
Faidherbia albida trees form a natural buffer against millet water stress in agroforestry parklands in Senegal
Résumé
Faidherbia albida forme un tampon naturel contre le stress hydrique du mil dans les parcs agroforestiers au Sénégal
Description du sujet. Au centre-ouest du Sénégal, les parcs agroforestiers dominés par Faidherbia albida pourraient protéger la culture du mil contre la sécheresse et augmenter la production agricole. Seules quelques études ont examiné les relations hydriques entre les arbres et le mil dans les conditions des champs des agriculteurs.
Objectifs. Notre objectif était d’explorer l’effet des arbres sur le potentiel hydrique foliaire du mil et les implications pour le rendement du mil dans les parcs à F. albida.
Méthode. Soixante-huit parcelles de F. albida dispersées dans cinq villages ont été sélectionnées pendant la saison des pluies 2019. Dans chaque parcelle, une paire de placettes de mil « placette-proche-arbre » et « placette ouverte » a été suivie. Nous y avons mesuré les potentiels hydriques des feuilles de mil, avant l’aube et à midi (Ψp, Ψm), la densité apparente, la teneur en eau et la température en surface sol, ainsi que les rendements en grain et en paille à la récolte.
Résultats. Les comparaisons par paires ont révélé un statut hydrique de la culture meilleur et plus stable, ainsi qu’une densité apparente et température des sols souvent plus faibles dans les placettes proches des arbres. L’humidité du sol était parfois plus basse dans les placettes proches des arbres, probablement parce que la croissance plus élevée du mil dans ces positions a entrainé une transpiration plus importante. Les effets de l’arbre sur le rendement en grains de mil et en paille ont été souvent positifs ou nuls. Les corrélations entre le rendement du mil et Ψp, Ψm mesurées autour de la floraison ont été faibles.
Conclusions. Tenant compte de leurs effets positifs fréquents sur le statut hydrique et le rendement du mil, les F. albida peuvent jouer un rôle clé pour une agriculture durable dans le contexte de changement climatique. L’augmentation du rendement du mil due à la proximité des arbres est probablement liée aux changements induits par les arbres sur de multiples ressources environnementales. Les interactions des conditions microclimatiques de l’élevage et de la gestion des arbres dans la région (densité, intensité d’élagage) sur les effets des arbres doivent être le centre d’intérêt des travaux futurs.
Abstract
Description of the subject. In central-west Senegal, agroforestry parklands dominated by Faidherbia albida trees might protect the millet crop against drought and increase agricultural production. Only few studies examined the water relations between the trees and the crop in farmers’ field conditions.
Objectives. Our aim was to explore the effect of F. albida trees on millet water potential and the implications for millet yield in F. albida parklands.
Method. Sixty-eight F. albida plots scattered in five villages were selected in the 2019 rainy season. In each plot, a pair of millet subplots, “close-to-tree subplot” and “open subplot”, was monitored. We measured millet predawn and midday leaf water potential (Ψp, Ψm), top soil bulk density, water content and temperature, and grain and straw yields at harvest.
Results. Pairwise comparisons of Ψp and Ψm of millet indicated better and more stable water status and lower soil temperature and bulk density in close-to-tree subplots. Soil water was sometimes lower in close-to-tree subplots, perhaps because the higher crop biomass in this location resulted in higher water loss through crop transpiration. Tree effects on millet grain and straw yield were positive or null. Correlations between millet yield and Ψp, Ψm measured around flowering were weak.
Conclusions. Given their common positive effects on millet water status and yield, F. albida trees could play a key role in promoting sustainable agriculture under the changing climate conditions. Millet yield increase due to the tree proximity was likely due to changes induced by the tree on multiple environmental resources. The interactions of micro-pedoclimatic conditions, livestock and tree management (e.g. density, pruning intensity) on the tree effects should be the focus of future studies.
Received 29 September 2022, accepted 26 September 2023, available online 5 October 2023.
This article is distributed under the terms and conditions of the CC-BY License (http://creativecommons.org/licenses/by/4.0)
1. Introduction
1Water constraints due to the climatic change are expected to increase in the world and particularly in sub-Saharan Africa (IPCC, 2022). Trees associated with crops in the same fields are currently recognized as valuable for mitigating the effect of climate change on the sustainability of cereal crops, in contrast to the recent past when they were seen as obstacles to agricultural intensification of annual food crops (Mbow et al., 2014; Zomer et al., 2014; Skole et al., 2021). In sub-Saharan Africa, the case of Faidherbia albida tree parklands particularly attracted the researchers’ attention. Most studies confirmed farmers’ perception of a beneficial effect of tree proximity on soil fertility and cereal yield (Sileshi et al., 2016; Sida et al., 2018a; Kuyah et al., 2019; Roupsard et al., 2020).
2Senegalese F. albida parklands are original examples of these agroforestry systems. Pelissier (1966) and Lericollais (1972) described them as the most constructive landscape of the agrarian civilization developed by the Sereer communities under the high increase rates of population and demand for food. The system is known to deliver good trade-offs between tree growth, staple food crop production, livestock feeding, and recycling of environmental resources. Faidherbia albida is defoliated during the rainy season when the associated millet/groundnut rotation receiving no or low fertilizer inputs, is cultivated in the same field. Therefore, the trees would not exert significant competition on the annual crops for light and other resources (Roupsard et al., 2020). Additionally, the trees provide various provisioning services as high quality forage and edible fruits with high nutritional quality for the farm livestock during the hunger period, wood, traditional medicine, pollen and nectar for bees and other insects (Sileshi et al., 2020). During the dry season, their canopies serve as a shelter for the livestock moving freely throughout the harvested fields. In return, livestock droppings improve soil fertility under the trees. As a legume, F. albida tree has the potential to symbiotically fix atmospheric nitrogen. Faidherbia albida fine roots are mostly concentrated under the tree with half of the fine roots located in the 40-100 cm soil layer, and almost half in the 100-200 cm (Siegwart et al., 2022). Only a small portion (less than 5%) of the F. albida fine roots is located in the top soil (0-40 cm soil layer), either under the tree (Siegwart et al., 2022, 2023). By contrast, the medium roots of F. albida are more likely distributed far from the tree at depth (100-150 cm) indicating a wide and deep root colonization around the trees (Siegwart et al., 2022, 2023), and suggesting a high potential facilitation role of F. albida tree in agroforestry systems (Gning et al., 2023). As a phreatophitic tree, we may expect deep fine roots of F. albida trees reaching the water table (Roupsard et al., 1999) to be useful to improve the carbon sequestration in deep soil layers (Siegwart et al., 2022), and to capture water and nutrients and recycling them through the litter of leaves, towards the top soil, at the benefit of annual intercrops. Yengwe et al. (2018) showed that tree litter decomposes at the soil surface enriching the soil with organic matter and nutrients, but also at various soil depths through tree and crop root litters (Siegwart et al., 2022). Various studies showed a significantly higher soil nutrients and carbon stock at the F. albida tree proximity (Sileshi et al., 2016; Siegwart et al., 2022). They suggested that such soil fertility improvement contributed to the millet yield increase around F. albida trees.
3Water is a major limiting factor for the cereal yield in sub-Saharan area. Depending on their shape, tree canopies in agroforestry systems may affect both the intensity of rainfall water reaching the ground, and redistribution into throughfall and stemflow (Antoneli et al., 2021). Soil water evaporation may be reduced because of the microclimate developed under the tree canopy (Padovan et al., 2018). Increased soil organic matter at the tree proximity may reduce soil bulk density, improve soil porosity and water infiltration and retention (Kuyah et al., 2019; Stephen et al., 2020). Some studies showed that changes in tree root distribution (Forey et al., 2017; Clermont-Dauphin et al., 2018), and/or in soil biological activity (Rivest et al., 2013) may improve the resilience of agroforestry systems to climatic risks. Faidherbia albida trees might improve the water status and the water use efficiency of the associated millet crop in the Senegalese parklands through hydraulic redistributions (Diongue et al., 2023; Gning et al., 2023). This would be particularly helpful during the late vegetative phase of millet, which corresponds to the period of highest crop water demand but also to the period of decreased rainfall in sub-Saharan regions. In this period, more than 90% of millet roots are concentrated in the first 0-40 cm depth, while most F. albida roots are deeper and may reach the water table (Siegwart et al., 2022).
4While numerous studies have examined the beneficial effect of F. albida tree on both soil chemical fertility and millet yield, the general effect of the tree on water status of the associated millet within the Sahelian parklands, and the implications for the millet yield largely remains unexplored. Mc Intyre et al. (1995) found millet predawn leaf water potential values (Ψp) between -0.1 and -0.6 MPa (Mega-Pascals) with water stress starting around -0.3 MPa, and midday values (Ψm) between -0.5 and -1.6 MPa with stress starting around -0.6 MPa. Do et al. (1996) reported midday values between -0.5 and -2.3 MPa with water stress starting around -1.2 MPa. The large range of midday leaf water potential suggested an anisohydric trend in pearl millet, which was supported by a recent study of Cai et al. (2020) on changes in transpiration versus leaf water potential. Anisohydric behavior also indicated that the leaf water potential would be a very sensitive indicator of soil drying in the root zone of millet crops.
5The overall objective of this study was to highlight the effects of trees on millet water status, and millet productivity in F. albida parklands. Specific objectives were to examine:
6– how the proximity of F. albida trees affected the water status of millet crop during short drought periods occurring in the reproductive phase of the crop cycle;
7– the links between the water stress intensity during the reproductive phase of the crop, and the millet yield.
8We used an original screening-approach with punctual and rapid measurements carried out in a large network of Senegalese farmers’ fields under one of the most common agroforestry systems in West Africa.
2. Materials and methods
2.1. Study area
9The study area covers approximately 45 km2 in central-west Senegal. It extended from the village of Touba Toul (14.819926°-16.661553°) to Niakhar (14.475930°-16.403219°). The climate is Sahelo-Sudanian with a long dry season between November and June, and a short rainy season between July and October receiving 600 mm·year-1, on average over the 2000-2020 period. This average derived from the records of the National Agency for Civil Aviation and Meteorology (ANACIM, unpublished) at Bambey station (14°42'N, 16°28'W). The soils are classified as arenosols (FAO-UNESCO, 1976). They have a sandy texture, a pH water slightly under neutral and low CEC. Tounkara et al. (2020) underlined the very low organic matter content and phosphorus availability of the soils.
2.2. The Senegalese F. albida parkland agro-system
10In the study area, 60 different native trees species have been inventoried, with F. albida being the most abundant with around 43% of trees (Leroux et al., 2022). Comparisons between the trees inventory made in 1965, 1985 and 2005 in one of the villages of the study area indicated that the number of trees decreased by 40% in forty years. Young trees were totally absent in the inventory of 1985 and regeneration was still poorly assured in 2005 (Delaunay al., 2009). Following a regression equation using the diameter at breast height (DBH) proposed by Sida et al. (2018b), the tree ages would range from 50 to 150 years.
11The tree distribution within a village is often highly heterogeneous, with tree density varying between 0-1 to 10-20 tree·ha-1 over short distances, as shown by Delaunay et al. (2009) for a village of the study area. Most F. albida trees are concentrated in the farthest ring of cultivated fields around the village, which is also the most degraded one. This ring is mostly that of the biennial rotation of millet/groundnut, receiving only few organic and/or inorganic fertilizer since farmers preferentially allocate their low availability of nutrient resources to fields close to the homestead (Tounkara et al., 2020).
12The main farmers’ intervention is tree pruning every two to four years, to provide fodder to the livestock. We are not aware of any recent quantitative evaluation of pruning intensity in our study area in Senegal, but based on our rough visual estimation, at least 50% of the canopy of most trees are pruned every two to four years with inappropriate methods causing injuries and predisposing the trees to attack by pathogens. This visual estimate crosschecks the quantitative evaluation of Depommier & Guerin (1996) in F. albida parklands of Burkina Faso. The millet crop is typically sown before the first rains of the season. Soil preparation before sowing consists in a superficial tillage with manual hoes. Farmers often use the Souna variety, a synthetic cultivar, obtained in the research station of Bambey by crossing various pure lines (CEDEAO, 2017). The sowing is made in hills at a density of 12,000 hills·ha-1, corresponding to distances of 90 x 90 cm between and within the rows respectively. About three seedlings per hill are maintained after an early thinning and weeding that occur about ten days after the seedlings emergence. Two manual weedings are generally performed at about one month and two months after seedlings emergence before harvesting. Sometimes tree density at the farmer’s plot level is high (Figures 1 and 2).

Figure 1. Illustration of the tree density in different villages in the study area (extracted with permission from Leroux et al., 2022) — Illustration de la densité d’arbres dans différents villages de la zone d’étude (extraite avec la permission de Leroux et al., 2022).

Figure 2. One of the most wooded Faidherbia albida parkland of the study area (45 trees.ha-1). This photo was taken on August 28, 2019 in the village of Thila Ounté (14.798932°, -16.685310°) — Un parc à Faidherbia albida parmi les plus boisés de la zone d’étude (45 arbres.ha-1). Cette photo a été prise le 28 aout 2019 dans le village de Thila Ounté (14.798932°, -16.685310°).
2.3. Experimental design
13The study was carried out during the 2019 wet season over a network of 68 farmers’plots selected in five villages of the study area, namely Thila Ounté, Toukar, Poultock-Diohine, Toucar and Diohine. Figure 3 shows a map of the location of the five villages in the center west of Senegal, extracted from Leroux et al. (2022). A plot was defined as a surface area of at least 100 m2, including F. albida trees scattered in the field as homogeneously as possible. All the selected plots had the typical millet crop management described above for the Senegalese F. albida parkland. Their topography, soil physical characteristics and general climate were similar. In each of the 68 plots, we monitored a pair of millet subplots, measuring 4 x 4 m each. One (referred to as “close-to-tree”) was installed at 1 m from the tree trunk. The other (referred to as “open subplot”) was at mid distance between two trees, generally more than 17 m away from all trees, which is considered the limit for tree influence on the millet crop in this area (Roupsard et al., 2020). Care has been taken to install the subplots following the orientation of the tree canopy. This was in often the South orientation. Throughout this network of plots, the average diameter of tree trunk at 1.30 height, total tree height and crown surface amounted 70 ± 21 cm, 12 ± 2 m, and 117 ± 85 m2 respectively.
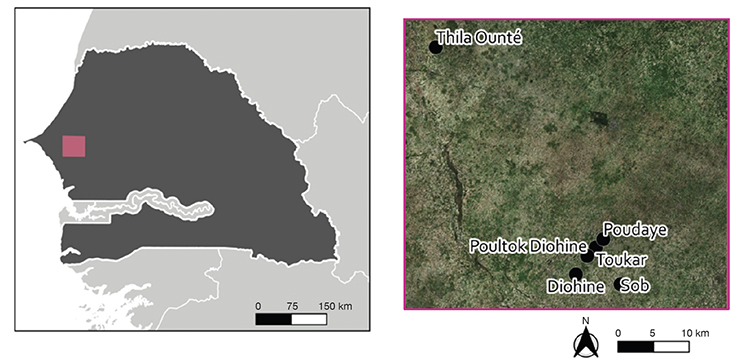
Figure 3. Map of the location of the study area in five villages of central Senegal — Carte de l’emplacement de la zone d’étude dans cinq villages du centre du Sénégal.
2.4. Measured variables
14Rainfall amount was measured from a meteorological station installed in the “Faidherbia-Flux” observatory in the village of Sob (FLUXNET, SN-Nkr; https://lped.info/wikiObsSN/?Faidherbia-Flux, N: 14°29'44.916''; W: 16°27'12.851'') (Roupsard et al., 2019). Moreover, we installed manual rain gauges in the village of Thila Ounté and Diohine respectively. The following variables were collected in each pair of subplots.
15Predawn leaf water potential (Ψp) and midday leaf water potential (Ψm) of millet crop. Millet crop water potentials were measured with a hydraulic press (Seghieri, 2010). Ψp measurements were carried out between 05:30 am and 06:30 am on the last liguled leaf of five main shoots of millet collected from five hills in each subplot. Ψm measurements were carried out the same day as Ψp, between 12:30 am and 01:30 pm. Dates of records corresponded to 35-40 days (before millet flowering), and 50-60 days (after millet flowering) after millet emergence (Figure 4). Ψp is the most used indicator of the plant water status versus soil water constraint. It is assumed in relative equilibrium with the soil water potential in the root zone (Novick et al., 2022; Rodriguez-Dominguez et al., 2022). Ψm is the most negative value of the day. It is induced by the transpiration and represents the highest water stress. Seghieri (2010) showed high correlation between the potential values recorded with a hydraulic press and the reference Scholander chamber. The small size and low weight of the hydraulic press, and the availability of one material per village and a staff of three operators per village, facilitated our approach of screening throughout a large network of farmer’s fields.
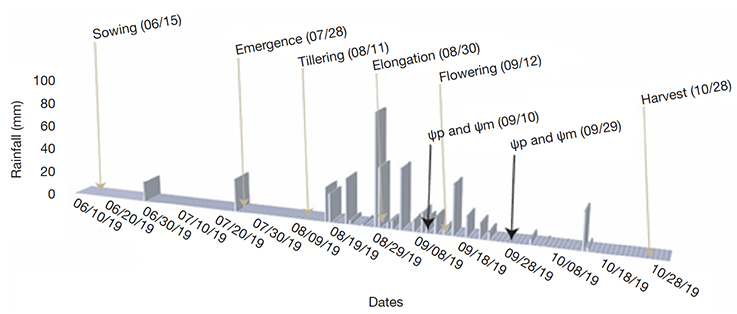
Figure 4. Rainfall distribution in 2019 at Sob village, dates of phenological stages of millet and dates of leaf water potential records — Répartition des précipitations en 2019 dans le village de Sob, dates des stades phénologiques du mil et dates des enregistrements du potentiel hydrique des feuilles.
16Surface soil bulk density water content and temperature. Soil bulk densities were measured in both 0-10 cm and 10-20 cm soil layers 47 days after millet emergence. We used a steel cylinder 5 cm high and 100 cm3 volume. We collected four replications on the diagonal of each subplot. Surface soil water content and temperature were measured in the 0-10 cm soil layer, with a portable direct reading-humidimeter (Fieldscout, TDR150) and provided calibration. The measurements were carried out at 47, 53 and 62 days after millet crop emergence, with four replications by subplot. Only the data of two villages (Thila Ounté and Diohine) were available. Surface soil water measurements in the 0-10 cm layer were taken as indicator of surface soil drying difference between pair of subplots. Indeed this measure cannot be considered fully representative of the water availability of millet during the crop cycle since, as shown by Siegwart et al. (2022) in a F. albida parkland of the study area, millet roots may extend until 200 cm depth, whereas most of them are concentrated in the first 0-40 cm depth.
17Grain and straw yields of millet at harvest. Grain and straw yields were determined from a sampling area (8 m2) located in the center part of each subplot. They were expressed in dry matter basis, by drying samples at 65 °C for 48 h.
2.5. Statistics
18The resulting design of our farmers’ field network was a 5 (Villages) by 2 (Date of measures) by 2 (Subplot positions “close-to-tree” or in “open subplot”) analysis of variance when Ψp and Ψm were the dependent variables, and it was a 2 (Villages) by 3 (Date of measures) by 2 (Subplot positions “close-to-tree” or in “open subplot”) analysis of variance when soil water content and temperatures were the dependent variables. It was a 5 (villages) by 2 (subplot positions “close-to-tree” or in “open subplot”) analysis of variance when millet grain and straw yields were the dependent variables. The date and the subplot position factors in the anovas are within-subject or repeated measures factors because they represent repeated measurements on the same subject. Linear regressions between variables were analyzed with GLM. We used the software STATISTICA (StatSoft. Inc. 2007).
3. Results
3.1. Rainfall distribution
19Rainfall amount reached 473 mm in 2019 at Sob village. After two isolated rainfall events in June and July, the rainy season started at mid-August. Farmers had to make two to three re-sowings. During the flowering period of millet, rainfall distribution was irregular, with several short episodes without rain during at least 5 days (Figure 4). In the village of Thila Ounté, which is the furthest to Sob village, total rainfall amounted to 441 mm. In the village of Diohine, which is very close to Sob, the total rainfall amounted to 495 mm.
3.2. Effect of tree proximity on millet leaf water potential
20Ψp and Ψm in close-to-tree were slightly but significantly higher than in open-field subplots at both dates of observations. Lowest values were reached at the second date of measurement, corresponding to the end of the millet reproductive phase, the Ψp values supported a significant water constraint in both subplot situations. The interactions between factors were statistically significant (Table 1a). An increasing difference in Ψp between close-to-tree and open subplots when open subplot conditions become more severe seems to be developing (Figures 5a and 5b). Ψm values showed similar trends (Figures 5c and 5d). Overall, trees had beneficial effects for millet Ψm and Ψp during the studied year, despite significant differences between villages (Table 1b).
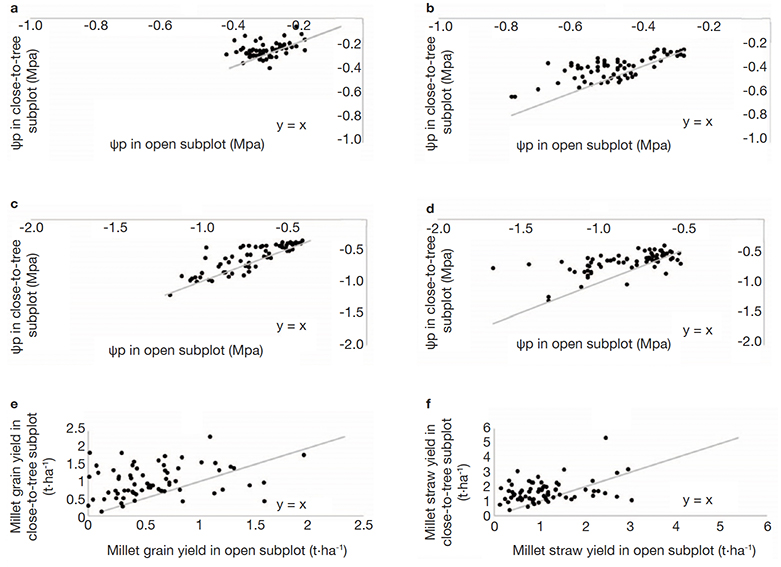
Figure 5. Pairwise comparisons of the following variables measured in open and close-to-tree subplots of five villages of central Senegal: a, b. Predawn leaf water potential of millet before and after millet flowering respectively; c, d. midday leaf water potential of millet before and after millet flowering respectively; e, f. millet grain and straw yield respectively. In a, b, c, d, each point is average of four repetitions collected in 68 pairs of subplot. In e, f, each point is from a 8 m2 sampling area located in the center part of each subplot — Comparaisons par paires des variables suivantes mesurées dans des sous-parcelles en plein soleil et proches des arbres de cinq villages du centre du Sénégal : a, b. potentiel hydrique foliaire avant l’aube du mil avant et après la floraison du millet respectivement ; c, d. potentiel hydrique foliaire du mil avant et après la floraison du millet, respectivement ; e, f. le rendement en grains et en paille de mil, respectivement. En a, b, c, d, chaque point est la moyenne de quatre répétitions recueillies dans 68 paires de sous-parcelle. En e, f, chaque point provient d’une zone d’échantillonnage de 8 m2 située dans la partie centrale de chaque sous-placette.
3.3. Effect of tree proximity on soil water, temperature and bulk density
21Overall topsoil water content and temperature varied significantly with main effects and interactions of all the studied factors (Tables 1c and 1d). In the village of Thila Ounté, close-to-tree subplots had slightly higher soil water content (Figure 6c) and lower soil bulk density and temperature (Figures 6a and 6e). In the village of Diohine where the soil water covered a larger range than in Thila Ounté, the soils in close-to-tree subplots are sometimes dryer than in open subplots (Figure 6d). This was particularly observed for the highest range values of soil water contents in open subplots. Close-to-tree subplots of Diohine showed no difference in soil bulk density (Figure 6b) and sometimes lower temperature (Figure 6f) in comparison to open subplots.
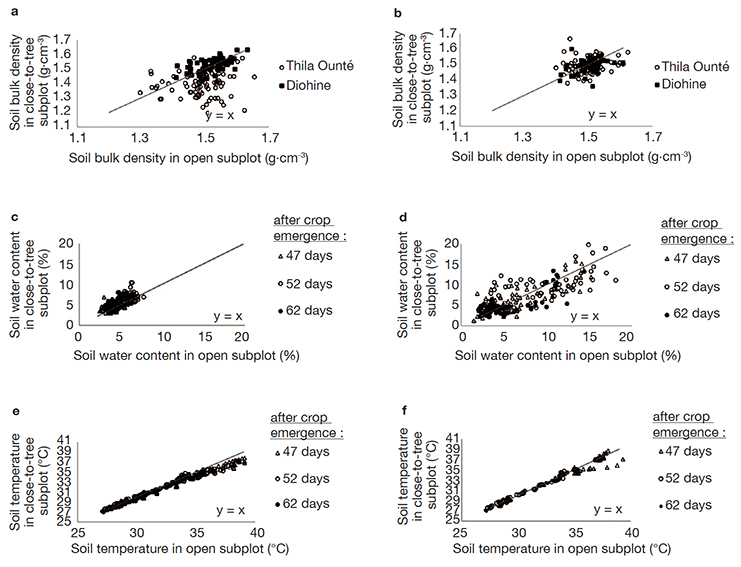
Figure 6. Pairwise comparisons of the following variables measured in open and close-to-tree subplots in two villages of central Senegal (Thila Ounté and Diohine): a, b. Soil bulk density at 47 days after emergence; c, d. Soil water content at three dates; e, f. Soil temperature at three dates. There were four repetitions collected in 35 pairs of subplot respectively. Each point is a raw data — Comparaison par paires des variables suivantes mesurées dans les sous-placettes en plein soleil et proches des arbres de deux villages du centre du Sénégal (Thila Ounté et Diohine) : a, b. Densité apparente du sol 47 jours après la levée ; c, d. Teneur en eau du sol à trois dates ; e, f. Température du sol à trois dates. Il y eut quatre répétitions recueillies dans 35 paires de sous-placettes respectivement. Chaque point est une donnée brute.
3.4. Effect of tree proximity on millet yield
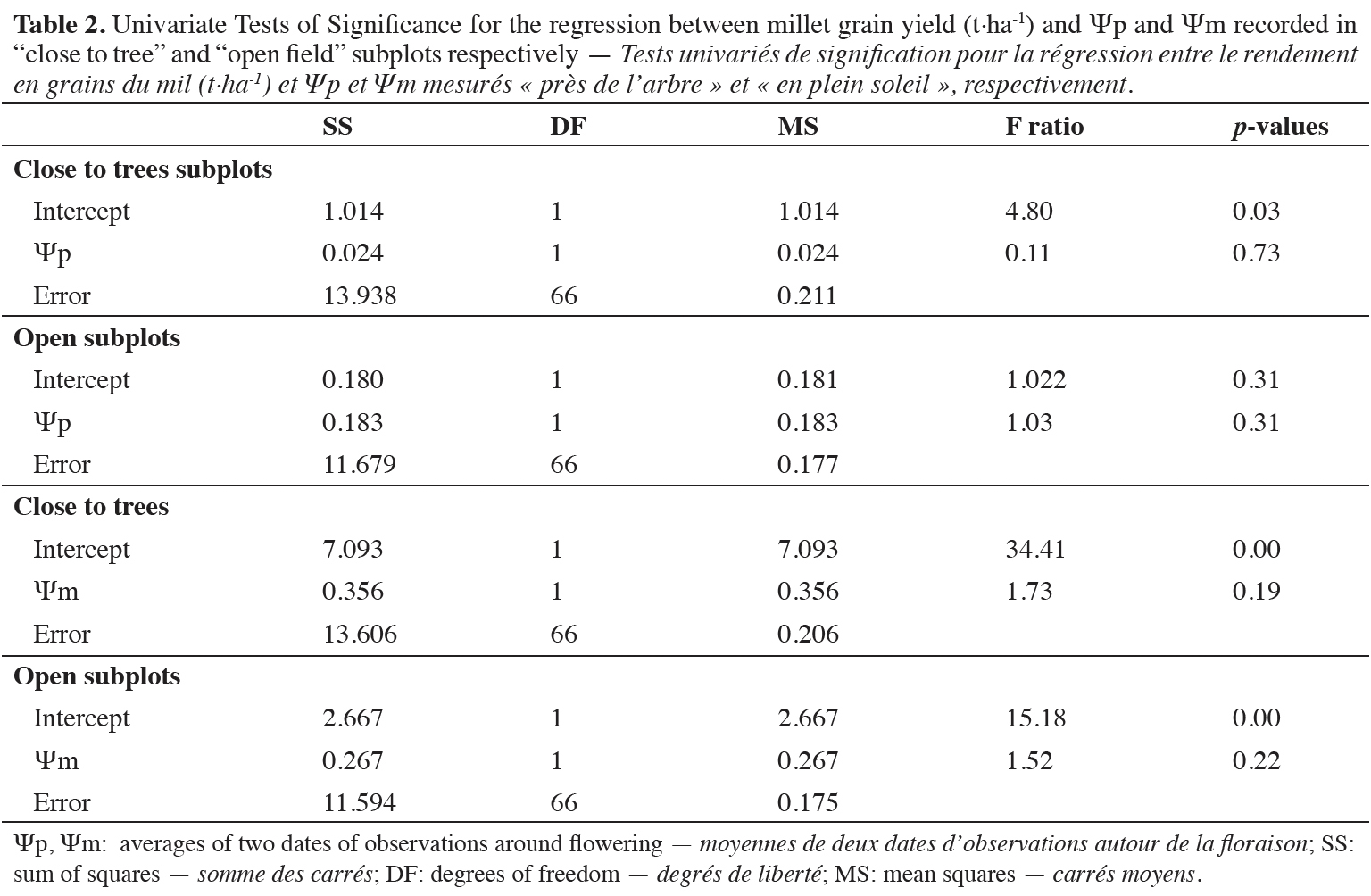
22Tree proximity increased millet grain and straw yields over the network (Figures 5e and 5f). Hot spots of higher millet growth are clearly visible around the tree in some situations (Figure 7). Correlations between millet yield and leaf water potentials measured around flowering were not statistically significant (Table 2).

Figure 7. Illustration of the “fertility islands” for millet growth around Faidherbia albida. This photograph has been taken on September 30, 2019 in the village Thila Ounte (14. 795532, -16. 69585) — Illustration des « îlots de fertilité » pour la croissance du mil autour de Faidherbia albida. Cette photographie a été prise le 30 septembre 2019 dans le village de Thila Ounte (14. 795532, -16. 69585). © Leroux L, 2019.
4. Discussion
23In this study, we show that F. albida trees have a positive effect on millet water status and yield. The study was done in an exceptionally dry year, underlining the power of F. albida in mitigating upcoming extreme climate change effects, which are expected to lead to more severe droughts in the study area. The year 2019 indeed was a very low and late rainfall year according to the records of ANACIM at Bambey. The rainfall of 2019 (528 mm) was lower than the median value (602 mm) of the 2000-2020 period (except 2019) and close to the lower quartile (476 mm). The first two rainy months of 2019 at Bambey station, June and July, had only 5 and 45 mm·month-1 respectively. These amounts were close to the lower quartile of June (2 mm) and far under that of July (88 mm) of the 2000-2020 period. This low rainfall year was particularly favorable to highlight the water relations between trees and the millet crop, and the potential implications for resilience of millet productivity towards drought in F. albida tree agroforestry systems.
24Whereas the villages were close to each other, and the millet crop management considered as homogenous over the network, the “village” effects were mostly statistically significant and interacted with the tree beneficial effect on yield. The heterogeneity of micropedoclimatic conditions over the network may have been in cause. Consistently, our rain gauges indicated differences of rainfall between the village of Thila Ounté and those of Sob and Diohine.
25The values of predawn leaf water potential at the flowering stage of the pearl millet were often under the critical threshold of -0.40 MPa. These results supported that the crop water stress was high (McIntyre et al., 1995). Hence, we showed that the crop water status as indicated by Ψp was improved close to trees. This advantage increased when the water stress increased in the open subplot plots (i.e. when Ψp decreases in open subplot). This suggested that the tree proximity contributed to improve and maintain millet water status relatively stable as drought intensity increased in open subplots. The crop water status at midday (Ψm) was also significantly improved by the tree proximity and remained in most cases above the value of -0.8 Mpa in the close-to-tree subplots, while it continued to decrease in the corresponding open subplots.
26Our results indicated that various processes may have contributed to improve millet water status at tree proximity. Increased water infiltration close to the trees was suggested by the observed decrease of soil bulk density in the close-to-tree subplots. Also, reduced soil evaporation was suggested by the lower soil temperature recorded in the close-to-tree subplots. These results were consistent with Sida et al. (2018a), De Frenne et al. (2019) and Kuyah et al. (2019). However, our results also suggested that the tree proximity effects were not always as favorable as expected. Whereas positive effects of tree proximity on the soil variables are common, null or even negative effects also occurred. The top soil moisture in the close-to-tree subplots sometimes decreased, particularly when the rainy season was well established. This might have been due to the higher water withdrawal by crops that have grown and yielded significantly more in the close-to-tree subplots than in the open subplots. Such mechanism might adversely affect the crop yield in some years when the drought is intensifying. A similar analysis was proposed by Tounkara et al. (2020) for the most manured millet fields in the study area. Clearly, more detailed monitoring of the soil water budget and temperature in close-to-tree and open subplots are needed, with special attention to identification of the interactions of period of the cropping season, time of the day, physiological stage and growth of the crop, tree pruning intensity and growth.
27Millet management was kept as homogenous as possible over our field network to not contribute to the variability of millet water status over the network. In contrast, various components of tree management (such as pruning intensity, trunk size and average distance between trees) were highly variable. These may have interacted with the tree effects on the crops as shown by Toib et al. (2021) for pruning practices in Ethiopia. Our results attracted the attention on the need to avoid rapid generalization of results acquired in one site and under one tree management. Moreover, we should be aware that interactions between the tree and the crop in the Senegalese F. albida parklands could involve not only on the tree canopy extent with its benefits in terms of microclimate, soil and plant water status and nutrient facilitations for the crop, but could also involve the underground root area, which may be wider than that of the canopy area (Siegwart et al., 2022; Diongue et al., 2023 ; Gning et al., 2023; Siegwart et al., 2023).
28The millet yields confirmed the beneficial effect of F. albida on the land use efficiency reported by Roupsard et al. (2020) from drone imagery data in farmers’ fields of this area. Whereas the trees appeared mostly severely pruned, the positive effect of F. albida trees on millet yield was still significant in average. Non-significant correlations between Ψp, Ψm and millet yield suggested that the beneficial effect of tree proximity on millet yield is not solely due to better water crop status. This interpretation was consistent with the results showing that other key resources of the environment particularly soil N and P availability have been also improved close to F. albida trees (Sileshi, 2016; Stephen et al., 2020; Siegwart et al., 2022).
29Overall, the positive effects of the tree on millet crop water status and yields might advocate for increasing tree densities and/or extending the parklands, but this is not simple. Indeed, the climate change and the increase of demographic pressure have deteriorated the socio-economic conditions of many farmers. Socio-economic factors of farms are keys in determining farmers’ management decisions. The reduction of tree density over the years reported by Delaunay et al. (2009) for this area may express the farmers’ priority to use more land for growing staple food crops. The choice of intense tree pruning practices, despite their negative effects on survival and regeneration of F. albida parklands (Sida et al., 2018b), may be that of farmers which considered direct tree provisioning services as priority comparing to other potential environmental services. It is not realistic to think that farmers will modify their practices to follow researchers’ recommendations when these have been exclusively based on the purpose of storing more carbon and mitigating climate change. It is likely that climate change mitigation would be just a nice bonus for many of them. As stated by Meijer et al. (2015), the diversity of farmers’ socio-economic ambitions, material resources, labor force, knowledge, as well as their global view of field characteristics should play a key role in targeting sustainable innovations.
5. Conclusions
30Mixing Faidherbia albida trees with millet crops appeared as a viable strategy to improve millet water status and yield in the changing climate conditions of Senegal. Our results suggested that the interactions between the tree, the soil and the crop components were complex, with various direct positive effects, and potential adverse feedback effects on the soil water dynamic and crop yield. Given this complexity, a systemic approach based on a conceptual model of the major mechanisms and associated variables that govern the performances of F. albida parklands is needed, so that we can effectively help farmers optimize their practices.
31The diversity of tree management practices in Senegalese parklands deserves further attention when evaluating the beneficial effects of the trees. This diversity is especially important to integrate in our recommendations since it both reflects the variation and diversity in farmers’ socio-economic constraints and individual farmers’ perceptions regarding the beneficial effects of F. albida trees. The climate change makes the installation of new parklands even more challenging than it has been in the past and should be taken into account.
32Planting more trees and managing them wisely and effectively not only increase millet production but also help in improving the wealth of ecological services of such landscapes. Together with farmers, we could pull on the blade of wool to unroll the ball of ideas for introducing more trees in the Senegalese agrosystems. Faidherbia albida parklands may turn into a source of inspiration for the design of innovative, but realistic strategies to regreen Senegalese landscapes and improve both their agricultural and non-agricultural ecological services.
Acknowledgements
33This work was supported by the UE H2020 SustainSAHEL project (Grant No. 861974) and the UE Eranet LEAP Agri-215 project Roles of Agroforestry in sustainable intensification of small farMs and food SEcurity for SocIetIes in West Africa (RAMSES II), https://www.ramsesiiagroforesterie.com/
34We are grateful to four master degree students who contributed to the field data collection: Adama Diouf, Franck Diatta, Alpha Sané and Mountaga Mboh. We are also grateful to the farmers of the study site for sharing their fields and knowledge during this research.
Bibliographie
Antoneli V., De Jesus F.C., Bednarz J.A. & Thomasz E.L., 2021. Stemflow and throughfall in agricultural crops: a synthesis. Rev. Ambiente Água, 16(1), doi.org/10.4136/ambi-agua.2528
Cai G. et al., 2020. Linear relation between leaf xylem water potential and transpiration in pearl millet during soil drying. Plant Soil, 447, 565-578, doi.org/10.1007/s11104-019-04408-z
CEDEAO-UEMOA-CILSS, 2017. Catalogue régional des espèces et variétés végétales.
Clermont-Dauphin C. et al., 2018. Intercrops improve the drought resistance of young rubber trees. Agron. Sustainable Dev., 38, 56, doi.org/10.1007/s13593-018-0537-z
de Frenne P. et al., 2019. Global buffering of temperatures under forest canopies. Nat. Ecol. Evol., 3(5), 744-749, doi.org/10.1038/s41559-019-0842-1
Delaunay V. et al., 2009. Dynamique démographique et dynamique du parc agroforestier à Faidherbia albida (Del.) A.Chev. en pays Serer (Sob, Sénégal). In : Actes de la XXVIè International Population Conference, IUSSP, Septembre 2009, Marrakech, Maroc, hal-01140561
Depommier D. & Guérin H., 1996. Émondage traditionnel de Faidherbia albida. Production fourragère, valeur nutritive et récolte de bois à Dossi et Watinoma (Burkina Faso). In : Peltier R., ed. Les parcs à Faidherbia = [Faidherbia Parklands]. Montpellier, France : CIRAD-Forêt, 55-84.
Diongue D.M. et al., 2023. A probabilistic framework for assessing the hydrological impact of Faidherbia albida in an arid area of Senegal. J. Hydrol., 622, 129717, doi.org/10.1016/j.jhydrol.2023.129717
Do F., Winkel T., Cournac L. & Louguet P., 1996. Impact of late season drought on water relations in a sparse canopy of millet (Pennisetum glaucum (L.) R. Br.). Field Crops Res., 48, 103-113, doi.org/10.1016/S0378-4290(96)01030-1
FAO-Unesco, 1976. Carte mondiale des sols 1: 5 000 000. Volume VI : Afrique. Paris : FAO Unesco.
Forey O. et al., 2017. Effect of combined deficit irrigation and grass competition at plantation on peach tree root distribution. Eur. J. Agron., 91, 16-24, doi.org/10.1016/j.eja.2017.08.008
Gning F. et al., 2023. Root distribution of Adansonia digitata, Faidherbia albida and Borassus akeassii along a climate gradient in Senegal. Agrofor. Syst., 97, 605-615, doi.org/10.1007/s10457-023-00813-7
IPCC, 2022. Climate change 2022: impacts, adaptation, and vulnerability. Contribution of working group II to the sixth assessment report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, doi.org/10.1017/9781009325844
Kuyah S. et al., 2019. Agroforestry delivers a win-win solution for ecosystem services in sub-Saharan Africa. A meta-analysis. Agron. Sustainable Dev., 39, 47, doi.org/10.1007/s13593-019-0589-8
Lericollais A., 1972. Sob, étude géographique d'un terroir Sérer (Sénégal). Paris : Mouton & Co.
Leroux L. et al., 2022. A spatialized assessment of ecosystem service relationships in a multifunctional agroforestry landscape of Senegal. Sci. Total Environ., 853, 158707, doi.org/10.1016/j.scitotenv.2022.158707
Mbow C. et al., 2014. Agroforestry solutions to address food security and climate change challenges in Africa. Curr. Opinion Environ. Sustainability, 6, 61-67, doi.org/10.1016/J.COSUST.2013.10.014
Mc Intyre B.D., Riha S.J. & Flower D.J., 1995. Water uptake by pearl millet in a semiarid environment. Field Crops Res., 43, 67-76, doi.org/10.1016/0378-4290(95)00035-o
Meijer S.S. et al., 2015. The role of knowledge, attitudes and perceptions in the uptake of agricultural and agroforestry innovations among smallholder farmers in sub-Saharan Africa. Int J. Agric. Sustainability, 13, 40-54, doi.org/10.1080/14735903.2014.912493
Novick K. et al., 2022. Confronting the water potential information gap. Nat. Geosci., 15, 158-164, doi.org/10.1038/s41561-022-00909-2
Padovan M.P. et al., 2018. Water loss by transpiration and soil evaporation in coffee shaded by Tabebuia rosea Bertol. and Simarouba glauca DC. compared to unshaded coffee in sub-optimal environmental conditions. Agric. For. Meteorol., 248, 1-14, doi.org/10.1016/j.agrformet.2017.08.036
Pelissier P., 1966. Les paysans du Sénégal. Les civilisations agraires du Cayor à la Casamance. Saint-Yrieix, France : Impr. Fabrègue.
Rivest D., Lorente M., Olivier A. & Messier C., 2013. Soil biochemical properties and microbial resilience in agroforestry systems: effects on wheat growth under controlled drought and flooding conditions. Sci. Total Environ., 463-464, 51-60, doi.org/10.1016/j.scitotenv.2013.05.071
Rodríguez Domínguez C.M. et al., 2022. Leaf water potential measurements using the pressure chamber: synthetic testing of assumptions towards best practices for precision and accuracy. Plant Cell Environ., 45(7), 2037-2061, doi.org/10.1111/pce.14330
Roupsard O. et al., 1999. Reverse phenology and dry‐season water uptake by Faidherbia albida (Del.) A. Chev. in an agroforestry parkland of Sudanese west Africa. Funct. Ecol., 13, 460-472, doi.org/10.1046/j.1365-2435.1999.00345.x
Roupsard O. et al., 2019. “Faidherbia-Flux”: adapting crops to climate changes in a semi-arid agro-sylvo-pastoral open observatory (Senegal). In: Dupraz Chr., Gosme M. & Lawson G., eds. 4th World Congress on Agroforestry, Book of abstracts, 20-22 May, 2019, Montpellier, France. Montpellier, France: CIRAD; INRA, 44.
Roupsard O. et al., 2020. How far does the tree affect the crop in agroforestry? New spatial analysis methods in a Faidherbia parkland. Agric. Ecosyst. Environ., 296, 106928, doi.org/10.1016/j.agee.2020.106928
Seghieri J., 2010. Comparison of the measurements of leaf water potential between a hydraulic press and a pressure chamber in six Sahelian woody species. Agrofor. Syst., 80, 185-190, doi.org/10.1007/s10457-009-9256-4
Sida T.S., Baudron F., Kim H.K. & Giller K.E., 2018a. Climate‐smart agroforestry: Faidherbia albida trees buffer wheat against climatic extremes in the Central Rift Valley of Ethiopia. Agric. For. Meteorol., 248, 339-347, doi.org/10.1016/j.agrformet.2017.1010.1013
Sida T.S. et al., 2018b. Excessive pruning and limited regeneration: are Faidherbia albida parklands heading for extinction in the central Rift Valley of Ethiopia? Land Degrad. Dev., 29, 1623-1633, doi.org/10.1002/ldr.2959
Siegwart L. et al., 2022. Root litter decomposition in a sub-Sahelian agroforestry parkland dominated by Faidherbia albida. J. Arid Environ., 198, 104696, doi.org/10.1016/j.jaridenv.2021.104696
Siegwart L., Bertrand I., Roupsard O. & Jourdan C., 2023. Contribution of tree and crop roots to soil carbon stocks in a sub-Sahelian agroforestry parkland in Senegal. Agric. Ecosyst. Environ., 352, 108524, doi.org/10.1016/j.agee.2023.108524
Sileshi G.W., 2016. The magnitude and spatial extent of influence of Faidherbia albida trees on soil properties and primary productivity in drylands. J. Arid Environ., 132, 1-14, doi.org/10.1016/J.JARIDENV.2016.03.002
Sileshi G.W., Teketay D., Gebrekirstos A. & Hadgu K., 2020. Sustainability of Faidherbia albida-based agroforestry in crop production and maintaining soil health. In: Dagar J.C., Gupta S.R. & Teketay D., eds. Agroforestry for degraded landscapes. Singapore: Springer, doi.org/10.1007/978-981-15-6807-7_12
Skole D.L. et al., 2021. Trees outside of forests as natural climate solutions. Nat. Clim. Change, 11, 1013-1016, doi.org/10.1038/s41558-021-01230-3
Stephen E.A., Evans K.D. & Akwasi A.A., 2020. Effects of Faidherbia albida on some important soil fertility indicators on agroforestry parklands in the semi-arid zone of Ghana. Afr. J. Agric. Res., 15(2), 256-268, doi.org/10.5897/AJAR2019.14617
Toib A.A. et al., 2021. To prune or not to prune Faidherbia albida: competing needs for water, wheat and tree products in semi-arid Ethiopia. Agrofor. Syst., 95, 1661-1673, doi.org/10.1007/s10457-021-00675-x
Tounkara A. et al., 2020. Inorganic fertilizer use efficiency of millet crop increased with organic fertilizer application in rainfed agriculture on smallholdings in central Senegal. Agric. Ecosyst. Environ., 294, 106878, doi.org/10.1016/j.agee.2020.106878
Yengwe J., Amalia O. & De Neve S., 2018. Quantifying nutrient deposition and yield levels of maize (Zea mays) under Faidherbia albida agroforestry system in Zambia. Eur. J. Agron., 99, 148-155, doi.org/10.1016/j.eja.2018.07.004
Zomer J.R. et al., 2014. Trees on farms: an update and reanalysis of agroforestry’s global extent and socio-ecological characteristics. Working Paper 179. Bogor, Indonesia: World Agroforestry Centre (ICRAF), Southeast Asia Regional Program, doi.org/10.5716/WP14064.PDF
Para citar este artículo
