


- Accueil
- Volume 14 (2010)
- numéro 1
- Restauration écologique : contexte, contraintes et indicateurs de suivi


Visualisation(s): 23918 (145 ULiège)
Téléchargement(s): 1800 (15 ULiège)

Restauration écologique : contexte, contraintes et indicateurs de suivi

Notes de la rédaction
Reçu le 15 janvier 2009, accepté le 8 juillet 2009
Résumé
Dans un contexte global de perturbations anthropiques toujours croissantes des écosystèmes naturels et semi-naturels, l'unique conservation de ces écosystèmes ne suffit plus. La restauration écologique peut ainsi se révéler un complément essentiel à la conservation s.l. L'article consiste en une synthèse de la littérature scientifique dédiée aux concepts de la restauration écologique. Nous abordons la notion d'objectifs de la restauration notamment au travers de la référence écologique. Outre la définition d'objectifs clairs de restauration, la référence permet ensuite d'évaluer les efforts de restauration, par le biais d'indicateurs de suivi. Les indicateurs peuvent par ailleurs fournir des informations quant aux attributs de l'écosystème restauré et/ou la trajectoire écologique empruntée. Les éventuelles contraintes rencontrées par les espèces au travers de filtres à la restauration pourront être détectées et des mesures de compensation adéquates pourront être proposées.
Abstract
Ecological restoration: context, constraints, and indicators. In a global context of growing anthropogenic disturbances on natural and semi-natural ecosystems, the only conservation of those ecosystems is not sufficient anymore. The ecological restoration may hence be an essential complement to conservation. The paper consists in a synthesis of scientific literature dedicated to the concepts of ecological restoration. We tackle the notion of restoration objectives, notably through the ecological reference. Besides the delimitation of clear objectives, the reference will enable the evaluation of restoration efforts by the use of indicators. The indicators may also provide information related to the attributes of the restored ecosystem and/or the followed ecological trajectory. Potential constraints faced by species through ecological filters could hence be detected and compensation measures could be proposed.
Table des matières
1. Introduction
1La perturbation d'un écosystème consiste en un (des) évènement(s) altérant – dans le temps et dans l'espace – les relations entre les organismes vivants et leurs habitats (Wali, 1987). Ces perturbations peuvent être d'amplitude et de nature diverses. Bien que nombre d'habitats soient intrinsèquement dynamiques (Burel et al., 1999) et subissent des perturbations naturelles, les actions humaines sont à présent à l'origine de la majeure partie des perturbations. La plupart des écosystèmes et des régions du monde (si pas tous) connaissent de tels phénomènes de perturbation d'origine anthropique (Godron et al., 1983 ; UNEP, 2007). Les activités perturbatrices vont de la déforestation au drainage des zones humides, en passant par l'introduction de polluants chimiques. Les impacts des modifications engendrées par les perturbations anthropiques sur la biodiversité ont été largement documentés. Parmi ceux-ci, la fragmentation et ses conséquences sont considérées comme des atteintes majeures à la biodiversité (Ouborg, 1993 ; Harrisson et al., 1999 ; Eriksson et al., 2001 ; Balmford et al., 2005). La fragmentation est la réduction d'un habitat continu de grande taille en fragments d'habitats plus petits et isolés les uns des autres. À l'échelle du paysage, ce processus se traduit par une réduction de la diversité en habitats ainsi que par une hétérogénéité plus importante des habitats. Par ailleurs, à l'échelle de l'espèce, la fragmentation peut résulter en une réduction de la taille des populations des fragments rémanents et une diminution du taux de colonisation des fragments d'habitat, du fait du manque de connectivité. La vulnérabilité de telles populations face aux modifications naturelles ou anthropiques s'en voit accrue (Sih et al., 2000 ; UNEP, 2007).
2L'importance de ces menaces à la biodiversité n'a toutefois reçu une attention sérieuse de la part des décideurs que depuis les années 1990 (Madgwick et al., 2002). Il est apparu que la seule protection et gestion des fragments rémanents de certains habitats naturels et semi-naturels peut ne pas garantir la conservation à long terme de tous les composants de la biodiversité (Bennett, 1997). Les surfaces protégées ne peuvent en réalité assumer leur fonction que si elles sont suffisamment grandes et connectées et si elles abritent un échantillon relativement complet de la biodiversité à l'échelle régionale (Soulé et al., 2001). Ainsi, outre un besoin de mesures de protection des habitats et des espèces, est né un besoin grandissant de restauration écologique visant à augmenter les surfaces des habitats menacés et créer un réseau écologique assurant la connectivité entre fragments. Ces restaurations font à présent l'objet de nombreux projets, plus ou moins ambitieux et … plus ou moins couteux. Il en résulte que, derrière une volonté de conservation et de préservation des écosystèmes, il existe une demande de la société quant à l'évaluation du succès des restaurations écologiques. Dans cette optique, l'article présente une synthèse bibliographique portant sur les notions générales liées à la restauration écologique d'une part, ainsi que sur les critères justifiant de la réussite d'une restauration et les indicateurs permettant de quantifier cette réussite.
2. Restauration écologique ?
3« La restauration écologique est une action intentionnelle qui initie ou accélère l'auto-réparation d'un écosystème qui a été dégradé, endommagé ou détruit, en respectant sa santé, son intégrité et sa gestion durable » (SER, 2004). Une restauration peut ainsi être passive, lorsque les forces de dégradations sont réduites, permettant aux processus naturels de récupération de diriger la restauration, ou active, lorsque non seulement les forces de dégradations sont réduites ou arrêtées, mais que le cours de la restauration est dirigé par des interventions humaines (Lake, 2001). On parle de restauration écologique indifféremment pour des écosystèmes naturels et des écosystèmes semi-naturels (ou culturels), ces derniers étant des habitats abritant des espèces natives et à colonisation spontanée, mais dépendant d'une méthode de gestion traditionnelle (Westhoff, 1983).
4Une étape cruciale d'un projet de restauration est la sélection d'un écosystème de référence. Celui-ci a été défini par Le Floc'h et al. (1995) comme une approximation de l'état souhaitable, une norme choisie parmi plusieurs états alternatifs possibles et accessibles par une succession d'étapes appelée trajectoire. L'écosystème de référence sera ensuite utilisé pour évaluer les efforts de restauration (White et al., 1997). Deux facteurs sont à intégrer à la notion d'écosystème de référence :
5– La variation naturelle ou anthropique des écosystèmes dans le temps et dans l'espace. Ainsi, par exemple, le contexte spatial (nature de la matrice paysagère, nature des lisières, taille, isolement) des écosystèmes faisant l'objet de restauration est bien souvent altéré par rapport à la situation historique (considérée comme référence dans de nombreux cas). Ces modifications du contexte spatial ont des conséquences sur la composition et les processus des écosystèmes (Turner, 1989), ainsi que sur les possibilités de restauration des sites (Aronson et al., 1996). Si le choix de la référence se porte sur un site, il faudra pouvoir admettre que cette référence comporte simplement des conditions, typiques des sites les moins endommagés de la région de restauration (Lake, 2001) ;
6– Par ailleurs, le paradigme de la variation spatiale régionale, i.e. le fait que la similarité entre deux sites décroît avec leur éloignement respectif du fait de la corrélation spatiale parmi les facteurs biotiques, environnementaux et historiques, suggère qu'aucun site de référence ne constitue un objectif de restauration parfait (Pickett et al., 1994 ; White et al., 1997). Quand la trajectoire désirée est réalisée, l'écosystème manipulé ne requiert plus d'assistance extérieure pour assurer sa santé et son intégrité future (hormis une éventuelle gestion courante dans le cas d'habitats semi-naturels), de plus l'écosystème est résilient ; dans ce cas, la restauration peut-être considérée comme achevée (SER, 2004).
7La restauration écologique se base largement sur la théorie de la succession et la théorie de l'assemblage (“assembly theory”), en considérant la restauration comme initiant ou accélérant l'assemblage d'une série d'espèces. La théorie des assemblages, en étudiant les ensembles de règles gouvernant l'assemblage des espèces, des communautés et des écosystèmes et leur sélection parmi un pool d'espèces plus large (Weiher et al., 1999), possède la capacité de déterminer les stratégies de gestion les plus pertinentes, permettant ainsi de diriger la succession vers l'état désiré. Toutefois, Chase (2003) suggère que, dans des environnements sujets à des perturbations de grande ampleur et ayant une faible connectivité, l'assemblage des communautés peut aboutir à des états d'équilibre multiples. De tels états stables, alternatifs (Hobbs et al., 1996 ; Temperton et al., 2004) peuvent représenter un challenge pour les responsables de restaurations écologiques.
8Notons enfin que, contrairement à la biologie de la conservation généralement centrée sur une (ou quelques) espèce(s) cible(s), la restauration visant l'échelle de l'écosystème et la reconstruction des fonctions et processus de cet écosystème, ce sont les communautés d'espèces, voire les écosystèmes entiers, qui font l'objet principal et central des études en écologie de la restauration.
3. Filtres à la restauration
9À différentes étapes du processus de restauration, des filtres (ou contraintes) vont intervenir – isolément ou conjointement – entravant la colonisation, l'établissement ou la persistance à long terme de certaines espèces (Tonn et al., 1990). Selon les auteurs, les catégories de filtres distinguées varient (e.g. Fattorini et al., 2004 ; Hobbs et al., 2004 ; Lortie et al., 2004). Nous considérerons trois grandes catégories :
10– les filtres liés à la dispersion,
11– les filtres liés aux conditions abiotiques du site,
12– les filtres d'ordre biotiques (Figure 1).
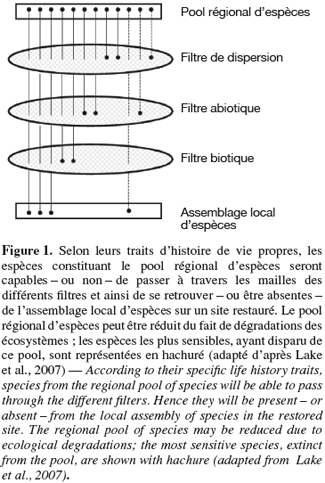
13Ces filtres vont ainsi induire un assemblage local d'espèces (sur le site restauré) constituant un sous-ensemble plus ou moins représentatif du pool d'espèces régional selon la perméabilité des filtres pour les espèces (Rahel, 2002). Par ailleurs, d'après cette perméabilité du filtre (i.e. ses caractéristiques de sélectivité), certains traits d'histoire de vie des espèces seront favorisés au sein de l'assemblage local (Keddy, 1992). L'identification des traits d'histoire de vie sensibles aux différents filtres liés à une restauration écologique peut améliorer la compréhension de la régulation de la distribution des espèces dans le paysage (Poff, 1997).
3.1. Filtres de dispersion
14Les végétaux et animaux se dispersent par le vent et l'eau, à l'aide d'autres espèces, ou par leurs propres mouvements. De façon plus précise, et en ce qui concerne les végétaux, la dispersion peut, en plus d'être spatiale (voie végétative ou diaspores propagées par des agents de dispersion), être temporelle via la banque de graines du sol (Leck et al., 1989 ; Dulière, 2001). Ces différents moyens de dispersion confèrent des capacités de dispersion spatio-temporelles variables selon les espèces leur permettant de franchir avec plus ou moins de facilité les filtres à la dispersion. Un site nouvellement restauré peut, au départ, n'accueillir qu'une proportion limitée, voire nulle, en espèces cibles (en ce compris la banque de graines). Dans ce cas, la colonisation du site restauré par les espèces « manquantes » ne sera possible que par l'arrivée d'individus ayant franchi les filtres à la dispersion, issus de populations voisines (Bakker et al., 1996), dans un contexte spatial satisfaisant, permettant la dispersion de ces espèces et résultant en un enrichissement des assemblages en espèces cibles sur le site restauré (Seabloom et al., 2003). Outre la restauration à proximité d'un site de référence, la connectivité, et donc la maille du filtre de dispersion, peut être améliorée par la mise en place de corridors écologiques entre le site restauré et un site pouvant jouer le rôle de source de propagules ou d'individus (Turner et al., 2001). Par ailleurs, la connectivité et dispersion d'individus entre écosystèmes similaires, restaurés et de référence, permettra par la suite la persistance à long terme de métapopulations à l'échelle du paysage (Hanski et al., 1997 ; Thomas et al., 2001). Par métapopulation, on entend un ensemble de sous-populations, appartenant à une espèce donnée, spatialement structurées et interconnectées entre elles par des flux d'individus. Ces sous-populations pouvant être confrontées à des évènements d'extinction et de recolonisation (via les flux d'individus), distribués dans le temps (Levins, 1969 ; Hanski, 1998). Enfin, la nature de la matrice paysagère est également considérée comme un filtre à la dispersion, pouvant influencer la connectivité fonctionnelle entre les fragments d'habitats cibles (Ricketts, 2001). En effet, selon l'organisme visé, la dispersion à travers la matrice peut générer des couts plus ou moins importants, selon sa perméabilité aux mouvements de l'organisme.
15Les échelles spatiales et temporelles auxquelles est lié un projet de restauration détermineront les possibilités d'actions sur le filtre de dispersion. Malheureusement, ces échelles sont fréquemment déterminées davantage par des contraintes logistiques, économiques et sociales que par une compréhension adéquate des échelles pertinentes pour les processus spécifiques de l'écosystème visé. Lorsqu'une action visant à réduire les filtres de dispersion est hors de portée du projet de restauration, l'introduction de propagules des espèces cibles peut être considérée comme une manière de passer outre le filtre de dispersion, ou tout au moins d'accélérer les successions menant à la restauration (Palmer et al., 1997). De même, des populations existantes sur le site restauré, mais jugées menacées d'extinction locale de par leur faible effectif, pourraient faire l'objet de renforcement des populations (Seabloom et al., 2003) pour autant qu'une attention particulière soit portée à l'introduction d'écotypes locaux, aux génotypes adaptés aux conditions environnementales locales (Montalvo et al., 1997). Néanmoins, avant toute introduction, il est nécessaire de définir la liste des espèces cibles avec précision. Des listes imprécises peuvent résulter en l'introduction d'espèces en dehors de leur niche naturelle de dispersion et étendre ainsi les limites de répartition de certaines (sous-) espèces (McKay et al., 2005).
3.2. Filtres abiotiques
16En général, les premières interventions d'un projet de restauration écologique visent à se rapprocher des conditions abiotiques de l'écosystème de référence (restauration du régime hydrique, dépollution, réduction des excès en nutriments dans le sol, etc.). En restaurant les conditions physico-chimiques propres à un écosystème, les espèces cibles sont susceptibles de retrouver dans le site restauré des conditions propres à leur niche écologique fondamentale. Toutefois, les conditions abiotiques nécessaires à une espèce peuvent grandement varier au cours de son ontogenèse. Ceci est bien connu pour le règne animal, mais s'applique également au règne végétal. Grubb (1977) a suggéré que pour de nombreuses espèces végétales, la période du cycle de vie définissant la niche réalisée de l'espèce soit la période s'étalant de la germination à l'établissement de l'espèce. La niche adulte étant souvent plus large que la niche de recrutement (Young et al., 2005). Ceci induit que les efforts de restauration doivent tenir compte de cette ontogénie de l'habitat, en restaurant les conditions propices au développement des différents stades de vie d'une espèce, mais également en assurant une connectivité suffisante entre les différents (micro-) habitats utilisés par l'espèce (Young et al., 2005). La non considération de cette notion de niches ontogéniques peut déboucher sur la mise en place de populations non-recrutantes. Ceci s'observe lorsque les limites déterminées par la niche de recrutement sont dépassées : e.g. plantation d'individus dans des sites où de jeunes individus et des adultes peuvent persister, mais où les propagules sont incapables de germer avec succès et de s'établir (« populations relictes », Eriksson, 1996).
3.3. Filtres biotiques
17Lorsqu'une espèce du pool régional parvient à franchir les filtres de dispersion (i.e. arrive sur le site restauré) et n'est pas contrainte par les filtres abiotiques, son inclusion dans l'assemblage d'espèces sera déterminée par les filtres biotiques. Ces filtres biotiques consistent en interactions rencontrées par les espèces cibles de la restauration avec le cortège d'espèces liées au site (i.e. espèces présentes sur le site ou pouvant y avoir un quelconque effet). La compétition interspécifique, les relations trophiques et les « espèces nexus » en sont les exemples classiques. Les espèces nexus sont des espèces pouvant n'apparaitre que transitoirement dans la trajectoire de restauration et de développement d'une communauté, mais dont la présence – ou l'absence – se répercute par des effets à long terme importants (Drake et al., 1996 ; Lockwood et al., 2004). À titre d'exemple, Lotus purshianus (Benth.) Clem. & Clem., espèce transitoire des plaines fluviales, fixe l'azote. La dénitrification qui en résulte initie des modifications dans la dynamique des espèces (Sheibley et al., 2006 ; Trowbridge, 2007). L'idée des « ingénieurs écologiques » est similaire. Il s'agit d'espèces dont l'activité peut créer des structures profitant à l'activité d'autres espèces (Jones et al., 1997). Favoriser ces espèces lors du processus de restauration peut ainsi faciliter et améliorer la colonisation par des espèces cibles (Wright et al., 2002).
18Comme le soulignait Poff (1997), il est important de quantifier la nature des filtres biotiques, mais dans la majorité des cas, l'exercice n'est pas aisé. Construire une matrice de coefficients d'interactions mutuelles entre chacune des espèces d'un assemblage, par exemple, est vraisemblablement illusoire. Par contre, se focaliser sur des espèces présentant de fortes interactions biotiques par nature – e.g. espèces dominantes ou ingénieurs écologiques – peut se révéler efficace. La connaissance de leur(s) impact(s) peut permettre d'appréhender la composition des assemblages d'espèces (Poff, 1997).
19Fattorini et al. (2004) ont introduit la notion de filtres dynamiques. De par les interactions mutuelles et continues entre les filtres en jeu lors du processus de restauration, les effets respectifs des filtres sont plus ou moins contraignants vis-à-vis du pool d'espèces. En étudiant un écosystème rivulaire, Fattorini et al. ont considéré la suppression des forces de dégradation de l'écosystème – et donc la diminution du stress environnemental – comme un élargissement des mailles du filtre abiotique permettant ainsi l'établissement d'un plus grand nombre d'espèces cibles. Toutefois, avec ce nombre croissant d'espèces, la compétition pour les ressources augmentait, entrainant un rétrécissement des mailles du filtre biotique. De façon similaire, l'arrivée d'une espèce sur le site restauré peut en modifier les conditions – et donc le filtre – abiotiques.
4. Comment évaluer la réussite d'une restauration ?
20Dans tout projet de restauration, il est important de définir des objectifs clairs et précis. Ces objectifs peuvent être définis à différentes échelles écologiques : populations, communautés, processus écologiques et services de l'écosystème. Le suivi des restaurations est essentiel pour en évaluer le succès et évaluer la réalisation de leurs objectifs. Malheureusement, un suivi insatisfaisant des projets de restauration est assez récurrent (Lake, 2001).
4.1. Une restauration réussie…
21Avant de quantifier la réussite d'une restauration écologique, il convient de préciser ce qu'est une restauration considérée comme réussie. Selon l'International Primer on Ecological Restoration (SER, 2004), un écosystème s'est régénéré (restauré), lorsqu'il possède les neuf attributs suivants :
22– l'écosystème restauré contient un ensemble caractéristique d'espèces de l'écosystème de référence qui procure une structure communautaire appropriée,
23– l'écosystème restauré est constitué pour la plupart d'espèces indigènes,
24– tous les groupes fonctionnels nécessaires à l'évolution continue et/ou à la stabilité de l'écosystème restauré sont représentés ou, s'ils ne le sont pas, les groupes manquant ont la capacité à le coloniser naturellement,
25– l'environnement physique de l'écosystème restauré est capable de maintenir des populations reproductrices d'espèces nécessaires à sa stabilité ou à son évolution continue le long de la trajectoire désirée,
26– l'écosystème restauré fonctionne en apparence normalement lors de sa phase écologique de développement et les signes de dysfonctionnement sont absents,
27– l'écosystème restauré est intégré comme il convient dans une matrice écologique plus large ou un paysage, avec qui il interagit par des flux et des échanges biotiques et abiotiques,
28– les menaces potentielles du paysage alentour sur la santé et l'intégrité de l'écosystème restauré ont été éliminées ou réduites autant que possible,
29– l'écosystème restauré est suffisamment résilient pour faire face à des évènements normaux de stress périodiques de l'environnement local, ce qui sert à maintenir l'intégrité de l'écosystème,
30– l'écosystème restauré se maintient lui-même au même degré que son écosystème de référence et a la capacité à persister indéfiniment sous les conditions environnementales existantes.
4.2. … quantifiée à l'aide d'indicateurs
31Différentes stratégies permettent l'évaluation de la réussite d'une restauration :
32– l'atteinte des objectifs peut être quantifiée par rapport aux attributs spécifiques de sites de référence (Hobbs et al., 1996), ou par rapport à de l'information de référence de source historique ou contemporaine (White et al., 1997) ;
33– une estimation du degré selon lequel les différents attributs (cfr 4.1.) d'un site restauré sont atteints ;
34– l'analyse de la trajectoire se base sur des indicateurs relevés périodiquement sur le site restauré et permet de vérifier la bonne trajectoire du site, dans la direction de la référence. Dans tous les cas, un « bon » indicateur répond à plusieurs critères : facile à mesurer et relativement peu couteux, ne pas présenter de difficultés taxonomiques ou de difficultés de mesures, être sensible aux mesures de restauration, i.e. possibilité d'être inclus directement dans les hypothèses à tester (Keddy, 1999). La facilité et le faible cout d'échantillonnage sont particulièrement importants dans les projets de suivi à long terme. En effet, bien que des budgets parfois importants soient alloués à la restauration écologique, ceux-ci concernent bien souvent des projets à durée restreinte. Les budgets attribués aux suivis, quant à eux, sont généralement peu conséquents, voire totalement inexistants.
35Nous allons brièvement caractériser des indicateurs qui, au vu des objectifs de restauration fixés, permettent d'évaluer la représentativité et la fonctionnalité du cortège d'espèces de l'écosystème restauré (i.e. évaluation des attributs 1-3, cfr supra). Classiquement, l'évaluation – biotique – d'une restauration repose soit sur une approche « espèce », soit sur une approche « communauté ».
36Approche espèce. L'approche espèce fait référence à une espèce cible de la restauration, souvent rare ou menacée, ou un groupe restreint de telles espèces (Simberloff, 1988 ; Brussard, 1991). Les indicateurs concernent, dans ce cas, directement l'état de la (les) population(s) (nombre d'individus, démographie, diversité génétique, etc.). Bien que son utilisation se justifie dans le cadre de suivi de projets de restauration visant explicitement une espèce en particulier, l'approche espèce a fait l'objet de critiques concernant le fait qu'elle n'apporte qu'une information restreinte par rapport aux diverses composantes de biodiversité de l'écosystème à évaluer (Franklin, 1993). Ces mêmes critiques suggéraient des approches considérant des niveaux d'organisation supérieurs tels que les écosystèmes ou les paysages (Noss, 1983 ; Salwasser, 1991). De ces critiques, est né le concept d'espèces parapluie (Frankel et al., 1981 ; Caro, 2003). Il s'agit d'espèces dont les exigences importantes sont supposées englober les exigences – moins étendues – d'un ensemble d'espèces. Ainsi, les espèces parapluie permettent d'appréhender la réponse de toute une série d'espèces face aux mesures de restauration, tout en conservant l'avantage (économique et main-d'œuvre) d'une collecte de données restreinte, puisque ne concernant qu'une espèce. Toutefois, Franklin (1994) souligne que des évaluations (et plus généralement des stratégies de conservation), centrées sur une espèce, même parapluie, peuvent passer à côté d'autres aspects critiques de l'écosystème dans lequel l'espèce se développe. La notion d'espèce parapluie reste utile mais devrait dès lors être inclue dans une approche multi-taxonomique identifiant un ensemble d'espèces (focales) dont les exigences spatiales, compositionnelles et fonctionnelles comprennent les exigences de toutes les autres espèces d'une région (Lambeck, 1997).
37Approche communauté. De son côté, l'approche communauté est supposée saisir l'intégrité, la diversité et la vulnérabilité d'une communauté donnée (Lambeck, 1997 ; Soulé et al., 2001). Des indicateurs classiques en sont la diversité et la richesse spécifique, souvent mises en relation avec les patrons paysagers (Saunders et al., 1991 ; Angelstam, 1992). La richesse spécifique est le nombre d'espèces correspondant à un nombre d'individus (ou une biomasse) donnés ou à une superficie donnée (Magurran, 2004). Notons qu'un problème lié aux estimateurs de richesse spécifique est leur dépendance vis-à-vis de l'effort d'échantillonnage, rendant notamment difficile la comparaison des richesses spécifiques entre études (Gaston, 1996). Certains indices permettent toutefois de remédier à ce problème (e.g. Clifford et al., 1975). La richesse spécifique peut être considérée dans son ensemble (nombre total d'espèces) ou être décomposée en compartiments. La richesse en espèces spécialistes de l'écosystème visé par la restauration est ainsi fréquemment utilisée. En effet, les espèces spécialistes sont davantage sensibles aux modifications environnementales que les espèces généralistes (New, 1997 ; Tscharntke et al., 2002 ; Krauss et al., 2003) et l'impact d'une restauration, de même que les vitesses de (re)colonisation du milieu restauré peuvent ainsi différer entre les différents compartiments de la richesse spécifique. D'autres approches vont plus loin et visent à générer des résultats permettant une meilleure compréhension des mécanismes écologiques sous-jacents aux réponses différenciées entre espèces et déterminant la composition en espèces dans un écosystème donné (Maurer et al., 2003 ; Hérault et al., 2005). Les groupes fonctionnels en sont un exemple. Il s'agit d'ensembles d'espèces ayant des combinaisons de traits d'histoire de vie morphologiques, physiologiques et phénologiques similaires (Lavorel et al., 1997). Dans le cadre d'une approche communauté, l'aspect multi-taxonomique trouve également son intérêt. Le recours à des indicateurs de suivi et d'évaluation répondant à différentes échelles temporelles est préférable pour une évaluation pertinente à long terme. En effet, différents taxons répondent à des vitesses diverses aux modifications environnementales liées à la restauration. Dans un projet de restauration de rivière, Trexler (1995) avait estimé la réponse des végétaux aquatiques à 3-8 ans, celle des invertébrés à 10-12 ans et celle des poissons à 12-20 ans. De telles différences de récupération par les différents taxons doivent être prises en compte et se refléter par un choix judicieux des différents indicateurs d'évaluation.
38Dette d'extinction et crédit de colonisation. Une fois les données de suivi collectées et les différents indices d'évaluation de l'écosystème restauré calculés, il convient de garder à l'esprit ce fait que certaines espèces répondent – presque – immédiatement aux modifications environnementales, alors que d'autres réagiront avec un certain délai. Deux situations de déséquilibre peuvent ainsi apparaitre. Du fait de relations bien établies entre la richesse spécifique et les propriétés spatiales (surface, connectivité, effet lisière, etc.) d'un fragment d'habitat (e.g. Rosenzweig, 1995 ; Bastin et al., 1999 ; Bruun, 2000), on peut théoriquement déterminer la richesse spécifique attendue dans ce fragment, pour un paysage donné. Toutefois, le délai de réponse des espèces peut résulter en un surplus d'espèces par rapport à la situation d'équilibre ; il s'agit d'espèces persistant dans le fragment, alors que leurs exigences ne sont plus rencontrées, et donc vouées à s'éteindre localement dans un avenir plus ou moins proche selon leur délai de réponse. Ce phénomène est connu sous le terme de dette d'extinction (Tilman et al., 1994) et concerne principalement des situations de dégradation d'habitat telles que la fragmentation. À l'inverse, lorsque la situation spatiale d'un fragment d'habitat est « améliorée » (restauration par augmentation de la surface ou de la connectivité), un crédit de colonisation peut être observé et est défini comme le nombre d'espèces dont une colonisation future est attendue (Cristofoli et al., 2009). Ces deux phénomènes ont des implications importantes en termes de conservation. Une dette d'extinction, à l'inverse d'un crédit de colonisation, peut mener à surestimer la richesse spécifique à long terme et de ce fait, à une sous-estimation du niveau de menace pesant sur les espèces (Hanski et al., 2002).
5. Conclusion
39L'écologie de la restauration est une science encore jeune. Bien que les exemples de restauration s'accumulent, ils sont encore bien souvent trop récents que pour bénéficier de suffisamment de recul et pouvoir évaluer leur efficacité de manière pertinente. Or, l'avancée de cette science passe par la compréhension des réussites, autant que des échecs, de restauration. Ceci nécessite un suivi sur le long terme de la réponse des écosystèmes à la restauration et passe par la définition précise d'indicateurs. Le choix des indicateurs est fonction des objectifs fixés ; ils permettent une approche d'autant plus globale qu'ils intègrent un maximum de compartiments de la diversité biologique.
Bibliographie
Angelstam P., 1992. Conservation of communities – the importance of edges, surroundings and landscape mosaic structure. In: Hansson L., ed. Ecological principles of nature conservation. Applications in temperate and boreal environments. London: Elsevier Applied Science, 9-70.
Aronson J. & Le Floc'h E., 1996. Vital landscape attributes: missing tools for restoration ecology. Restoration Ecol., 4, 377-387.
Bakker J.P. et al., 1996. Seed bank and seed dispersal: important topics in restoration ecology. Acta Bot. Neerl., 45, 461-490.
Balmford A. et al., 2005. The convention on biological diversity's 2010 Target. Science, 307, 212-213.
Bastin L. & Thomas C.D., 1999. The distribution of plant species in urban vegetation fragments. Landscape Ecol., 14, 493-507.
Bennett A.F., 1997. Habitat linkages: a key element in an integrated landscape approach to conservation. Parks, 7, 43-49.
Brussard P.F., 1991. The role of ecology in biological conservation. Ecol. Appl., 1(1), 6-12.
Bruun H.H., 2000. Patterns of species richness in dry grassland patches in an agricultural landscape. Ecography, 23, 641-650.
Burel F. & Baudry J., 1999. Écologie du paysage. Concepts, méthodes et applications. Paris : Lavoisier Tec & Doc.
Caro T.M., 2003. Umbrella species: critique and lessons from East Africa. Anim. Conserv., 6, 171-181.
Chase J.M., 2003. Community assembly: when should history matter? Oecologia, 136, 489-498.
Clifford H.T. & Stephenson W., 1975. An introduction to numerical classification. London: Academic Press.
Cristofoli S. et al., 2009. Colonization credit in restored wet heathlands. Restoration Ecol., doi: 10.1111/j.1526-100.2008.00495.x., in press.
Drake J.A., Hewitt C.L., Huxel G.R. & Kolasa J., 1996. Diversity and higher levels of organization. In: Gaston K.J., ed. Biodiversity: a biology of numbers and differences. Cambridge, UK: Oxford Press, 149-166.
Dulière J.F., 2001. L'étude de la banque de graines du sol est un outil efficace qui doit orienter les actions de restauration des milieux semi-naturels menacés. Thèse annexe de doctorat : Faculté universitaire des Sciences agronomiques de Gembloux (Belgique).
Eriksson O., 1996. Regional dynamics of plants: a review of evidence for remnant, source-sink and metapopulations. Oikos, 77, 248-258.
Eriksson O. & Ehrlén J., 2001. Landscape fragmentation and the viability of plant populations. In: Silvertown J. & Antonovics J., eds. Integrating ecology and evolution in a spatial context. Oxford, UK: Blackwell Publications, 157-175.
Fattorini M. & Halle S., 2004. The dynamic environmental filter model: how do filtering effects change in assembling communities after disturbance. In: Temperton V.M., Hobbs D.A., Nuttle T. & Halle S., eds. Assembly rules and restoration ecology: bridging the gap between theory and practice. Washington, DC, USA: Island Press, 94-116.
Frankel O.H. & Soulé E., 1981. Conservation and evolution. Cambridge, UK: Cambridge University Press.
Franklin J.F., 1993. Preserving biodiversity: species, ecosystems or landscapes? Ecol. Appl., 3, 202-205.
Franklin J.F., 1994. Preserving biodiversity: species in landscapes. Response to Tracy and Brussart. Ecol. Appl., 4, 208-209.
Gaston K.J., 1996. Spatial covariance in the species richness of higher taxa. In: Hochberg M.E., Clobert J. & Barbault R., eds. Aspects of the genesis and maintenance of biological diversity. Oxford, UK: Oxford University Press, 221-242.
Godron M. & Forman R.T.T., 1983. Landscape modification and changing ecological characteristics. In: Mooney H.A. & Godron M., eds. Disturbance and ecosystems: components of response. New York, USA: Springer-Verlag, 12-28.
Grubb P., 1977. The maintenance of species richness in pant communities: the importance of the regeneration niche. Bot. Rev., 52, 107-145.
Hanski I., 1998. Metapopulation dynamics. Nature, 396(5), 41-49.
Hanski I. & Ovaskainen O., 2002. Extinction debt at extinction threshold. Conserv. Biol., 16(3), 666-673.
Hanski I. & Simberloff D., 1997. The metapopulation approach: its history, conceptual domain, and application to conservation. In: Hanski I.A. & Gilpin M.E., eds. Metapopulation biology. San Diego, CA, USA: Academic Press, 5-25.
Harrisson S. & Bruna E., 1999. Habitat fragmentation and large-scale conservation: what do we know for sure? Ecography, 22, 225-232.
Hérault B. & Honnay O., 2005. The relative importance of local, regional and historical factors determining the distribution of plants in fragmented riverine forests: an emergent group approach. J. Biogeogr., 32, 2069-2081.
Hobbs R.J. & Norton D.A., 1996. Towards a conceptual framework for restoration ecology. Restoration Ecol., 4, 93-110.
Hobbs R.J. & Norton D.A., 2004. Ecological filters, thresholds, and gradients in resistance to ecosystem reassembly. In: Temperton V.M., Hobbs R.J., Nuttle T. & Halle S., eds. Assembly rules and restoration ecology: bridging the gap between theory and practice. Washington, DC, USA: Island Press, 72-95.
Jones C.G., Lawton J.H. & Shachak M., 1997. Positive and negative effects of organisms as physical ecosystem engineers. Ecology, 78, 1946-1957.
Keddy P.A., 1992. Assembly and response rules: two goals for predictive community ecology. J. Veg. Sci., 3, 157-164.
Keddy P.A., 1999. Wetland restoration: the potential for assembly rules in the service of restoration. Wetlands, 19, 716-732.
Krauss J., Steffan-Dewenter I. & Tscharntke T., 2003. How does landscape context contribute to effects of habitat fragmentation on diversity and population density of butterflies? J. Biogeogr., 30, 889-900.
Lake P.S., 2001. On the maturing of restoration: linking ecological research and restoration. Ecol. Manage. Restoration, 2, 110-115.
Lake P.S., Bond N. & Reich P., 2007. Linking ecological theory with stream restoration. Freshwater Biol., 52(4), 597-615.
Lambeck R.J., 1997. Focal species: a multi-species umbrella for nature conservation. Conserv. Biol., 11(4), 849-856.
Lavorel S., Mcintyre S., Landsberg J. & Forbes T.D.A., 1997. Plant functional classifications: from general groups to specific groups based on response to disturbance. Trends Ecol. Evol., 12, 474-478.
Le Floc'h E. & Aronson J., 1995. Écologie de la restauration. Définition de quelques concepts de base. Nat. Sci. Soc., 3, 29-35.
Leck M.A., Parker V.T. & Simpson R.L., 1989. Ecology of soil seed banks. San Diego, CA, USA: Academic Press Inc.
Levins R., 1969. Some demographic and genetic consequences of environmental heterogeneity for biological control. Bull. Entomol. Soc. Am., 15, 236-240.
Lockwood J.L. & Samuels C.L., 2004. Assembly models and the practice of restoration. In: Temperton V.M., Hobbs R.J., Nuttle T. & Halle S., eds. Assembly rules and restoration ecology: bridging the gap between theory and practice. Washington, DC, USA: Island Press, 34-54.
Lortie C.J. et al., 2004. Rethinking plant community theory. Oikos, 107(2), 433-438.
Madgwick F.J. & Jones T.A., 2002. Europe. In: Perrow M.R. & Davy A.J., eds. Handbook of ecological restoration. Principles of restoration. Vol. 2. Cambridge, UK: Cambridge University Press, 32-56.
Magurran A.E., 2004. Measuring biological diversity. Cornwall, ON, Canada: Blackwell Publishing.
Maurer K., Durka W. & Stöcklin J., 2003. Frequency of plant species in remnants of calcareous grassland and their dispersal and persistence characteristics. Basic Appl. Ecol., 4(4), 307-316.
McKay J., Christian C., Harrison S.P. & Rice K.J., 2005. How local is local? A review of practical and conceptual issues in the genetics of restoration. Restoration Ecol., 13(3), 432-440.
Montalvo A.M. et al., 1997. Restoration biology: a population biology perspective. Restoration Ecol., 5, 277-290.
New T.R., 1997. Are the Lepidoptera an effective “umbrella group” for biodiversity conservation? J. Insect Conserv., 1, 5-12.
Noss R.F., 1983. A regional landscape approach to maintain biodiversity. Bioscience, 33, 700-706.
Ouborg N.J., 1993. Isolation, population size and extinction: the classical and metapopulation approaches applied to vascular plants along the Dutch Rhine-system. Oikos, 66, 298-308.
Palmer M.A., Ambrose R.F. & Poff N.L., 1997. Ecological theory and community restoration ecology. Restoration Ecol., 5, 291-300.
Pickett S.T.A. & Parker V.T., 1994. Avoiding old pitfalls: opportunities in a new discipline. Restoration Ecol., 2, 75-79.
Poff N.L., 1997. Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology. J. North Am. Benthological Soc., 16(2), 391-409.
Rahel F.J., 2002. Homogeneization of freshwater faunas. Ann. Rev. Ecol. Syst., 33, 291-315.
Ricketts T.H., 2001. The matrix matters: effective isolation in fragmented landscapes. Am. Nat., 158(1), 88-99.
Rosenzweig M.L., 1995. Species diversity in space and time. Cambridge, UK: Cambridge University Press.
Salwasser H., 1991. In search of an ecosystem approach to endangered species conservation. In: Kohm K.A., ed. Balancing on the brink of extinction: the endangered species act and lessons for the future. Washington, DC, USA: Island Press.
Saunders D.A., Hobbs R.J. & Margules C.R., 1991. Biological consequences of ecosystem fragmentation: a review. Conserv. Biol., 5(1), 18-32.
Seabloom E.W. & van der Valk A.G., 2003. Plant diversity, composition, and invasion of restored and natural prairie pothole wetlands: implications for restoration. Wetlands, 23, 1-12.
SER, 2004. The SER (Society for Ecological Restoration International Science & Policy Working Group) international primer on ecological restoration, http://www.ser.org/content/ecological_restoration_primer.asp, (30/06/09).
Sheibley R.W., Ahearn D.S. & Dahlgren R.A., 2006. Nitrate loss from a restored floodplain in the lower Cosumnes River, California. Hydrobiologia, 571, 261-272.
Sih A., Jonsson B.G. & Luikart G., 2000. Habitat loss: ecological, evolutionary and genetic consequences. Trends Ecol. Evol., 15, 132-134.
Simberloff D.S., 1988. The contribution of population community biology to conservation science. Ann. Rev. Ecol. Syst., 19, 473-511.
Soulé M.E. & Orians G.H., 2001. Conservation biology research: its challenges and contexts. In: Soulé M.E. & Orians G.H., eds. Conservation biology, research priorities for the next decade. Washington, DC, USA: Island Press, 271-285.
Temperton V.M. & Hobbs R.J., 2004. The search for ecological assembly rules and its relevance to restoration ecology. In: Temperton V.M., Hobbs R.J., Nuttle T. & Halle S., eds. Assembly rules and restoration ecology: bridging the gap between theory and practice. Washington, DC, USA: Island Press, 34-54.
Thomas J.A. et al., 2001. The quality and isolation of habitat patches both determine where butterflies persist in fragmented landscapes. Proc. R. Soc. London Ser. B, Biol. Sci., 268(1478), 1791-1796.
Tilman D., May R.M., Lehman C.L. & Nowak M.A., 1994. Habitat destruction and the extinction debt. Nature, 371, 65-66.
Tonn W.M., Magnuson M., Rask M. & Toivonen J., 1990. Intercontinental comparison of small-lake fish assemblages: the balance between local and regional processes. Am. Nat., 136, 345-375.
Trexler J.C., 1995. Restoration of the Kissimmee river: a conceptual model of past and present fish communitites and its consequences for evaluating restoration success. Restoration Ecol., 3, 195-210.
Trowbridge W.B., 2007. The role of stochasticity and priority effects in foodplain restoration. Ecol. Appl., 17(5), 1312-1324.
Tscharntke T., Steffan-Dewenter I., Kruess A. & Thies C., 2002. Characteristics of insect populations on habitat fragments: a mini-review. Ecol. Res., 17, 229-239.
Turner M.G., 1989. Landscape ecology: the effect of pattern on process. Ann. Rev. Ecol. Syst., 20, 171-197.
Turner M.G., Gardner R.H. & O'Neill R.V., 2001. Landscape ecology in theory and practice: pattern and process. New York, USA: Springer Verlag.
UNEP, 2007. Global environmental outlook - GEO 4. Valletta, Malta: Progress Press.
Wali M.L., 1987. The structure, dynamics, and rehabilitation of drastically disturbed ecosystems. In: Khoshoo T.N., ed. Perspectives in environmental management. New Delhi, India: Oxford and IBH Publishing, 163-183.
Weiher E. & Keddy P., 1999. Assembly rules as general constraints on community composition. In: Weiher E. & Keddy P., eds. Assembly rules: perspectives, advances, retreats. Cambridge, UK: Cambridge University Press, 251-271.
Westhoff V., 1983. Man's attitude towards vegetation. In: Holzner W., Werger M.J.A. & Ikusima I., eds. Man's impact on vegetation. The Hague, The Netherlands: Dr. W. Junk Publishers, 7-21.
White P.S. & Walker J.L., 1997. Approximating nature's variation: selecting and using reference information in restoration ecology. Restoration Ecol., 5(4), 338-349.
Wright J.P., Jones C.G. & Flecker A.S., 2002. An ecosystem engineer, the beaver, increases species richness at the landscape scale. Oecologia, 132, 96-101.
Young T.P., Petersen D.A. & Clary J.J., 2005. The ecology of restoration: historical links, emerging issues and unexplored realms. Ecol. Lett., 8, 662-673.
Pour citer cet article
A propos de : Sara Cristofoli
ULg - Gembloux Agro-Bio Tech. Laboratoire d'Écologie. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : cristofoli.s@gmail.com
A propos de : Grégory Mahy
ULg - Gembloux Agro-Bio Tech. Laboratoire d'Écologie. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : G.Mahy@ulg.ac.be
