


- Startpagina tijdschrift
- volume 9 (2005)
- Effets biocides des alcaloïdes, des saponines et des flavonoïdes extraits de Capsicum frutescens L. (Solanaceae) sur Bemisia tabaci (Gennadius) (Homoptera : Aleyrodidae)


Weergave(s): 15006 (51 ULiège)
Download(s): 1072 (5 ULiège)

Effets biocides des alcaloïdes, des saponines et des flavonoïdes extraits de Capsicum frutescens L. (Solanaceae) sur Bemisia tabaci (Gennadius) (Homoptera : Aleyrodidae)

Nota's van de redactie
Reçu le 9 avril 2004, accepté le 10 septembre 2005
Résumé
Dans la perspective d’élaborer une stratégie de gestion intégrée des ravageurs associés aux cultures maraîchères au Maroc, l’activité biocide des alcaloïdes, des saponines et des flavonoïdes extraits des fruits de Capsicum frutescens L. sur la survie des œufs et des adultes de Bemisia tabaci infestant les plants de tomates, Lycopersicon esculentum Mill. var. Daniella, a été étudiée en conditions contrôlées. Les alcaloïdes, saponines et flavonoïdes dissous dans de l’éthanol à 1 % ont été appliqués à 0, 5, 10 et 20 g.l-1. Par rapport aux témoins, les trois composés affectent significativement la survie des stades suivis, selon la concentration et la durée d’exposition. Pour les œufs, la mortalité corrigée s’échelonne de 35 % à 59 % pour les alcaloïdes, de 14 % à 31 % pour les saponines et de 10 % à 14 % pour les flavonoïdes. Chez les adultes, de 29 % à 86 %, de 14 % à 48 % et de 6 % à 29 % meurent intoxiqués respectivement par les alcaloïdes, les saponines et les flavonoïdes. Oeufs et adultes répondent aux effets toxiques des composés testés linéairement dans le même sens ; les œufs requièrent cependant des concentrations plus élevées que les adultes, la CL50 est de 13,78 contre 6,83 g.l-1 pour les alcaloïdes, de 98,63 contre 32,28 g.l-1 pour les saponines et non toxique contre 120,65 g.l-1 pour les flavonoïdes. Ce sont les alcaloïdes qui se sont avérés les plus toxiques par rapport aux saponines et aux flavonoïdes. Le TL50, estimé pour les composés provoquant plus 50 % de morts, ne permet pas d’éviter l’inoculation du TYLCV par des adultes virulifères. L’utilisation de ces composés en gestion intégrée des ravageurs associés aux cultures maraîchères mérite d’être approfondie.
Abstract
Biocide effects of alkaloids, saponins and flavonoids extracted from Capsicum frutescens L. (Solanaceae) on Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae). In an attempt to elaborate a strategy of integrated pest management on vegetable crops in Morocco, insecticidal activities of alkaloids, saponins and flavonoids extracted from Capsicum frutescens L. fruits against eggs and adults of Bemisia tabaci infesting tomato plants, Lycopersicon esculentum Mill. var. Daniella, were carried out under controlled conditions. Alkaloids, saponins and flavonoids, dissolved in ethanol at 1%, were tested at 0, 5, 10 and 20 g.l-1. The three compounds affected egg and adult survival, significantly. The mortality of embryo and adult varied according to the compound considered and for each compound according to concentration and exposure duration. For eggs, the corrected mortality varied from 35 to 59% for alkaloids, 14 to 31% for saponins and 10 to 14% for flavonoids. In adults, the mortalities were spread from 29% to 86%, 14 to 48% and 6 to 29% with alkaloids, saponins and flavonoids, respectively. Significant correlation between egg and adult mortality, due to the compounds tested, were observed. However, eggs required higher concentrations than adults; the LC50 was 13.78 vs. 6.83 g.l-1 for alkaloids, 98.63 vs. 32.28 g.l-1 for saponins and non-toxic vs. 120.65 g.l-1 for the flavonoids. Therefore, alkaloids were shown more effective against B. tabaci than saponins or flavonoids. However, the LT50 estimated for compounds, killing more than 50% of the population studied, was sufficient for viruliferous adults to inoculate TYLCV to plants before they died. The use of these compounds in integrated pest management merit further study.
Inhoudstafel
1. Introduction
1La mouche blanche du cotonnier Bemisia tabaci est devenue un véritable fléau qui préoccupe de plus en plus les producteurs de cultures maraîchères dans le monde entier (Naranjo, Ellsworth, 2001). En plus des dégâts trophiques directs, l’insecte véhicule de nombreux virus sur différentes cultures. Par l’excrétion du miellat, B. tabaci affecte aussi la fonction photosynthétique et la qualité des produits récoltés (Brown, 1994 ; Traboulsi, 1994 ; Schulten, 1997).
2Au Maroc, en véhiculant le TYLCV (Tomato Yellow Leaf Curl Virus), B. tabaci génère des dégâts économiquement importants aux cultures de tomate sous serre et en plein champ (Jabbour et al., 2001). A l’instar des autres pays, la lutte contre B. tabaci repose principalement sur des pulvérisations foliaires d’insecticides de synthèse. L’utilisation intensive de ces composés conduit souvent à l’élimination d’ennemis naturels de la mouche blanche (Liu et al., 1996 ; Stansly, Liu, 1997) et à la perte de sensibilité chez la mouche (Bathee et al., 1991 ; Brown, 1994) et ce, même vis-à-vis des nouveaux insecticides tels que les nicotinoïdes et les RCI (Palumbo et al., 2001), sans oublier l’effet sur l’environnement (Van der Werf, 1997).
3Pour contrôler le ravageur sans l’inconvénient des pesticides de synthèse, il est intéressant de trouver d’autres méthodes, alternatives, en protection phytosanitaire. En effet, de nouveaux produits sont recherchés pour, d’une part, assurer une protection efficace de la production agricole, et d’autre part, contribuer à une gestion durable de l’environnement. Dans cette optique, l’utilisation d’extraits de plantes dotées d’activités insecticides offre une certaine potentialité (Larew, Lock, 1990 ; Gomez et al., 1997). De nombreux travaux ont mis en évidence les effets négatifs des extraits de plantes sur des ravageurs phytophages. Ainsi par exemple, des extraits de Melia azaderach L., Azadirachta indica A. Juss. affectent la fécondité et la mortalité de B. tabaci (Coudriet et al., 1985 ; Nardo et al., 1997 ; DeSouza, Vandramin, 2000) ; les huiles de soja et des grains du cotonnier se sont aussi montrés toxiques à la mouche blanche (Butler et al., 1988).
4Le piment fort, Capsicum frutescens, est caractérisé par l’existence de plusieurs variétés se distinguant par leurs formes, leurs couleurs et leurs pouvoirs piquants. D’un point de vue chimique, en plus des composés primaires, le genre Capsicum contient divers métabolites secondaires tels que des alcaloïdes sous formes de capsaicidine (Saber, 1976), de capsaicine (Cordell, Araujo, 1993) et de solanine (Newall et al., 1996), des saponines (De Lucca et al., 2002) et des flavonoïdes avec d’autres composés phénoliques (Materska et al., 2003 ; Zang, Hamauzu, 2003).
5En thérapeutique, plusieurs utilisations à base des fruits de C. frutescens ont été préconisées contre diverses pathologies humaines (Deall et al., 1991 ; Lopez et al., 1994 ; Therry, 1994 ; Abdel Salam et al., 1995 ; Chichewicz, Thorpe, 1996 ; Wang et al., 1999 ; Tolan et al., 2001 ; Iorizzi et al., 2002).
6En protection des plantes, la poudre et les extraits de C. frutescens ont montré un pouvoir répulsif contre Callosobruchus maculatus (F.) (Zibokere, 1994 ; Onu, Aliyu, 1995 ; Ofuya, 1986), Rhyzopertha dominica (L.) (El-Lakwah et al., 1997), Sitophilus zeamaïs Motsch et Tribolium castaneum (Herbst) (Morallo-Rejesus, 1987 ; Trematerre, Sciarretta, 2001). La toxicité des extraits des fruits du piment fort a aussi été notée chez R. dominica, S. oryzae (L.) et T. confusum J. du Val (Williams, Mansingh, 1993 ; Gakuru, Foua, 1996).
7De notre côté, et dans la perspective de rechercher d’autres produits alternatifs biodégradables et non nocifs à l’environnement, compatibles avec la gestion intégrée des ravageurs notamment en agriculture biologique, nous avons mis en évidence un effet négatif des composés ethéré, éthanolique et aqueux extraits de C. frutescens, sur l’éclosion, la fécondité et la survie des adultes de B. tabaci (Bouchelta et al., 2003). Nous avons ensuite procédé à l’extraction et au criblage de trois groupes de composés présumés actifs présents dans les fruits du piment fort. Dans ce travail, nous présentons les résultats relatifs aux effets des alcaloïdes, des saponines et des flavonoïdes, extraits des fruits de C. frutescens, sur la survie des œufs et des adultes du phytophage généraliste B. tabaci.
2. Matériels et méthodes
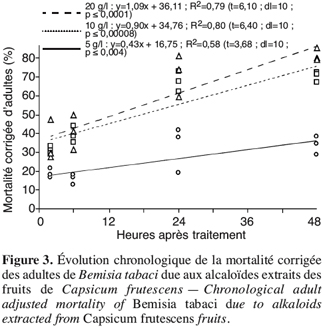
2.1. Bemisia tabaci
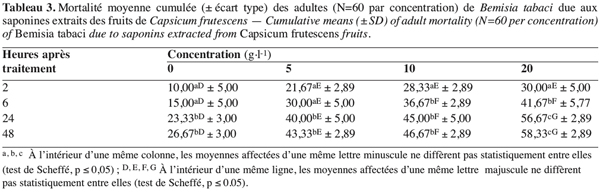
8La souche de B. tabaci, utilisée dans cette étude, a été collectée sur tomate, L. esculentum Mill., cultivée sous abri. Les adultes ont été lâchés sur des plants de la même plante, variété Daniella, en culture dans des chambres d’élevage réglées à 26 ± 1 °C, 65 ± 5 % d’humidité relative et à une photopériode de L/D : 14/10 heures. Les plants de tomate utilisés ont été cultivés dans les mêmes conditions précitées ; chaque plant avec huit feuilles, âgé de 4–5 semaines, a été transplanté dans un pot en plastique (vol = 3 l) rempli du sol local.
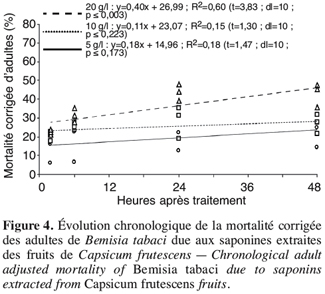
2.2. Extraction des composés à partir des fruits de Capsicum frutescens
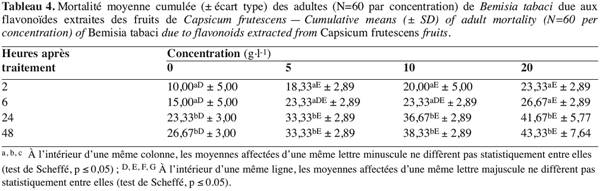
9Les fruits secs de C. frutescens ont été achetés au marché local de Meknès. Pour chaque composé ciblé (alcaloïdes, saponines ou flavonoïdes), 50 g de fruits de C. frutescens équeutés ont été séchés à l’étuve réglée à 40 °C jusqu’au poids constant ; ils ont été ensuite broyés. Des extractions sélectives des broyats ont été effectuées spécifiquement à chaque famille des composés étudiés (voir ci-après). Pour mesurer le rendement en alcaloïdes, saponines et flavonoïdes dans les fruits de C. frutescens, trois répétitions ont été effectuées pour chaque groupe de composés. Les extraits secs finaux ont été pesés et les teneurs en composés recherchés exprimées en g de composé par 100 g du poids sec de fruits équeutés. Pour répondre aux besoins des tests biologiques, plusieurs extractions ont été réalisées. Pour chaque composé, les concentrations testées sont 0, 5, 10 et 20 g d’alcaloïdes, de flavonoïdes ou de saponines par litre d’éthanol à 1 %.
10Alcaloïdes totaux. Ils ont été obtenus par triple extraction liquide – liquide selon la méthode de Ross et Rain (1977) in Harborne (1998), la poudre du fruit de C. frutescens a été extraite au soxhlet par 250 ml d’éthanol absolu pour analyse (LABOSI) durant cinq heures. L’extrait éthanolique a été ensuite évaporé à sec sous vide à 40 °C par un rotavapor type Bruchi à vitesse 4. Le résidu sec a été repris par 150 ml de chloroforme pour analyse (reag. ISO type Riedel-de Haen) et acidifié par l’HCl (type A – Fisher) à 5 % au pH 3 ; il a été laissé reposer pendant 30 minutes à la température ambiante. La phase aqueuse acide a été extraite par 150 ml de chloroforme, basifiée par le NaHCO3 (type BDH) à 5 % au pH 9 et laissée reposer pendant 15 minutes à la température ambiante. La phase chloroformique a été évaporée à sec sous vide. Le résidu sec, constitué d’alcaloïdes totaux, a été pesé puis repris par l’éthanol à 1 % pour les tests biologiques.
11Saponines. Les saponines ont été extraites selon la méthode élaborée par Applebaum et al. (1969), légèrement modifiée. Le broyat du fruit de C. frutescens a été délipidé durant deux heures par 250 ml du n-hexane pur pour analyse (reag. ACS Riedel-de Haen). Après élimination de la phase organique, le précipité obtenu a été macéré dans 300 ml d’éthanol absolu sous agitation magnétique à la température ambiante pendant 24 heures. La phase éthanolique a été évaporée à sec sous vide à 40 °C par le rotavapor. Le résidu sec a été extrait trois fois par 100 ml du mélange eau distillée/éther de pétrole (v:v) chauffé à 50 °C dans un bain-marie pendant 30 minutes. Les phases aqueuses ont été mélangées puis reprises par 150 ml de n-butanol pour analyse (type Riedel-de Haen) pendant 30 minutes. La phase organique, évaporée à sec à 40 °C par le rotavapor, a été pesée et reprise par l’éthanol à 1 % pour les tests biologiques.
12Flavonoïdes. L’extraction des flavonoïdes a été faite selon la méthode modifiée de Lee et al. (1995). Les fruits de C. frutescens réduits en poudre ont été chauffés à 90 °C sous reflux dans un mélange eau distillée/éthanol absolu pour analyse marque A Fisher (100 ml:100 ml) pendant quatre heures. L’extrait a été filtré à travers le papier filtre de 0,45 µm de porosité. La phase aqueuse a été ensuite extraite par 200 ml de n-butanol pour analyse marque Ridel-de Haen puis acidifiée par HCl à10 % au pH 3. La phase butanolique a été évaporée à sec sous pression réduite par le rotavapor à 40 °C et à vitesse 4. Le résidu sec a été extrait trois fois par 200 ml du mélange eau distillée/acétate d’éthyle pour analyse marque A Fisher (100 ml : 100 ml) pendant une heure. La phase organique a été basifiée par NaHCO3 au pH 9. Après 15 minutes de repos la phase organique (= flavonoïdes) a été évaporée à sec à 40 °C, pesée et reprise par de l’éthanol à 1 % pour les tests biologiques.
2.3. Tests biologiques
13Effets des alcaloïdes, saponines ou des flavonoïdes sur les œufs. Vingt femelles de B. tabaci, nouvellement émergées (<24 h), ont été libérées sur chaque plant de tomate avec huit feuilles, âgé de 4-5 semaines, repiqué dans un pot en plastique d’environ 3 l rempli de sol local et couvert d’une toile montée sur une armature en bois de dimensions 20 cm20 cm50 cm (longueurlargeurhauteur), pendant 48 heures. Elles ont ensuite été retirées des plants. Les feuilles de ces derniers avec les œufs de l’insecte ont été plongées dans différents extraits d’alcaloïdes, de saponines ou de flavonoïdes aux concentrations de 0, 5, 10, et 20 g d’extrait par litre d’éthanol à 1 % pendant 20 secondes puis séchées à l'air libre pendant deux heures pour évaporer l’éthanol. Les pots ont été remis dans la chambre d’élevage sous les mêmes conditions d’élevage de B. tabaci. Les plants de tomate ont été régulièrement arrosés. Après huit à dix jours, l’effectif total d’œufs éclos et non éclos a été dénombré pour chaque traitement. En moyenne 105 œufs ont été utilisés pour chaque concentration du composé testé.
14Effets des alcaloïdes, saponines ou des flavonoïdes sur les adultes. Vingt adultes, nouvellement émergés (<24 h), ont été libérés sur un plant de tomate appartenant au même stade de développement et repiqué de la même manière que précédemment. Chaque plant a été trempé au préalable, avant la mise en contact avec les adultes, dans une solution du produit testé pendant 20 secondes puis séché à l’air libre durant deux heures pour évaporer l’éthanol. Les plants avec les adultes ont été remis dans la chambre d’élevage. Les individus morts et vivants après deux, six, 24 et 48 heures ont été dénombrés ; un insecte est considéré comme mort lorsque, légèrement secoué par un pinceau, il ne bouge plus. Pour chaque concentration (i.e. 0, 5, 10 et 20 g.l-1), les essais ont été répétés trois fois. La proportion des adultes morts (= effectif de morts/effectif utilisé 100) a été calculée pour chaque concentration du composé testé.
2.4. Analyses des données
15La présentation graphique a porté sur des pourcentages des adultes morts ou des œufs non éclos. L’analyse de la variance à un ou deux facteur(s) a été conduite sur des données brutes dans le cas des variables quantitatives et normalisées (arcsin ÷(x/n)) dans le cas des proportions, au moyen d’Excel. Quand F s’est montré significatif, le classement des moyennes a été réalisé par le test de Scheffé au seuil de 5 %. Pour déterminer le temps létal 50 % (TL50), la chronologie de la mortalité de chaque stade étudié a été estimée au moyen de la régression (Excel) ; il en est de même pour les relations entre concentration - effet et entre les mortalités des stades.
16Les CL50, CL90, les pentes de droites et les intervalles de confiance ont été déterminés selon la méthode des probits (Finney, 1971) au moyen du logiciel « EPA Probit analysis program Version 1.5 » ; elles sont exprimées en grammes du composé considéré par litre d’éthanol à 1 %. Les mortalités ont été corrigées selon la méthode d’Abbott (1925).
3. Résultats
3.1. Teneurs en composés
17Les extractions spécifiques des fruits de C. frutescens ont permis de préciser les teneurs en composés recherchés (Tableau 1). En poids sec, ce sont les alcaloïdes qui sont les plus abondants (F = 1053,50 ; dl = 2 ; 3 ; p≤ 5,36E-5), avec une teneur égale à 2,36 % du poids sec de fruit. Les saponines et les flavonoïdes représentent respectivement en moyenne 0,34 et 0,17 % du poids sec du même fruit.
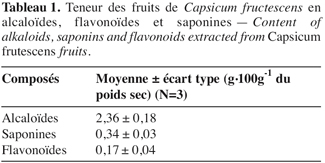
3.2. Effets des alcaloïdes, saponines ou flavonoïdes sur les œufs
18L’effet des composés, extraits des fruits de C. frutescens, sur la viabilité des embryons de B. tabaci varie entre les composés testés (F = 227,39 ; dl = 2 ; 6 ; p≤ 2,52E-16) et, pour un même composé, le pourcentage d’œufs non éclos varie selon la concentration testée (F = 316,02 ; dl = 3 ; 6 ; p≤ 2,04E-19). Les trois composés testés réduisent significativement l’éclosion des œufs par rapport au témoin (= éthanol à 1 %) (Figure 1). Le nombre d’œufs non éclos est plus élevé quand ceux-ci sont exposés aux alcaloïdes ; en moyenne, ces composés inhibent, à eux seuls, l’éclosion des œufs traités de 36 % avec 5 g.l-1 à 58 % avec 20 g.l-1. Selon la concentration considérée, les saponines et les flavonoïdes provoquent respectivement en moyenne la mortalité corrigée de 16 % à 30 % et de 12 % à 13 % des embryons traités.
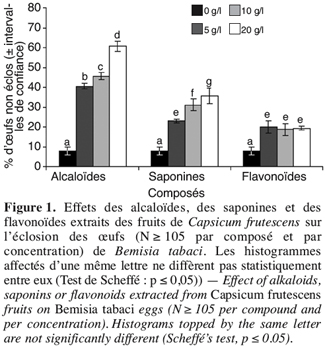
19Par ailleurs, avec les alcaloïdes et les saponines, il existe un effet concentration dépendant, les pourcentages des œufs non éclos augmentent linéairement au fur et à mesure que la concentration s’accroît. Pour les flavonoïdes, la relation entre le pourcentage des œufs non éclos et les concentrations testées n’est pas significativement corrélée. Au regard de la pente de la droite de régression, ce sont toujours les alcaloïdes qui possèdent le pouvoir toxique le plus fort vis-à-vis des œufs de B. tabaci ; viennent ensuite les saponines ; les flavonoïdes se sont montrés très peu toxiques (Figure 2).
3.3. Effets des alcaloïdes, saponines ou flavonoïdes sur les adultes
20La réponse des adultes de B. tabaci varie entre les composés, et avec un même composé suivant la concentration testée. Comme dans le cas des œufs, ce sont les alcaloïdes qui provoquent la mortalité la plus élevée, les saponines et les flavonoïdes affectent relativement moins la survie des adultes de B. tabaci.
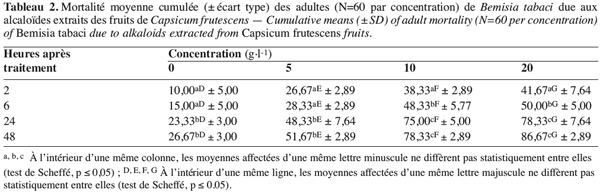
21Effet des alcaloïdes. Quand les adultes de B. tabaci sont traités aux alcaloïdes, leurs réponses varient selon la concentration testée (F = 108,44 ; dl = 3 ; 32 ; p≤ 7,57E-17), la durée d’exposition (F = 199,97 ; dl = 3 ; 32 ; p≤ 8,43E-21), avec une interaction entre ces deux paramètres (F = 3,91 ; dl = 9 ; 32 ; p≤ 0,002). Comme le montre le tableau 2, les alcaloïdes provoquent une mortalité des adultes de B. tabaci significativement plus élevée que celle obtenue avec les adultes libérés sur des plants traités à l’éthanol à 1 %. Le nombre de morts augmente ensuite avec la concentration appliquée, il passe d’environ 12 à 42 %, de 29 à 73 % et de 28 à 86 % des adultes traités respectivement avec 5, 10 et 20 g.l-1. Il faut aussi noter qu’il y a une forte variabilité individuelle quant à la réponse aux concentrations appliquées et à la durée d’exposition à celles-ci, les coefficients de variation varient de 12 à 37 %, de 4 à 11 % et de 4 à 31 % respectivement pour 5, 10 et 20 g.l-1.
22Par ailleurs, pour chaque concentration testée, la mortalité des adultes de l’insecte augmente linéairement avec la durée de l’exposition (Figure 3). Le TL50 est estimé à 77, 17 et 13 heures respectivement pour 5, 10 et 20 g.l-1, il diminue au fur et mesure que la concentration testée augmente.
23Effet des saponines. Comme dans le cas des alcaloïdes, la mortalité des adultes de B. tabaci due aux saponines varie avec la concentration appliquée (F = 62,89 ; dl = 3 ; 32 ; p≤ 1,64E-13) et la durée d’exposition (F = 93,78 ; dl = 3 ; 32 ; p≤ 6,15E-16), mais sans interaction entre ces deux paramètres (F = 0,64 ; dl = 9 ; 32 ; p≤ 0,75). Les composés saponiques affectent plus les adultes de la mouche blanche que le témoin (Tableau 3). Ici aussi, la proportion des adultes morts s’amplifie avec les concentrations étudiées ; celle-ci varie d’environ 5 à 28 % avec 5 g.l-1, de 17 à 37 % avec 10 g.l-1 et de 21 à 48 % avec 20 g.l-1.Vis-à-vis des saponines, les adultes de B. tabaci répondent d’une manière très hétérogène selon les concentrations et le temps de séjour avec ces composés, les coefficients de variation varient de 31 à 56 %, de 8 à 30 % et de 5 à 15 % respectivement pour 5, 10 et 20 g.l-1.
24Par contre, la mortalité des adultes de l’insecte est chronologiquement moins dépendante (Figure 4) ; la mortalité due aux saponines n’atteint pas 50 % des individus testés, le Tl50 n’a pas pu être estimé.
25Effet des flavonoïdes. Quand les adultes de B. tabaci sont mis en contact avec les plants de tomate traités aux flavonoïdes, leurs réponses varient selon la concentration testée (F = 50,48 ; dl = 3 ; 32 ; p≤ 3,10E-12) et la durée d’exposition (F = 28,52 ; dl = 3 ; 32 ; p≤ 3,62E-9) mais sans interaction entre ces deux paramètres (F = 0,30 ; dl = 9 ; 32 ; p≤ 0,969).
26Comme il ressort du tableau 4, les flavonoïdes provoquent une mortalité des adultes de B. tabaci significativement plus élevée que celle obtenue avec les adultes libérés sur des plants traités seulement à l’éthanol. En revanche, le nombre de morts reste généralement comparable entre les concentrations testées, il passe d’environ 0 à 19 %, de 6 à 19 % et de 11 à 29 % des adultes traités respectivement avec 5, 10 et 20 g.l-1. En fait, il y a une grande variabilité individuelle vis-à-vis des concentrations utilisées aux différentes durées d’exposition, les coefficients de variation s’échelonnent de 37 à 90 %, de 10 à 149 % et de 18 à 32 % respectivement pour 5, 10 et 20 g.l-1.
27Pour chaque concentration testée, le traitement aux flavonoïdes n’accroît pas la mortalité des adultes de B. tabaci au fil du temps, la relation entre la mortalité des adultes et la durée de l’exposition est très faible (Figure 5). Là aussi, moins de 30 % des adultes meurent en contact avec les flavonoïdes, le TL50 ne peut pas être déterminé.
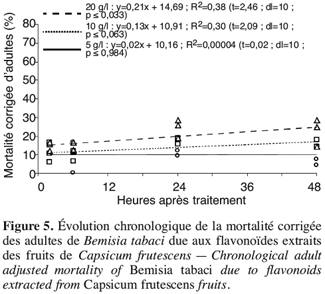
3.4. Analyse comparative de l’effet des alcaloïdes, saponines et flavonoïdes sur les œufs et les adultes de B. tabaci
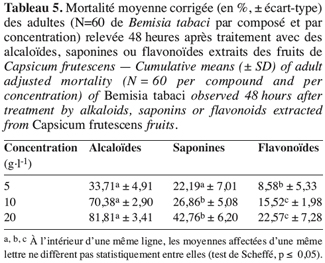
28Au terme de l’expérimentation, 48 heures après application des composés testés à différentes concentrations sur les adultes, ce sont donc les alcaloïdes qui se sont avérés les plus toxiques et ce, quelle que soit la concentration considérée ; les saponines le sont relativement moins, alors que les flavonoïdes se sont montrés très peu nocifs aux adultes de B. tabaci (Tableau 5).
29Par ailleurs, en comparant la réponse des œufs et des adultes aux composés testés dans cette étude, les deux stades de développement de B. tabaci répliquent dans le même sens, les pourcentages des œufs et des adultes morts sont positivement corrélés (r = 0,95 ; r8 ; 0,001 = 0,52) ; mais les œufs sont relativement moins vulnérables que les adultes.
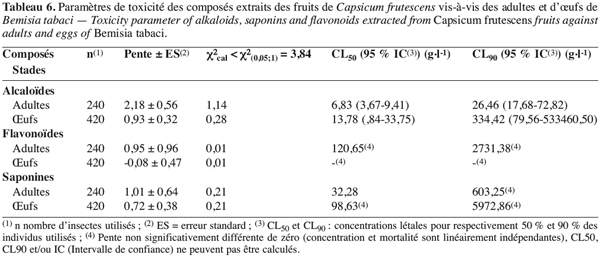
30A court terme, la toxicité des alcaloïdes, des saponines ou des flavonoïdes varie selon le composé et le stade de B. tabaci (Tableau 6). En regard des pentes des droites, des CL50 et CL90, les alcaloïdes apparaissent plus toxiques que les deux autres composés testés. En outre, traités séparément aux alcaloïdes, aux saponines ou aux flavonoïdes, les œufs requièrent plus de deux fois la concentration que les adultes pour affecter 50 % des populations embryonnaires, les adultes se sont montrés plus vulnérables que les œufs. Les deux stades n’ont pas manifesté de concentration dépendance quand ils sont traités aux flavonoïdes (Tableau 6).
4. Discussion
4.1. Teneur en composés
31Les teneurs en composés testés varient selon le produit considéré, les alcaloïdes paraissent plus abondants dans les fruits secs de C. frutescens que les saponines ou les flavonoïdes. Les teneurs trouvées dans notre travail s’insèrent dans l’intervalle de celles rapportées dans la littérature. Ainsi par exemple, des teneurs en alcaloïdes variant de 0,2 à 5,7 % du poids ont été trouvées dans les fruits de C. frutescens par Mathur et al., 2000 et de 0 à 5779 ppm (= 0-0,578 g/100 g) du poids frais par Jarret et al., 2002. Dans les fruits secs de C. annuum, espèce voisine de C. frutescens, les teneurs en principaux alcaloïdes varient de 4,33 à 4,79 g/100 g du poids sec (Contreras-Padilla, Yahia, 1998). Ce sont surtout la capsaicine et la dihydrocapsaicine qui constituent les fractions alcaloïdiques dans les fruits de Capsicum (op. cit.).
32Concernant les saponines associées aux fruits de Capsicum, elles sont concentrées principalement dans les graines ; elles se présentent sous forme d’oligoglycosides (= Capsicosides E1, F2, G5), de stéroïdes (CAY-1) ou de benzaldéhyde, chitosane et 2-deoxy-D-glucose ; ils ont été isolés respectivement par Iorizzi et al. (2002), De Lucca et al. (2002) avec Renault et al. (2003), et par (Duke et al., 2003), ils ont tous une activité anti fongique et anti microbienne. A défaut des ordres de grandeurs de ces composés dosés dans les fruits de Capsicum disponibles dans la littérature, des teneurs avoisinant 5 % du poids des arilles ont été trouvées dans les graines de Swartizia langsdorffii (Magalhaes et al., 2003) ; elles sont beaucoup plus élevées que celles trouvées dans les fruits de Capsicum, objet de notre travail.
33Les flavonoïdes extraites des fruits de Capsicum sont constitués principalement de quercétine et de lutéoline (Lee et al., 1995 ; Materska et al., 2003). Les teneurs en ces composés varient selon les variétés. Elles oscillent de 0,072 à 0,097 g/100 g du poids sec des fruits (Perucka, Materska, 2003) et avoisine 0,004 g/100 g dans les fruits murs de C. frutescens (Howard et al., 2000), elles sont relativement inférieures à celles extraites des fruits de C. frutescens étudiés dans le présent travail.
34En fait, en accord avec Zang, Hamauzu (2003), la teneur en composés phytochimiques dans les fruits murs de Capsicum, comme d’ailleurs dans d’autres espèces de plantes, peut être affectée par le génotype, les conditions du développement et de croissance, la maturité, le conditionnement, les conditions de stockage et par les méthodes d’extraction.
4.2. Effets des alcaloïdes, saponines et flavonoïdes sur les œufs et les adultes de B. tabaci
35Dans les conditions d’étude, les alcaloïdes, les saponines et les flavonoïdes, extraits des fruits de C. frutescens, affectent la viabilité des œufs et la survie des adultes de B. tabaci. Comme cela a été observé chez Trialeurodes vaporariorum (Choi et al., 2003), les adultes se sont montrés plus sensibles que les œufs. Les alcaloïdes se sont avérés plus efficaces que les saponines et les flavonoïdes.
36Les alcaloïdes peuvent avoir des effets toxiques par contact chez les embryons, par contact et/ou par ingestion chez les adultes de B. tabaci. L’effet insecticide du fruit de C. frutescens a aussi été observé chez C. maculatus (Gakuru, Foua, 1996). Vis-à-vis des adultes, il n’est pas non plus impossible que les insectes soient morts par inanition consécutive à des effets répulsifs ou anti-appétant comme dans le cas d’autres invertébrés (El-Lakwah et al., 1997 ; Tang et al., 2000 ; Trematerra, Sciarretta, 2002).
37Le faible effet des saponines chez B. tabaci, notamment sur les œufs, peut être lié à la masse moléculaire élevée empêchant ces composés de traverser le chorion de l’œuf ; comme le suggèrent Magalhaes et al. (2003) dans le cas de Biomphalaria glabrata traité par des saponines extraites des graines de Swartizia langsdorffii. L’effet des composés saponiques semble être toxique ; ainsi appliquées contre Callosobruchus chinensis, les saponines inhibent le développement de l’insecte (Applebaum et al., 1969). L’incorporation des saponines extraites des racines et des feuilles de la luzerne à 0,5 ou 1,6 mg/g du poids frais dans le milieu d’élevage de Ostrinia nubilalis, affecte aussi significativement la croissance et le développement des chenilles de l’insecte (Nozzolillo et al., 1997).
38Les flavonoïdes, testés dans ce travail, se sont montrés sans ou à faibles effets sur l’éclosion des œufs et la survie des adultes de B. tabaci, ce résultat rappelle celui observé chez Manduca sexta avec la rutine (Barbosa et al., 1991). L’effet de ces composés peut être anti-appétant ou/et toxique, comme par exemple chez les chenilles de Utetheisa ornatrix (Gronquit et al., 2001). In vitro, certains flavonoïdes inhibent la réductase d’aldose, la CI50 a été estimée à 0,0005 mg/ml dans le cas de la quercitrine (Lee, 2002).
39Dans le cas des adultes non virulifères, les alcaloïdes à 20 g/l peuvent réduire le niveau des populations de B. tabaci à un seuil tolérable en culture sous serre ; mais, le TL50, déterminé dans ce travail pour les composés efficaces à fortes concentrations, ne semble pas suffisamment court pour tuer les adultes virulifères avant de transmettre le TYLC aux plantes saines ; en effet, le temps minimum requis pour l’inoculation du virus avoisine 40 min (Rubinstein et al., 1999). Les alclaoïdes apparaissent donc comme potentiellement utilisables en gestion intégrée des ravageurs.
5. Conclusions
40Les concentrations utilisées dans cette étude sont nettement supérieures à celles trouvées naturellement dans les fruits (des essais préliminaires conduits avec des concentrations égales aux teneurs naturelles n’ont pas donné de résultats significativement différents des lots témoins). Les résultats obtenus montrent que les alcaloïdes sont plus actifs que les saponines et les flavonoïdes sur B. tabaci. A court et moyen termes, les alcaloïdes, à 20 g.l-1, peuvent être intégrés dans un programme de gestion de B. tabaci en culture sous serre, surtout en situation à risque de résistance aux autres produits. Reste cependant à évaluer leurs éventuels effets sur les auxiliaires naturels (ennemis et pollinisateurs) associés à la culture protégée, et ce, en dépit de l’innocuité de la capsaicine vis-à-vis de certains parasitoïdes, par exemple Encarsia citrina (Rebek, Sadof, 2003). En effet, les produits biologiques les plus intéressants, utilisés en protection des plantes, sont ceux qui ont un impact minimal sur l’ensemble des composantes de l’agro-écosystème sauf pour les ravageurs ciblés (NAP, 1996). L’idéal serait d’exploiter les propriétés anti appétant non toxiques des composés testés, comme le suggèrent Akhtar et Isman (2004).
41Par ailleurs, sachant que les extraits de plantes perdent leur activité biologique sous les radiations solaires (Scott et al., 2003), les modes d’action, les modalités d’application et l’impact des facteurs physiques sur la dégradation des composés botaniques testés doivent aussi être étudiés.
Bibliographie
Abbott W. (1925). A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18, p. 265–267.
Abdel Salam OMB., Moszik O., Szolcsanyi J. (1995). Studies on the effect of intragastric capsaicin on gastric ulcer and on the prostacyclin-induced cytoprotection in rats. Pharmacol Res. 32 (4), p. 209–215.
Akhtar Y., Isman MB. (2004). Comparative growth inhibitory and antifeedant effects of plant extracts and pure allelochemicals on four phytophagous insect species. J. Appl. Entomol. 128, p. 32–38.
Applebaum SW., Marco S., Birk Y. (1969). Saponins as possible factors of resistance of legume seeds to the attack of insects. J. Agr. Food Chem. 17 (3), p. 618–622.
Barbosa P., Gross P., Kemper J. (1991). Influence of plant allelochemicals on tobacco hornworm and its parasitoid, Cotesia congregate. Ecology 72 (5), p. 1567–1575.
Bathee JA., Paine TD., Nuessly GS. (1991). Comparative biology, morphometrics and development of two populations of Bemisia tabaci (Homoptera: Aleyrodidae) on cotton and poinsettia. Ann. Entomol. Soc. Am. 84 (4), p. 407–411.
Bouchelta A., Blenzar A., Beavougui AJP., Lakhlifi T. (2003). Etude de l’activité insecticide des extraits de Capsicum frutescens (Solanacées) sur Bemisia tabaci (Gennadius) (Homoptera : Aleyrodidae). Rev. Méd. Pharm. Afr. 17, p. 19–28.
Brown JK. (1994). Current status of Bemisia tabaci as a plant pest virus vector in agro-ecosystems worldwide. FAO Plant Prot. Bull. 42 (1–2), p. 3–32.
Butler GD. Jr., Coudriet DL., Henneberry TJ. (1988). Toxicity and repellency of soybean and cotton seed oil to the sweetpotato whitefly and the cotton aphid on cotton in greenhouse studies. Southwest. Entomol. 13, p. 81–86.
Chichewicz RH., Thorpe PA. (1996). The antimicrobial properties of chile peppers (Capsicum species) and their uses in Mayan medecine. J. Ethnopharm. 52, p. 61–70.
Choi WI., Lee EH., Choi BR., Park HM., Ahn YJ. (2003). Toxicity of plant essential oils to Trialeurodes vaporariorum (Homoptera: Aleyrodidae). J. Econ. Entomol. 96 (5), p. 1479–1484.
Cordell GA., Araujo OE. (1993). Capsaicin: identification, nomenclature and pharmacotherapy. Ann. Pharmacother. 27, p. 383–395.
Contreras-Padilla M., Yahia EM. (1998). Changes in capsaicinoids during development, maturation, and senescence of chile peppers and relation with peroxidase activity. J. Agric. Food Chem. 46, p. 2075–2079.
Coudriet LD., Prabhaker N., Meyerdik DE. (1985). Sweetpotato whitefly (Homoptera: Aleyrodidae): effects of neem seed extract on oviposition and immature stages. Environ. Entomol. 14 (6), p. 77–83.
Deall CC., Schnitzer TJ., Lipstein E., Seibold JR., Stevens RM., Levy MD., Albert D., Renold F. (1991). Treatment of arthritis with topical capsaicin: A double-blind trial. Clin. Ther. 13 (3), p. 383–395.
De Lucca AJ., Bland JM., Vigo CB., Cushion M., Selitrennikoff CP., Peter J., Walsh TJ., (2002). CAY –I. a fungicidal saponin from Capsicum sp. fruit. Med. Mycol. 40 (2), p. 131–137.
de Souza AP., Vendramim JD. (2000). Efeito de axtratos aquosos de Meliaceas sobre Bemisia tabaci biotipo B em tomateiro. Bragantia 59 (2), p. 173–179.
Duke SO., Baerson SR., Dayan FE., Rimando AM., Scheffler BE., Tellez MR., Wedge DE., Schrader KK., Akey DH., Arthur FH., De Lucca AJ., Gibson DM., Harrison HF. Jr, Peterson JK., Gealy DR., Tworkoski T., Wilson CL., Morris JB. (2003). Pest Manage. Sci. 59 (6-7), p. 708–817.
EL-Lakwah F., Khaled OM., Kattab MM., Abdel-Rahman TA. (1997). Effectiveness of some plant extracts and powders against the lesser grain borer Ryzopertha dominica (F.). Ann. Agric. Sci. 35 (1), p. 567–578.
Finney DJ. (1971). Probit analysis. 3th ed. Cambridge University Press. IBSN 0-521080421 X, 333 p.
Gakuru S., Foua BK. (1996). Effects of plant extracts on the cowpea weevil (Callosobruchus maculatus Fab.) and the rice weevil (Sitophilus oryzae L.). Cah. Agric. 5 (1), p. 39–42.
Gomez P., Cubillo D., Mora GA., Hilje L. (1997). Evalucion de posiblies repelentes de Bemisia tabaci. II. Extractos vegetales. Manejo Integrado de Plagas (Costa Rica) 29, p. 17–25.
Gronquist M., Bezzerides A., Attygalle A., Meinwald J., Eisner M., Eisner T. (2001). Attractive and defensive functions of the ultraviolet pigments of a flower (Hypericum calycinum). PNAS 98, p. 13745–13750.
Harborne JB. (1998). Phytochemical methods: a guide to modern techniques of plant analysis. (3rd ed.) London: Chapman & Hall. ISBN: 0-412-57270-2, 302 p.
Howard LR., Talcott ST., Brenes CH., Villalon B. (2000). Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum Sp) as influenced by maturity. J. Agric. Food Chem. 48, p. 1713–1720.
Iorizzi M., Lanzotti V., De Marino GRS., Zollo F. (2002). Antimicrobial furostanol saponins from the seeds of Capsicum annuum L. var. acuminatum. J. Agric. Food Chem. 50 (15), p. 4310–4316.
Jabbour F., Abaha L., Kaak H., Moryane R. (2001). La maladie des feuilles jaunes en cuillère de la tomate (TYLCV) au Maroc. Proceedings AMPP. Rabat, Maroc, p. 41–46.
Jarret RL., Perkins B., Fan T., Prince A., Guthrie K., Skoczenski B. (2002). Screening Capsicum sp. for capsaicinoids using EIA. Proceedings of the 16th International Pepper Conference Tampico, Tamaulipas, Mexico. November 10–12. (Abstract).
Jenks CW. (2002). Extraction studies of Thernanthe iboga and Voacanga africana. Nat. Prod. Lett. 16 (1), p. 71–76.
Lee HS. (2002). Inhibitory activity of Cinnamomum cassia bark-derived component against rat lens aldose reductase. J. Parm. Pharmaceut. Sci. 5 (3), p. 226–230.
Lee Y., Howard LR., Villalon B. (1995). Flavonoids and antioxidant activity of fresh pepper (Capsicum annuum) cultivars. J. Food Sci. 60 (3), p. 473–476.
Liu TX., Stansly PA., Couner JM. (1996). Toxicological effects of selected insecticide to Nephaspis occulatus (Balatchley) (Col. Coccinellidae), a predator of Bemisia argentifolii (Homoptera : Aleyrodidae). J. Appl. Ent. 120, p. 369–373.
Larew HG., Locke JC. (1990). Repellency and toxicity of horticultural oil against whitefly on Chrysanthemum. HortScience 25 (11), p. 1406–1407.
Lopez-Carrillo L., Avila M., Dubrow R. (1994). Chilli pepper consumption and gastric cancer in Mexico: A case-control study. Am. J. Epidem. 139 (3), p. 263–271.
Magalhaes AF., Goulant AM., Tozzi de A., Santos CC., Serrano DR., Zanotti-Magalhaes EM., Magalhaes EG., Magalhaes AL. (2003). Saponins from Swartizia langsdorffii : Biological activities. Mem. Inst. Oswaldo Cruz, Rio de Janeiro 98 (5), p. 713–718.
Materska M., Piacenteb S., Stochmalc A., Pizzab C., Oleszekc W., Peruckaa I. (2003). Isolation and structure elucidation of flavonoid and phenolic acid glycosides from pericarp of hot pepper fruit Capsicum annuum L. Phytochemistry 63 (8), p. 893–898.
Mathur R., Dangi RS., Dass SC., Malhotra RC. (2000). The hottest chilli variety in India. Curr. Sci. 79 (3), p. 287–288.
Morallo-Rejesus B. (1987). Botanical pest control research in the Philippines. Philipp. Entomol. 7, p. 1–30.
NAP (1996). Ecologically based pest management – New solution for new century. Board on agriculture. National Research Council. ISBN 0-309-05330-7. NAP Washington D.C. 160 p.
Naranjo SE., Ellsworth PC. (2001). Challenges and opportunities for pest management of Bemisia tabaci in the new century. Crop Prot. 20, p. 77.
Nardo EAD., De Costa AS., Lorencao AL. (1997). Melia azadirach extract as an antifeedent to Bemisia tabaci (Homoptera : Aleyrodidae). Florida Entomol. 80 (1), p. 92–94.
Newall CA., Anderson LA., Philipson JD. (1996). Herbal Medicines: a Guide for heath care professionals. London, England: The pharmaceutical Press. p. 44.
Nostro A., Germano MP., D’Angelo V., Marino A., Cannatelli MA. Extraction methods and bioautography for evaluation of medicinal plant antimicrobial activity.
Nozzolillo C., Arnason JT., Campos F., Donskov N., Jurzysta M. (1997). Alfalfa leaf saponins and insect resistance. J. Chem. Ecol. 23 (4), p. 995–1002.
Ofuya TL. (1986). Use of word ash, dry chilli pepper fruits and onion scale leaves for reducing Callosobruchus maculates (Fabricius) damage in cowpea seeds during storage. J. Agr. Sci. 107 (2), p. 467–468.
Onu I., Aliyu M. (1995). Evaluation of powdered fruits of four peppers (Capsicum spp.) for the control of Callosobruchus maculatus (F.) on stored cowpea seed. Int. J. Pest Manag. 41 (3), p. 143–145.
Palumbo JC., Horowitz AR., Prabhaker N. (2001). Insecticidal control and resistance management for Bemisia tabaci. Crop Prot. 20, p. 739–765.
Perucka I., Materska M. (2003). Antioxidant activity and content of capsaicinoids isolated from paprika fruits. Pol. J. Food Nutr. Sci. 12/53 (2), p. 15–18.
Rebek EJ., Sadof CS. (2003). Effects of pesticide applications on the Euonymus Scale (Homoptera: Diaspididae) and its parasitoid, Encarsia citrina (Hymenoptera: Aphelinidae). J. Econ. Entomol. 96 (2), p. 446–452.
Renault S., De Lucca AJ., Boue S., Bland JM., Vigo CB., Selitrennikoff CP. (2003). CAY-1, a novel antifungal compound from cayenne pepper. Med. Mycol. 41 (1), p. 75–81.
Rubinstein G., Morin S., Czosnek H. (1999). Transmission of tomato yellow leaf curl geminivirus to imidacloprid treated tomato plants by the whitefly Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 92 (3), p. 658–662.
Saber MS. (1976). Antimicrobial substance in certain members of Solanaceae. IV. Detection of active principle in pepper. Zentralbl. Bakteriol. Parasitenkd Infektionkr. Hyg. 131 (2), p. 110–112.
Schulten GGM. (1997). Overview of the whitefly - Virus problem: objectives of the workshop. FAO Plant Prod. Prot. Paper. 143, p. 7–10.
Scott IM., Jensen H., Scott JG., Isman MB., Arnason JT., Philogène1 BJR. (2003). Botanical insecticides for controlling agricultural pests: Piperamides and the Colorado potato beetle Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae). Arch. Insect Bioch. Physio. 54, p. 212–225.
Stansly PA., Liu TX. (1997). Selectivity of insecticides to Encarsia pergandiella (Hymenoptera: Aphelinidae), an endoparasite of Bemisia argentifolii (Hemoptera: Aleyrodidae). Bull. Entomol. Res. 87, p. 525–531.
Tang D., Wang C., Luo L., Qin J. (2000). Comparative study on the responses of maxillary sensilla styloconica of cotton bollworm Helicoverpa armigera and oriental tobacco budworm H. assulta larvae to phytochemicals. Science China (Series C) 43 (6), p. 606–612.
Therry V. (1994). Capsaïcine en dermatologie. Ann. Pharm. 45 (2), p. 20–23.
Tolan I., Ragoobirsingh D., Morisson EY. (2001). The effect of capsaicin on blood glucose, plasma insulin levels and insulin binding in dog models. Phytother. Res. 15 (5), p. 391–394.
Traboulsi R. (1994). Bemisia tabaci: a report on its pest status with particular reference to the Near East. FAO Plant Prot. Bull. 42 (1-2), p. 33–58.
Trematerra P., Sciarretta A. (2002). Activity of chilli, Capsicum annuum L. var. acuminatum, on stored product insects Oryzaephilus surinamensis (L.), Sitophilus oryzae (L.) and Tribolium castaneum (Herbst). IOBC/wprs Bull. 25 (3), p. 177–182.
Van der Werf HMG. (1997). Evaluer l’impact des pesticides sur l’environnement. Courr. Environ. 31, p. 5–22.
Wang W., Schultz HD., Ma R. (1999). Cardiac sympathetic afferent sensitivity is enhanced in heart failure. Am. J. Physiol. Heart Circ. Physiol. 277 (2), p. H812–H817.
Williams LAD., Mansingh A. (1993). Pesticidal potential of tropical plants - I. Insecticidal activity of leaf extracts of sixty plants. Insect Sci. Applic. 14 (5), p. 697–700.
Zang D., Hamauru Y. (2003). Phenolic compounds, ascorbic acid, carotenoids and antioxidant properties of grebe, red and yellow bell peppers. Food, Agric., Environ. 1 (2), p. 22–27.
Zibokere DS. (1994). Insecticidal potency of red pepper (Capsicum annum) on pulse beetle (Callosobruchus maculatus) infesting cowpea (Vigna unguiculata) seeds during storage. Indian J. Agr. Sci. 64 (10), p. 727–728.
