Les insectes dans le système post-récolte
5Beaucoup de travaux font référence à l'attaque et à la déperdition des stocks de céréales et de légumineuses par les insectes (Philogène et al., 1989 ; Ratnadass et al., 1989 ; Ashamo, 2006). Ils sont responsables sous les tropiques de pertes pouvant dépasser 30 %. Un autre aspect inhérent à l'établissement des insectes est la colonisation des stocks par des champignons producteurs de toxines à l'image d'Aspergillus flavus, producteur d'aflatoxine. Ainsi, au Bénin, Lamboni et al. (2009) ont recensé, sur du maïs stocké, neuf espèces de champignons, principalement des genres Penicillium, Rhizoctonia et Aspergillus associés aux ravageurs. Cathartus quadricollis (Guérin) est apparu comme l'insecte ravageur le plus actif dans ce rôle de « transporteur de champignon ».
2.1. Les principaux ravageurs des céréales et légumineuses au Sénégal et en Afrique occidentale
6Deux catégories de ravageurs sont à distinguer : ceux qui attaquent les récoltes dès le champ à l'image de Caryedon serratus (Olivier) sur arachide et de Callosobruchus maculatus (F.) sur niébé et ceux qui sont exclusivement inféodés aux stocks, tels que Corcyra cephalonica (Stainton) et Tribolium castaneum (Herbst) (Tableau 1). Au Sénégal, les travaux se rapportant au stockage des vivriers, que sont les céréales et légumineuses, ont essentiellement trait au mil, à l'arachide et au niébé. Seck et al. (1992) ont mis en évidence l'importance du taux de brisures sur les pertes subies par le mil stocké. À travers d'autres études, Seck (1991, 1992) a abordé la dynamique des populations de quelques insectes ravageurs du mil et a ensuite essayé des méthodes de lutte intégrée pour la gestion de ces populations. Ces études ont permis une meilleure compréhension des mécanismes de pullulation de ces insectes. Guèye et al. (1999) ont quant à eux examiné la sensibilité du mil, de ses dérivés et du fonio vis-à-vis de quatre insectes des stocks. Cette étude a mis en évidence l'importance de l'intégrité et de la finesse des grains sur leur résistance à l'infestation.
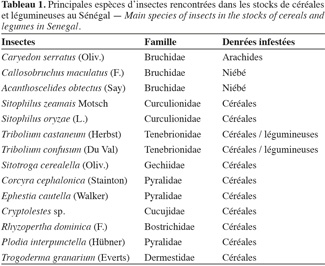
7L'arachide et le niébé sont plus sensibles à l'infestation des déprédateurs. Sur arachide, Guèye (2000) situe vers avril la période où l'émergence d'adultes de C. serratus (Olivier) est à son niveau maximal, qui coïncide avec une forte canicule. Considérant la perte en poids, des tests en stocks fermés (Ndiaye, 1991) montrent que la pré-infestation au champ des arachides par C. serratus, même si elle est faible, peut suffire à détruire complètement des stocks destinés à fournir des semences en quatre mois de conservation seulement.
8La sensibilité du niébé aux ravageurs est sans doute la cause du nombre important d'études consacrées à cette denrée. Acanthoscelides obtectus (Say), Callosobruchus rhodesianus (Pic) et surtout C. maculatus sont les insectes majeurs responsables de la quasi-totalité des dégâts observés sur niébé. Il arrive qu'il y ait compétition entre eux dans les mêmes stocks. La plus grande voracité de C. maculatus a été relatée dans plusieurs travaux ; Amevoin et al. (2005) ont décrit la dynamique de cohabitation des deux Callosobruchus. En effet C. rhodesianus atteint son pic et disparait en trois mois, coïncidant avec une explosion des populations de C. maculatus qui est responsable de l'essentiel des dégâts.
2.2. Cas de Prostephanus truncatus (Horn), ravageur du maïs émergeant au Sénégal
9P. truncatus est un ravageur redouté sur maïs et cossettes de manioc. Depuis son introduction accidentelle à la fin des années 1970, d'abord en Tanzanie et par la suite en Afrique occidentale, au Togo, en 1984 (Farrell et al., 2002), P. truncatus ne cesse de progresser. Une étude de Nansen et al. (2002a) indiquait sa présence dans au moins 14 pays africains. Il vient d'être découvert pour la première fois au Sénégal (Guèye et al., 2008a) dans deux sites du département de Kolda à 12°50'57"N 15°02'36"W et 12°54'35"N 14°57'01"W et dans la nouvelle région de Kédougou à 12°29'20"N 12°17'26"W. Tous ces sites sont à moins de 20 km des frontières de la Guinée où il a été signalé depuis 1998 (Aman et al., 2007). Il est probable que P. truncatus a été introduit au Sénégal à partir de la Guinée Conakry par le biais des transactions qui s'effectuent notamment au niveau des marchés hebdomadaires frontaliers.
10Verstraeten et al. (1987), menant des essais sur les possibilités de migration de P. truncatus dans du maïs égrené, ont pu mettre en évidence sa mobilité, sa rapidité de pénétration et sa voracité. Déjà, les auteurs prévenaient sur les risques d'envahissement de régions entières en quelques années, eu égard à son adaptation au manioc, au sorgho et de ses possibilités de vivre dans le bois tendre des silos. Nansen et al. (2002b) ont établi par piégeage une corrélation positive entre l'activité d'individus de P. truncatus et la proximité de plantations de teck (Tectona grandis). Une étude modélisant les types de dégâts entre S. zeamais et P. truncatus dans des magasins en milieu rural ouest africain a conclu à la plus grande voracité de P. truncatus qu'ils imputent à son comportement : le forage de l'adulte détruit quatre fois plus que la consommation de la larve et de l'adulte (Holst et al., 2000). Les dégâts peuvent s'amplifier lorsque l'infestation commence dès le champ (Vowotor et al., 2005). Cependant, depuis l'introduction de son ennemi naturel Teretrius nigrescens (Lewis), Hill et al. (2003) constatent en cinq ans une réduction drastique (plus de 80 %) des populations de P. truncatus au Kenya.
3. Moyens de lutte
11Cette revue n'a pas pour objet de s'étendre sur les différents groupes de pesticides, sur leurs modes d'action et efficacité. Son ambition est de mettre en évidence les dangers liés aux pesticides qui justifient la recherche de méthodes alternatives. Actuellement, près de 750 000 personnes contractent, chaque année, une maladie chronique telle que les cancers suite à une exposition à des pesticides. Plus de 20 000 décès accidentels et 3 millions d'empoisonnements liés aux pesticides sont annuellement recensés dans le monde (PAN Africa, 2003).
3.1. La lutte chimique
12Insecticides utilisés dans la protection des stocks au Sénégal. Étant membre du Comité Permanent Inter-états de Lutte contre la Sècheresse au Sahel (CILSS) qui regroupe en plus les pays suivants, Burkina Faso, Cap Vert, Gambie, Guinée Bissau, Mali, Mauritanie, Niger et Tchad, le Comité Sahélien des Pesticides (CSP) autorise les produits dans le traitement des denrées alimentaires. Le CSP a pour tâche de prendre des décisions communes à l'ensemble des pays du CILSS en matière de circulation des pesticides et en vue de leur utilisation judicieuse. La dernière actualisation de la liste globale des pesticides autorisés a eu lieu en juillet 2009. Toutefois, dans le domaine de la protection des denrées alimentaires, il n'a été retenu que quatre molécules différentes (Tableau 2).
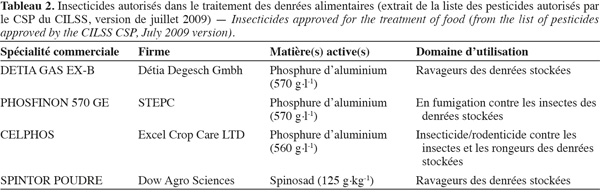
13Comme l'indique le tableau 2, seuls trois fumigants et un insecticide de contact (Spintor) sont dédiés aux denrées stockées, sur un total de 101 produits. Toutefois, il convient de noter que dans la pratique, d'autres insecticides sont encore couramment utilisés. Il s'agit entre autres de l'Actellic (m.a. pirimiphos-méthyle), de la K-Othrine (m.a. deltaméthrine 250 g.kg-1) et du Dursban (chlorpyriphos-éthyl 50 g.kg-1). Ces produits ne sont pas interdits dans les pays du CILSS, mais ils ne sont autorisés que pour d'autres applications. En guise d'exemple, la deltaméthrine est autorisée dans la lutte contre Helicoverpa armigera, le chlorpyriphos-éthyl (dursban) contre les sauteriaux, les termites du manguier, la cyperméthrine contre les chenilles, les insectes piqueurs-suceurs du cotonnier, etc. Les raisons de la restriction de ces produits dans le traitement des denrées stockées ne sont pas exposées, mais il reste évident que leur liberté de circulation est synonyme de leur usage dans les denrées stockées. Une enquête récente menée par Guèye et al. (2008b) a fait ressortir l'usage de raticides, fongicides, herbicides ou encore des liquides non déterminés sur du maïs destiné à la consommation humaine. En outre, l'étude a relevé une ignorance totale des matières actives et doses à utiliser. La couleur du sachet semble être le critère le plus retenu dans le choix du pesticide.
14Sur le plan financier, au Sénégal, outre les quantités fabriquées localement et consommées dans le pays, le CORAF (2007) estimait les importations en insecticides (matières actives, produits finis) à 4,4 milliards de CFA (6 707 317 euros). Ces importations ne se font pas sur autorisation préalable du Service de la Protection des Végétaux ou du secrétariat permanent du comité national de gestion des pesticides, qui permettrait d'instaurer un contrôle pour une meilleure gestion de ces produits.
15Résistance des ravageurs aux insecticides. La résistance des insectes aux pesticides de synthèse est l'un des principaux méfaits de l'application répétée des produits contre les ravageurs. De nombreux cas de résistance sont révélés partout. Actuellement, aucun groupe parmi les organophosphorés, organochlorés, pyréthrinoïdes ou encore fumigants n'échappe à la résistance des insectes. Avec le retrait au niveau mondial du bromure de méthyle en 2015 (Bell, 2000), le problème de la fumigation est un des plus préoccupants en Afrique. Bell et al. (1995) puis Benhalima et al. (2004) rapportent respectivement des souches de Trogoderma granarium (Everts) provenant du Burkina et des populations de Sitophilus zeamais (Motsch) du Maroc ayant un fort degré de tolérance à la phosphine, produit réputé efficace et ne laissant pas de résidus. Les espèces du genre Sitophilus sont parmi celles qui sont le plus citées dans la résistance aux pesticides. Pour pallier cette résistance, une association avec d'autres gaz a été prônée. À cet égard, Athié et al. (1998) suggèrent la combinaison de la phosphine au dioxyde de carbone (10 à 20 %) pour maitriser des souches résistantes de S. oryzae et de Rhyzopertha dominica (F.). Pimentel et al. (2009), sur base de la méthode FAO de détection standard de la résistance à la phosphine, ont aussi découvert pour la première fois au Brésil la résistance à la phosphine de vingt populations de S. zeamais issues des LC50 jusqu'à 86,6 fois plus élevées. Ces auteurs expliquent la base physiologique de la résistance à la phosphine par la capacité de ces souches de S. zeamais à réduire leur taux de respiration et non par une exclusion active du gaz. L'examen de ces différents travaux montre que l'acquisition de la tolérance à la phosphine est dans bien des cas associée à de mauvaises pratiques de fumigation dont le manque d'étanchéité des silos, le manque de suivi de la concentration du gaz lors des opérations, le temps d'exposition, mais surtout l'utilisation à grande échelle d'un seul produit. Zettler et al. (2000) faisaient déjà état de la menace qui pèse sur l'utilisation de ces fumigants, raison pour laquelle ils évoquent l'intérêt de développer de nouvelles alternatives dans le cadre de programmes de lutte intégrée. À ce sujet, plusieurs pistes sont proposées. Herron et al. (1996), se référant à des lieux de stockage où des grains traités ont été mélangés à des reliques de vieux grains infestés, ont constaté une augmentation des niveaux d'infestation et de la résistance au fénitrothion et au pirimiphos-méthyl chez des populations de Oryzaephilus surinamensis (L.). Ainsi, les mesures d'hygiène, bien que n'ayant que peu d'effet direct, s'avèrent nécessaires pour ralentir le développement de la résistance. Mohandas et al. (2006) suggèrent l'hydroprène, un analogue de l'hormone juvénile présenté comme alternative aux insecticides conventionnels de par sa spécificité contre les formes immatures des insectes de stocks et une non-toxicité vis-à-vis des mammifères. Selon Mbata et al. (2009), une faible oxygénation découlant de l'application d'une faible pression (32,5 ± 1,0 mmHg) lors du stockage des variétés résistantes à C. maculatus peut constituer une alternative aux fumigants.
16L'ensemble des cas évoqués ci-dessus montre la nécessité d'associer la lutte chimique à d'autres techniques qui seront à même de confiner les dégâts des insectes dans des limites économiquement supportables, tout en assurant un environnement moins exposé aux pollutions chimiques.
3.2. Les alternatives à la lutte chimique
17Face aux nuisances de la lutte chimique nonobstant les succès enregistrés, il a été développé plusieurs autres formes de lutte contre les insectes. Nous ne citerons que les principales.
18La résistance variétale. Des variétés plus tolérantes aux insectes ont été développées dans le but de limiter les pertes. Le maïs a été particulièrement étudié dans les pays tropicaux (Kumar, 2002) et en Afrique occidentale, suite à la récente apparition de P. truncatus dans ce continent. Au Nigéria, Appleby et al. (2003) résument les principaux atouts issus de variétés résistantes de niébé sur C. maculatus en un allongement du développement pré-imaginal, à la forte mortalité et au faible poids des adultes émergeants. Des caractéristiques telles que la rugosité de la surface et le poids des grains, ainsi que la dureté peuvent réduire les pertes dues à C. maculatus à cause d'un faible taux de multiplication et de croissance des populations (Sulehrie et al., 2003).
19Lutte physique
20L'irradiation et la lutte par le froid. Ces méthodes, bien que procurant de bons résultats, ne sont guère présentes en Afrique du fait du cout de l'énergie et de la lourdeur des installations de base.
21Le stockage hermétique. Ce type de stockage est de plus en plus pratiqué en milieu rural. Le niébé est l'une des denrées les plus concernées du fait de sa forte sensibilité aux ravageurs. Ce sont souvent des fûts d'huile récupérés, des bidons en plastique et des jarres commodes pour emmagasiner jusqu'à 100 voire 200 kg de graines. Il est devenu actuellement pratique courante de rajouter quelques comprimés de phostoxin. Le stockage hermétique de graines de niébé combiné à l'ajout de fruits de B. senegalensis à 1,2 g.l-1 (poids échantillon/volume enceinte) réduit l'émergence de C. maculatus, tandis qu'une concentration de 2,4-4,8 g.l-1 inhibe complètement l'avènement d'une nouvelle génération (Seck et al., 1996).
22L'insolation. C'est une pratique effectuée le plus souvent avant emmagasinage des récoltes. Elle permet d'achever le séchage et de faire fuir les insectes grâce à la chaleur et à l'incidence directe des rayons solaires. Des essais conduits sur niébé ont donné une mortalité totale des bruches C. maculatus et C. subinnotatus (Pic) au bout de 6 h d'exposition à 50 °C (Lale et al., 2003). D'après Sembène et al. (2006), sur arachide, si la température externe au sol est supérieure à 33 °C, une heure d'exposition au soleil dans un dispositif de type Murdock et al. (1991) suffit pour obtenir la mort de tous les stades de Caryedon serratus. Il faut dire que le renouvellement périodique de cette opération lors des journées ensoleillées pourrait être à même d'assurer un stockage adéquat. Le cas des semences est particulièrement intéressant à considérer.
23L'enfumage. Il est surtout pratiqué en milieu rural autant pour les vivres que les semences. Dans le cas des semences, essentiellement de mil et de maïs, les épis sont suspendus au-dessus du foyer, lui proférant une immunité contre les insectes grâce à la chaleur et à la fumée. L'accumulation potentielle de substances délétères issues des fumées n'a pas non plus fait l'objet d'une attention ou de recherches particulières.
24Poudres minérales et terres de diatomées. Les poudres à action abrasive à l'image de la cendre et du sable fin ont été traditionnellement utilisées dans la conservation des récoltes. Avec du maïs, Akob et al. (2007) ont démontré l'efficacité au bout de six mois de cendres de feuilles de Eucalyptus grandis sur S. zeamais à une dose de 20 g par 2 kg par une réduction de la F1 avec un seul émergeant et des pertes de poids de 2,4 %. De façon plus élaborée, les terres de diatomées ont connu un regain d'intérêt. Carlson et al. (1962) pensent que les terres de diatomées sont appropriées pour certains pays sous-développés qui connaissent des problèmes perpétuels de stockage à cause d'équipements inadéquats couplés avec de hautes températures et/ou humidité. Les avantages de leur utilisation : absence de résidus, efficacité dans le contrôle des insectes sans développement de résistance, facilité de manipulation. Korunic et al. (1996) éclairent leur mode d'action par un effet abrasif et une absorption des lipides cuticulaires entrainant la mort des insectes par dessiccation, conséquence d'une perte d'eau importante. Toutefois, leur efficacité est meilleure aux fortes températures et aux basses humidités et dépend en outre de la denrée traitée et de l'insecte considéré (Vayias et al., 2009). Deux formulations naturelles de zéolithes à des doses allant de 0,25 à 0,75 g.kg-1 se sont révélées efficaces (94-100 %) au bout de trois semaines avec S. oryzae (L.) et T. castaneum, alors qu'à la plus forte dose, soit à 0,75 g.kg-1, la mortalité avec R. dominica a été au meilleur des cas de 74 % (Kljajic et al., 2010).
25Lutte biologique. Ce mode de lutte s'articule dans la majeure partie des cas sur l'utilisation de parasitoïdes, parasites et prédateurs. Elle a été particulièrement étudiée en Afrique dans le cas de la bruche du niébé C. maculatus (Ketoh et al., 2002 ; Jaloux et al., 2004). L'introduction d'espèces comme Dinarmus basalis (Rondani) et Eupelmus vuilleti (CRW.) dans les greniers permit de limiter les populations de C. maculatus au Togo. Toutefois, il est relevé dans ces études d'une part, des compétitions interspécifiques entre parasitoïdes dont l'activité dépend de la densité de l'hôte et d'autre part, une sensibilité plus forte des auxiliaires aux traitements. Au Burkina, Sanon et al. (2005) ont constaté que l'introduction de feuilles de Boscia senegalensis dans les greniers montre une toxicité aussi bien pour les bruches C. maculatus que C. rhodesianus au cours des trois premiers mois de stockage. Du fait que les larves se développant à l'intérieur des graines sont peu affectées par le traitement et compte tenu de la diminution des composés volatils émis par les feuilles, les pertes au bout de six mois ne sont pas significativement différentes du contrôle. Une étude dynamique des populations de C. serratus au Sénégal a révélé un fort parasitisme des œufs de C. serratus sur Piliostigma reticulatum et une totale incapacité du parasitoïde à exercer la même prédation sur les œufs pondus sur arachide (Guèye, 2000). Sur céréales, nous n'avons pas connaissance de l'usage de parasites pour le contrôle des ravageurs dans la sous-région.
26Des micro-organismes entomopathogènes, bactéries, champignons, nématodes, protozoaires et virus ont intéressé les chercheurs dans la lutte biologique. L'espèce la plus connue dans ce domaine est la bactérie Bacillus thuringiensis. Par ailleurs, dans la lutte contre C. serratus, Ekesi et al. (2001) ont obtenu de l'application de conidies de Metarizhium anisopliae sur gousses d'arachide un contrôle comparable à celui obtenu avec le pyrimiphos-méthyl à 10 ppm. L'isolat le plus actif CPD 4 a même montré des effets répulsifs contre cette bruche. Dans le cas du maïs, une expérimentation conduite au Bénin en laboratoire et dans les champs a révélé les possibilités d'utilisation d'isolat de conidies de Beauveria bassina contre P. truncatus. L'efficacité d'un tel traitement sur la densité de P. truncatus proviendrait d'une forte mortalité au niveau larvaire (Meikle et al., 2001). Les auteurs pensent aussi à une possible persistance des champignons dans les cadavres des adultes par une sporulation lorsque les conditions deviennent défavorables. Ce qui serait bénéfique pour la persistance du traitement au cours du stockage.
27Malgré les chances de succès, le constat est que la lutte biologique tarde à prendre son essor dans beaucoup de pays ouest-africains. Certains facteurs concourent à limiter l'emploi de techniques assez exigeantes voire sophistiquées dans les zones rurales africaines particulièrement concernées par les pertes post-récolte. Ekesi et al. (2001) soulignent que l'efficacité des entomopathogènes peut être affectée par de fortes intensités lumineuses et les rayons ultraviolets. Ainsi, un stockage de longue durée dans de telles localités s'avère difficile, surtout si l'on sait que beaucoup de localités sont dépourvues d'énergie et d'installations adéquates pour la conservation de tels organismes. De plus, les champignons ont besoin d'une humidité élevée pour être actifs, corollaire d'une détérioration des grains à stocker. Ces quelques limites sont de nature à nous inciter à rechercher d'autres moyens utilisant des techniques moins couteuses et dont la mise en œuvre par les producteurs est moins contraignante.
4. Utilisation des plantes insecticides
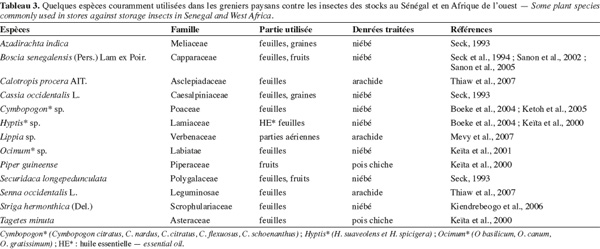
28L'usage des plantes indigènes dans la conservation des récoltes a été pratiqué avant même l'apparition des insecticides de synthèse. Les plantes sont utilisées contre les ravageurs pour leurs effets répulsif, de contact ou fumigant. Les molécules actives peuvent varier d'une famille à une autre et à l'intérieur d'une même famille et la sensibilité peut différer pour un insecte donné d'un stade à un autre. Boeke et al. (2004) ont relevé une action répulsive et toxique de Tephrosia vogelii Hook f. sur C. maculatus, alors que Blumea aurita (L.) DC, qui ne présente aucune toxicité, possède un fort pouvoir répulsif. Par ailleurs, Dracaena arborea du Mono au Bénin est répulsif et la même espèce récoltée dans le Borgou n'est guère efficace. Paul et al. (2009) fournissent plusieurs exemples de plantes avec une efficacité différente entre les feuilles entières ou réduites en poudre et les graines dans le contrôle de C. maculatus et Zabrotes subfasciatus (Boheman). Ces auteurs ont en outre mis en exergue avec Chenopodium ambrosioides une variabilité de l'efficacité des traitements en fonction des zones de collecte et des stades phénologiques. Le tableau 3 indique quelques espèces employées en Afrique occidentale dans la lutte contre les insectes.
4.1. Toxicité de contact chez les plantes insecticides
29Le neem Azadirachta indica est sans doute la plante la plus étudiée pour ses propriétés antiappétantes, répulsives, toxiques et inhibiteur de la croissance des insectes. Plusieurs composés ont été mis en évidence dans l'activité du neem et les principaux sont des limonoïdes. L'azadirachtine en est le composé majoritaire ; la salanine, la nimbine et leurs analogues sont aussi significativement présents (Lale et al., 1999). Cependant, seuls l'azadirachtine A (Aza A) et le 3-thigloyazadirachtol (Aza B) ont une activité significative. Philogène et al. (2002) font mention de l'utilisation du neem dans le sous-continent indien depuis plus de 4 000 ans contre les insectes des denrées stockées. Dans le cas du haricot, Facknath (2006) suggère aux fermiers pauvres comme moyen simple et bon marché la combinaison du neem et du brassage des grains (source de vibrations). Cette méthode donne un contrôle plus satisfaisant que l'un ou l'autre système utilisé seul dans le contrôle des populations de A. obtectus, S. oryzae, O. surinamensis et Cryptolestes ferrugineus (Stephens). En fait, la perturbation physique a montré les mêmes capacités que le neem à induire une réduction de la F1 et un allongement du cycle chez tous les insectes testés.
30Il est intéressant de mentionner les travaux de Seck et al. (1994) qui ont obtenu en application directe de fruits et de feuilles de B. senegalensis à des doses de 2 à 4 % (P/P) 80 à 100 % de mortalité des adultes de C. maculatus et réduit significativement à la fois l'émergence et les dégâts de la F1. Kiendrebeogo et al. (2006) ont eu une approche novatrice par l'utilisation d'une plante parasite, Striga hermonthica (Del.) Benth sur C. maculatus. Toutefois, la fraction éther de pétrole qui a donné le meilleur résultat permet à peine un effet ovicide de 51 % et larvicide de 72 % à la dose de 0,4 % (P/P).
4.2. Les huiles essentielles
31Les huiles essentielles des plantes font partie ces dernières années des voies les plus explorées dans la régulation des ravageurs. Leur application dans la protection des stocks a fait l'objet de nombreux travaux. Leur toxicité s'exprime de différentes manières : activités ovicide, larvicide, antinutritionnelle et inhalatoire (Kéïta et al., 2000 ; Regnault-Roger, 2002). Mis à part l'inhibition de l'éclosion des œufs, les vapeurs d'huiles essentielles accroissent la mortalité des larves. Papachristos et al. (2002) ont démontré la toxicité de Lavandula hybrida, Rosmarinus officinalis et Eucalyptus globulus sur les œufs de A. obtectus avec une différence de sensibilité significativement corrélée à l'âge. C'est au-delà de trois jours que la sensibilité est la plus forte, probablement à cause d'une plus grande perméabilité du chorion ou de la membrane vitelline facilitant ainsi la diffusion des vapeurs. Ogendo et al. (2008) ont quant à eux démontré la toxicité des huiles essentielles de Ocimum gratissimum L. à 1 µl.l-1 sur R. dominica, O. surinamensis et C. chinensis (L.) avec des taux de mortalité de 98 à 100 % en 24 h. Ils ont identifié le méthyle eugénol comme composant majoritaire et précisent une très forte variation dans la composition chimique de neuf chémotypes de O. gratissimum en relation avec la saison, le stade récolte ainsi que l'origine géographique. Habiba (2007) a obtenu une toxicité par contact équivalente avec des huiles essentielles d'O. gratissimum ainsi que celles de Xylopia aethiopica sur S. zeamais. L'étude menée par Noudjou-Wandji (2007) semble montrer un effet knock-down de l'huile essentielle de la poudre de X. aethiopica sur C. maculatus. Cet auteur a obtenu une mortalité maximale lors des trois premiers jours au-delà desquels la teneur en huile baisse, résultant d'une quasi-absence de molécules volatiles libérées entrainant une baisse accrue de la mortalité.
32Les huiles essentielles de certaines plantes sont utilisées pour leurs activités de contact et inhalatoire qui n'offrent pas souvent le même degré d'efficacité selon la cible visée. Citons, à titre d'exemple, Tapondjou et al. (2002) qui ont obtenu de la poudre de feuilles de C. ambrosioides en 24 h une mortalité totale des adultes de S. granarius (L.) et S. zeamais à une dose de 6,4 % (P/P). À 0,4 %, il n'a été noté aucune émergence à la F1 pour les bruches C. chinensis, C. maculatus et A. obtectus. Par contre, la fumigation par l'huile essentielle de cette plante (0,2 µl.cm-2) tue 80 à 100 % des adultes de C. chinensis, A. obtectus, S. zeamais et P. truncatus, tandis qu'avec C. maculatus et S. granarius, la mortalité n'a été respectivement que de 20 et 5 %. Selon ces derniers, il est probable que l'activité des feuilles soit due à une forte teneur en huiles essentielles. Liu et al. (1999) relatent une plus grande sensibilité des adultes de S. zeamais (DL50 = 0,043 mg.mg-1 de poids corporel) que T. castaneum (DL50 = 0,118 mg.mg-1 de poids corporel) au contact de l'huile essentielle de Evodia rutaecarpa (Hook.f. et Thomas), alors qu'en fumigation S. zeamais (LC50 = 41 mg.l-1 air) se révèle plus tolérant que T. castaneum (LC50 = 11,7 mg.l-1 air). Par une mortalité de contact équivalente à celle des témoins, George et al. (2009) ont apporté la preuve que les huiles essentielles de Thymus vulgaris (L.) et de Mentha pulegium (L.), actives sur l'acarien des poules Dermanyssus gallinae, ne le sont que par inhalation (CL99 < 0,30 mg.cm-3). Ketoh et al. (2002) évoquent une possible absorption des huiles par les graines de niébé. De plus, le parasitoïde D. basalis semble plus sensible que son hôte aux terpènes émis particulièrement par Cymbopogon schoenanthus (L.) et Ocimum basilicum (L.).
5. Perspectives
33Les différentes méthodes présentées comme alternatives aux pesticides présentent chacune des avantages, mais aussi quelques limites. C'est là tout le sens d'une gestion intégrée basée sur la combinaison de plusieurs procédés pour circonscrire l'activité des insectes, redoutables compétiteurs de l'homme. Les différences de sensibilité des insectes aux produits devraient nous amener à considérer la lutte de manière plus spécifique. En effet, le choix de l'insecticide (naturel) devra en priorité reposer sur l'insecte majeur à combattre. Il a été prouvé que la réponse à l'intensité de la toxicité d'un produit dépendait dans bien des cas de manière spécifique du ravageur et pour un ravageur donné du stade considéré. L'efficacité des plantes n'est pas non plus toujours garantie avec tous ses organes et dépend du stade phénologique. De plus, l'efficacité des biocides est en partie associée aux conditions pédoclimatiques.
34Dans cette lutte destinée aux fermiers dont la plupart en Afrique sont illettrés ou à faible niveau d'éducation, il est impératif de tenir compte des choix de la technique et des modèles proposés. En effet, les cadres de vie en milieu rural souvent rudimentaires rendent complexe la mise en pratique de certaines méthodes. La promotion de variétés résistantes, des techniques telles que l'insolation ainsi que l'application directe de plantes à action inhalatoire sont à prendre avec plus de considération, compte tenu de leur simplicité de mise en œuvre. Certaines pratiques empiriques encore utilisées dans la conservation des récoltes mériteraient d'être évaluées afin d'établir scientifiquement leur efficacité réelle et leur innocuité sur la santé des populations, compte tenu des conditions de stockage. Dans cette perspective, nous entamerons un travail de recherche sur l'effet de diverses plantes potentiellement insecticides à la fois sur céréales et légumineuses sur les insectes S. zeamais, T. castaneum, C. maculatus, C. serratus et P. truncatus. Les molécules actives seront recherchées, de même que les résidus sur les denrées stockées destinées à l'alimentation.
Bibliographie
Adda C., Borgemeister C., Biliwa A. & Meikle W.G., 2002. Integrated pest management in post-harvest maize: a case study from the Republic of Togo (West Africa). Agric. Ecosyst. Environ., 93, 305-321.
Akob C.A. & Ewete F.K., 2007. The efficacy of ashes of four locally used plant materials against Sitophilus zeamais (Coleoptera: Curculionidae) in Cameroon. Int. J. Trop. Insect Sci., 27(1), 21-26.
Aman O.B., Schulthess F., Masiga D. & Van den Berg J., 2007. On the diversity of Teretrius nigrescens, predator of the Larger Grain Borer. In: 17e Conférence de l'Association des Entomologistes Africains et de la Société entomologique du Sénégal, Towards food security, health and protection of the environment in Africa: the role of insect sciences, 11-15 juin 2007, Dakar, Sénégal.
Amevoin K., Glitho I.A., Monge J.P. & Huignard J., 2005. Why Callosobruchus rhodesianus causes limited damage during storage of cowpea seeds in a tropical humid zone in Togo? Entomol. Exp. Appl., 116, 175-182.
Appleby J.H. & Credland P.F., 2003. Variation in responses to susceptible and resistant cowpeas among west African populations of Callosobruchus maculatus (Coleoptera: Bruchidae). J. Econ. Entomol., 96(2), 489-502.
Ashamo M.O., 2006. Relative susceptibility of some local and elite rice varieties to the rice weevil, Sitophilus oryzae L. (Coleoptera: Curculionidae). J. Food Agric. Environ., 4(1), 249-252.
Athié I. et al., 1998. Effects of carbon dioxide and phosphine mixtures on resistant populations of stored-grain insects. J. Stored Prod. Res., 34(1), 77-32.
Ba A. et al., 2005. L'arachide. In : Bilan de la Recherche Agricole et Agroalimentaire au Sénégal. Dakar : Institut Sénégalais de Recherches Agricoles (ISRA), Institut de Technologie Alimentaire (ITA), CIRAD, 163-188.
Bell C.H., 2000. Fumigation in the 21st century. Crop Prot., 19, 563-569.
Bell C.H. & Wilson S.M., 1995. Phosphine tolerance and resistance in Trogoderma granarium Everts (Coleoptera: Dermestidae). J. Stored Prod. Res., 31(3), 199-205.
Benhalima H., Chaudhry M.Q., Mills K.A. & Price N.R., 2004. Phosphine resistance in stored-product insects collected from various grain storage facilities in Morocco. J. Stored Prod. Res., 40, 241-249.
Boeke S.J. et al., 2004. Toxicity and repellence of African plants traditionally used for the protection of stored cowpea against Callosobruchus maculatus. J. Stored Prod. Res., 40, 423-438.
Carlson S.D. & Ball H.J., 1962. Mode of action and insectidal value of a diatomaceous earth as a grain protectant. J. Econ. Entomol., 55(6), 964-970.
CILSS, 2009. Liste globale actualisée des pesticides autorisés par le CSP en juillet 2009, http://www.insah.org/protectiondesvegetaux/csp/pesticidautorise.html, (05.01.2010).
Coly E.V., 2007. Accent renforcé sur les céréales et légumineuses. Agrovision, 3, 31-32.
CORAF, 2007. Programme de productivité agricole en Afrique de l'ouest. Plan de gestion des pestes et pesticides. Rapport E1553, v2. Dakar : CORAF ; Banque Mondiale.
Ekesi S., Egwurube E.A., Akpa A.D. & Onu I., 2001. Laboratory evaluation of the entomopathogenic fungus, Metarhizium anisopliae for the control of the groundnut bruchid, Caryedon serratus on groundnut. J. Stored Prod. Res., 37, 313-321.
Facknath S., 2006. Combination of neem and physical disturbance for the control of four insect pests of stored products. Int. J. Trop. Insect Sci., 26(1), 16-27.
Fall A.A. & Dièye P.N., 2008. Impact des cours mondiaux du riz sur la sécurité alimentaire au Sénégal. Réflexions Perspect. ISRA, 6(6).
Farrell G. & Schulten G.G.M., 2002. Larger grain borer in Africa; a history of efforts to limit its impact. Integr. Pest Manage. Rev., 7, 67-84.
George D.R. et al., 2009. Mode of action and variability in efficacy of plant essential oils showing toxicity against the poultry red mite, Dermanyssus gallinae. Vet. Parasitol., 161, 276-282.
Glitho I.A., 2002. Annexe : post-récoltes et biopesticides en Afrique. In : Regnault-Roger C., Philogène B.J.R. & Vincent C., coord. Biopesticides d'origine végétale. Paris : Lavoisier-Éditions Tec & Doc., 314-321.
Guèye M.T., 2000. Structure et évolution des peuplements de C. serratus, déprédateur de l'arachide entreposée en milieu sahélien. Thèse de doctorat : Université Cheikh Anta Diop, Dakar (Sénégal).
Guèye M.T. & Delobel A., 1999. Relative susceptibility of stored pearl millet products and fonio to insect infestation. J. Stored Prod. Res., 35, 277-283.
Guèye M.T. et al., 2008a. First report on occurrence of the larger grain borer Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) in Senegal. Afr. Entomol., 16(2), 309-311.
Guèye M.T. et al., 2008b. La protection des stocks de maïs au Sénégal : enquêtes sur les pratiques d'utilisation des pesticides et plantes à effet insecticide en milieu paysan. ITA Échos, 3.
Habiba K., 2007. Étude des potentialités d'utilisation d'huiles essentielles pour le contrôle de deux insectes ravageurs des grains Callosobruchus maculatus(Coleoptera : Bruchidae) et Sitophilus zeamais (Coleoptera : Curculionidae) au Nord Cameroun. Thèse de doctorat : Faculté des Sciences, Centre de Recherche sur la Biodiversité, Université Catholique de Louvain (Belgique).
Herron A.G., Clift A.D., White G.G. & Greening H.G., 1996. Relationships between insecticide use, grain hygiene and insecticide resistance in Oryzaephilus surinamensis (L.) (Coleoptera: Silvanidae) on grain-producing farms. J. Stored Prod. Res., 32(2), 131-136.
Hill M.G., Nang'ayo F.L.O. & Wright D.J., 2003. Biological control of the larger grain borer Prostephanus truncatus (Coleoptera:Bostrichidae) in Kenya using a predatory beetle Teretrius nigrescens (Coleoptera: Histeridae). Bull. Entomol. Res., 93, 299-306.
Holst N., Meikle W.G. & Markham R.H., 2000. Grain injury models for Prostephanus truncatus (Coleoptera: Bostrichidae) and Sitophilus zeamais (Coleoptera: Curculionidae) in rural maize stores in West Africa. J. Econ. Entomol., 93(4), 1338-1346.
IRD, 2009. Le mil, aliment du futur au Sahel. Actualité scientifique, fiche n°35.
Jaloux B., Sanon A., Huignard J. & Monge J.P., 2004. Interspecific relationships between the solitary ectoparasitoid, Eupelmus vuilleti (Crw.) (Eupelmidae), and its sympatric species, Dinarmus basalis (Rond.) (Pteromalidae), in the presence of their host, Callosobruchus maculatus Pic (Coleoptera Bruchidae). J. Insect Behav., 17(6), 793-808.
Keïta S.M. et al., 2000. Effect of various essential oils on Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). J. Stored Prod. Res., 36, 355-364.
Keïta S.M. et al., 2001. Efficacy of essential oil of Ocimum basilicum L. and O. gratissimum L. applied as an insecticidal fumigant and powder to control Callosobruchus maculatus (Fab.) (Coleoptera: Bruchidae). J. Stored Prod. Res., 37, 339-349.
Ketoh G.K., Glitho I.A. & Huignard J., 2002. Susceptibility of the bruchid Callosobruchus maculatus (Coleoptera: Bruchidae) and its parasitoid Dinarmus basalis (Hymenoptera: Pteromalidae) to three essential oils. J. Econ. Entomol., 95(1), 174-182.
Ketoh G.K., Koumaglo H.K. & Glitho I.A., 2005. Inhibition of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) development with essential oil extracted from Cymbopogon schoenanthus L. Spreng. (Poaceae), and the wasp Dinarmus basalis Rondani (Hymenoptera: Pteromalidae). J. Stored Prod. Res., 41, 363-371.
Kiendrebeogo M., Ouedraogo A.P. & Nacoulma O.G., 2006. Activités insecticides de Striga hermonthica (Del.) Benth (Scrophulariaceae) sur Callosobruchus maculatus (Fab.) (Coleoptera : Bruchidae). Biotechnol. Agron. Soc. Environ., 10(1), 17-23.
Kljajic P. et al., 2010. Laboratory assessment of insecticidal effectiveness of natural zeolite and diatomaceous earth formulations against three stored-product beetle pests. J. Stored Prod. Res., 46(1), 1-6.
Korunic Z. et al., 1996. The effect of diatomaceous earth on grain quality. Postharvest Biol. Technol., 9, 373-387.
Kumar H., 2002. Resistance in maize to the Larger Grain Borer, Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae). J. Stored Prod. Res., 38, 267-280.
Labeyrie V., 1992. Problèmes fondamentaux posés par les insectes des denrées. In : Foua-Bi K. & Philogène B.J.R., éds. Actes du seminaire international à Abidjan, Côte d'Ivoire, 29 janvier-1 février 1992, 9-14.
Lale N.E.S. & Abdulrahman H.T., 1999. Evaluation of neem (Azadirachta indica A. Juss) seed oil obtained by different methods and neem powder for the management of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) in stored cowpea. J. Stored Prod. Res., 35, 135-143.
Lale N.E.S. & Vidal S., 2003. Simulation studies on the effects of solar heat on egg-laying, development and survival of Callosobruchus maculatus (F.) and Callosobruchus subinnotatus (Pic) in stored bambara groundnut Vigna subterranea (L.) Verdcourt. J. Stored Prod. Res., 39, 447-458.
Lamboni Y. & Hell K., 2009. Propagation of mycotoxigenic fungi in maize stores by post-harvest insects. Int. J. Trop. Insect Sci., 29(1), 31-39.
Liu Z.L. & Ho S.H., 1999. Bioactivity of the essential oil extracted from Evodia rutaecarpa Hook f. et Thomas against the grain storage insects, Sitophilus zeamais Motsch. and Tribolium castaneum (Herbst). J. Stored Prod. Res., 35, 317-328.
Mbata G.N., Phillips T.W. & Payton M.E., 2009. Effects of cowpea varietal susceptibility and low pressure on the mortality of life stages of Callosobruchus maculatus (Coleoptera: Bruchidae). J. Stored Prod. Res., 45, 232-235.
Meikle W.G. et al., 2001. The effects of an entomopathogenic fungus, Beauveria bassiana (Balsamo) Vuillemin (Hyphomycetes), on Prostephanus truncatus (Horn) (Col.: Bostrichidae), Sitophilus zeamais Motschulsky (Col.: Curculionidae), and grain losses in stored maize in the Benin republic. J. Invertebr. Pathol., 77, 198-205.
Mevy J.P. et al., 2007. Chemical composition and some biological activities of the volatile oils of a chemotype of Lippia chevalieri Moldenke. Food Chem., 101, 682-685.
Mohandas S.M., Arthur F.H., Zhua K.Y. & Throneet J.E., 2006. Hydroprene: mode of action, current status in stored-product pest management, insect resistance, and future prospects. Crop Prot., 25, 902-909.
Murdock L.L. & Shade R.E., 1991. Eradication of cowpea weevil (Coleoptera: Bruchidae) in cowpeas by solar heating. Am. Entomol., 37(4), 228-231.
Nansen C. & Meikle W.G., 2002a. The biology of the larger grain borer, Prostephanus truncatus (Horn)(Coleoptera: Bostrichidae). Integr. Pest Manage. Rev., 7, 91-104.
Nansen C., Meikle W.G. & Korié S., 2002b. Spatial analysis of Prostephanus truncatus (Bostrichidae: Coleoptera) flight activity near maize stores and in different forest types in southern Benin, West Africa. Ann. Entomol. Soc. Am., 95(1), 66-74.
Ndiaye S., 1991. La bruche de l'arachide dans un agrosystème du Centre-Ouest du Sénégal : contribution à l'étude de la contamination en plein champ et dans les stocks de l'arachide (Arachis hypogea L.) par Caryedon serratus (Ol.) (Coleoptera, Bruchidae) ; rôle des légumineuse hôtes sauvages dans le cycle de cette bruche. Thèse de doctorat : Université de Pau et des Pays de l'Adour (France).
Ndiaye A. et al., 2005. Les céréales. In : Bilan de la Recherche agricole et agroalimentaire au Sénégal. Dakar : Institut Sénégalais de Recherches Agricoles (ISRA), Institut de Technologie Alimentaire (ITA), CIRAD, 241-256.
Noudjou-Wandji F., 2007. Utilisation des huiles essentielles pour la protection des grains contre les insectes ravageurs au nord du Cameroun. Thèse de doctorat : Faculté des Sciences agronomiques de Gembloux (Belgique).
Ogendo J.O. et al., 2008. Bioactivity of Ocimum gratissimum L. oil and two of its constituents against five insect pests attacking stored food products. J. Stored Prod. Res., 44, 328-334.
PAM (Programme Alimentaire Mondial), 2008. Rapport d'analyse de marché. Sénégal : commerce du Riz. World Food Programm.
PAN Africa (Pesticide Action Network), 2003. Les pesticides au Sénégal. 2e éd. Dr Abou Thiam & Dr Alassane Sarr, eds.
Papachristos D.P. & Stamopoulos D.C., 2002. Toxicity of vapours of three essential oils to the immature stages of Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res., 38, 365-373.
Paul U.V., Lossini J.S., Edwards P.J. & Hilbeck A., 2009. Effectiveness of products from four locally grown plants for the management of Acanthoscelides obtectus (Say) and Zabrotes subfasciatus (Boheman) (both Coleoptera: Bruchidae) in stored beans under laboratory and farm conditions in Northern Tanzania. J. Stored Prod. Res., 45, 97-107.
Philogène B.J.R, Armson J.T. & Lambert J.D.H., 1989. Facteurs contribuant à la protection du maïs contre les attaques de Sitophilus et Prostephanus. In : Parmentier M. & Kouahou F.-B., eds. Céréales en régions chaudes. Montréal, Canada : Aupelf-Uref ; Paris : John Libbey Eurotext, 47-56.
Philogène B.J.R, Regnault-Roger C. & Vincent C., 2002. Produits phytosanitaires insecticides d'origine végétale : promesses d'hier et d'aujourd'hui. In : Philogène B.J.R, Regnault-Roger C. & Vincent C., coord. Biopesticides d'origine végétale. Paris : Lavoisier-Éditions Tec & Doc, 1-17.
Pimentel M.A.G. et al., 2009. Phosphine resistance in Brazilian populations of Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). J. Stored Prod. Res., 45, 71-74.
Ratnadass A. & Sauphanor B., 1989. Les pertes dues aux insectes sur les stocks paysans de céréales en Côte d'Ivoire. In : Parmentier M. & Kouahou F.-B., eds. Céréales en régions chaudes. Montréal, Canada : Aupelf-Uref ; Paris : John Libbey Eurotext, 141-150.
Regnault-Roger C., 2002. De nouveaux phyto-insecticides pour le troisième millénaire ? In : Philogène B.J.R, Regnault-Roger C. & Vincent C., coord. Biopesticides d'origine végétale. Paris : Lavoisier-Éditions Tec & Doc, 19-39.
Sanon A., Garbab M., Augerb J. & Huignard J., 2002. Analysis of the insecticidal activity of methylisothiocyanate on Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) and its parasitoid Dinarmus basalis (Rondani) (Hymenoptera: Pteromalidae). J. Stored Prod. Res., 38, 129-138.
Sanon A. et al., 2005. Combining Boscia senegalensis Lamarck (Capparaceae) leaves and augmentation and the larval parasitoid Dinarmus basalis Rondani (Hymenoptear: Pteromalidae) for bruchids control in stored cowpea. J. Entomol., 2(1), 40-45.
Seck D., 1991. Observations préliminaires sur les fluctuations saisonnières des populations d'insectes ravageurs du mil stocké en grenier traditionnel au Sénégal. Tropicultura, 9, 92-94.
Seck D., 1992. Importance économique et développement d'une approche de lutte intégrée contre les insectes ravageurs des stocks de maïs, de mil et de niébé en milieu paysan. In : La post-récolte en Afrique. Montréal, Canada : Aupelf-Uref ; Paris : John Libbey Eurotext, 155-160.
Seck D., 1993. Développement de méthodes alternatives de contrôle des principaux insectes ravageurs des denrées emmagasinées au Sénégal par l'utilisation de plantes indigènes. Thèse de doctorat : Faculté des Sciences agronomiques de Gembloux (Belgique).
Seck D., Sidibé B. & Fall A., 1992. Observations sur le développement de Tribolium castaneum Herbst sur mil (Pennisetum thyphoïdes L.), en fonction du taux de brisure dans le substrat. Mém. Soc. R. Belge Entomol., 35, 471-475.
Seck D. et al., 1993. Biological activity of the shrub Boscia senegalensis (Pers.) Lam ex Poir. (Capparaceae) on stored grain insects. J. Chem. Ecol., 19(2), 377-389.
Seck D. et al., 1996. Alternative protection of cowpea seeds against Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) using hermetic storage or in combination with Boscia senegalensis (Pers.) Lam ex Poir. (Capparaceae) on stored grain insects. J. Stored Prod. Res., 32(1), 39-44.
Sembène M. et al., 2006. Désinsectisation des stocks d'arachide infestés par Caryedon serratus (OL.) : utilisation de la solarisation en zone sahélienne. J. Sci., 6(1), 1-11.
Sulehrie M.A.Q., Golob P., Tran B.M.D. & Farrell G., 2003. The effect of attributes of Vigna spp. on the bionomics of Callosobruchus maculatus. Entomol. Exp. Appl., 106, 159-168.
Tapondjou L.A., Adler C., Bouda H. & Fontem D.A., 2002. Efficacy of powder and essential oil from Chenopodium ambrosioides leaves as post-harvest grain protectants against six-stored product beetles. J. Stored Prod. Res., 38, 395-402.
Thiaw C. et al., 2007. Ovicid and adulticid effects of powders and extracts of Calotropis procera AIT. and of Senna occidentalis L. on Caryedon serratus (Ol.) destroyer of groundnut stocks. J. Sci., 7(3), 1-15.
Vayias B.J. & Vassiliki K.S., 2009. Factors affecting the insecticidal efficacy of an enhanced diatomaceous earth formulation against three stored-product insect species. J. Stored Prod. Res., 45, 226-231.
Verstraeten C. & Haubruge E., 1987. Les processus de migration de Prostephanus truncatus Horn (Coleoptera : Bostrichidae) dans un stock de maïs égrené. Med. Fac. Landbouww. Rijksuniv. Gent, 52(2a), 235-239.
Vowotor K.A., Meikle W.G., Ayertey J.N. & Markham R.H., 2005. Distribution of and association between the larger grain borer Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) and the maize weevil Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae) in maize stores. J. Stored Prod. Res., 41, 498-512.
Zettler J.L. & Arthur F.H., 2000. Chemical control of stored product insects with fumigants and residual treatments. Crop Prot., 19, 577-582.
Para citar este artículo
Acerca de: Momar Talla Guèye
Institut de Technologie alimentaire. BP 2765. SN-Hann-Dakar (Sénégal). E-mail : gueyemt@gmail.com
Acerca de: Dogo Seck
Centre Régional de Recherche en Écotoxicologie et Sécurité environnementale (CERES/Locustox). Route de Rufisque, km 15. BP 3300. SN-Dakar (Sénégal).
Acerca de: Jean-Paul Wathelet
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Chimie analytique. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
Acerca de: Georges Lognay
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Chimie générale et organique. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
