


- Portada
- volume 16 (2012)
- numéro 1
- Ecological review of black-grass (Alopecurus myosuroides Huds.) propagation abilities in relationship with herbicide resistance


Vista(s): 5760 (54 ULiège)
Descargar(s): 403 (7 ULiège)

Ecological review of black-grass (Alopecurus myosuroides Huds.) propagation abilities in relationship with herbicide resistance

Notes de la rédaction
Received on May 6, 2010; accepted on November 14, 2011
Résumé
Revue de l'écologie des capacités de propagation du vulpin des champs (Alopecurus myosuroides Huds.) en relation avec la résistance aux herbicides. Alopecurus myosuroides Huds. (vulpin des champs) a toujours été une préoccupation majeure pour les céréaliculteurs et le développement de la résistance aux herbicides n'améliore pas la situation. Cet article de synthèse résume les différents critères impliqués dans les modèles de dispersion d’individus de vulpin des champs au sein d’un champ ou d’une population sensible. Pour ce faire, le cycle complet du vulpin est décrit de la semence à la semence. Depuis le développement végétatif précoce jusqu’à la chute de la graine, chaque étape est décrite en prenant en compte comment la résistance aux herbicides peut influencer ou exercer un impact différent par rapport à des plantes sensibles.
Abstract
Alopecurus myosuroides Huds. (black-grass) has always been a major concern for cereal growers, and the development of herbicide resistance does not improve the situation. This review article summarizes the different traits involved in the dispersal pattern of herbicide resistant black-grass individuals within a susceptible field population. Therefore, the whole life cycle of black-grass is depicted from the seed to the seed. From the early vegetative development to the seed falling, every stage is described, taking into account how herbicide resistance can influence or exert a different impact compared to susceptible plants.
Tabla de contenidos
1. Introduction
1Alopecurus myosuroides Huds. (black-grass) is one of the commonest grass weeds of winter cereals in North-Western Europe. The control of this weed has always been a major concern for farmers, particularly because of its high reproduction rate and its strong competition towards the crops. In order to tackle its proliferation and limit the dispersal of this grass weed, cereal growers used to spray large quantities of herbicides. But soon herbicide resistant plants developed, thus inducing a new source of worries. Furthermore, the spread of herbicide resistance, combining genetic evolution and seed dispersal, has important consequences for the sustainability of cereal production.
2This review article aims to gather and explain the different traits involved in the dispersal pattern of black-grass individuals within a field population. Therefore, the whole life cycle of black-grass is depicted here from the seed to the seed.
3Before aiming to contribute to the next generation and get a positive fitness, the plant has first to grow in its environment, in the best way to produce the maximum of flowers that might be pollinated. It implies a good germination, correct early vegetative development and sufficient height to struggle with neighboring plants for light and nutrient, and in this case of a gramineous plant, a large number of tillers carrying fertile ears. Then, for this anemochorous species, a lot of factors are involved in pollen spread addition to wind force and relative humidity, such as ears’ height but also crop’s height. Indeed, it is in this particular boundary layer above the crop cover, that wind takes over the pollen to carry it to the next fertile ear. Afterwards, the plant has to provide to its offspring, the best conditions to survive once they will be released within the environment, carried by the wind, animals or simply falling to the ground. And thereafter starts the cycle again.
4As a matter of fact, herbicide resistance is usually not considered as an induced mechanism in the plant but more likely as a heritable trait, transmitted through generations (Letouzé et al., 2001). Therefore, we can assume that the resistance gene(s) can be conveyed by the pollen. The brackets for the “s” of gene are due to the fact that we still do not know exactly neither how many, nor which genes are involved in resistance, especially for enhanced metabolism resistance. In the case of target-site resistance, due to a single mutation in the gene coding for the enzyme targeted by the herbicide, it is simple as only one gene seems to be responsible for the resistance (e.g. Délye et al., 2007; Marshall et al., 2008).
5Dispersal is one of the central processes in the dynamics and evolution of plant populations. The spatial dynamics of plant populations are determined to a large degree by the movement of seeds. At regional scales, seed dispersal ranges will set the possibilities for colonization of new sites (Ouborg et al., 1999). A better understanding of these dispersal patterns, linked to herbicide resistance, is crucial in order to implement good strategies to limit and control the extension of herbicide resistance within the fields or even over a larger area.
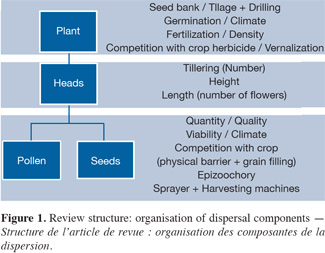
6Figure 1 presents the structure of this review paper, from the seed to the seed. Mother plant has first to germinate, to start vegetative growth, then reproductive growth, which gives some heads. These will emit and receive a certain amount of pollen, and then produce seeds that will finally fall back down to the soil.
2. Vegetative development
7Several characteristics are involved in the propagation process of one plant. Before the pollen emission and the seed shedding which are the point of this paper, the plant has first to develop. Therefore, seeds have to be present in the field’s seed bank, the dormancy has to be lifted, and then the tillage and the drilling have to be performed so as to allow the seeds to germinate in good conditions (Colbach et al., 2002; Colbach et al., 2003). Climatic conditions are very important during the whole development cycle of the plant, and especially in the early stages with the soil humidity and temperature. The intra and inter-specific competition (Chauvel et al., 2005) with the crop is really crucial during the first days. Hereafter, each factor conditioning the good vegetative development is described.
2.1. Dormancy
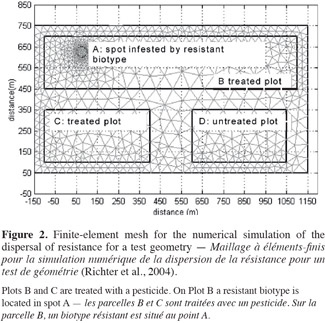
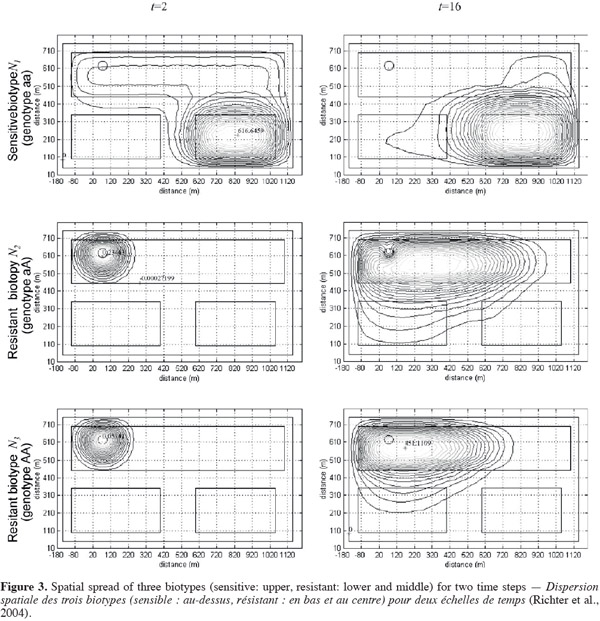
8Variation in primary dormancy among populations is a common feature in wild plant species and has been explained, for example, by differing weather conditions during seed maturation, light environment of the mother plant and genetic differences. Duration of primary dormancy indicates to what extent fresh seeds will germinate the first autumn after dispersal. Appreciable variation in initial dormancy among different collections was shown for A. myosuroides, with germination percentage ranging from 15% to 68% in light at normal autumn temperature (16 ⁄ 6°C) (Andersson et al., 2009). Colbach et al. (2002) demonstrated a similar variation among seed collections, with germination of fresh black-grass seeds ranging from 38% to 70%. Swain et al. (2006) found large variations over years in A. myosuroides and concluded that variations could be related to the temperature during seed maturation on the mother plant. The humidity levels to which plants were exposed during panicle formation and seed maturation affected the dormancy of seeds produced. Lower relative humidity was associated with a higher proportion of non-dormant seeds, and vice versa. Nevertheless, temperature during seed maturation had generally a greater effect on seed dormancy than soil moisture (Swain et al., 2006). There is less initial dormancy in seeds produced under warm and dry conditions, than under cool and wet ones.
9Froud-Williams et al. (1984a) demonstrated that seeds did not enter in a deep secondary dormancy during the summer. Indeed, germination of A. myosuroides was promoted by Red and Far-Red during the autumn of sowing, but germination only occurred in response to high levels of Pfr (physiologically active form of phytochrome) during the spring. However, in the subsequent autumn increased sensitivity to low levels of Pfr was regained. This is consistent with a total loss of dormancy during periods of natural emergence. Re-imposition of dormancy and light sensitivity confers a survival advantage to seeds by preventing germination at times hazardous for establishment.
2.2. Germination
10A. myosuroides populations commonly display two distinct emerging cohorts, a major one during autumn, and a minor one during spring (Barralis, 19701 cited in Menchari et al., 2007). So, it can develop either in winter crops, such as winter wheat or rape, or in spring crops, like sugar beet or maize. This ability, which limits the efficiency of control by rotation, occurs simultaneously with a change in the rate of leaf appearance due to sensitivity towards combined environmental factors (Chauvel et al., 2000) in order to complete its life cycle in a shorter period. Even in winter crops, a second range of germination can occur after the winter, though in a rather limited proportion. Therefore, it is really important to control the weed population as well as possible to avoid an overwhelming increase of black-grass in the seed bank. Indeed, with more or less 500 viable seeds produced by one plant, the expansion can be really fast (Moss, 1985).
11In case of autumn germination, plants have to grow sufficiently before the first low temperatures to resist the rudeness of winter. Moreover, they benefit from these cold temperatures to fill the need of vernalization. Emerging roughly at the same time as the crop, black-grass can synchronize with the crop cycle. Thus, weeds can grow together with it to be able to compete for light. As with other annual weeds, A. myosuroides must complete its life cycle earlier than the crop.
12In case of spring germination, plants get a better climate as soon as their emergence. They can start really fast and develop in good conditions and catch up with the crop. On the other hand, they are not vernalized enough, but the sums of temperature are more rapidly completed. The higher appearance rate of black-grass leaves for early spring germination facilitates a faster development and an earlier seed production in spring cereals (Chauvel et al., 2000). It has been suggested that the autumn and spring cohorts could result from a selection of two different biotypes with different germination and flowering requirements (Wellington et al., 19662 cited in Chauvel et al., 2002).
13The effect of burial on germination of A. myosuroides seeds has been shown by several authors (i.e. Froud-Williams et al., 1984b; Cussans et al., 1996). Froud-Williams et al. (1984b) showed that A. myosuroides emerged only from relatively shallow depth (< 50 mm), with an optimum at 0-20 mm. At the soil surface, emergence occurs mainly in autumn, and in case of soil cultivation practices, a smaller proportion can also germinate during the following spring. When buried at 5 cm depth, seeds did not germinate until the following year.
2.3. Vernalization
14The vernalization is one of the main factors that reduced the vegetative period and enhanced flowering induction. Chauvel et al. (2002) showed that vernalized plant apices were at the double-ridge stage after 670 day-degrees (base 0°C), whereas at the same time, all unvernalized plant apices were still at the vegetative stage. If there was no vernalization, the reproductive process was induced later, and 12% of the plants remained vegetative after 1,200 day-degrees. In addition, vernalized plants appear to be morphologically different (taller, producing fewer tillers but with more panicles emerging earlier) from unvernalized ones.
2.4. Phyllochron
15Figuring the thermal time between the initiation of two successive leaves on the main stem (expressed in day-degrees), the phyllochron of A. myosuroides does not seem significantly to be affected by vernalization. Mean values of the phyllochron were 90 day-degrees (base 0°C). According to the sowing dates, phyllochron values vary from 60 to 99 day-degrees per leaf, whereas tiller developmental rates vary from 66 to 95 day-degrees. The combined action of cumulative radiation and photoperiod can modify phyllochron values. Day length and vernalization appeared to have cumulative effects on final leaf number and flowering time. The rate of development of leaves within A. myosuroides appears to be faster than that observed for wheat or barley. This allows the weed to complete its life cycle before the harvest (Chauvel et al., 2002).
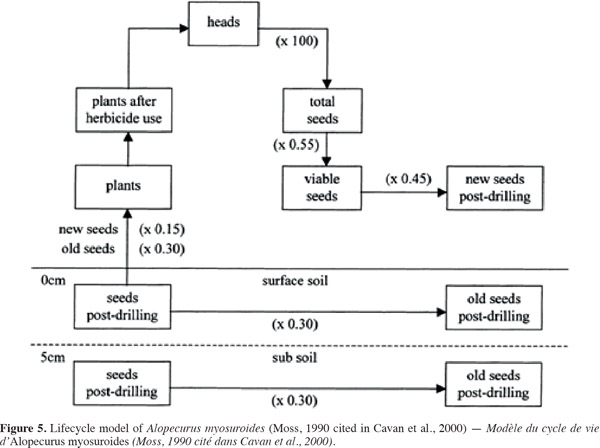
16There is a variation in leaf appearance with sowing date which cannot solely be related to temperature. Certainly, as for cultivated plants, the black-grass phyllochron may be influenced by a number of factors such as water, soil compaction or light. Kirby et al. (1985) demonstrated that leaf appearance rate varies with sowing date, and that leaves of late sown crop plants generally appear more rapidly. Leaf emergence rate seems to be determined early in the plant life cycle, and is dependent on environmental conditions during the emergence stage. Influence of day length has also been demonstrated for wheat, and other authors have suggested that use of a photothermal scale can improve leaf number prediction (Masle et al., 19893 cited in Chauvel et al., 2000). Global solar radiation seems to be one of the main factors explaining phyllochron variation (Chauvel et al., 2000).
2.5. Intra-specific competition
17Density effects due to intra-specific competition are very strong on the growth and vegetative development of blackgrass. Chauvel et al. (2005) worked on blackgrass plots, sowed in two densities (D1-D2) and with two nitrogen supply (N1-N2), to assess the plants’ development when exposed to intra-specific competition. The high density (D2) of A. myosuroides affected plant morphology; plants in D2 density were higher than those in D1 density. The high density (D2) affected negatively the shoot weight and the total number of tillers: this tendency tends to be more important with the higher nitrogen supply (N2). Without light competition, nitrogen influenced positively A. myosuroides growth. Early nitrogen deficiencies reduced early growth and components such as tillering (Chauvel et al., 19964 cited in Chauvel et al., 2005). A. myosuroides suppression by the crop was bigger at high nitrogen level. These results indicated that this weed’s development and growth were strongly influenced by intra-species competition. Unlike Lolium rigidum, A. myosuroides did not appear as a strong competitive weed (Chauvel et al., 2005).
3. Flowering stage
18The number of heads per plant is conditioned by the number of tillers. Sometimes, in greenhouse conditions, the number of tillers can rise up to more than 100 per plant (Chauvel et al., 2005). Their height is crucial to guarantee good pollen propagation at first, but also for the seed shedding. Indeed, if the apex of the ear is fairly high and above the crop cover, there is no physical barrier for the wind to carry the pollen, and for the seed to fall to the ground at a longer distance.
19The dispersal unit of A. myosuroides is the whole spikelet which is single-flowered. The head length (HL) is known to be correlated to the total number of spikelets (SNH, Spikelets Number per Head): SNH = 1.74 HL - 35.97 (R² = 0.86) (Dalbiès-Dulout et al., 2001). Chauvel’s results (2005) indicated that inflorescence length is more likely influenced by nitrogen availability, while the number of spikelets by inflorescence is not modified. As low differences were observed on inflorescence length, seed production differences were especially linked to the tiller number. A. myosuroides plant under reduced light produced less mature inflorescence. Roché et al. (1994) have shown for another weed, Centaurea solstitialis L., that reduction of light could decrease significantly the reproductive capacity, for example the number of flowers. This could be explained by the fact that light reduction during meiosis phenomenon altered outcrossing fertilisation. The anthesis date appears not to be modified by neither light nor nitrogen availabilities (Chauvel et al., 2005).
4. Propagation abilities
4.1. Pollen
20More than 70% of A. myosuroides pollen dispersal was found to occur within 1 m, although fecundation can occur at a distance of at least 60 m from the pollen donor plant in the absence of physical barriers to pollen propagation (Chauvel, 1991). So, one field could include several subpopulations. Considering the mean heterozygosity calculated for seven loci, black-grass populations fit with panmixia equilibrium: it seems that genes are mostly exchanged at random within a field (Chauvel et al., 1994).
21Menchari et al. (2007) observed a total expected heterozygosity of 0.21 and a mean FST value of 0.023. The low differentiation among populations and the absence of a significant relationship between genetic and geographical distances suggest that there may be extensive gene flow connecting black-grass populations. However, the self-dispersal capacity of A. myosuroides seeds and pollen were shown to be limited.
22Studies of genetic variation of weeds (and invaders generally) after their introduction (Warwick et al.,19875 cited in Chauvel et al., 1994) in a new geographical area have shown that an out crossing mating system reduces the effect of genetic drift in comparison with a strong inbreeding mating system (Warwick, 1991a6 and Bosbach et al., 19817 cited in Chauvel et al., 1994).
23Richter et al. (2004) presented a model to describe the spatial spread of pollen over a closed canopy with a Neumann boundary condition at the surface of the canopy. The wind profile above the canopy causes long-range transport in the upper layers, whereas particles near the boundary layer are transported only a short range because of gravitational settling and low velocities within the boundary layer. Then they linked these data to model the spatial spread of pesticide resistant pest (or weed) populations. They used the modeling framework above to study the spread of pesticide resistance using an artificial simulation experiment.
24The hypothetical study area consisted of three plots “B”, “C”, and “D” (Figure 2). A pesticide (or herbicide) is applied to plots “B” and “C”. Plot “D” remained untreated. Starting from an infested spot “A” on plot “B” a resistant biotype begins to spread in the study area. The resultant spatial distributions of resistant (AA and aA) and sensitive (aa) biotypes are shown in figure 3 for two time steps. The resistant biotypes invade the treated plots, whereas the sensitive biotype retreats to the untreated plot, which may serve as a refuge (Richter et al., 2004).
25Amalgamation of all the processes (dispersal, growth and genetics) into the same mathematical structure (i.e. partial differential equations) allows a broad spectrum of applications that encompass developments of anti-resistance strategies, as well as risk assessment of spread of transgenic properties. However, for a detailed mapping of the life cycle of plants, it is more appropriate to use time discrete models that are embedded into cellular automaton models (Seppelt, 2003, chapter 38 cited in Richter et al., 2004). A major drawback of cellular automaton models is that the range of dispersal in one time step is limited by the order of the Moore radius. Cellular automata models are therefore not capable of modeling pollen dispersal on a large (landscape) scale. In order to achieve both a realistic description of plant (or insect) development in terms of a time discrete scheme and a realistic simulation of long-range dispersal, both systems have to be integrated into a hybrid model (Richter et al., 2004).
26Busi et al. (2008) conducted a landscape level field experiment to try to assess the pollen-mediated gene flow at long distance. They used Lolium rigidum herbicide-susceptible individuals placed at increasing distance of a large field infested with herbicide-resistant L. rigidum. Herbicide resistance was used as a marker to quantify the distance and the rate of pollen-mediated gene flow. Resistance genes were found at the maximum tested distance of 3,000 m. This was the first reported case of long-distance gene flow for L. rigidum, and these data are fairly outstanding. We now have to know if this distance can be transposed to A. myosuroides.
4.2. Seeds
27Seed production differences were especially linked to the tiller number. The effect of nitrogen fertilizer did not always increase weed seed production. Furthermore, the germination rate was reduced for seeds produced under high level of nitrogen (Colbach et al., 2003), while it is increased with seed weight. Caryopsis weight decreases with higher density (D2) and the quality of the seed produced in competition was partly altered (Chauvel et al., 2005).
28Chauvel et al. (2005) observed that black-grass behaves as typical allogamous species showing high seed viability in high plant density conditions. They explained this by the number of flowering plants in each greenhouse that produced such a pollen quantity that out crossing fertilization values were strongly higher in comparison with field data.
29The dispersal unit of A. myosuroides is the whole spikelet which is single-flowered. The mean dispersal distance of black-grass seeds is 51 cm (Colbach et al., 2001). Wind speed (significant at 0.0001, whatever the analyzed parameter) increases both the number of dispersed seed and the dispersal distance with the highest seed density. The region (Figure 4) also influences these two parameters, and it appears that both the number of collected seeds and the distance at which seeds were dispersed were higher in the downwind region (II). Region strongly interacts with another significant input variable, i.e. the number of ears per plant. This variable increases the number of dispersed seeds, independently of the regions. However, the number of ears per plant effect on dispersal distance depended on the regions: it was nil in the upwind region (which was the region with little dispersal) and highly significant in the downwind region (which was the region with most dispersal); in the latter case, dispersal distances were lowest for plants with large number of ears (Colbach et al., 2001).

30Plant height only influenced the distance at which the largest number of seeds was found; the higher the plants, the further away from the plant the seeds fell. Lighter seeds are carried farther because gravity interacts less with their horizontal movement (Colbach et al., 2001). As expected, the density of collected seeds formed a leptokurtic distribution with a peak near the seed-producing plant and a monotonic decrease with distance (Schupp et al., 19959 cited in Colbach et al., 2001). Of course, dispersal by harvest tools carries seeds over larger distances than “natural” seed dispersal (Colbach et al., 2001).
31Seeds of black-grass have been efficiently eliminated from cereal seeds for at least 70 years in France, so that dispersal via cereal seed lots used for sowing is very unlikely at present. However, seeds may occasionally be transported by farm machinery, especially combine-harvesters and tractors, over a few kilometers, as shown for other grass weed species (Légère et al., 2000). Since contemporary gene flow among agricultural fields seems rather restricted, the low genetic differentiation at AFLP markers may result from important gene flow that occurred in the past. The historical distribution and demography of A. myosuroides are not well known (Menchari et al., 2007). In France, its presence has regularly been recorded in botanical reports from the end of the 19th century to the mid-20th century, although it was not considered as a harmful weed at that time. At the end of the 19th century, black-grass was sown in meadows as a minor species bulked with other grasses, and appeared in commercial seed lots (Heuze, 191110 cited in Menchari et al., 2007). This could have resulted in long-distance transportation of seeds.
32After the shedding, seeds can be transported at a further distance by small animals such as mice and voles. This dispersion mechanism called epizoochory, whose widely studied cases are those mediated by herbivores, concerns adhesion to animal fur (Sorensen, 1986; Davidson, 199311 cited in Benvenuti, 2007). The plant height contributes to the effectiveness of epizoochory as the degree of dispersal is greatest when the height of the plant is approximately similar to that of the animal with which the plant most frequently comes into contact.
33Dactylis glomerata L. and Bromus erectus Huds. are likewise highly successful in achieving lasting adhesion to ovine fur and can be transported for prolonged distances. Other examples of epizoochory are found in Knautia arvensis (L.) Coulter (Dipsaceae), some species of the genus Medicago (Fabaceae), and almost all species of the genus Setaria (Poaceae). In the latter case, the modifications of the spikelet allow adhesion. The botanical family of Graminaceae has numerous species that undergo epizoochorous dispersal, even though they have no particular specialization. The elongated and frequently pointed shape of their seeds, as in the case of Bromus sterilis L., Lolium multiflorum Lam., and Alopecurus myosuroides Huds., facilitates adhesion (Benvenuti, 2007).
34In the case of black-grass growing in a crop, especially wheat, shedding occurs when the vegetation is very dense and few animals go through it, enough to be in contact with the heads. Nevertheless, when seeds fall to the ground, small mammals surrounding on the soil, or birds like pheasant or partridge, nesting under the cover, might get these on their feathers or fur and then make them travel away.
5. Models
5.1. Life cycle
35S.R. Moss developed in 1990 a life cycle model of A. myosuroides presented in figure 5 (Cavan et al., 2000), which relates the number of inflorescences produced per square meter, h, to the number of plants per square meter surviving herbicide treatment, p, by the density-dependent relation:
36h = 3,88p / (1,0 + 0,0018 p).
37The model assumes that 55 viable seeds are produced on each head and shed on to the soil surface, where 55% of seeds are lost by predation, decay and germination before cultivation. The soil seed bank is divided into two levels: the top 5 cm “surface seed bank” from which seedlings can emerge and the lower “deep seed bank” from which they cannot. Seed predation and decay remove 70% of the seed bank annually at both levels. The plough (mould board to 25 cm deep) moves 95% of seeds from the surface seed bank to the deep seed bank and 35% from the deep to the surface seed bank. Tine cultivation to 10 cm deep moves 20% of seeds from the surface seed bank to the deep seed bank but does not move any of the deep seed bank upwards. Annually 15% of newly shed seeds and 30% of seeds that are at least 1 year old produce seedlings which emerge (from the shallow seed bank). A proportion of susceptible seedlings (set at either 80%, 90% or 95% “herbicide kill rate”) are killed by herbicide before maturing to produce heads. The initial seed bank contains 100 newly shed seeds per m², distributed evenly to a depth of 25 cm. Single-gene resistance was incorporated into the model with a mutation rate of 10-6 per generation, conferring total resistance to ACCase inhibitors herbicides in both homozygous and heterozygous states but not affecting the kill rate of other herbicide groups. Randomly, spatially homogeneous pollination was assumed. Infestations of A. myosuroides do not start to impact significantly on cereal yields until they exceed 10 plants·m-² (Ingle et al., 199712 cited in Cavan et al., 2000) but populations can increase extremely rapidly unless checked by herbicide. Consequently, this population level (10 plants·m-²) was used as a threshold to define field resistance (Cavan et al., 2000).
5.2. ALOMYSYS
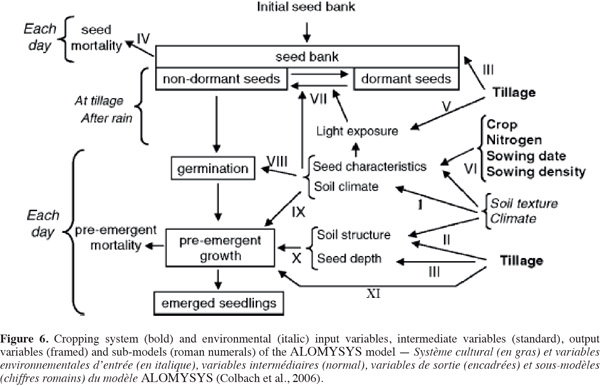
38ALOMYSYS is another model developed by Colbach et al. (2006) that can also be really useful in the understanding and prediction of black-grass dispersal within a field (Figure 6). This model gathers and integrates information from 11 sub-models (roman numerals in figure 6), predicting (a) soil environment (climate, structure) resulting from the cropping system and external climate, (b) vertical soil seed distribution after tillage, and (c) seed survival, dormancy, germination and pre-emergent growth depending on soil environment, seed depth, characteristics and past history. Consequently, the model can be used to simulate the effects of cropping systems on black-grass emergence.
6. Conclusion
39Since early 1980’s and even before, research on black-grass ecology has really went up, especially because of the development of herbicide resistance. The first case was reported in 1983. The aim of this research was to get as much information as possible on this plant to try to get rapidly rid of it in our crops. Unfortunately, for farmers, thirty years after, black-grass still remains a major concern. During the last fifty years, we could assist to a wide spread of resistant populations, first in the UK, and now in the whole Northern part of Europe. Although herbicide resistant genes can explain in great part its recent spread, combined effects of different practices due to intensive agriculture seem also to be involved. To try to avoid or limit a wider extension of theses resistant biotypes it is essential to develop and implement integrated cropping systems.
40As these new systems introduce changes in cultural practices, such as modifications in crop sowing dates or the introduction of new crops in the rotation, knowledge about the effects of these practices on weed demography are important to improve weed management and prevent weed spread. Demographic models for A. myosuroides, such as ALOMYSYS, take into account the number of days to flowering to predict the possibility of seed production in new cropping systems. For example, reduced row spacing or selecting for more competitive wheat varieties could help to reduce weed seed production and could manage weed infestation. Every topic presented in this paper can be aimed at, in a way to limit as much as possible the development and the extension of this weed, even before using herbicides.
41Unfortunately, when some individuals manage to slip through the net, and get to flowering stage, black-grass, especially resistant ones, are able to scatter in the environment, either by pollen flow, or by the seed. Wind direction and speed are most important for daily seed dispersal. The total number of seeds increases with wind speed as more seeds are detached from the ear; they are carried farther away from the plant, in the windward direction, by the force of the wind. Nevertheless, most seeds are dispersed no further than the immediate neighborhood of their mother plant (50-60 cm) (Colbach et al., 2001). However, Busi showed in 2008 that Lolium rigidum pollen can cover up to 3 km. Some research has to be conducted to verify if these data can be transposed to A. myosuroides, but it is clear that such a distance is really important for resistance gene dispersion, even at a landscape level.
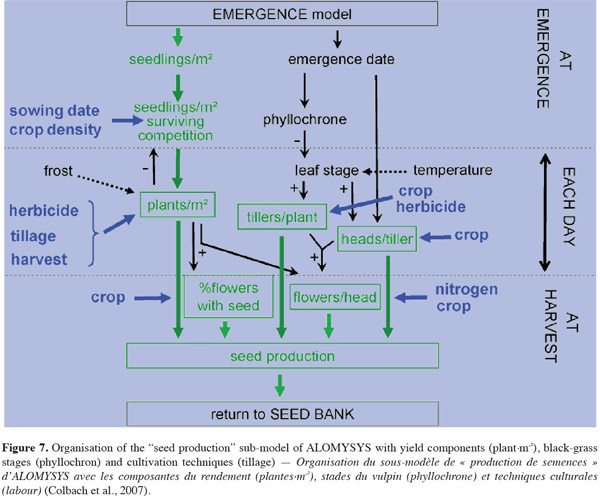
42To sum up the article, figure 7 presents the “seed production” sub-models of ALOMYSYS, taking into account the whole of the major steps depicted through this article.
Bibliographie
Andersson L.L. & Åkerblom Espeby L., 2009. Variation in seed dormancy and light sensitivity in Alopecurus myosuroides and Apera spica-venti. Weed Res., 49, 261-270.
Benvenuti S., 2007. Weed seed movement and dispersal strategies in the agricultural environment. Weed Biol. Manage., 7, 141-157.
Busi R., Yu Q. & Barrett-Lennard R., 2008. Long distance pollen-mediated flow of herbicide resistance genes in Lolium rigidum. Theor. Appl. Genet., 117, 1281-1290.
Cavan G., Cussans J. & Moss S.R., 2000. Modelling different cultivation and herbicide strategies for their effect on herbicide resistance in Alopecurus myosuroides. Weed Res., 40, 561-568.
Chauvel B., 1991. Polymorphisme et sélection pour la résistance aux urées substituées chez Alopecurus myosuroides Huds. Thèse de doctorat : Université Paris XI, Orsay (France).
Chauvel B. & Gasquez J., 1994. Relationships between genetic polymorphism and herbicide resistance within Alopecurus myosuroides Huds. Heredity, 72, 336-344.
Chauvel B., 1996. Variabilité de la production de semences chez le vulpin (Alopecurus myosuroides Hudson) en fonction de la culture. In : 10e Colloque international sur la Biologie des mauvaises herbes, 11-13 septembre 1996, Dijon, France, 43-49. Alfortville, France : AFPP.
Chauvel B., Munier-Jolain N., Letouzé A. & Grandgirard D., 2000. Developmental patterns of leaves and tillers in a black-grass population (Alopecurus myosuroides Huds.). Agronomie, 20, 247-257.
Chauvel B., Munier-Jolain N.M., Grandgirard D. & Gueritaine G., 2002. Effect of vernalization on the development and growth of Alopecurus myosuroides. Weed Res., 42, 166-175.
Chauvel B., Guillemin J.P. & Letouzé A., 2005. Effect of intra-specific competition on development and growth of Alopecurus myosuroides Hudson, Europ. J. Agronomy, 22, 301-308.
Colbach N. & Sache I., 2001. Black-grass (Alopecurus myosuroides Huds.) seed dispersal from a single plant and its consequences on weed infestation. Ecol. Model., 139, 201-219.
Colbach N., Chauvel B., Dürr C. & Richard G., 2002. Effect of environmental conditions on Alopecurus myosuroides germination. I. Effect of temperature and light. Weed Res., 42, 210-221.
Colbach N. & Dürr C., 2003. Effects of seed production and storage conditions on black-grass (Alopecurus myosuroides) germination and shoot elongation. Weed Sci., 51, 708-717.
Colbach N. et al., 2006. AlomySys: modelling black-grass (Alopecurus myosuroides Huds.) germination and emergence, in interaction with seed characteristics, tillage and soil climate. I. Construction. Eur. J. Agron., 24, 95-112.
Colbach N., Chauvel B., Gauvrit C. & Munier-Jolain N.M., 2007. Construction and evaluation of ALOMYSYS modelling the effects of cropping systems on the black-grass life-cycle: from seedling to seed production. Ecol. Model., 201, 283-300.
Cussans G.W., Randonius S., Brain P. & Cumberworth S., 1996. Effects of depth of seed burial and soil aggregate size on seedling emergence of Alopecurus myosuroides, Galium aparine, Stellaria media and wheat. Weed Res., 36, 133-141.
Dalbiès-Dulout A. & Doré T., 2001. Management of inflorescence and viable seed production of blackgrass (Alopecurus myosuroides) on set-aside in France. Crop Prot., 20, 221-227.
Délye C. et al., 2007. Status of black grass (Alopecurus myosuroides) resistance to acetyl-coenzyme A carboxylase inhibitors in France. Weed Res., 47, 95-105.
Froud-Williams R.J., Drennan D.S.H. & Chancellor R.J., 1984a. The influence of burial and dry-storage upon cyclic changes in dormancy, germination and response to light in seeds of various arable weeds. New Phytol., 96, 473-481.
Froud-Williams R.J., Chancellor R.J. & Drennan D.S.H., 1984b. The effects of seed burial and soil disturbance on emergence and survival of arable weeds in relation to minimal cultivation. J. Appl. Ecol., 21, 629-641.
Kirby E.J.M., Appleyard M. & Fellowes G., 1985. Effect of sowing date and variety on main shoot leaf emergence and number of leaves in barley. Agronomie, 5(2), 117-126.
Légère A., Beckie H.J., Stevenson F.C. & Thomas A.G., 2000. Survey of management practices affecting the occurrence of wild oat (Avena fatua) resistance to acetyl-CoA carboxylase inhibitors. Weed Technol., 14, 366-376.
Letouzé A. & Gasquez J., 2001. Inheritance of fenoxaprop-P-ethyl resistance in a black-grass (Alopecurus myosuroides Huds.) population. Theor. Appl. Genet., 103, 288-296.
Marshall R. & Moss S.R., 2008. Characterisation and molecular basis of ALS inhibitor resistance in the grass weed Alopecurus myosuroides. Weed Res., 48, 439-447.
Masle J., Doussinault G., Farquhar G.D. & Sun B., 1989. Foliar stage in wheat correlates better to photothermal time than to thermal time. Plant Cell Environ., 12, 235-247.
Menchari Y., Délye C. & Le Corre V., 2007. Genetic variation and population structure in black-grass (Alopecurus myosuroides Huds.), a successful, herbicide-resistant, annual grass weed of winter cereal fields. Mol. Ecol., 16, 3161-3172.
Moss S.R., 1985. The survival of Alopecurus myosuroides Huds. seeds in soil. Weed Res., 25, 201-211.
Ouborg N.J., Piquot Y. & Van Groenendael J.M., 1999. Population genetics, molecular markers and the study of dispersal in plants. J. Ecol., 87, 551-568.
Richter O. & Seppelt R., 2004. Flow of genetic information through agricultural ecosystems: a generic modelling framework with application to pesticide-resistance weeds and genetically modified crops. Ecol. Model., 174, 55-66.
Roché Jr B.F., Roché C.T. & Chapman R.C., 1994. Impacts of grassland habitat on yellow starthistle (Centaurea solstitialis L.) invasion. Northwest Sci., 68, 86-96.
Sorensen A.E., 1986. Seed dispersal by adhesion. Ann. Rev. Ecol. Syst., 17, 443-446.
Swain A.J., Hughes Z.S., Cook S.K. & Moss S.R., 2006. Quantifying the dormancy of Alopecurus myosuroides seeds produced by plants exposed to different soil moisture and temperature regimes. Weed Res., 46, 470-479.
Notes
Para citar este artículo
Acerca de: Pierre-Yves Maréchal
Univ. Liege - Gembloux Agro-Bio Tech. Unit of Plant Science of the moderate Regions. Passage des Déportés, 2. B-5030 Gembloux (Belgium). E-mail: pymarechal@doct.ulg.ac.be
Acerca de: François Henriet
Centre wallon de Recherches agronomiques. Département Sciences du Vivant (D1). Unité Protection des Plantes et Écotoxicologie (U4). Rue du Bordia, 11. B-5030 Gembloux (Belgium).
Acerca de: Françoise Vancutsem
Univ. Liege - Gembloux Agro-Bio Tech. Unit of Plant Science of the moderate Regions. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Acerca de: Bernard Bodson
Univ. Liege - Gembloux Agro-Bio Tech. Unit of Plant Science of the moderate Regions. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
