
Évaluation de l'impact des cours d'eau sur l'eutrophisation dans la bande côtière : nécessité de raisonner sur les concentrations et sur les flux
Agrocampus-Ouest. 65 rue de Saint Brieuc. F-35042 Rennes Cedex (France). E-mail : pierre.aurousseau@cseb-bretagne.fr
Résumé
Le raisonnement de la qualité des eaux en concentration a été privilégié jusqu’ici principalement à cause des normes de potabilité qui sont définies en concentration (par exemple, 50 mg·l-1 pour le nitrate) et bien que cette norme s’appuie en fait sur une absorption, par l’eau de boisson, plafonnée à 20 mg d’azote par jour. Le double raisonnement en flux et concentration est encore plus incontournable quand on s’intéresse aux impacts environnementaux car l’intensité de ces impacts est liée en grande part à la quantité de nutriments apportés, c’est-à-dire au flux de sels nutritifs parvenant dans un compartiment de l’environnement. Cette liaison entre flux de nutriments et impacts environnementaux est particulièrement bien établie pour les milieux marins côtiers qui sont exposés aux phénomènes de marées vertes, blooms de diatomées ou de Phaeocystis. À l’échelle d’un bassin versant, l’évaluation quantitative des flux n’est pas sans poser quelques problèmes de métrologie. Le raisonnement en flux offre l’avantage de pouvoir mieux comparer l’impact (à l’échelle d’une bande côtière) de plusieurs bassins versants. À l’échelle du bassin versant, pour réduire les flux sortants, il importe d’intervenir sur les flux entrants en réduisant l’excédent des bilans N et P. Cette façon de raisonner est illustrée avec l’exemple des phénomènes d’eutrophisation des eaux marines côtières en Manche-Est et à l’entrée de la Mer du Nord.
Abstract
Assessing the impact of streams on eutrophication in the coastal strip: the need to consider concentrations and loads. The approach of using measurements of water quality concentrations has been favored to date because of the current standards for drinking water, which are defined in terms of concentration. For example, the standard for the concentration of nitrate is 50 mg·l-1, although this standard, in fact, derives from a limited quantity of nitrate absorbed by drinking water with a concentration of 20 mg nitrogen per day. The dual assessment of flow and concentration is essential when studying environmental impacts because the intensity of these impacts is mainly linked to the quantity of nutrients reaching a compartment of the environment. This link between nutrient load and environmental impact is particularly established in coastal marine environments exposed to the phenomena of green tides, diatoms or Phaeocystis blooms. At the watershed scale, flow control presents some metrological problems but assessment based on flow has the advantage of providing a better comparison of the overall impact (at the coastal band scale) of several watersheds. At the catchment scale, in order to reduce the outflow, it is important to intervene at the level of the inflow by reducing any excesses in the N and P balance. This approach is illustrated by the example of the eutrophication of coastal marine water in the Eastern English Channel and at the mouth of the North Sea.
1. Introduction
1La norme de 50 mg NO3-·l-1 d’eau considérée comme norme de potabilité résulte en réalité d’une norme de flux maximum d’ingestion par l’homme située à 20 mg N par jour dans deux litres de boisson, soit 10 mg N·l-1 ou 44 mg NO3-·l-1 (arrondi in fine à 50 mg NO3-·l-1). Ce même raisonnement en flux devra être adopté pour évaluer les conséquences des apports de nitrate en termes d’eutrophisation des eaux.
2. Relation des impacts environnementaux avec les flux
2Précocement, les scientifiques ont attiré l'attention des élus et de l'administration sur le fait qu'il ne fallait pas être obnubilé par la norme des 50 mg·l-1 de nitrate dans l'eau et sur le raisonnement de la qualité des eaux en termes de concentration. Les travaux scientifiques ont insisté dès les années 1980 sur le fait, par exemple, que l’on pouvait en un lieu établir une relation entre le flux en nitrate pendant les mois de photosynthèse et le développement des algues vertes dans les sites à marées vertes en Bretagne (Chapelle et al., 1994). D'autres travaux ont aussi rapidement démontré que les objectifs à atteindre pour limiter de manière suffisamment forte ce phénomène de marées vertes allaient être plus contraignants que la norme des 50 mg·l-1 de nitrate dans l'eau (Ménesguen et al., 1995 ; Perrot et al., 2007). On a compris que le respect d'un certain nombre de règles agronomiques suffirait pour descendre en dessous de cette norme des 50 mg·l-1 sans remettre obligatoirement en cause les systèmes agricoles pratiqués sur les bassins versants. Par contre, descendre en dessous de 15 mg·l-1, voire moins (10, voire 5 mg·l-1) conduirait à remettre profondément en cause les systèmes agricoles tels que nous les connaissons actuellement.
3Il est évidemment logique que l'eutrophisation dans un site donné soit liée à un flux de sels nutritifs arrivant dans ce site. Il y a bien en effet une relation entre la quantité de matière végétale produite conséquemment à l'eutrophisation et la quantité de sels nutritifs arrivant dans le site. On peut illustrer ce mécanisme par plusieurs exemples :
4– celui des marées vertes (déjà cité) ;
5– celui d'un bloom de diatomées dans un site sensible à l'eutrophisation par phytoplancton comme l'estuaire de la Vilaine (Sud Bretagne) ou comme l'estuaire de la Seine (Guillaud et al., 2000 ; Cugier et al., 2005a ; Cugier et al., 2005b ; Billen et al., 2009 ; Thieu et al., 2010) ;
6– celui d'un bloom de Phaeocystis (Figure 1) dans un site sensible comme la Manche-Est ou l'entrée de la Mer du Nord (Lancelot et al., 2007 ; Thieu et al., 2009 ; Lancelot et al., 2011).
7Il en est de même si on s'intéresse non plus à un phénomène d'eutrophisation en eaux marines côtières, mais à un phénomène d'eutrophisation en eaux douces comme, par exemple, aux blooms de cyanobactéries dans des lacs ou des retenues eutrophes.

8En conclusion partielle, nous dirons que l'analyse des impacts environnementaux nécessite le plus souvent d'aborder le diagnostic de la qualité des eaux en flux et non plus seulement en concentration et que souvent les objectifs de réduction des impacts environnementaux imposeront des objectifs de qualité des eaux plus contraignants que les objectifs pour les eaux brutes, les eaux distribuées ou les eaux de consommation.
3. L’évaluation des flux
9L'évaluation des flux pose de multiples problèmes, parmi lesquels on citera :
10– la nécessité de disposer de fréquences suffisantes de mesures de concentration (Birgand et al., 2009 ; Moatar et al., 2009) :
11– la mise en relation des stations de suivi qualité (où la concentration est mesurée) avec des stations de suivi des débits (appelées aussi station débit ou station limnimétrique).
12En Bretagne, les stations de suivi qualité ne sont pas obligatoirement situées au même endroit que les stations limnimétriques, car elles appartiennent le plus souvent à des réseaux de suivi différents qui ont été établis par des organismes différents et obéissant à des logiques différentes. Quand pour une rivière donnée, un fleuve donné, un bassin versant donné, la station de suivi qualité est distincte de la station de suivi limnimétrique, il faut faire appel à des techniques d'extrapolation (le plus souvent, on extrapole les données de débit au niveau de la station de suivi qualité où l'on décide de calculer les flux). Si l'on souhaite disposer d'une évaluation des flux sortants en un point exutoire du bassin versant ou du sous-bassin versant (CSEB, 2005b), on réalisera une seconde extrapolation en ce point (CSEB, 2005c). Ce point exutoire est par exemple le point de confluence d'un affluent avec une rivière ou un fleuve plus important ou encore un exutoire « à la mer ».
13Les flux sont la plupart du temps très dépendants des débits, car la variabilité dans le temps des débits est le plus souvent beaucoup plus grande que la variabilité dans le temps des concentrations (par exemple, on peut rencontrer couramment des situations où la variabilité des débits est d'un facteur 10, alors que la variabilité des concentrations est d'un facteur 3). Il existe plusieurs techniques pour s'abstraire – autant que faire se peut – de cette variabilité temporelle :
14– calculer des moyennes pluriannuelles sur des périodes suffisamment longues couvrant le plus possible la variabilité annuelle ;
15– faire des corrections du flux en fonction des débits ou de l'hydraulicité de l'année (CSEB, 2005a).
16Le flux sortant d'un bassin versant est également très dépendant de la superficie du bassin versant, c'est la raison pour laquelle on calcule souvent simultanément le flux sortant et le flux dit « spécifique » qui est le flux sortant ramené par unité de surface du bassin versant. Le flux sortant est habituellement exprimé en tonnes (de N par exemple) et le flux spécifique en kg·km-2 ou kg·ha-1. Les hydrologues expriment habituellement le flux spécifique d’azote en kg·km-2 et les agronomes l'expriment le plus souvent en kg·ha-1. Par exemple, dans la synthèse réalisée par Howarth et al. (1996) sur l'Océan atlantique nord élargi, le flux spécifique de l'Amazone est de 505 kg N·km-2 (ou 5,05 kg·ha-1).
17Le flux spécifique exprimé en kg·ha-1 présente l'intérêt de pouvoir être comparé à l'excédent du bilan azoté sur le bassin versant (appelé aussi en conséquence flux entrant dans le bassin versant) et à d'autres indicateurs agronomiques intermédiaires (azote potentiellement lessivable, etc.).
18Le calcul du flux sortant et du flux spécifique constitue un outil pertinent pour comparer des bassins versants entre eux (Figure 2). À titre d’exemple, le flux spécifique moyen des principales rivières et fleuves de Bretagne est de l’ordre de 25 kg N·ha-1. Ce flux moyen cache cependant une variabilité allant de 10 à 97 kg N·ha-1. Les bassins versants bretons sont des bassins côtiers de petite superficie avec des temps courts de séjour de l'eau dans le réseau hydrographique ; ils sont occupés par une agriculture intensive qui conduit à des flux spécifiques plus élevés que les bassins de la Manche-Est et de l'entrée de la Mer du Nord (Seine : 19 kg N·ha-1, Somme : 14 kg N·ha-1, Escaut : 23 kg N·ha-1).
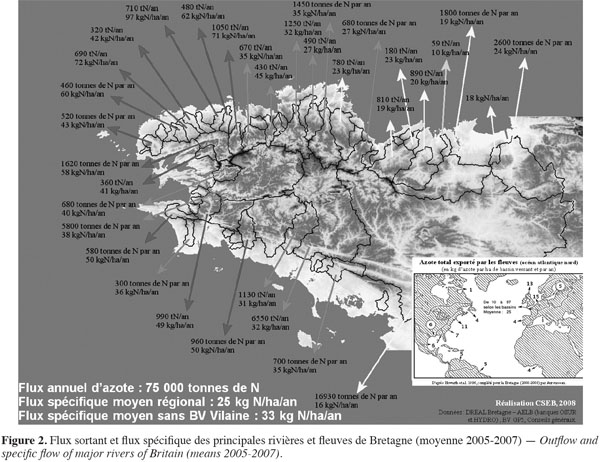
19Dans une phase de reconquête de l'environnement et de la qualité des eaux, on est amené à se fixer des objectifs de réduction du flux sortant et du flux spécifique qu'il s'agit ensuite de convertir en objectifs de réduction du flux entrant dans le bassin versant afin de réduire ces excédents sortant.
4. Le double raisonnement en concentration et en flux
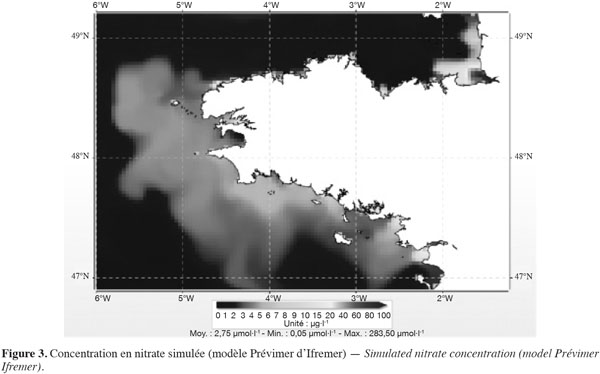
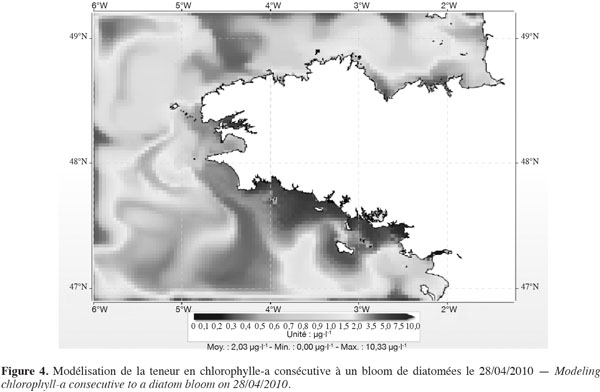
20Le phénomène des marées vertes constitue un excellent exemple pour comprendre l'intérêt de ce double raisonnement. D'un côté, en un lieu donné, il y a une relation entre la biomasse produite et la quantité de sels nutritifs apportés au site pendant les mois de photosynthèse. D'un autre, les ulves (qu'il s'agisse de Ulva lactuca comme en Bretagne ou de Ulva prolifera anciennement dénommée Enteromorpha prolifera comme en Chine) sont des végétaux assez rudimentaires qui ne disposent pas de l'équipement enzymatique pour s'alimenter en azote à partir d'une eau peu chargée en azote inorganique. Même si le panache des fleuves peut avoir des concentrations de 10 à plusieurs dizaines de µmoles NO3-·l-1, soit de 0,6 à 6 mg·l-1 NO3- (Figure 3), les algues vertes ne savent pas s’alimenter en azote à partir de telles eaux. Avec des eaux oligotrophes (de 1 à 10 µmoles, soit de 0,06 à 0,6 mg·l-1 NO3- (Figure 4), c’est aussi encore plus impossible, même si à cause des volumes considérables d’eaux marines à ces concentrations, la quantité d’azote disponible dans ces eaux est en fait très importante. En langage courant, on dit que l'algue « ne voit pas » cet azote inorganique. Ces algues ne pourront s'alimenter en azote que si elles baignent dans une eau fortement chargée en azote inorganique (entre 100 et 1 000 µmoles·l-1). Cet intervalle de concentration ne peut s'observer qu'à proximité immédiate de l'exutoire des fleuves. Pour simplifier et en prenant l'exemple de la Bretagne, on admettra que la plupart des fleuves et rivières ont à l'exutoire des concentrations de l'ordre de 500 à 1 000 µmoles·l-1 (soit environ 30 à 60 mg·l-1 de nitrate). Descendre à 100 µmoles·l-1 implique alors un taux de dilution de 5 à 10. En deçà de 100 µmoles·l-1, l'azote inorganique constituera de manière prépondérante la cause des blooms phytoplanctoniques.
21Cet exemple que nous venons de détailler constitue une excellente démonstration de l'intérêt du double raisonnement de la qualité des eaux en flux et en concentration.
5. Application de ce double raisonnement en concentration et en flux aux cas des bassins de la Manche-Est et de l'entrée de la Mer du Nord
22Billen et al. (2009) ont comparé d'un côté les flux sortants de la Seine, de la Somme et de l'Escaut (respectivement 19,50, 14,30 et 23,10 kg N·ha-1) pour l'année 2000, considérée comme une année hydrologique moyenne. D'un autre côté, ils ont comparé la part impactée par l’homme de l’excédent du bilan azoté (respectivement 59,65, 60,60 et 112,15 kg N·ha-1). Howarth et al. (1996), Alexander et al. (2002), Boyer et al. (2002) et Howarth et al. (2006) ont montré que le NANI est un bon indicateur du flux sortant des bassins versants (le NANI est la somme des apports atmosphériques, des apports de fertilisants minéraux, de la fixation atmosphérique et du solde du bilan dit « food and feed ». Dans le langage francophone, le NANI serait appelé un bilan entrée-sortie). Dans ces trois cas, on est face à des bilans azotés fortement excédentaires et il y a un défaut de flux élevé (différence entre l'excédent du bilan et le flux sortant). Ce type de situation avec des défauts de flux élevés peut être mis en relation avec le fait que ces bassins versants sont d'assez grande taille (en comparaison avec des petits bassins versants côtiers comme ceux de Bretagne où le temps de séjour de l'eau dans le réseau hydrographique est plus court), avec une partie aval du réseau hydrographique où la circulation est lente et parfois canalisée, favorable au développement d'une eutrophisation interne des eaux douces susceptible de consommer une partie de l'azote transférée.
23Dans ces trois cas, le flux sortant est inférieur à très inférieur aux flux sortants de bassins versants intensifs de Bretagne (moyenne de 25 kg N·ha-1, maximum de 97 kg N·ha-1).
24Sous l'effet de ces flux nutritifs, la Manche et l'entrée de la Mer du Nord sont affectées par des phénomènes d'eutrophisation phytoplanctonique. Une des caractéristiques de la Manche-Est et de l'entrée de la Mer du Nord est l'existence de blooms de Phaeocystis.
25Les blooms de Phaeocystis sont interprétés comme étant le résultat d'un déséquilibre dans la flore phytoplanctonique consécutif à une dystrophie (déséquilibre dans les rapports N:P:Si). En situation pristine, ces rapports auraient oscillé autour de 40:1:400 (400 fois moins de P que de Si et 10 fois moins de N que de Si). Ceci signifie qu'en conditions pristines, ces trois éléments se présentaient dans l'ordre suivant de concentration (Si > N > P). Aujourd'hui, les apports terrigènes sont plus riches à beaucoup plus riches en N et en P par rapport au Si (N > P > Si). Comme les sédiments marins ont une faible contribution à la nutrition des blooms phytoplanctoniques, l'enrichissement des apports telluriques en N et en P joue un rôle important sur les blooms. Le déséquilibre avec le Si conduit à des efflorescences de certaines espèces phytoplanctoniques qui seraient dominées en conditions naturelles par les diatomées dont les besoins en Si sont élevés.
26La réduction des blooms de Phaeocystis en Manche-Est et à l'entrée de la Mer du Nord devrait conduire les politiques de reconquête de la qualité de l'eau dans les bassins versants qui alimentent ces fleuves (Seine, Somme et Escault) à être non seulement fondées sur des objectifs en concentration, mais aussi sur des objectifs de réduction des flux. Se fixer des objectifs de réduction des flux sortants à l'exutoire des bassins versants conduit à se fixer des objectifs de réduction des flux entrants dans les bassins. Appliqué au cas des composés azotés, cela conduit à se fixer des objectifs de réduction de l'excédent du bilan azoté dans les bassins.
6. Conclusion
27Le caractère spectaculaire des eutrophisations à algues vertes et leurs conséquences sanitaires ont conduit à faire admettre un concept prôné par les scientifiques depuis longtemps déjà : celui du double raisonnement de la qualité des eaux en concentration et en flux. Ce double raisonnement constitue un des fondements des politiques de reconquête de la qualité des eaux. Ce double raisonnement a non seulement un intérêt dans le cas de la lutte contre l'eutrophisation à macro-algues (marées vertes), mais aussi dans ceux de la lutte contre l'eutrophisation à phytoplancton. Ce double raisonnement s'applique donc aussi bien à des cas comme ceux de la Vilaine (Sud Bretagne) dont l'estuaire est un site sensible aux blooms de diatomées, à l'hypoxie et à l'anoxie, que dans les cas de la Manche-Est et de l'entrée de la Mer du Nord qui sont des sites sensibles aux blooms de Phaeocystis.
Bibliographie
Alexander R.B., Johnes P.J., Boyer E.W. & Smith R.A., 2002. A comparison of models for estimating the riverine export of nitrogen from large watersheds. Biogeochemistry, 57/58, 295-339.
Billen G., Thieu V., Garnier J. & Silvestre M., 2009. Modelling the N cascade in regional watersheds: the case study of the Seine, Somme and Scheldt rivers. Agric. Ecosyst. Environ., 133, 234-246.
Birgand F. et al., 2009. Une approche quantitative du rôle de la fréquence d'échantillonnage sur les incertitudes associées aux calculs de flux et des concentrations moyennes en nitrate en Bretagne. Ingénieries, 59-60, 23-37.
Boyer E.W., Goodale C.L., Jaworski N.A. & Howarth R.W., 2002. Anthropogenic nitrogen sources and relationships to riverine nitrogen export in the Northeastern USA. Biogeochemistry, 57/58, 137-169.
Chapelle A., Lazure P. & Ménesguen A., 1994. Modelling eutrophication events in a coastal ecosystem. Sensitivity analysis. Estuarine Coastal Shelf Sci., 39, 529-548.
CSEB (Conseil scientifique de l'environnement de Bretagne), 2005a. Fiche E-5. Hydraulicité, correction de l’effet de la pluviosité : flux pondéré par l’hydraulicité, http://www.cseb-bretagne.fr/downloads/CSEB_Recueil%20Fiches_Tomes1+2.pdf, (16.01.13).
CSEB (Conseil scientifique de l'environnement de Bretagne), 2005b. Fiche E-7. Bassin versant, sous-bassin versant, tronçon et calcul de flux, http://www.cseb-bretagne.fr/downloads/CSEB_Recueil%20Fiches_Tomes1+2.pdf, (16.01.13).
CSEB (Conseil scientifique de l'environnement de Bretagne), 2005c. Fiche E-9. Méthode d’extrapolation spatiale des données de débit, http://www.cseb-bretagne.fr/downloads/CSEB_Recueil%20Fiches_Tomes1+2.pdf, (16.01.13).
Cugier P., Ménesguen A. & Guillaud J.F., 2005a. Three-dimensional (3D) ecological modelling of the Bay of Seine (English Channel, France), 2005. J. Sea Res., 54, 104-124.
Cugier P. et al., 2005b. Modelling the eutrophication of the Seine Bight (France) under historical, present and future riverine nutrient loading. J. Hydrol., 304, 381-396.
Guillaud J.F., Andrieux F. & Ménesguen A., 2000. Biogeochemical modelling in the Bay of Seine/ France: an improvement by introducing phosphorus in nutrient cycles. J. Mar. Syst., 25, 369-386.
Howarth R.W. et al., 1996. Regional nitrogen budgets and riverine N & P fluxes for the drainages to the North Atlantic Ocean: natural and human influences. Biogeochemistry, 35, 75-139.
Howarth R.W. et al., 2006. The influence of climate on average nitrogen export from large watersheds in the Northeastern United States. Biogeochemistry, 79, 163-186.
Lancelot C. et al., 2007. Testing an integrated river–ocean mathematical tool for linking marine eutrophication to land use: the Phaeocystis-dominated Belgian coastal zone (Southern North Sea) over the past 50 years. J. Mar. Syst., 64, 216-228.
Lancelot C. et al., 2011. Cost assessment and ecological effectiveness of nutrient reduction options for mitigating Phaeocystis colony blooms in the Southern North Sea: an integrated modeling approach. Sci. Total Environ., 409, 2179-2191.
Ménesguen A. & Piriou J.Y., 1995. Nitrogen loadings and macroalgal (Ulva sp.) mass accumulation in Brittany (France). Ophelia, 42, 227-237.
Moatar F. et al., 2009. Incertitudes sur les métriques de qualité des cours d'eau (médianes et quantiles de concentrations, flux, cas des nutriments) évaluées à partir de suivis discrets. Houille Blanche, 3, 68-76.
Perrot Th., Ménesguen A. & Dumas F., 2007. Modélisation écologique de la marée verte sur les côtes bretonnes. Houille Blanche, 5, 49-55.
Thieu V., Garnier J. & Billen G., 2009. Assessing the effect of nutrient mitigation measures in the watersheds of the Southern Bight of the North Sea. Sci. Total Environ., 408, 1245-1255.
Thieu V., Billen G., Garnier J. & Benoît M., 2010. Nitrogen cycling in a hypothetical scenario of generalised organic agriculture in the Seine, Somme and Scheldt watersheds. Reg. Environ. Change., 11, 359-370.
Pour citer cet article
