


- Portada
- Volume 11 (2007)
- numéro 2
- Homologies entre les deux rétrovirus BLV et HTLV-1 et développement d’une nouvelle approche thérapeutique basée sur la levée de la latence virale


Vista(s): 5164 (67 ULiège)
Descargar(s): 110 (2 ULiège)

Homologies entre les deux rétrovirus BLV et HTLV-1 et développement d’une nouvelle approche thérapeutique basée sur la levée de la latence virale

Notes de la rédaction
Reçu le 3 avril 2007, accepté le 18 avril 2007.
Résumé
Les virus BLV (bovine leukemia virus) et HTLV-1 (human T-cell leukemia virus) sont des rétrovirus apparentés induisant leucémies (BLV et HTLV-1) ou maladies inflammatoires (HTLV-1) chez leur hôte que sont respectivement le bovin et l’humain. Ces maladies se caractérisent par une absence apparente d’expression virale dans beaucoup de cellules infectées in vivo. Une réponse immune humorale et cellulaire dirigée contre les antigènes viraux est continuellement présente. Ainsi, BLV et HTLV-1 ont développé une série de mécanismes régulant très finement leur expression, leur permettant d’échapper à la surveillance immune. Parmi ceux-ci, l’état de compaction de la chromatine joue un rôle primordial dans leur régulation transcriptionnelle. La condensation de la chromatine est régie par l’action antagoniste de deux familles d’enzymes : les HATs (histone acétyltransférases) et les HDACs (histone désacétylases) qui, respectivement, incorporent ou retirent les groupements acétyles des histones. L’utilisation d’inhibiteurs d’HDACs induit la décondensation de la chromatine et de ce fait, augmente la transcription du virus. Ainsi, les cellules infectées latentes sont forcées d’exprimer les protéines virales et sont détruites par la réponse immune. Cette stratégie d’activation de l’expression virale pourrait constituer une nouvelle approche thérapeutique contre les maladies induites par les virus BLV et HTLV-1.
Abstract
Homologies between BLV and HTLV-1 retroviruses and development of a novel therapeutic approach based on induction of viral expression. BLV (bovine leukemia virus) and HTLV-1 (human T-cell leukemia virus) are related retroviruses inducing leukemia (BLV and HTLV-1) or inflammatory diseases (HTLV-1) in the bovine and the human species respectively. The diseases are characterized by an apparent lack of viral expression in many infected cells in vivo. A humoral and a cellular immune response directed against viral antigens is continuously present. In fact, these two viruses have developed a series of mechanisms tightly controlling their expression, thereby allowing escape from the immune surveillance. Among these, chromatin compaction plays an essential role in the transcriptional regulation of the virus. The level of chromatin compaction is the result of an interplay between antagonistic actions of two families of enzymes: HATs (histone acetyltransferases) and HDACs (histone deacetylases) which, respectively, incorporate or withdraw histone acetyl groups. HDAC inhibitors induce a decondensation of chromatin and consequently increase the transcription. Thus, latent infected cells are forced to express viral proteins and are destroyed by the immune response. This viral gene activation therapy might be a novel approach for the treatment of BLV and HTLV-1 induced diseases.
Tabla de contenidos
1. Historique, épidémiologie et modes de transmission des virus BLV et HTLV-1
1.1. BLV
1Sur base de leur épidémiologie, Bendixen (1965) distingue deux types de leucémies bovines, l’une est contagieuse : la leucose bovine enzootique (LBE) et l’autre n’est pas associée à un agent pathogène : la leucose bovine sporadique (LBS). La présence de particules virales fut observée dans des cultures de lymphocytes d’animaux atteint de LBE (Miller et al., 1969) et la transmission horizontale de cette maladie fut réalisée par le transfert de cellules infectées (Mammerickx, 1972). Il fut ensuite démontré que l’agent infectieux, alors appelé BLV pour bovine leukemia virus, est un rétrovirus exogène à l’espèce bovine (Kettmann et al., 1976). La transmission naturelle s’avèrera se faire principalement par le lait (Ferrer, Piper, 1981).
1.2. HTLV-1
2En 1980, Poiesz et ses collègues réalisent pour la première fois le lien entre un rétrovirus et un cancer humain chez un patient atteint d’un lymphome cutané à cellules T (Poiesz et al., 1980). Quelques années auparavant, une entité clinique appelée ATL (pour adult T-cell leukemia, ou leucémie à cellules T de l’adulte) avait été décrite au Japon (Uchiyama et al., 1977). Il fut ensuite démontré que l’ATL était due à la présence d’un rétrovirus appelé ATLV (Hinuma et al., 1981 ; Yoshida et al., 1982), qui n’était autre que le virus mis en évidence par Poiesz et un nom unique, HTLV-1, fut retenu (Popovic et al., 1982). L’infection par HTLV-1 touche de 10 à 20 millions de personnes à travers le monde mais est restreinte géographiquement dans des zones endémiques telles que le Japon méridional, les îles des Caraïbes, l’Afrique centrale et l’Amérique du Sud. La transmission horizontale du virus dans des régions endémiques se réalise par l’allaitement maternel ou via des rapports sexuels. HTLV-1 peut également être transmis par transfusion sanguine ou consécutivement au partage d’aiguilles souillées permettant le transfert de cellules infectées vivantes. L’individu infecté est porteur du virus durant toute sa vie et 95 % des séropositifs restent porteurs asymptomatiques. Seulement 5 % des personnes infectées développent la maladie après une longue phase médicalement asymptomatique : 2 à 3 % développent une forme agressive et rapidement mortelle de leucémie (l’ATL), alors que 2 à 3 % développent des maladies inflammatoires, telles que l’uvéite, l’arthropathie, la myosite ou, le plus généralement, la HAM/TSP (pour HTLV-1 associated myelopathy/tropical spastic paraparesis ou myélopathie associée à HTLV-1/paraparésie spastique tropicale) (revu par Proietti et al., 2005).
3Comme nous allons le voir ci-après, les deux rétrovirus BLV et HTLV-1 sont très similaires quant à leur pathogenèse, leur structure génomique, et leurs interactions avec le système immunitaire de l’hôte. L’existence de virus humains apparentés au BLV, comme l’est HTLV-1, marque le début de l’intérêt pour le BLV comme modèle d’étude et de développement thérapeutique in vivo.
2. Pathologies associées aux virus
2.1. BLV
4Chez l’hôte naturel : le bovin. Les lymphocytes B constituent la cible essentielle du virus BLV dans le sang périphérique (Levy et al., 1987). Le provirus y est intégré en divers sites non préférentiels dans leur génome (Kettmann et al., 1980). Les bovins infectés par BLV peuvent présenter trois stades distincts : une phase asymptomatique, une lymphocytose persistante (LP) et une phase tumorale (revu par Willems et al., 2000).
5La phase asymptomatique correspond au premier stade de la maladie qui peut être considéré comme une phase de latence, la présence d’anticorps dirigés contre les protéines virales constituant une des rares manifestations de l’infection par BLV (Meiron et al., 1985). Cet état peut persister durant toute la vie de l’animal.
6La lymphocytose persistante (LP) se traduit par une augmentation du nombre de lymphocytes B circulants. Le rapport entre les cellules B et T est modifié (voire inversé) et, selon l’animal, la population de lymphocytes B peut représenter 40 à 90 % de la population lymphocytaire totale, contre 15 à 20 % chez un animal sain. La lymphocytose peut se stabiliser pendant de très longues périodes mais peut également progresser pour atteindre des valeurs très élevées ou même disparaître subitement.
7Enfin, une faible fraction des bovins infectés (< 5 %) développent une leucémie, un lymphome ou un lymphosarcome. Généralement, la phase tumorale survient chez les animaux en LP mais peut aussi se développer chez les bovins aleucémiques ne présentant aucun signe clinique (Ferrer et al., 1979). Le développement de tumeurs peut s’accompagner d’une augmentation du nombre de lymphocytes B circulants, allant jusqu’à 1 million de lymphocytes B par mm3 de sang (contre 3 à 5 mille lymphocytes par mm3 de sang chez un animal normal). Les tumeurs sont toujours issues d’une cellule lymphoïde de type B où le provirus est intégré en un (ou quelques) site(s) dans le génome cellulaire (Kettmann et al., 1980). Cette transformation maligne aboutit inexorablement à la mort de l’animal dans l’année.
8Chez un modèle animal : le mouton. Chez les ovins, la pathogenèse est comparable à celle des bovins en ce qui concerne la cellule cible (lymphocytes B), la faible expression virale et la phase tumorale. Mais elle présente également certaines différences et notamment, au niveau de la LP que ne développe pas le mouton. Chez ces animaux, le nombre de lymphocytes B infectés croît plus régulièrement après une période de latence de 1 à 5 ans. De plus, la phase tumorale atteint tous les ovins infectés au cours de leur existence (Mammerickx et al., 1987 ; 1988).
9Par ces différences, le mouton constitue un excellent système pour l’étude de la pathogenèse induite par le BLV.
2.2. HTLV-1
10La grande majorité des individus infectés par le virus HTLV-1 demeure des porteurs asymptomatiques, alors qu’une petite proportion développe la maladie après une longue période de latence (Uchiyama, 1997 ; Bangham, 2000).
11Leucémie à cellules T de l’adulte. L’ATL est une leucémie agressive à lymphocytes T caractérisée par une lympho-adénopathie, une hypercalcémie et par des lésions de la peau (Matsuoka, 2003). Les lymphocytes T leucémiques porteurs du provirus sont presque toujours CD4+, peuvent avoir un noyau multilobé et résultent d’une prolifération oligoclonale ou monoclonale (Yoshida et al., 1984). Les sites d’intégration de HTLV-1 dans le génome de la cellule semblent être aléatoires (Seiki et al., 1984 ; Zoubak et al., 1994) et n’influencent apparemment pas la pathogenèse. La période de survie médiane d’un individu diagnostiqué avec une ATL aiguë et progressive est de seulement 6 mois (Bangham, 2000 ; Matsuoka, 2003). La durée de survie médiane pour une ATL chronique est d’environ 2 ans. Après une réponse en première ligne, l’ATL devient fréquemment réfractaire aux traitements de chimiothérapie classique (cyclophosphamide, adriamycine, vincristine et prednisolone) (revu par Taylor et Matsuoka, 2005).
12HAM/TSP. L’HAM/TSP est une maladie inflammatoire chronique du système nerveux central caractérisée par une démarche altérée, une lombalgie, un dysfonctionnement urinaire et une faiblesse spastique progressive des membres inférieurs. L’infiltration de lymphocytes T CD4+ infectés dans le système nerveux central est très probablement la cause des facteurs inflammatoires produits par l’hôte. Le développement de la maladie est fortement variable entre individus. L’HAM/TSP peut se développer rapidement, progresser très lentement pendant des décennies ou même arrêter d’évoluer dans certains cas. L’âge moyen au début de l’HAM/TSP est de 44 ans. La maladie affecte davantage les femmes (rapport homme/femme de 1 pour 2, l’inverse de l’ATL). Il n’y a actuellement aucun traitement satisfaisant (revu par Proietti et al., 2005).
13Comme nous venons de le voir ici, les deux rétrovirus BLV et HTLV-1 ciblent tous les deux des lymphocytes. Chez leur hôte naturel, ils n’induisent des pathologies que dans environ 5 % des individus infectés et ceci après une longue période asymptomatique. Ces deux virus sont leucémogènes (provoquant la LBE ou l’ATL) mais seul HTLV-1 semble responsable du développement de maladies inflammatoires.
3. Virion et provirus
14Les virus BLV et HTLV-1 appartiennent à la famille des rétrovirus et au genre des delta-rétrovirus caractérisés par une structure génomique complexe (région « X ») (revu par Willems et al., 2000).
3.1. Structure générale du virion
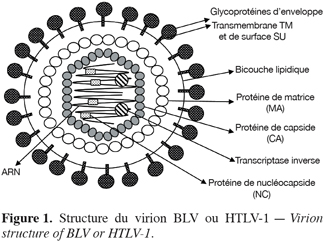
15Les génomes de BLV et de HTLV-1 sont constitués de deux molécules d’ARN identiques, chacune associée à la protéine de nucléocapside ainsi qu’à la transcriptase inverse. Ce complexe ribonucléoprotéique est entouré par une capside (constituée des protéines CA) qui est reliée à l’enveloppe virale par les protéines de matrice (protéines MA) (Figure 1). Cette enveloppe, d’origine cellulaire, est acquise lors du bourgeonnement du virus à la surface de la cellule productrice. Elle est constituée d’une bicouche phospholipidique dans laquelle sont insérés les complexes glycoprotéiques d’enveloppe TM et SU d’origine virale qui interviennent dans l’attachement du virus à la surface de la cellule cible.
3.2. Structure générale du provirus
16Le génome viral (8,7 kb pour BLV ou 9,0 kb pour HTLV-1) comporte les gènes structuraux classiques des rétrovirus (gag, prt, pol, env) auxquels s’ajoute une région supplémentaire, la région « X », codant pour les protéines de régulation (Figure 2). Il présente à chaque extrémité une séquence redondante appelée long terminal repeat (LTR). Les LTRs contiennent des éléments de régulation nécessaires à l’expression. Bien que les deux LTRs aient une structure identique, leur fonction est toutefois différente. Le LTR5’ comprend les promoteurs nécessaires à la régulation du niveau d’initiation de la transcription en interagissant avec des facteurs spécifiques. Tandis que le LTR3’ contient les séquences nécessaires à l’addition d’une queue poly A aux ARNs messagers viraux. Il en résulte trois principaux transcrits d’ARN messager : doublement épissé, simplement épissé et non épissé.
17L’ARN messager non épissé code pour les protéines de structure Gag (MA, CA, NC) et pour les enzymes Prt (protéase) et Pol (polymérase). Les protéines Prt et Pol sont exprimées comme protéines de fusion Gag-Prt et Gag-Prt-Pol. Prt se sépare de manière autocatalytique du précurseur Gag-Prt-Pol et est responsable du clivage de maturation de Gag et Pol. Pol a trois domaines fonctionnels que sont la transcriptase inverse, la RNaseH et l’intégrase. La transcriptase inverse (ADN polymérase-ARN dépendante) permet la transcription de l’ARN viral en ADN proviral ; la RNAse H dégrade l’ARN de la molécule hybride ARN/ADN présente au cours de la rétrotranscription et l’intégrase est nécessaire pour l’intégration du provirus dans l’ADN cellulaire.
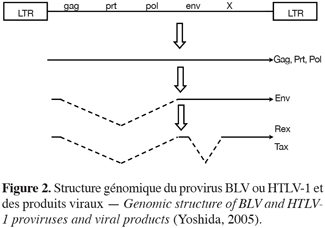
18L’ARN messager simplement épissé code pour l’enveloppe. Env est une grande glycoprotéine qui génère, après clivage, les deux glycoprotéines d’enveloppe : SU, qui est une protéine de surface fortement glycosylée impliquée dans la reconnaissance du récepteur cellulaire et TM, qui est une protéine transmembranaire ancrant le complexe SU-TM dans la membrane virale d’origine cellulaire (Schultz et al., 1984 ; Johnston, Radke, 2000 ; Gatot et al., 2002 ; Johnston et al., 2002).
19L’ARNm doublement épissé code pour deux protéines de régulation : Tax et Rex. Tax est une protéine phosphorylée à localisation nucléaire. Elle est impliquée dans la régulation de la transcription virale : elle accroît la transcription des gènes viraux via des séquences activatrices présentes dans le LTR5’. Tax ne se lie toutefois pas directement à ces séquences mais agit par l’intermédiaire de facteurs cellulaires (cf. paragraphe 4). Rex est une phosphoprotéine nucléaire intervenant en tant que régulateur post-transcriptionnel. Cette protéine stabilise et permet l’export vers le cytoplasme des ARNs génomiques et des ARNs messagers codant pour les protéines structurales Gag, Prt, Pol et Env.
4. Réponse immune et régulation de l’expression virale
20Les pathologies induites par les virus BLV ou HTLV-1 se développent alors qu’aucun virion, aucun ARNm ou aucune protéine virale n’est détectée dans la majorité des cellules sanguines infectées. La détection de l’expression virale ne peut se faire que par des techniques très sensibles de RT-PCR (reverse transcriptase polymerase chain reaction). Cependant, même si ces virus paraissent silencieux, on observe chez les sujets ou animaux infectés une réponse immune efficace et continue dirigée contre les épitopes viraux. Cette réponse immune se caractérise par la présence d’anticorps et de lymphocytes cytotoxiques dirigés contre les antigènes viraux (revu par Bangham, Osame, 2005 et par Florins et al., 2007). Pour échapper à cette surveillance immune permanente ces virus ont élaboré des stratégies de régulation fine de leur expression dans lesquelles la protéine Tax joue un rôle majeur (Willems et al., 2000 ; Kashanchi, Brady, 2005).
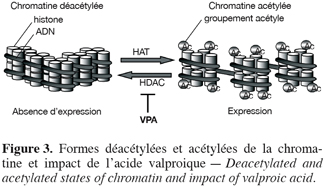
21Pour comprendre les mécanismes par lesquels Tax régule la transcription il est important de préciser que l’expression de gènes peut être régulée suite à la variation de l’état de condensation de la chromatine. Cet état est régi par l’action antagoniste de deux familles d’enzymes : les HATs (pour histone acétyltransférases) et les HDACs (pour histone désacétylases) qui, respectivement, incorporent ou retirent les groupements acétyles des résidus lysines des queues d’histones (Figure 3). L’enlèvement par les HDACs des groupements acétyles restaure une charge positive sur les histones et ainsi augmente leur affinité vis-à-vis de l’ADN, mène à la compaction de la chromatine et inhibe la transcription. A l’inverse, les HATs acétylent les résidus lysine, diminuent la compaction et permettent la transcription (revu par Marks et al., 2003).
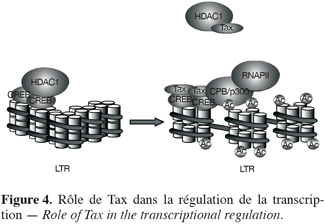
22Ainsi, les protéines Tax ne se lient pas directement à l’ADN mais recrutent plusieurs partenaires cellulaires pour activer la transcription virale (Figure 4). Tax se lie au LTR via les protéines CREB (pour cyclic AMP-responsive element binding protein) (Adam et al., 1994) et provoque le détachement de HDAC1 (pour histone désacétylase 1) (Ego et al., 2002 ; Lu et al., 2004). Tax formera alors un complexe protéique incluant notamment l’acétyltransférase p300/CBP (pour CREB binding protein) et la RNA polymerase II (Azran et al., 2004 ; Lu et al., 2004). De cette façon, Tax modifie l’état de la chromatine, permettant sa décondensation et ainsi la transcription des gènes situés en aval.
5. Thérapie basée sur la levée de la latence virale
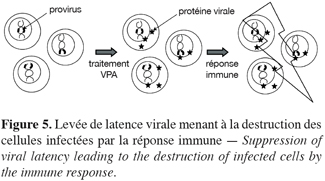
23L’acide valproique (VPA), utilisé depuis des décennies pour le traitement à long terme de l’épilepsie (Henry, 2003), est un inhibiteur d’HDACs (Gottlicher et al., 2001 ; Phiel et al., 2001). Le VPA présente une faible toxicité chez l’adulte et avec une demi-vie de 16-17 heures, a des propriétés pharmacocinétiques adéquates in vivo (Wong et al., 2001 ; Blaheta, Cinatl Jr., 2002 ; Blaheta et al., 2002). Les inhibiteurs d’HDACs favorisent l’acétylation de la chromatine et augmentent dès lors potentiellement l’expression génique (Drummond et al., 2005 ; Monneret, 2005) (Figure 3). Ainsi, le VPA induit l’expression des virus BLV et HTLV-1 en cultures cellulaires (Merezak et al., 2002 ; Achachi et al., 2005). Un traitement par injection intraveineuse de VPA à des moutons leucémiques infectés par le BLV a conduit à une réduction du nombre de cellules malignes dans le sang circulant de ces animaux (Achachi et al., 2005). De la même manière, un traitement au VPA administré à des patients infectés par HTLV-1 et atteints de HAM/TSP a entraîné une importante diminution de la charge virale. Bien que le mécanisme incriminé ne soit pas formellement démontré, il nous semble que le mode d’action du VPA repose sur l’activation de l’expression virale et la destruction des cellules infectées par le système immunitaire (Figure 5). Toutefois, un mécanisme complémentaire reposant sur l’induction de la mort programmée des cellules infectées (apoptose) n’est pas à exclure.
6. Conclusion
24Dans cet article, nous avons présenté une comparaison entre deux rétrovirus apparentés : BLV et HTLV-1. Nous avons aussi rendu compte d’une nouvelle approche thérapeutique basée sur la levée de latence virale. Nous l’avons développée à partir d’essais in vitro qui ont été étendus par la suite à l’expérimentation animale grâce au modèle de moutons infectés par le BLV et enfin appliquée dans le cadre d’un essai clinique à des patients infectés par HTLV-1.
25Remerciements
26Nous remercions le Fonds national de la Recherche scientifique (FNRS), le Télévie, le 6e programme cadre INCA de la Commission européenne, la Fondation belge contre le cancer, la Fondation Bekales et le CGRI (Commissariat général des relations internationales) pour le support financier ainsi que toutes les personnes qui ont collaboré aux études menées sur l’acide valproique et son application thérapeutique : Becca Asquith, Catherine Burteau, Charles Bangham, Hervé Balon, Gilda Belrose, Mathieu Boxus, Arsène Burny, Arnaud Florins, Agnès Lézin, Annamaria Nigro, Angelina Mosley, Patrice Urbain, Fabian Vandermeers et Olivier Verlaeten. Nicolas Gillet (boursier Télévie), Richard Kettmann (Directeur de Recherches) et Luc Willems (Directeur de Recherches) sont membres du FNRS.
Bibliographie
Achachi A., Florins A., Gillet N., Debacq C., Urbain P., Foutsop GM., Vandermeers F., Jasik A., Reichert M., Kerkhofs P., Lagneaux L., Burny A., Kettmann R., Willems L. (2005). Valproate activates bovine leukemia virus gene expression, triggers apoptosis, and induces leukemia/lymphoma regression in vivo. Proc. Natl Acad. Sci. USA 102 (29), p. 10309–10314.
Adam E., Kerkhofs P., Mammerickx M., Kettmann R., Burny A., Droogmans L., Willems L. (1994). Involvement of the cyclic AMP-responsive element binding protein in bovine leukemia virus expression in vivo. J. Virol. 68 (9), p. 5845–5853.
Azran I., Schavinsky-Khrapunsky Y., Aboud M. (2004). Role of Tax protein in human T-cell leukemia virus type-I leukemogenicity. Retrovirology 1 (1), p. 20–44.
Bangham CR. (2000). HTLV-1 infections. J. Clin. Pathol. 53 (8), p. 581–586.
Bangham CR., Osame M. (2005). Cellular immune response to HTLV-1. Oncogene 24 (39), p. 6035–6046.
Bendixen HJ. (1965). Bovine enzootic leukosis. Adv. Vet. Sci. 10, p. 129–204.
Blaheta RA., Cinatl J. Jr. (2002). Anti-tumor mechanisms of valproate: a novel role for an old drug. Med. Res. Rev. 22 (5), p. 492–511.
Blaheta RA., Nau H., Michaelis M., Cinatl J. Jr. (2002). Valproate and valproate-analogues: potent tools to fight against cancer. Curr. Med. Chem. 9 (15), p. 1417–1433.
Drummond DC., Noble CO., Kirpotin DB., Guo Z., Scott GK., Benz CC. (2005). Clinical development of histone deacetylase inhibitors as anticancer agents. Annu. Rev. Pharmacol. Toxicol. 45, p. 495–528.
Ego T., Ariumi Y., Shimotohno K. (2002). The interaction of HTLV-1 Tax with HDAC1 negatively regulates the viral gene expression. Oncogene 21 (47), p. 7241–7246.
Ferrer JF., Marshak RR., Abt DA., Kenyon SJ. (1979). Relationship between lymphosarcoma and persistent lymphocytosis in cattle: a review. J. Am. Vet. Med. Assoc. 175 (7), p. 705–708.
Ferrer JF., Piper CE. (1981). Role of colostrum and milk in the natural transmission of the bovine leukemia virus. Cancer Res. 41 (12), p. 4906–4909.
Florins A., Gillet N., Asquith B., Boxus M., Burteau C., Twizere JC., Urbain P., Vandermeers F., Debacq C., Sanchez-Alcaraz MT., Schwartz-Cornil I., Kerkhofs P., Jean G., Thewis A., Hay J., Mortreux F., Wattel E., Reichert M., Burny A., Kettmann R., Bangham C., Willems L. (2007). Cell dynamics and immune response to BLV infection: a unifying model. Front Biosci. 12, p. 1520–1531.
Gatot JS., Callebaut I., Van Lint C., Demonte D., Kerkhofs P., Portetelle D., Burny A., Willems L., Kettmann R. (2002). Bovine leukemia virus SU protein interacts with zinc, and mutations within two interacting regions differently affect viral fusion and infectivity in vivo. J. Virol. 76 (16), p. 7956–7967.
Gottlicher M., Minucci S., Zhu P., Kramer OH., Schimpf A., Giavara S., Sleeman JP., Lo CF., Nervi C., Pelicci PG., Heinzel T. (2001). Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. EMBO J. 20 (24), p. 6969–6978.
Henry TR. (2003). The history of valproate in clinical neuroscience. Psychopharmacol. Bull. 37 (2), p. 5–16.
Hinuma Y., Nagata K., Hanaoka M., Nakai M., Matsumoto T., Kinoshita KI., Shirakawa S., Miyoshi I. (1981). Adult T-cell leukemia: antigen in an ATL cell line and detection of antibodies to the antigen in human sera. Proc. Natl Acad. Sci. USA 78 (10), p. 6476–6480.
Johnston ER., Radke K. (2000). The SU and TM envelope protein subunits of bovine leukemia virus are linked by disulfide bonds, both in cells and in virions. J. Virol. 74 (6), p. 2930–2935.
Johnston ER., Albritton LM., Radke K. (2002). Envelope proteins containing single amino acid substitutions support a structural model of the receptor-binding domain of bovine leukemia virus surface protein. J. Virol. 76 (21), p. 10861–10872.
Kashanchi F., Brady JN. (2005). Transcriptional and post-transcriptional gene regulation of HTLV-1. Oncogene 24 (39), p. 5938–5951.
Kettmann R., Portetelle D., Mammerickx M., Cleuter Y., Dekegel D., Galoux M., Ghysdael J., Burny A., Chantrenne H. (1976). Bovine leukemia virus: an exogenous RNA oncogenic virus. Proc. Natl Acad. Sci. USA 73 (4), p. 1014–1018.
Kettmann R., Cleuter Y., Mammerickx M., Meunier-Rotival M., Bernardi G., Burny A., Chantrenne H. (1980). Genomic integration of bovine leukemia provirus: comparison of persistent lymphocytosis with lymph node tumor form of enzootic. Proc. Natl Acad. Sci. USA 77 (5), p. 2577–2581.
Levy D., Kettmann R., Marchand P., Djilali S., Parodi AL. (1987). Selective tropism of bovine leukemia virus (BLV) for surface immunoglobulin-bearing ovine B lymphocytes. Leukemia 1 (5), p. 463–465.
Lu H., Pise-Masison CA., Linton R., Park HU., Schiltz RL., Sartorelli V., Brady JN. (2004). Tax relieves transcriptional repression by promoting histone deacetylase 1 release from the human T-cell leukemia virus type 1 long terminal repeat. J. Virol. 78 (13), p. 6735–6743.
Mammerickx M. (1972). La transmission horizontale et verticale de la leucose bovine enzootique, premiers résultats après cinq ans d’expérimentation. Ann. Med. Vet. 116, p. 647–665.
Mammerickx M., Portetelle D., de Clercq K., Burny A. (1987). Experimental transmission of enzootic bovine leukosis to cattle, sheep and goats: infectious doses of blood and incubation period of the disease. Leuk. Res. 11 (4), p. 353–358.
Mammerickx M., Palm R., Portetelle D., Burny A. (1988). Experimental transmission of enzootic bovine leukosis to sheep: latency period of the tumoral disease. Leukemia 2 (2), p. 103–107.
Marks PA., Miller T., Richon VM. (2003). Histone deacetylases. Curr. Opin. Pharmacol. 3 (4), p. 344–351.
Matsuoka M. (2003). Human T-cell leukemia virus type I and adult T-cell leukemia. Oncogene 22 (33), p. 5131–5140.
Meiron R., Brenner J., Gluckman A., Avraham R., Trainin Z. (1985). Humoral and cellular responses in calves experimentally infected with bovine leukemia virus (BLV). Vet. Immunol. Immunopathol. 9 (2), p. 105–114.
Merezak C., Reichert M., Van Lint C., Kerkhofs P., Portetelle D., Willems L., Kettmann R. (2002). Inhibition of histone deacetylases induces bovine leukemia virus expression in vitro and in vivo. J. Virol. 76 (10), p. 5034–5042.
Miller JM., Miller LD., Olson C., Gillette KG. (1969). Virus-like particles in phytohemagglutinin-stimulated lymphocyte cultures with reference to bovine lymphosarcoma. J. Natl Cancer Inst. 43 (6), p. 1297–1305.
Monneret C. (2005). Histone deacetylase inhibitors. Eur. J. Med. Chem. 40 (1), p. 1–13.
Phiel CJ., Zhang F., Huang EY., Guenther MG., Lazar MA., Klein PS. (2001). Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen. J. Biol. Chem. 276 (39), p. 36734–36741.
Poiesz BJ., Ruscetti FW., Gazdar AF., Bunn PA., Minna JD., Gallo RC. (1980). Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl Acad. Sci. USA 77 (12), p. 7415–7419.
Popovic M., Reitz MS. Jr., Sarngadharan MG., Robert-Guroff M., Kalyanaraman VS., Nakao Y., Miyoshi I., Minowada J., Yoshida M., Ito Y., Gallo RC. (1982). The virus of Japanese adult T-cell leukaemia is a member of the human T-cell leukaemia virus group. Nature 300 (5887), p. 63–66.
Proietti FA., Carneiro-Proietti AB., Catalan-Soares BC., Murphy EL. (2005). Global epidemiology of HTLV-I infection and associated diseases. Oncogene 24 (39), p. 6058–6068.
Schultz AM., Copeland TD., Oroszlan S. (1984). The envelope proteins of bovine leukemia virus: purification and sequence analysis. Virology 135 (2), p. 417–427.
Seiki M., Eddy R., Shows TB., Yoshida M. (1984). Nonspecific integration of the HTLV provirus genome into adult T-cell leukaemia cells. Nature 309 (5969), p. 640–642.
Taylor GP., Matsuoka M. (2005). Natural history of adult T-cell leukemia/lymphoma and approaches to therapy. Oncogene 24 (39), p. 6047–6057.
Uchiyama T. (1997). Human T cell leukemia virus type I (HTLV-I) and human diseases. Annu. Rev. Immunol. 15, p. 15–37.
Uchiyama T., Yodoi J., Sagawa K., Takatsuki K., Uchino H. (1977). Adult T-cell leukemia: clinical and hematologic features of 16 cases. Blood 50 (3), p. 481–492.
Willems L., Burny A., Collete D., Dangoisse O., Dequiedt F., Gatot JS., Kerkhofs P., Lefebvre L., Merezak C., Peremans T., Portetelle D., Twizere JC., Kettmann R. (2000). Genetic determinants of bovine leukemia virus pathogenesis. AIDS Res. Hum. Retroviruses 16 (16), p. 1787–1795.
Wong H., Rurak DW., Kumar S., Kwan E., Abbott FS., Riggs KW. (2001). Dose-dependent pharmacokinetics and metabolism of valproic acid in newborn lambs and adult sheep. Drug Metab. Dispos. 29 (5), p. 664–675.
Yoshida M., Miyoshi I., Hinuma Y. (1982). Isolation and characterization of retrovirus from cell lines of human adult T-cell leukemia and its implication in the disease. Proc. Natl Acad. Sci. USA 79 (6), p. 2031–2035.
Yoshida M., Seiki M., Yamaguchi K., Takatsuki K. (1984). Monoclonal integration of human T-cell leukemia provirus in all primary tumors of adult T-cell leukemia suggests causative role of human T-cell leukemia virus in the disease. Proc. Natl Acad. Sci. USA 81 (8), p. 2534–2537.
Zoubak S., Richardson JH., Rynditch A., Hollsberg P., Hafler DA., Boeri E., Lever AM., Bernardi G. (1994). Regional specificity of HTLV-I proviral integration in the human genome. Gene 143 (2), p. 155–163.
Para citar este artículo
Acerca de: Nicolas Gillet
Faculté universitaire des Sciences agronomiques de Gembloux. Unité de Biologie cellulaire et moléculaire. Avenue Maréchal Juin, 13. B-5030 Gembloux (Belgique). E-mail : gillet.n@fsagx.ac.be
Acerca de: Richard Kettmann
Faculté universitaire des Sciences agronomiques de Gembloux. Unité de Biologie cellulaire et moléculaire. Avenue Maréchal Juin, 13. B-5030 Gembloux (Belgique).
Acerca de: Luc Willems
Faculté universitaire des Sciences agronomiques de Gembloux. Unité de Biologie cellulaire et moléculaire. Avenue Maréchal Juin, 13. B-5030 Gembloux (Belgique).
