


- Accueil
- volume 13 (2009)
- numéro 2
- Etude du comportement de Baillonella toxisperma Pierre (moabi) dans les trouées d'abattage enrichies


Visualisation(s): 3114 (68 ULiège)
Téléchargement(s): 158 (6 ULiège)

Etude du comportement de Baillonella toxisperma Pierre (moabi) dans les trouées d'abattage enrichies

Notes de la rédaction
Reçu le 1 septembre 2008, accepté le 4 novembre 2008
Résumé
Le moabi (Baillonella toxisperma Pierre) est une essence commerciale d'Afrique centrale. Bien que les chablis soient généralement considérés comme propices à la régénération de cette essence, le recensement préliminaire effectué dans le cadre de cette étude suggère que l'espèce est peu présente dans les trouées d'abattage provenant d'une exploitation sélective dans le district du Dja dans le sud-est du Cameroun. En effet, un inventaire effectué dans 174 trouées d'abattage n'y a relevé que 12,7 plants.ha-1. Afin d'étudier le comportement de l'espèce dans les trouées d'abattage, 795 graines ont été semées dans 15 trouées et 410 plants issus de pépinière ont été introduits dans 15 autres trouées. Un suivi semestriel pendant 30 mois a mis en évidence des taux de survie élevés de 75,9 % pour les semis et 95,5 % pour les plants. Bien que la croissance en hauteur lors du premier semestre diffère significativement selon les deux modalités d'introduction, après 30 mois de suivi, les accroissements notés sont similaires (167,5 et 160,6 cm pour les semis et les plants respectivement). En moyenne, 6,1 % des plants de B. toxisperma émergent naturellement de la végétation concurrente et connaissent une croissance supérieure aux autres individus. Les facteurs explicatifs des différences de croissance en hauteur des plants sont l'orientation de la trouée (N-S défavorable) et la densité en Macaranga spp. dont l'abondance inhibe l'évolution des tiges de B. toxisperma. En conclusion, le taux de survie élevé de B. toxisperma dans les trouées et ses valeurs de croissance dix fois supérieures à celles observées dans le sous-bois plaident pour la mise en œuvre de techniques sylvicoles valorisant les trouées d'abattage pour des opérations de reboisement.
Abstract
Study of the behavior of Baillonella toxisperma Pierre (moabi) in enriched logging gaps. The moabi (Baillonella toxisperma Pierre) is a commercial tree in central Africa. Although forest gaps are generally considered as favorable to the regeneration of this species, in this study, a preliminary inventory conducted within a selective exploitation located in the Dja district, south-eastern Cameroon, found a low density of 12.7 seedlings.ha-1 within logging gaps. This suggests that the species was in fact poorly represented in logging gaps. In order to further study the behavior of the moabi in logging gaps, 795 seeds were planted in 15 logging gaps and 410 saplings from a tree nursery were introduced in 15 other gaps. A biannual monitoring spanning a period of 30 months showed a 75.9% survival rate for seedlings and 95.5% for saplings. Despite our observation of a significant difference in growth rates between both methods during the first semester, after 30 months of monitoring, seedlings and saplings had reached similar growth rates (167.5 cm and 160.6 cm for seedlings and saplings, respectively). On average, 6.1% of B. toxisperma saplings emerged naturally from the competitive vegetation and had a higher growth rate than other plants species. Growth differences between plants were mainly explained by the geographical orientation of the logging gap (N-S unfavorable) and the density of Macaranga spp. whose abundance inhibits the development of B. toxisperma. In conclusion, both the high survival rate of B. toxisperma in logging gap and its 10 fold higher growth rate in the latter environment when compared to canopy cover suggest the implementation of forestry methods using logging gap during reforesting operations.
Table des matières
1. Introduction
1Le moabi, Baillonella toxisperma Pierre (Sapotaceae), est une espèce hautement symbolique de la forêt dense humide d'Afrique centrale. Parmi les plus grands arbres du continent (Vivien et al., 1985), B. toxisperma est recherchée par les exploitants forestiers pour la qualité de son bois ainsi que par les populations locales pour ses fruits et ses graines, desquelles une huile de qualité peut être extraite. Selon Debroux (1998), cette double convoitise pourrait engendrer une raréfaction de l'espèce à moyen terme. Répartie du sud du Nigeria à la République Démocratique du Congo, on peut en effet estimer que B. toxisperma est exploitée à raison de 100 000 m³ par année (ATIBT, 2006). Les données concernant les récoltes de fruits demeurent par contre beaucoup plus rares (Vermeulen et al., 2005).
2B. toxisperma est une espèce monoïque dont la fructification est abondante en moyenne tous les trois ans, lors de la petite saison sèche (Debroux, 1998). L'espèce fructifie régulièrement à partir de 70 cm de diamètre. Le fruit est une baie globuleuse d'environ 6 cm de diamètre. Les graines ellipsoïdes de 4 x 2 x 2,5 cm présentent un pouvoir germinatif élevé mais décroissant très rapidement (Pangou, 1989 ; Schmidt, 1992 ; Debroux et al., 1998). La dispersion est assurée principalement par l'homme, l'éléphant (Loxodonta africana Cuvier), le chimpanzé (Pan troglodytes Oken), le gorille (Gorilla gorilla Savage) et quelques espèces de rongeurs. La germination est semi-hypogée et les importantes réserves présentes dans les cotylédons permettent une croissance initiale d'environ 40 cm (Debroux, 1998 ; Nimbot Mamba, 2005). B. toxisperma s'installe et survit sous une large gamme de conditions lumineuses. Les semis sont tolérants à l'ombre mais y présentent une croissance très lente. Ainsi, en moyenne 119 ans seraient nécessaires à l'obtention d'individus de 10 cm de diamètre sous couvert (Debroux, 1998). La croissance des semis est également freinée dans les sites très éclairés sous l'effet de l'ombrage créé par les espèces concurrentes plus héliophiles et dont la croissance est plus rapide. Ces caractéristiques classeraient B. toxisperma parmi les espèces qualifiées de struggling gamblers par Oldeman et al. (1991) ou encore parmi les semi-héliophiles (Doucet, 2003). Les trouées et leurs abords pourraient par conséquent constituer un environnement idéal pour l'épanouissement des plantules (Debroux, 1998).
3Le rôle des trouées dans la dynamique des forêts tropicales a été largement documenté (Denslow, 1980 ; Putz, 1983 ; Brokawn, 1985 ; 1987 ; Uhl et al., 1988). Toutefois, peu d'études se sont focalisées sur la régénération des espèces à vocation commerciale dans cet environnement. Selon plusieurs études, les trouées forestières favoriseraient la régénération des essences pionnières au détriment des essences de valeurs (Brokawn, 1985 ; Denslow, 1987 ; Kyereh et al., 1999 ; Dalling et al., 2002 ; Babaasa et al., 2004). Toutefois, Uhl et al. (1988) ont noté très peu de nouvelles colonisations dans les trouées en forêt amazonienne, les espèces tirant profit du regain de lumière étant celles déjà présentes.
4La lumière serait le facteur principal qui affecterait l'installation et la croissance des semis (Denslow et al., 1994). Les conditions lumineuses prévalant au niveau du sol sont étroitement liées à la taille de la trouée, à sa forme, à la hauteur de la canopée adjacente et à la latitude (Canham et al., 1990).
5L'exploitation forestière en Afrique centrale se caractérise par l'abattage sélectif de 0,5 à 2 pieds par hectare (Fargeot et al., 2004). L'ouverture de la canopée qui en résulte pourrait s'apparenter aux chablis naturels avec toutefois une différence notable liée au passage des engins d'exploitation, lesquels détruisent la végétation à proximité de la souche et induisent une certaine compaction du sol.
6Dans cet article, nous avons cherché à évaluer l'intérêt d'une approche sylvicole de B. toxisperma basée sur la prise en compte des trouées forestières artificiellement créées par l'abattage sélectif (Coates et al., 1997). A cette fin, le comportement de plants de B. toxisperma provenant de pépinière ou de semis directs a été étudié dans les trouées d'abattage choisies aléatoirement au sein d'une exploitation forestière de 176 000 ha située dans le district de Dja, au sud-est du Cameroun. Plus spécifiquement, nous avons quantifié le succès de la régénération et les taux de croissance des plants en fonction des conditions locales.
2. Matériel et méthodes
2.1. Site d'étude
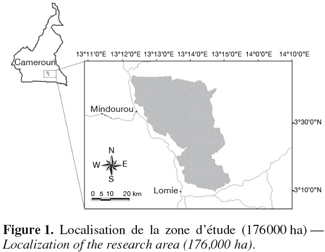
7L'étude s'est déroulée dans la concession forestière attribuée à la société Pallisco, en République du Cameroun, dans la province de l'Est, district du Dja (Mindourou) (Figure 1). La zone d'étude est située entre les latitudes 3°10' et 3°44' Nord et entre les longitudes 13°20' et 13°52' Est, sur une superficie de 176 000 ha.
8Les précipitations s'élèvent en moyenne à 1 650 mm par an et sont réparties en quatre périodes : une grande saison des pluies (d'août à novembre), une saison sèche très marquée (de décembre à février), une petite saison des pluies (de mars à juin) et une petite saison sèche (juillet) dont l'ampleur est très variable d'une année à l'autre. La température moyenne est de 24 °C.
9Le relief est doux et assez uniforme. Les dépressions sont faibles mais bien marquées. Le réseau hydrographique dessine un maillage régulier de petites vallées à fond plat et marécageux. L'altitude varie de 600 à 760 m. La plus grande partie de la zone d'étude repose sur le complexe de base du précambrien inférieur dont les principales roches sont les micaschistes gris sombres argentés. Du point de vue pédologique, les sols rencontrés dérivent de roches métamorphiques. Sur la terre ferme, ce sont principalement des sols ferrallitiques rouges ou jaunes typiques. Ils sont en général profonds, argileux, meubles, perméables et présentent souvent peu d'humus. Leur teneur en bases échangeables est faible. Ce sont des sols pauvres, acides et fragiles.
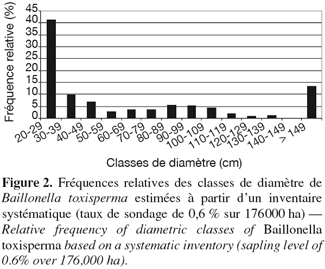
10La forêt est une forêt de transition entre les types sempervirents et semi-sempervirents. Elle se caractérise par la rareté des Caesalpiniaceae, et la présence de nombreuses Euphorbiaceae et Olacaceae. Parmi les arbres de grande taille, les Meliaceae ainsi que diverses Sterculiaceae et Ulmaceae sont bien représentées (Letouzey, 1968). B. toxisperma y est présent à raison de 0,1 pied.ha-1 (diamètre à hauteur de poitrine, dhp ≥ 20 cm). La structure de population obtenue après un inventaire systématique réalisé au taux de 0,6 % sur les 176 000 ha est présentée par la figure 2 (Hubert et al., 2003). Les pieds de diamètre compris entre 20 et 29 cm totalisent plus de 40 % des observations. Les individus matures (dhp ≥ 70 cm) sont présents à raison de 0,03 pied.ha-1.
2.2. Dispositif d'étude
11Inventaire de la régénération naturelle de B. toxisperma dans les trouées d'abattage. Afin d'évaluer la présence de B. toxisperma dans les trouées d'abattage, 174 trouées âgées de 6 à 9 mois ont été choisies de façon aléatoire au sein d'une assiette annuelle de coupe de 2 500 ha. L'espèce abattue et son diamètre ont systématiquement été notés. L'orientation de la trouée a été mesurée en considérant l'azimut de son axe principal. Les trouées ont ensuite été classées en quatre catégories : N-S (nord-sud), SE-NO (sud est-nord ouest), E-O (est-ouest), NE-SO (nord est-sud ouest). Le contour des trouées a été déterminé à partir de la projection des cimes des arbres de dhp ≥ 10 cm. La superficie a été calculée par la juxtaposition d'une grille de 2 x 2 m matérialisée sur le terrain par des jalons. Elle a été scindée en deux parties : la partie proximale et la partie distale (Oldeman, 1978). La partie proximale est la zone où se trouvait le tronc de l'arbre abattu et débardé par les engins d'exploitation. La partie distale correspond à l'emplacement délimité par la cime de l'arbre gisant au sol. Considérant l'amas de branches recouvrant la partie distale et inhibant toute régénération avant décomposition, seule la partie proximale a été inventoriée. La hauteur des plants de B. toxisperma rencontrés a été mesurée en cm.
12Comportement de B. toxisperma dans les trouées d'abattage. Trente trouées ont été choisies aléatoirement parmi les 174 précédentes. Des graines de B. toxisperma (N = 795) ont été semées dans 15 d'entre elles selon un quadrillage de 2 x 1 m établi dans la partie proximale. Dans les 15 autres trouées, des plants de B. toxisperma (N = 410) éduqués pendant 7 mois en pépinière ont été introduits tous les 2 x 4 m. La hauteur moyenne des plants était de 73 cm (σ = 18,4 cm). Préalablement au semis et à la plantation, la végétation concurrente a été éliminée à la machette. Par la suite, aucun autre dégagement n'a été opéré.
13Deux mois après les enrichissements, le taux de germination a été relevé dans les parcelles comportant les graines ainsi que le taux de reprise dans les parcelles enrichies en plants. Des suivis semestriels de la croissance ont été effectués pendant 30 mois. Lors de ceux-ci, les plants ont été mesurés en hauteur avec un mètre ruban (en cm) et en diamètre (au collet) avec un pied à coulisse (en mm). Le statut des plants a été examiné à 12 et 24 mois, il exprime la position de B. toxisperma par rapport à la végétation voisine. Trois classes ont été prises en compte : dominant (lorsque le plant dépasse la végétation voisine), codominant (lorsque le plant est à la même hauteur que la végétation voisine) et dominé (lorsque le plant est de taille inférieur à la végétation voisine). Les espèces dominantes ont été également identifiées dans un rayon d'un mètre autour de chaque tige de B. toxisperma.
2.3. Analyse des données
14Les valeurs moyennes obtenues ont été comparées à l'aide de test t-Student et d'analyses de la variance. Lorsque les conditions d'application n'étaient pas satisfaites (normalité et égalité des variances) après transformations de variables, des tests non paramétriques ont été utilisés (Kruskal-Wallis ANOVA H-test, Mann-Whitney U-test). La recherche de corrélation entre deux paramètres a été réalisée à l'aide du coefficient de corrélation de Pearson (r).
3. Résultats
3.1. Caractéristiques des trouées d'abattage et régénération naturelle de B. toxisperma
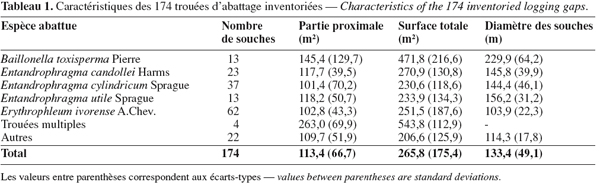
15Onze espèces ont fait l'objet d'une exploitation industrielle sélective à raison de 0,8 pied par hectare (Tableau 1). Quatre trouées parmi les 174 proviennent de l'abattage de deux pieds voisins. La superficie moyenne d'une trouée est de 265,8 m2 (σ = 175,4 m2). Les tailles moyennes des trouées des cinq espèces les plus représentées (B. toxisperma, E. cylindricum, E. candollei, E. utile, E. ivorense) diffèrent significativement (Kruskal-Wallis ANOVA, H = 18,40, p < 0,001). Les diamètres moyens des souches différant significativement selon les espèces (Kruskal-Wallis ANOVA, H = 77,5, p < 0,001) et une corrélation positive entre la taille de la trouée et le diamètre des souches (r = 0,364 ; p < 0,001), contribuent à expliquer les différences observées. La taille moyenne de la partie proximale est de 113,4 m2 (σ = 66,9 m2). Pour les cinq mêmes espèces, les différences dans ce cas ne sont plus significatives (KrusKal-Wallis ANOVA, H = 7,1, p = 0,13).
16Seulement 25 plantules de B. toxisperma ont été inventoriées sur une surface totale de 2,0 ha, soit 12,7 plants.ha-1 de trouée. Leur hauteur moyenne est de 65,9 cm (σ = 20,2 cm). Septante-huit pourcent de ces plantules se situent dans une trouée provoquée par la chute de B. toxisperma.
3.2. Comportement de B. toxisperma dans les trouées d'abattage
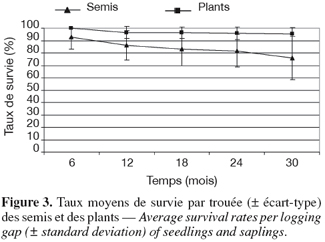
17Le taux de germination a été de 69,4 % (σ = 23,6 %) dans les trouées semées et le taux de reprise des plants introduits a été de 100 %. Après 30 mois de suivi, les taux de survie demeurent élevés, ils sont respectivement de 75,9 % (σ = 7,4 %) et de 95,3 % (σ = 5,4 %) pour les semis et les plants (Figure 3), les différences étant néanmoins significatives entre les deux modalités (t = 5,9 ; dl = 28 ; p < 0,001).
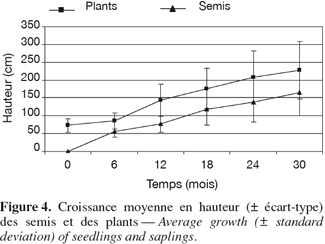
18La croissance en hauteur lors du premier semestre diffère significativement selon les deux modalités d'introduction (test U de Mann-Whitney : U = 5 238 ; Z = 25,1 ; p < 0,001), elle atteint en moyenne 55,5 cm (σ = 16,9 cm) pour les semis et seulement 13,2 cm (σ = 11,5 cm) pour les plants. Au cours des trois semestres suivants, la situation s'équilibre et la croissance moyenne des plants (136,0 cm ; σ = 64,8 cm) devient semblable à celle des semis (138,5 cm ; σ = 56,3 cm) (test U de Mann-Whitney : U = 85 168 ; Z = -0,1 ; p = 0,989). Finalement, après 30 mois de suivi, les accroissements s'avèrent être similaires (test U de Mann-Whitney : U = 79 746 ; Z = -0,9 ; p = 0,380), ils sont respectivement de 167,5 cm (σ = 65,7 cm) pour les semis et de 160,6 cm (σ = 70,6 cm) pour les plants. Au terme de deux années et demi de suivi, les plants conservent néanmoins l'avantage conféré par leur hauteur initiale (test U de Mann-Whitney : U = 47 582 ; Z = 11,63 ; p < 0,001) en atteignant en moyenne 229,3 cm (σ = 80,0 cm) de hauteur contre 167,5 cm (σ = 65,7 cm) pour les semis, soit 61,8 cm qui est proche de la hauteur moyenne des plants lors de leur introduction (73,0 cm) (Figure 4). Les diamètres des plants (16,8 mm ; σ = 5,3 mm) sont également supérieurs à ceux des semis (12,5 mm ; σ = 3,7 mm) (t = 4,93 ; p < 0,001).
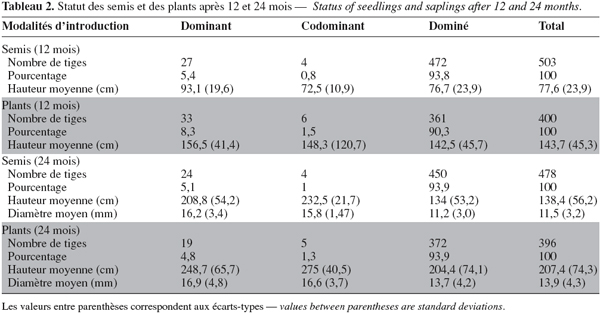
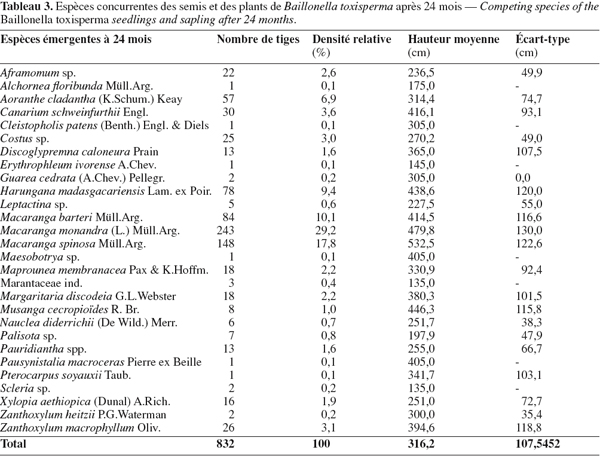
19Les plants de B. toxisperma sont majoritairement dominés par d'autres espèces végétales (Tableau 2). En effet, à 12 mois, 90,3 % des tiges sont dominées dans les plantations contre 93,8 % dans les semis. Ce pourcentage se stabilise à 93,9 % après 24 mois, quelle que soit la méthode d'introduction. La plupart des espèces concurrentes de B. toxisperma sont des espèces ligneuses héliophiles peu longévives (par exemple : Macaranga spp., Musanga cecropioides, Harungana madagascariensis) dont la croissance est exceptionnelle et comprise entre 2 et 2,5 m par an (Tableau 3).
20Après deux ans, tant les plants dominants, codominants que dominés issus de plantations sont plus grands que ceux provenant de semis (test U de Mann-Whitney : U = 141, p < 0,05 ; U = 37 256, p < 0,001).
3.3. Influences des caractéristiques des trouées
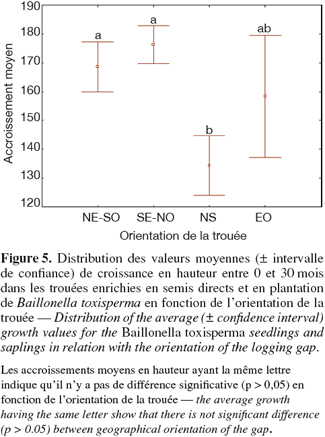
21Les croissances moyennes en hauteur des plants et des semis sont significativement différentes selon les trouées (Kruskal-Wallis : H = 246,95 ; p < 0,001). Afin d'expliquer les différences de croissance entre les trouées, des comparaisons ont été réalisées selon les facteurs suivants : la taille de la trouée, l'orientation de la trouée et la densité en Macaranga spp., principales espèces concurrentes de B. toxisperma (Tableau 4). Parmi ces trois paramètres, la taille de la trouée montre une corrélation positive avec la croissance en hauteur des plants et des semis, mais le test n'est que marginalement significatif (r = 0,34 ; p = 0,07). Les différences de performance de B. toxisperma constatées entre trouées sont liées à l'orientation de la trouée (Kruskal-Wallis : H = 38,96 ; p < 0,001) et à la densité en Macaranga spp. dont l'abondance dans les trouées inhibe l'évolution des tiges (r = - 0,56 ; p < 0,001). En ce qui concerne l'orientation de la trouée, la figure 5 montre que les meilleures croissances en hauteur de B. toxisperma ont été enregistrées dans les trouées orientées dans les directions E-0, NE-SO, SE-NO. Les trouées orientées dans ces dernières directions reçoivent en effet plus de lumière que celles situées dans la direction N-S qui sont perpendiculaires à la trajectoire du soleil et ne sont donc illuminées qu'au zénith ou à la mi journée. La disponibilité en lumière dans les trouées d'abattage orientées N-S est donc insuffisante pour assurer une croissance rapide des plants et des semis de B. toxisperma.

4. Discussion et conclusions
22Notre inventaire préliminaire a montré que les plantules de B. toxisperma (moabi) sont peu présentes naturellement dans les trouées d'abattage. En outre, elles se retrouvent principalement dans les trouées où un moabi a été abattu, indiquant une dispersion limitée des semences. Ces résultats sont en accord avec d'autres relevés effectués dans des trouées d'âges divers en forêt gabonaises (Degueret, 2002 ; Gobert, 2002 ; Cobut, 2005). Cette faible densité pourrait peut-être s'expliquer par la relative rareté des pieds adultes et par les dégâts provoqués par l'abattage et le débardage aux éventuelles plantules présentes sous les semenciers (Debroux, 1998 ; Makana et al., 2005). Néanmoins, dans la présente étude, nous avons observé une croissance régulière des plants de B. toxisperma introduits dans les trouées d'abattage, ce qui confirme que cet environnement lui est favorable. Son taux de survie y est élevé et sa croissance en hauteur et diamètre est dix fois supérieure à celle observée dans le sous-bois par Debroux (1998). Notons aussi que les valeurs d'accroissement obtenues pour les individus dominants ou codominants sont proches de celles observées par Koumba Zaou et al. (1998) dans des plantations monospécifiques régulièrement dégagées. Les principaux facteurs sylvicoles explicatifs des différences de croissance de B. toxisperma entre les trouées sont la concurrence engendrée par les espèces pionnières et l'orientation de la trouée. Bien que nos résultats ne mettent pas en évidence un effet significatif de la taille de la trouée, des comparaisons plus précises devraient être réalisées, via par exemple la prise en compte de la hauteur des arbres voisins, afin de mieux quantifier la lumière parvenant aux plants. De même, des facteurs édaphiques, dont la richesse et la compaction du sol résultant du passage des engins d'exploitation, devraient être intégrés dans les analyses car ils pourraient contribuer à expliquer la forte variabilité des taux de croissance entre trouées (Schulze, 2008).
23En conclusion, les différences de croissance constatées entre trouées sont la résultante de plusieurs facteurs combinés dont particulièrement la disponibilité en lumière (Paul et al., 2004). Bien que la hauteur des individus provenant de plantation soit finalement plus élevée que celle des individus issus de semis directs, la croissance de B. toxisperma est similaire dans les deux cas, avec une proportion d'individus dominants ou codominants atteignant 6,1 % après deux ans. Le développement d'une approche sylvicole basée sur la valorisation des trouées d'abattage (Coates et al., 1997) parait en conséquence des plus intéressante. Les trouées d'abattage pourraient être indifféremment enrichies en B. toxisperma par semis directs ou par plants éduqués en pépinière, les deux méthodes donnant des résultats similaires. Toutefois, en vertu du caractère récalcitrant des graines, une intervention par semis ne pourrait être envisagée que sur des courtes périodes. Enfin, l'intérêt d'un ou de plusieurs dégagements des plants devrait être évalué en fonction des contraintes rencontrées par les plants au cours de leur développement, mais également en fonction des couts supplémentaires qu'ils pourraient occasionner.
24Remerciements
25Les auteurs remercient la société Pallisco, plus particulièrement MM. Rougeron, Fétéké, Douaud, Zok, Djopandé, Gassang ; M. Ayol de l'asbl Nature + et MM. D. Monticelli, L. Delahaye, P. Lejeune de la Faculté universitaire des Sciences agronomiques de Gembloux.
Bibliographie
ATIBT, 2006. Gestion forestière durable : adapter, conserver, produire. Formation Aménagiste, trouver les hommes, statistique 2005. Paris : ATIBT.
Babaasa D. et al., 2004. Gaps characteristics and regeneration in Bwindi Impenetrable National Park, Uganda. Afr. J. Ecol., 42, 217-224.
Brokawn N.V.L., 1985. Treefalls, regrowth, and community structure in tropical forests. In: Pickett S.T.A. & White P.S, eds. The ecology of natural disturbance and patch dynamics. London: Academic Press, 53-69.
Brokawn N.V.L., 1987. Gap-phase regeneration of three pioneer tree species in a tropical forest. J. Ecol., 75, 9-19.
Canham C.D. et al., 1990. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can. J. Forest Res., 20, 620-631.
Coates K.D. & Burton P.J., 1997. A gap-based approach for the development of silvicultural systems to address ecosystem management objectives. Forest Ecol. Manage., 99, 339-356.
Cobut P., 2005. Etude de la régénération naturelle dans les chablis d'exploitation en forêt dense humide gabonaise. Mémoire : Faculté universitaire des Sciences agronomiques de Gembloux (Belgique).
Dalling J.W. & Hubbell S.P., 2002. Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species. J. Ecol., 90, 557-568.
Debroux L., 1998. L'aménagement des forêts tropicales fondé sur la gestion des populations d'arbres : l'exemple du moabi (Baillonella toxisperma Pierre) dans la forêt du Dja, Cameroun. Thèse de doctorat : Faculté universitaire des Sciences agronomiques de Gembloux (Belgique).
Debroux L., Delvingt W., Mbolo M. & Amougou A., 1998. Régénération du moabi et du mukulungu au Cameroun, perspective pour l'aménagement. Bois Forêts Trop., 255, 5-17.
Degueret A., 2002. Etude de la régénération des espèces commerciales dans les trouées d'exploitation au Gabon. Mémoire : ENGREF, Montpellier (France).
Denslow J.S., 1980. Patterns of species diversity during succession under different disturbance regimes. Oecologia, 46, 18-31.
Denslow J.S., 1987. Tropical rain forest gaps and tree species diversity. Ann. Rev. Ecol. Syst., 18, 431-451.
Denslow J.S. & Hartshorn G.S., 1994. Tree-fall gap environments and forest dynamic processes. In: McDade L.A., Bawa K.S., Hespenheide H.A. & Hartshorn G.S., eds. La Selva. Ecology and natural history of a neotropical rain forest. Chicago, USA: University Chicago Press, 120-127.
Doucet J.-L., 2003. L'alliance délicate de la gestion forestière et de la biodiversité dans les forêts du centre du Gabon. Thèse de doctorat : Faculté universitaires des Sciences agronomiques de Gembloux (Belgique).
Fargeot C., Forni E. & Nasi R., 2004. Réflexions sur l'aménagement des forêts de production du bassin du Congo. Bois Forêts Trop., 281(3), 19-34.
Gobert A., 2002. Etude de la régénération naturelle de quelques essences commerciales au Gabon. Mémoire : Faculté universitaire des Sciences agronomiques de Gembloux (Belgique).
Hubert D., Fétéké F. & NKolong E., 2003. Plan d'aménagement de l'unité forestière d'aménagement 10 039. Document final. Douala, Cameroun : Pallisco.
Koumba Zaou P., Mapaga D., Nze Nguema & Delporte P., 1998. Croissance de 13 essences de bois d'oeuvre plantées en forêt gabonaise. Bois Forêts Trop., 256, 21-33.
Kyereh B., Swaine M.D. & Tompson J., 1999. Effect of light on the germination of forest trees in Ghana. J. Ecol., 87, 772-783.
Letouzey R., 1968. Etude phytogéographique du Cameroun. Paris : Edition Paul Lechevalier.
Makana J.-R. & Thomas S.C., 2005. Effects of light gaps and litter removal on the seedling performance of six African timber species. Biotropica, 37(2), 227-237.
Nimbot Mamba M., 2005. Etude de la germination, de la croissance et de la morphologie des plantules d'espèces ligneuses des forêts denses humides tropicales de la province Ogoué-Lolo. Mémoire : Ecole Nationale forestière d'Ingénieurs, Salé (Maroc).
Oldeman R.A.A., 1978. Architecture and energy exchange of dicotyledonous trees in the forest. In: Tomlinson P.B. & Zimmerman M.H., eds. Tropical trees as living systems. London; New York, USA: Cambridge University Press.
Oldeman R.A.A. & Van Dijk J., 1991. Diagnosis of the temperament of rain forest tree. In: Gomez-Pompa A., Whitmore T.C. & Hardley M., eds. Rain forest regeneration and management. Paris: MAB, UNESCO, 21-65.
Pangou S.V., 1989. Régénération naturelle sur 8 hectares de la forêt dense du Mayombe Congolais : exemple de 5 espèces ligneuses pionnières, post-pionnières et forestières. Thèse de doctorat : Université Nancy I (France).
Paul J.R., Randle A.M., Chapman C.A. & Chapman L.J., 2004. Arrested succession in logging gaps: is tree seedling growth and survival limiting. Afr. J. Ecol., 42, 245-251.
Putz F.E., 1983. Treefall pits and mounds, buried seeds and the importance of soil disturbance to pioneer trees on Barro Colorado Island, Panama. Ecology, 64, 1069-1074.
Uhl C., Clark K., Dezzon N. & Marguirin P., 1988. Vegetation dynamics in amazonian treefall gaps. Ecology, 69, 98-106.
Schmidt K., 1992. Rapport final du projet « Reboisement Bokoué ». Feldkichen, Germany : Deutsche Forstservice.
Schulze M., 2008. Technical and financial analysis of enrichment planting in logging gaps as a potential component of forest management in the eastern Amazon. Forest Ecol. Manage., 255, 866-879.
Vermeulen C. & Doucet J.-L., 2005. Populations locales et aménagement forestier en Afrique. Calcul du besoin en moabi (Baillonnella toxisperma Pierre) des populations riveraines de l'UFA 10039, Cameroun. Bull. AIGX, 4, 27-29.
Vivien J. & Faure J.J., 1985. Arbres des forêts denses d'Afrique Centrale. Paris : ACCT.
Pour citer cet article
A propos de : Yao Lambert Kouadio
Gembloux Agricultural University – FUSAGx. Laboratoire de Foresterie des Régions tropicales et subtropicales. Unité de Gestion des Ressources forestières et des Milieux naturels. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : kouadio.y@fsagx.ac.be
A propos de : Jean-Louis Doucet
Gembloux Agricultural University – FUSAGx. Laboratoire de Foresterie des Régions tropicales et subtropicales. Unité de Gestion des Ressources forestières et des Milieux naturels. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
