


- Portada
- Volume 14 (2010)
- numéro 2
- Synthèse bibliographique : l’écologie chimique des coccinelles


Vista(s): 6849 (84 ULiège)
Descargar(s): 439 (6 ULiège)

Synthèse bibliographique : l’écologie chimique des coccinelles

Notes de la rédaction
Reçu le 15 mai 2009, accepté le 11 septembre 2009
Résumé
Cet article présente une synthèse de l’écologie chimique des coccinelles (Coleoptera : Coccinellidae) en vue de présenter le rôle des sémiochimiques impliqués dans les relations plante-coccinelle, proie-coccinelle et prédateur-coccinelle. Les coccinelles utilisent ces composés pour localiser leurs proies, s’accoupler, se protéger de la prédation ou du cannibalisme, trouver un abri où passer l’hiver ou assurer une meilleure survie pour leur descendance. L’étude approfondie des comportements des coccinelles vis-à-vis de ces molécules pourrait aboutir à leur utilisation en lutte biologique en maximisant l’efficacité des coccinelles dans le contrôle des ravageurs, comme les pucerons ou les cochenilles.
Abstract
Review: chemical ecology of ladybird beetles. This paper reviews the chemical ecology of ladybird beetles (Coleoptera: Coccinellidae) to present the role of semiochemicals involved in plant-ladybird, prey-ladybird and predator-ladybird interactions. Ladybird beetles use these compounds to locate their prey, mate, protect themselves from predation or cannibalism, find a shelter to overwinter or ensure a better survival for their offspring. Thorough studies on ladybird behaviors towards these compounds could lead to their practical implementation in integrated strategies using ladybirds to control pests, like aphids or mealybugs.
Tabla de contenidos
1. Introduction
1De récentes avancées scientifiques ont permis de comprendre l’importance de l’utilisation des molécules sémiochimiques dans le domaine de la lutte biologique. Les insectes communiquent entre eux notamment par le biais de ces molécules odorantes. Les substances sémiochimiques impliquées dans ces relations sont généralement de deux types : les phéromones et les substances allélochimiques (Figure 1). Les substances allélochimiques possèdent une action interspécifique, tandis que les phéromones ont une action intraspécifique (Brossut, 1996). Trois groupes peuvent être distingués au sein des substances allélochimiques : les allomones, les kairomones et les synomones. Les allomones ne sont bénéfiques qu’à l’organisme qui les émet, les kairomones procurent un avantage uniquement à l’organisme qui les reçoit et les synomones sont favorables à la fois à l’organisme qui les émet et à celui qui les reçoit. Les phéromones, quant à elles, sont classifiées en fonction de leur rôle. On distingue entre autres les phéromones sexuelles, les phéromones d’agrégation, les phéromones de piste, les phéromones d’espacement et les phéromones d’alarme.
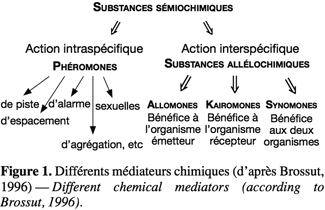
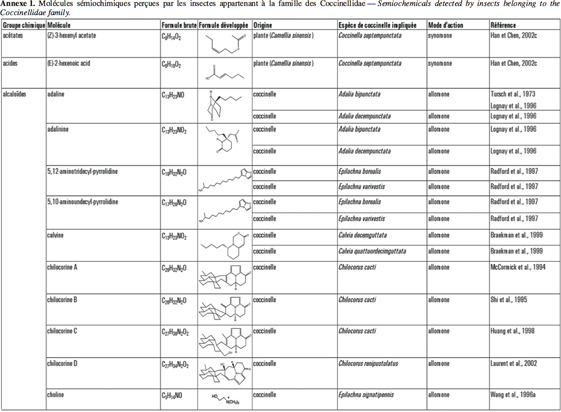
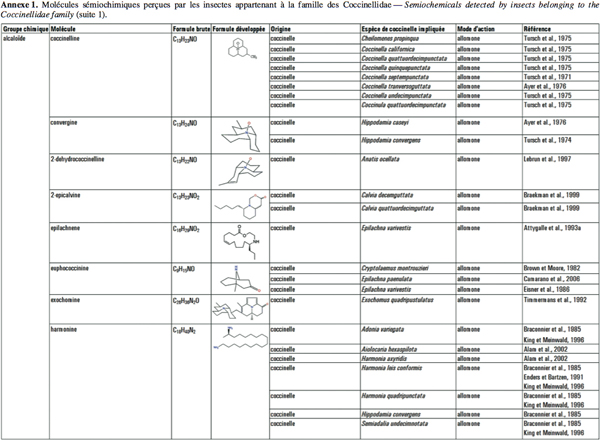
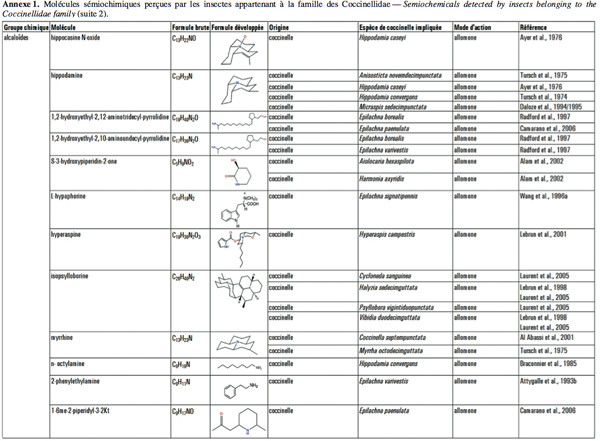
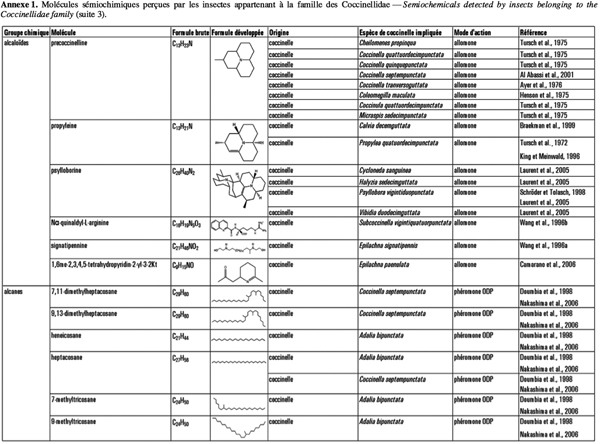
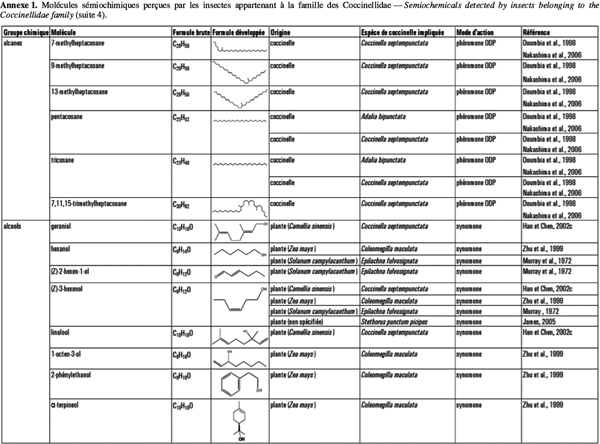
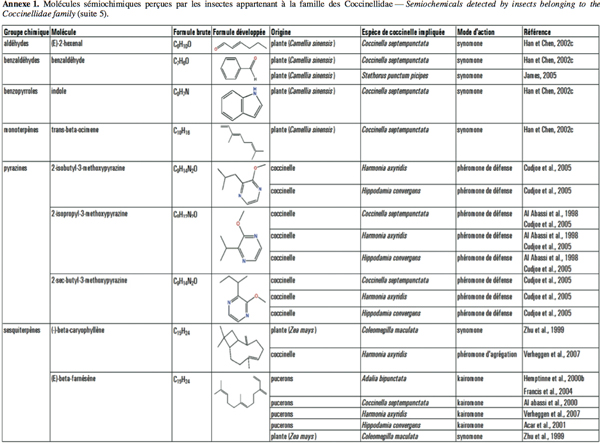
2Cet article fait le bilan de l’écologie chimique des coccinelles pour comprendre le rôle des messagers chimiques dans les interactions qu’elles entretiennent avec leur environnement en vue d’utiliser ces prédateurs dans des méthodes de lutte biologique de façon plus efficiente. L'annexe 1 reprend les molécules sémiochimiques perçues par les insectes appartenant à la famille des Coccinellidae.
2. Communication entre plantes et coccinelles
3Les plantes soumises à un stress, comme l’attaque par un herbivore, peuvent modifier le profil des substances volatiles qu’elles émettent et ce, en vue de faire fuir ce parasite et/ou de devenir plus attractives envers les ennemis naturels (prédateurs ou parasitoïdes) de l’herbivore en question. Plusieurs études ont démontré que diverses espèces de coccinelles sont capables d’utiliser ces sémiochimiques pour localiser l’hôte de leurs proies dans un environnement pourtant riche en signaux olfactifs.
4Les adultes de la coccinelle à sept points Coccinella septempunctata L. détectent les composés chimiques émis par certaines espèces végétales de la famille des Brassicaceae [Brassica rapa L. cultivar ‘Turnip purple top’, Brassica juncea L. cultivar ‘red giant mustard’, Brassica napus L. cultivar ‘Courage’ et Arabidopsis thaliana (L.) Heynh.] infestées ou blessées par le puceron vert du pêcher, Myzus persicae Sulzer. Il semble que ces composés aient un effet immobilisant, voire attractif, sur C. septempunctata (Girling et al., 2008). De même, Ponsonby et al. (1995) ont mis en évidence le caractère attractif des odeurs émises par la combinaison Solanum tuberosum L./Abgrallaspis cyanophylli Sign. envers les coccinelles Chilocorus nigritus F. femelles. Une augmentation de la vitesse de déplacement de ces dernières et du temps qu’elles passent dans les zones où ces odeurs sont présentes a également été observée (Ponsonby et al., 1995).
5D’autres études ont démontré que C. septempunctata est attirée par les substances volatiles émises par des plants d’orge (Hordeum vulgare L.) infestés par le puceron du merisier à grappes, Rhopalosiphum padi L., ainsi que par le cis-jasmone (ou (Z)-jasmone), composé volatil produit par certaines plantes lorsque celles-ci sont endommagées par des insectes herbivores, par exemple (Birkett et al., 2000 ; Ninkovic et al., 2001). C. septempunctata, tout comme la coccinelle asiatique, Harmonia axyridis Pallas, est également attirée par les molécules à action synomone (benzaldéhyde) produites par les plantes de thé (Camellia sinensis L.) attaquées par le puceron Toxoptera aurantii Boyer (Han et al., 2002a ; 2002b).
6La plante Glycine max (L.) Merr. produit plus de méthylsalicylates lorsqu’elle est attaquée par le puceron Aphis glycines L. (par comparaison à une plante saine ou une plante blessée artificiellement). Des expérimentations réalisées en champs ont démontré le pouvoir attractif de ce composé envers C. septempunctata (Zhu et al., 2005). H. axyridis ne semble pas montrer de préférence particulière pour ce composé mais elle est capable d’utiliser les volatils émis par d’autres plantes. En effet, Heit et al. (2005) ont démontré qu’elle est attirée par les composés émis par des feuilles de Solanum sisymbriifolium Lam. attaquées par M. persicae. Les femelles Exochomus flaviventris Mader, quant à elles, sont capables d’évaluer la qualité d’un site de ponte en utilisant les volatils émis par les plantes de manioc infestées par Phenacoccus manihoti Matile-Ferrero (le Rü et al., 2001).
7Les coccinelles peuvent également être attirées par des plantes qui ne sont ni infestées ni blessées (Hamilton et al., 1999). En effet, des observations réalisées en champ ont révélé que C. septempunctata était plus fréquemment présente dans les parcelles contenant de grandes quantités des deux plantes messicoles Cirsium arvense (L.) Scop. et Elytrigia repens (L.) Nevski que dans les parcelles ne contenant que de l’orge (Ninkovic et al., 2003). Des essais olfactométriques ont montré que la coccinelle adulte était plus attirée par le mélange des odeurs provenant des trois plantes que par l’odeur de l’orge seul.
8De même, une réponse électroantennographique significative est observée pour les antennes de Coleomegilla maculata DeGeer lorsque celles-ci sont stimulées avec les composés volatils provenant du maïs dont l’(E)-β-farnésène, l’α-terpineol, le 2-phényléthanol, le β-caryophyllène, le (Z)-3-hexenol, l’hexanol et le 1-octen-3-ol (Zhu et al., 1999).
9Ces résultats montrent que les coccinelles peuvent interagir avec les caractéristiques botaniques de leur environnement en répondant aux odeurs provenant de l’habitat du ravageur pour localiser leurs proies.
10Malgré l’importance grandissante des plantes transformées génétiquement, peu d’études concernant le sujet ont été réalisées. Quoi qu’il en soit, les espèces généralistes comme C. maculata semblent particulièrement compatibles avec les cultures transgéniques où des proies ou du pollen sont disponibles (Riddick et al., 1998).
3. Communication entre les coccinelles et leurs proies
11En plus de détecter les substances volatiles émises par les plantes, il a été reporté que les coccinelles se dirigent également en fonction des molécules volatiles émises par leurs proies (Obata, 1986 ; Sengonca et al., 1994 ; Hamilton et al., 1999).
12Lorsqu’il est perturbé, un puceron sécrète via ses cornicules des gouttelettes contenant une phéromone d’alarme, ce qui provoque la fuite de ses congénères. La constitution chimique de ces sécrétions varie d’une espèce de puceron à l’autre, mais est constituée essentiellement, ou en totalité, d’(E)-β-farnésène (EBF) (Francis et al., 2005). Il a été démontré qu’Adalia bipunctata L. (Hemptinne et al., 2000b ; Francis et al., 2004), Hippodamia convergens Guérin-Méneville (Acar et al., 2001), C. septempunctata (Al Abassi et al., 2000) et H. axyridis (Verheggen et al., 2007) étaient attirées par l’EBF.
13L’(E)-β-farnésène est également un composé sécrété par les plantes. Mais dans ce cas, la réponse des prédateurs diffère à cause de la détection d’autres composés, dont le (-)-β-caryophyllène (Dawson et al., 1984). La réponse d’alarme des pucerons est caractérisée par une augmentation rapide de l’(E)-β-farnésène par rapport au (-)-β-caryophyllène et les coccinelles semblent pouvoir différencier l’émission continue d’EBF d’origine végétale de l’émission instantanée d’origine aphidienne.
14Une expérience réalisée en olfactomètre à quatre voies par Francis et al. (2004) a démontré qu’A. bipunctata n’était attirée par l’(E)-β-farnésène que lorsque la dose de cette molécule déposée dans l’olfactomètre excédait 2 µg. Ceci peut expliquer pourquoi les coccinelles ne sont pas attirées par des pucerons non stressés : la quantité d’EBF produite est trop faible pour être détectée par ces prédateurs (Francis et al., 2004).
15La phéromone d’alarme n’est pas la seule molécule sémiochimique des pucerons utilisée par les coccinelles pour les localiser. En effet, Zhu et al. (1999) ont démontré que les antennes de C. maculata répondaient significativement aux deux composés constitutifs des phéromones sexuelles de pucerons, (4aS, 7S, 7aR)-nepetalactone et (1R, 4aS, 7S, 7aR)-nepetalactol (Dawson et al., 1989).
16Les proies homoptères, comme les pucerons et les cochenilles, se nourrissent de la sève élaborée des plantes et excrètent l’excès de sucre dans des gouttes de miellat. Les coccinelles peuvent dès lors utiliser ces gouttes comme kairomones lorsqu’elles sont en recherche de proies. En effet, les coccinelles passent plus de temps à chercher leurs proies sur des plantes où du miellat est présent que sur les plantes dépourvues de miellat (Carter et al., 1984 ; Heidari et al., 1993 ; Han et al., 2002b ; Ide et al., 2007).
17Il a également été mis en évidence que le miellat avait un effet immobilisant à la fois sur les larves et les adultes de coccinelles (Greany et al., 1981 ; Carter et al., 1984 ; van den Meiracker et al., 1990). De plus, Evans et al. (1997) ont mis en évidence une augmentation de la densité des coccinelles aphidiphages sur les parcelles traitées avec du miellat artificiel (solution de saccharose dissoute dans de l’eau).
18Le miellat permet donc aux coccinelles de localiser leurs proies mais en plus de cela, comme il représente un mélange complexe de carbohydrates, d’acides aminés et de métabolites secondaires (Wiedemann et al., 2004 ; Kazana et al., 2007), il constitue une source énergétique importante pour les coccinelles. La valeur nutritive d’une même espèce de puceron peut cependant varier en fonction de la plante hôte de laquelle le puceron s’est nourri (Malcolm, 1992 ; Martos et al., 1992 ; Francis et al., 2001 ; Giles et al., 2002). Quelques espèces de pucerons se révèlent même être toxiques pour certaines coccinelles en raison de composés allélochimiques puisés chez leur plante hôte (Hodek et al., 1996). Ceci a notamment été observé chez Aphis nerii Boyer de Fonscolombe infestant le laurier-rose (Rothschild et al., 1970 ; Malcolm, 1990). La toxicité de cette plante est due à la présence de cardénolides (dont les principaux sont l’oléandrine et la nériine). Lorsqu’A. nerii se nourrit de la sève élaborée du laurier-rose, il ingère ces composés toxiques, les stocke et les excrète par la suite dans le miellat.
19Bhatkar (1982) a démontré que C. septempunctata, Coccinella undecimpunctata L. et A. bipunctata étaient également attirées par les trainées de fourmis (Formica polyctena Förster) et obtenaient de ce fait des informations sur la distance et la direction des sources de proies. En effet, plusieurs espèces de pucerons entretiennent une relation de mutualisme avec les fourmis. Ces dernières protègent les pucerons en échange de nourriture fournie sous forme de miellat (Jiggins et al., 1993 ; Majerus et al., 2007).
20Si les molécules émises par les pucerons peuvent agir comme kairomones, il en va de même des composés émis par les coccinelles et plus particulièrement ceux présents dans leurs traces larvaires. En effet, Kunert et al. (2005) ont démontré l’importance des antennes chez Acyrthosiphon pisum (Harris) pour l’induction de la production d’individus ailés en présence d’ennemis naturels, dont les larves et les adultes de coccinelles. Ils suggèrent donc que les substances chimiques émises par ces dernières soient responsables de l’augmentation du nombre d’individus ailés. Dixon et al. (1999) confortent cette hypothèse en démontrant que le puceron du pois, A. pisum, produit plus d’individus ailés en présence de traces larvaires d’A. bipunctata. Ceci lui permet de ce fait de réduire le risque de prédation. Ils ont également observé une augmentation de l’activité des pucerons ainsi qu’une réduction de leur fécondité. En revanche, Megoura viciae Buckton, qui est toxique pour A. bipunctata et Aphis fabae Scopoli, qui est une proie de moindre qualité pour A. bipunctata et qui est fréquemment protégé des coccinelles par les fourmis, ne présentent pas la même réponse qu’A. pisum aux traces larvaires de la coccinelle à deux points (Dixon et al., 1999).
4. Communication entre les coccinelles et leurs prédateurs/parasitoïdes
21Les coccinelles se défendent en émettant certaines substances chimiques : leur couleur voyante indique qu’elles contiennent des alcaloïdes toxiques (King et al., 1996 ; Bezzerides et al., 2007). Pour repousser leurs prédateurs, elles ont également recours à l’autohémorrhée. En effet, lorsqu’elles sont dérangées, les coccinelles émettent via leurs articulations de l’hémolymphe (liquide jaune orangé, à l'odeur et au gout repoussants). Cette substance est chargée de ces alcaloïdes. Qui plus est, des alkylméthoxypyrazines volatils sont associés à ces alcaloïdes et agissent comme signal d’alerte olfactif (Moore et al., 1990 ; Cai et al., 2007). D’après Guilford et al. (1987), les coccinelles synthétisent seules leurs pyrazines, elles ne les puisent pas des plantes comme d’autres insectes tels que le papillon monarque Danaus plexippus L. Elles peuvent cependant stocker des composés toxiques ou répulsifs provenant de leurs proies. En effet, Rothschild et al. (1973) ont démontré que C. undecimpunctata séquestre des glycosides cardiaques lorsqu’elle se nourrit du puceron du laurier rose, A. nerii, qui est lui-même connu pour stocker ces composés (Rothschild et al., 1970). En revanche, les coccinelles à sept points, C. septempunctata, qui viennent d’émerger et qui se sont nourries de ce puceron durant leurs stades larvaires sont dépourvues de ces glycosides cardiaques (Rothschild et al., 1973). Les larves d’Hyperaspis trifurcata Schaeffer stockent quant à elles de l’acide carminique à partir de leurs proies cochenilles (Eisner et al., 1994). Lorsqu’elles sont perturbées, ces larves sécrètent de l’hémolymphe qui contient de l’acide carminique en concentrations suffisantes pour être répulsif envers les fourmis. Les coccinelles sont également capables de séquestrer des alcaloïdes d’origine végétale. Les alcaloïdes pyrrolizidines (PA) constituent un groupe de composés végétaux secondaires fréquemment retrouvés chez différents genres tels les Astéracaeae, Boraginaceae et Fabaceae. Les coccinelles se nourrissant de pucerons élevés sur des plantes produisant ces alcaloïdes sont capables d’accumuler des quantités considérables de ces PA (Witte et al., 1990). Ces composés peuvent atteindre de 10 à 50 % de la quantité des alcaloïdes produits par la coccinelle elle-même.
22Tous les stades (de l’œuf à la pupe) de nombreuses espèces de la sous-famille des Coccinellinae produisent des alcaloïdes pour se protéger de la prédation des autres coccinelles (Agarwala et al., 1991 ; Hemptinne et al., 2000a) mais aussi des oiseaux (Marples et al., 1989) et des fourmis (Pasteels et al., 1973 ; Ayer et al., 1977 ; Marples, 1993). Cependant, dans certains cas, les alcaloïdes endogènes peuvent avoir une action kairomonale en attirant les ennemis naturels des coccinelles. En effet, al Abassi et al. (2001) ont mis en évidence le pouvoir attractif de la precoccinelline et de la myrrhine, présents chez C. septempunctata, envers Dinocampus coccinellae Schrank (Hymenoptera : Braconidae), principal parasitoïde d’un nombre important d’espèces de coccinelles.
23D’autres composés chimiques interviennent dans la protection des coccinelles. En effet, des alcanes sont présents en surface des œufs de C. septempunctata, A. bipunctata (Hemptinne et al., 2000a), Calvia quatuordecimguttata L. (Ware et al., 2008), Menochilus sexmaculata (Fabricius) et Coccinella transversalis (Fabricius) (Agarwala et al., 2001) pour éviter la prédation intraguilde et le cannibalisme. Des alcènes ont également été rencontrés en surface des œufs mais selon Ware et al. (2008), ce ne serait le cas que pour C. quatuordecimguttata.
24Il a été démontré que les œufs étaient mieux protégés lorsque leur revêtement contenait de plus grandes quantité et diversité d’hydrocarbures, c'est-à-dire d’alcanes (Ware et al., 2008). Il apparait également que la force de cette défense chimique augmente significativement lorsque les œufs sont déposés en groupes par rapport à celle observée lorsque les œufs sont déposés isolément (Agarwala et al., 2001). En effet, déposer ses œufs en groupes confère un avantage pour la coccinelle en concentrant les substances défensives présentes en surface des œufs et ceci aide à éloigner les prédateurs intraguildes.
25Les coccinelles mangent plus volontiers leurs propres œufs que des œufs hétérospécifiques. Ceci a été observé chez C. septempunctata, A. bipunctata (Agarwala et al., 1991 ; Hemptinne et al., 2000a), Propylea dissecta (Mulsant), C. transversalis (Omkar et al., 2004), Cheilomenes sexmaculata (Fabricius) (Agarwala et al., 1998), H. axyridis et Cycloneda sanguinea L. (Michaud, 2002). Hemptinne et al. (2000a) avancent que cette asymétrie serait due au fait que le danger de prédation intraguilde est plus grand pour les coccinelles que celui de cannibalisme. Les substances chimiques présentes en surface des œufs semblent jouer un rôle prépondérant dans cette préférence car le comportement cannibale des larves diffère lorsque la composition chimique des œufs conspécifiques est modifiée (Omkar et al., 2004). Les composés chimiques présents en surface des œufs conspécifiques agissent apparemment comme stimulant pour l’alimentation, ce qui pourrait être moins efficace ou absent chez les œufs hétérospécifiques (Omkar et al., 2004).
26Des alcanes, similaires à ceux présents sur la surface des œufs, se retrouvent dans les traces laissées par les larves ou les adultes et sur les élytres des adultes. Ceux présents sur les élytres ont un rôle de reconnaissance lors de l’accouplement (Hemptinne et al., 1998), tandis que ceux présents dans les traces larvaires empêchent les femelles de pondre à un endroit où des larves conspécifiques sont présentes (Doumbia et al., 1998). Certains ennemis naturels de pucerons (Aphidius eadyi Stary, Aphidius ervi Haliday, Praon volucre Haliday) utilisent les composés présents dans les traces laissées par les coccinelles adultes pour éviter de pondre aux endroits où des coccinelles seraient présentes (Nakashima et al., 2004 ; 2006). Les composés responsables de ce comportement sont essentiellement le n-tricosane, le n-pentacosane et le n-heptacosane. Les deux espèces d’Aphidius précitées ne réagissent qu’aux deux premiers composés, tandis que P. volucre répond aux trois hydrocarbones (Nakashima et al., 2006). Ceci s’explique par le fait que P. volucre est un parasitoïde plus généraliste que les deux autres espèces étudiées, aussi bien en ce qui concerne les espèces hôtes que les plantes hôtes (Pennacchio, 1989). Dans ce cas, une sensibilité générale aux traces chimiques est plus avantageuse étant donné que les parasitoïdes possédant une gamme d’habitats plus étendue peuvent rencontrer un plus grand nombre d’espèces prédatrices.
5. Communication intraspécifique
5.1. Phéromones sexuelles
27Les phéromones sexuelles des coccinelles n’ont pas été identifiées jusqu’à présent mais diverses études aboutissent à la conclusion que de telles substances existent.
28Tout d’abord, les mâles de Propylea dissecta (Mulsant) sont capables de s’accoupler avec des femelles mortes (Omkar et al., 2005). Le temps d’examen de la femelle par le mâle augmente avec le nombre de jours de mort de la femelle, les femelles mortes depuis un mois étant délaissées (Omkar et al., 2005). Ceci suggère que des molécules chimiques sont responsables de l’attraction des mâles lors de l’accouplement. Ces composés seraient donc toujours présents chez les femelles mortes mais ils s’estomperaient au fur et à mesure du temps.
29Ensuite, Zhu et al. (1999) ont démontré que les extraits de femelles de C. maculata induisaient une réponse antennaire significative chez les mâles de cette espèce. Ceci suggère que les substances extraites des femelles pourraient constituer la phéromone sexuelle de C. maculata.
30Qui plus est, les mâles de l’espèce Leptothea galbula (Mulsant) peuvent garder une pupe femelle quiescente jusqu’à l’émergence de la femelle adulte (Richards, 1980). Ceci semble être lié à la production d’une phéromone sexuelle par les femelles, aussi bien au stade pupal qu’au stade adulte. Pendant tout le temps que le mâle garde la femelle, il ne se nourrit pas, nettoie constamment ses tarses et palpe le thorax de la pupe (Richards, 1980). Ceci suggère que la phéromone sexuelle produite agit comme un inhibiteur pour la locomotion une fois que le mâle a été en contact avec la femelle.
31Pour finir, Hemptinne et al. (1996) avancent le fait que les mâles de la coccinelle à deux points reconnaissent les femelles grâce à la présence d’une phéromone sécrétée par des glandes situées sur ou dans les élytres, mais cette phéromone ne serait détectée que par contact.
5.2. Phéromones d’agrégation
32En automne, de nombreuses espèces de coccinelles migrent pour former des amas d’agrégation (Majerus, 1994). Les coccinelles se dirigent le plus souvent vers des caractéristiques proéminentes du paysage, tels les affleurements de colline (comme H. convergens) ou les troncs d’arbres (comme C. maculata) mais il apparait que les agrégats ont tendance à se retrouver aux mêmes sites d’une année à l’autre (Hagen, 1962). Ceci suggère l’intervention de phéromones dans le comportement agrégatif des coccinelles. Il est avancé que les alkylméthoxypyrazines, en plus de leur rôle d’avertissement envers les prédateurs, constituent également des composants essentiels de la phéromone d’agrégation (al Abassi et al., 1998). Le 2-Isopropyl-3-methoxypyrazine a été identifié chez C. septempunctata, H. axyridis et H. convergens et des essais réalisés en laboratoire ont mis en évidence le pouvoir attractif et immobilisant de ce composé sur les coccinelles à sept points adultes (al Abassi et al., 1998 ; Cudjoe et al., 2005). Le caractère persistant des alkylméthoxypyrazines et la présence des mêmes pyrazines chez diverses espèces de coccinelles (Moore et al., 1990) confortent l’idée que ces composés sont impliqués dans la formation des agrégats de coccinelles aux mêmes endroits année après année et que fréquemment, plusieurs espèces de coccinelles se retrouvent dans le même agrégat (Roach et al., 1991 ; Majerus, 1994). D’autres composés pourraient également être impliqués dans le comportement agrégatif des coccinelles car les femelles de la coccinelle asiatique, H. axyridis, arrivent en premier sur les sites d’agrégation (Nalepa et al., 1996 ; 2000) et les femelles de cette espèce produisent des sesquiterpènes qui ne sont pas détectables chez des individus collectés au printemps (Aldrich, 1999). Lorsque Brown et al. (2006) ont réalisé des prélèvements d’odeurs sur des individus H. axyridis élevés en laboratoire sous des conditions automnales, ils ont constaté que les femelles émettaient du (-)-β-caryophyllène. Par la suite, le pouvoir attractif du (-)-β-caryophyllène a été démontré à la fois pour les mâles et pour les femelles chez H. axyridis (Alhmedi et al., 2007 ; Verheggen et al., 2007). Il se pourrait donc que ce composé soit impliqué dans le phénomène d’agrégation de cette coccinelle asiatique en automne.
5.3. Phéromones d’anti-oviposition (ODP)
33Les femelles de certaines espèces de coccinelles, telles que A. bipunctata (Hemptinne et al., 1992 ; Doumbia et al., 1998), Aphidecta obliterata (L.) (Oliver et al., 2006), C. sexmaculata (Ruzicka, 2006), C. septempunctata (Ruzicka, 1997 ; 2001 ; 2002 ; Doumbia et al., 1998), Cryptolaemus montrouzieri Mulsant (Merlin et al., 1996), Cycloneda limbifer Casey (Ruzicka, 2001), E. flaviventris (le Rü et al., 2001), H. axyridis (Yasuda et al., 2000), Semiadalia undecimnotata Schneider (Ruzicka, 2001 ; 2002), évitent de pondre aux endroits où des larves conspécifiques sont présentes et réduisent de ce fait les risques de cannibalisme et de compétition pour la nourriture. Ce phénomène s’explique par la présence, dans les traces larvaires des coccinelles, d’une phéromone d’anti-oviposition spécifique à l’espèce. Cette phéromone est déposée via le disque anal présent sur le dixième segment abdominal des larves (Laubertie et al., 2006) et est détectée par les femelles adultes grâce aux chémorécepteurs de contact présents sur les palpes maxillaires de ces dernières (Ruzicka, 2003). Une étude, réalisée par Hemptinne et al. (2001), a révélé que la phéromone d’anti-oviposition d’A. bipunctata est constituée d’un mélange d’alcanes similaire à celui retrouvé sur la surface des œufs conspécifiques. Ces alcanes se dispersent facilement sur la cuticule hydrophobe des plantes. Ceci permet au message laissé d’avoir une plus grande portée. Qui plus est, ces alcanes ne s’oxydent pas rapidement, ils sont donc stables et persistent dans l’environnement. En effet, la durée de vie du signal peut atteindre un mois pour certaines espèces de coccinelles, comme pour C. limbifer (Ruzicka, 2002).
34Cette phéromone peut aussi influencer le comportement d’oviposition d’autres espèces de coccinelles. En effet, les femelles C. sexmaculata évitent de pondre aux endroits où des traces larvaires de C. limbifer et de Ceratomegilla undecimnotata (Schneider) sont présentes (Ruzicka, 2006). De même, C. limbifer dépose un plus faible nombre d’œufs sur des sites précédemment exposés à des larves de S. undecimnotata et de Leis dimidiata Mulsant ; S. undecimnotata présente un plus faible taux d’oviposition en présence de traces larvaires de L. dimidiata (Ruzicka, 2001). Magro et al. (2007) ont étudié l’impact mutuel des traces larvaires chez trois espèces de coccinelles : A. bipunctata, Adalia decempunctata L. et C. septempunctata. Il apparait que les femelles d’A. bipunctata et d’A. decempunctata évitent de pondre en présence de traces larvaires de chaque espèce. En revanche, les femelles de C. septempunctata n’évitent de pondre que lors de la présence de traces larvaires conspécifiques. Ceci s’explique par le fait que C. septempunctata attaque les colonies de pucerons plus tard que les deux autres espèces considérées (Smith, 1966). Il est donc possible que C. septempunctata éprouve plus de difficultés à trouver une colonie de pucerons non exploitée. Le cannibalisme ou la prédation intraguilde pourraient dès lors être moins couteux en termes de fitness que ne pas déposer d’œufs du tout.
35En plus de détecter les traces larvaires de coccinelles conspécifiques ou hétérospécifiques, les coccinelles sont capables d’utiliser les traces larvaires laissées par d’autres compétiteurs. En effet, C. septempunctata dépose moins d’œufs sur des sites exposés à des larves de Chrysopa oculata Say (Ruzicka, 2001 ; 2002) et Chrysopa perla L. (Ruzicka, 2001).
36Inversement, des insectes ne faisant pas partie de la famille des Coccinellidae peuvent utiliser les traces larvaires de coccinelles afin de réduire le risque de prédation. En effet, Takizawa et al. (2000) ont démontré que le parasitisme dû à Aphidius colemani Viereck était réduit en présence de traces larvaires de C. septempunctata. Étant donné que les pucerons parasités sont également soumis à la prédation par les coccinelles, il est avantageux pour les femelles parasitoïdes d’éviter de pondre dans des sites où ces prédateurs seraient présents.
37Ruzicka et al. (2008) ont démontré que les traces larvaires de coccinelles pouvaient également influencer le comportement des larves conspécifiques. En effet, ils ont mis en évidence, grâce à une expérience réalisée en choix double, que les larves C. limbifer de 4e stade passaient plus de temps dans la zone dépourvue de traces de larves conspécifiques que dans celle où ces traces étaient présentes. La phéromone d’anti-oviposition serait donc aussi à l’origine d’une distribution active des larves. Ceci permet d’améliorer la distribution initiale de la descendance parmi les sites d’alimentation et de ce fait, d’augmenter les chances de survie de la progéniture.
38Agarwala et al. (2003) ont démontré que les excréments peuvent également servir de facteur d’évaluation de la qualité d’un site en termes de risque de prédation. En effet, ils ont remarqué que les femelles gravides de Propylea japonica (Thunberg) présentent un taux plus faible d’alimentation et d’oviposition en présence de fèces conspécifiques et de fèces d’H. axyridis. En revanche, les femelles gravides d’H. axyridis ne présentent une réduction de l’alimentation et de l’oviposition qu’en présence de fèces conspécifiques. Ceci s’explique par le fait qu’H. axyridis est un prédateur généraliste, alors que P. japonica est plus spécialiste et que ce dernier a beaucoup plus de risques d’être la proie intraguilde lors d’une rencontre avec la coccinelle asiatique.
6. Conclusion
39Les exemples donnés ci-dessus illustrent la capacité des coccinelles à interagir avec les molécules présentes dans leur environnement et ce, aussi bien pour localiser leurs proies, s’accoupler, se protéger de la prédation ou du cannibalisme, trouver un abri pour passer l’hiver ou assurer la meilleure survie pour leur descendance. Une excellente compréhension des mécanismes expliquant les comportements des coccinelles envers ces molécules sémiochimiques pourrait permettre l’utilisation de ces composés dans des méthodes de lutte intégrée. En effet, des pièges à base de ces volatils pourraient attirer les coccinelles aux endroits infestés par leurs proies (pucerons, cochenilles, etc.) tout en les éloignant des lieux où sa présence est indésirable (au sein des habitations en hiver par exemple). Mais avant la mise en place de telles méthodes de lutte, l’effet des molécules sur les autres organismes présents dans la nature devra être étudié.
Bibliographie
Acar E.B., Medina J.C., Lee M.L. & Booth G.M., 2001. Olfactory behaviour of convergent lady beetles (Coleoptera: Coccinellidae) to alarm pheromone of green peach aphid (Hemiptera: Aphididae). Can. Entomol., 133(3), 389-397.
Agarwala B.K. & Dixon A.F.G., 1991. Cannibalism and interspecific predation in ladybirds. In: Polgar L., Chambers R.J., Dixon A.F.G. & Hodek I., eds. Behaviour and impact of aphidophaga. The Hague, The Netherlands: SPB Academic Publishing, 95-102.
Agarwala B.K., Bhattacharya S. & Bardhanroy P., 1998. Who eats whose eggs? Intra-versus inter-specific interactions in starving ladybird beetles predaceous on aphids. Ethology Ecol. Evol., 10, 361-368.
Agarwala B.K. & Yasuda H., 2001. Overlapping oviposition and chemical defense of eggs in two co-occurring species of ladybird predators of aphids. J. Ethology, 19, 47-53.
Agarwala B.K., Yasuda H. & Kajita Y., 2003. Effect of conspecific and heterospecific feces on foraging and oviposition of two predatory ladybirds: role of fecal cues in predator avoidance. J. Chem. Ecol., 29(2), 357-376.
al Abassi S. et al., 1998. Ladybird beetle odour identified and found to be responsible for attraction between adults. Cell. Mol. Life Sci., 54, 876-879.
al Abassi S. et al., 2000. Response of the seven-spot ladybird to an aphid alarm pheromone and an alarm pheromone inhibitor is mediated by paired olfactory cells. J. Chem. Ecol., 26(7), 1765-1771.
al Abassi S. et al., 2001. Response of the ladybird parasitoid Dinocampus coccinellae to toxic alkaloids from the seven-spot ladybird, Coccinella septempunctata. J. Chem. Ecol., 27(1), 33-43.
Alam N. et al., 2002. A new alkaloid from two coccinellid beetles Harmonia axyridis and Aiolocaria hexaspilota. Bull. Korean Chem. Soc., 23, 497-499.
Aldrich J.R., 1999. Part II. Beneficials: predators. In: Hardie J. & Minks A.K., eds. Pheromones of non-lepidopteran insects associated with agricultural plants. Oxon, UK: CAB International, 357-381.
Alhmedi A., Francis F. & Haubruge E., 2007. Olfactory responses of the multicolored Asian lady beetle, Harmonia axyridis to aphid and host plant volatile releases. In: Proceedings of the 23rd International Society of Chemical Ecology, 22-26 July, Jena, Germany, 232.
Attygalle A.B. et al., 1993a. Azamacrolides: a family of alkaloids from the pupal defensive secretion of a ladybird beetle (Epilachna varivestris). Proc. Natl Acad. Sci. USA, 90, 5204-5208.
Attygalle A.B., Xu S.-C., McCormick K.D. & Meinwald J., 1993b. Alkaloids of the Mexican bean beetle, Epilachna varivestris (Coccinellidae). Tetrahedron, 49(41), 9333-9342.
Ayer W.A., Bennett M.J., Browne L.M. & Purdham J.T., 1976. Defensive substances of Coccinella transversoguttata and Hippodamia caseyi, ladybugs indigenous to western Canada. Can. J. Chem. Ecol., 54, 1807-1813.
Ayer W.A. & Browne L.M., 1977. The ladybug alkaloids including synthesis and biosynthesis. Heterocycles, 7, 1.
Bezzerides A.L., McGraw K.J., Parker R.S. & Husseini J., 2007. Elytra color as a signal defense in the Asian ladybird beetle Harmonia axyridis. Behav. Ecol. Sociobiol., 61, 1401-1408.
Bhatkar A.P., 1982. Orientation and defense of ladybeetles (Coleoptera, Coccinellidae) following ant trail in search of aphids. Folia Entomol. Mex., 53, 75-85.
Birkett M.A. et al., 2000. New roles for cis-jasmone as an insect semiochemical and in plant defense. Proc. Natl Acad. Sci. USA, 97(16), 9329-9334.
Braconnier M.F., Braekman J.C., Daloze D. & Pasteels J.M., 1985. (Z)-1,17-diaminooctadec-9-ene, a novel aliphatic diamine from Coccinellidae. Experientia, 41, 519-520.
Braekman J.-C. et al., 1999. New piperidine alkaloids from two ladybird beetles of the genus Calvia (Coccinellidae). Eur. J. Org. Chem., 7, 1749-1755.
Brossut R., 1996. Phéromones. La communication chimique chez les animaux. Paris : CNRS Éditions, 143.
Brown W.V. & Moore B.P., 1982. The defensive alkaloids of Cryptolaemus montrouzieri (Coleoptera: Coccinellidae). Aust. J. Chem., 35, 1255-1261.
Brown A.E., Riddick E.W., Aldrich J.R. & Holmes W.E., 2006. Identification of (-)-β-caryophyllene as a gender-specific terpene produced by the multicoloured Asian lady beetle. J. Chem. Ecol., 32, 2489-2499.
Cai L., Koziel J.A. & O’Neal M.E., 2007. Determination of characteristic odorants from Harmonia axyridis beetles using in vivo solid-phase microextraction and multidimentional gas chromatography-mass spectrometry-olfactometry. J. Chromatogr., 1147, 66-78.
Camarano S., González A. & Rossini C., 2006. Chemical defense of the ladybird beetle Epilachna paenulata. Chemoecology, 16, 179-184.
Carter M.C. & Dixon A.F.G., 1984. Honeydew: an arrestant stimulus for coccinellids. Ecol. Entomol., 9, 383-387.
Cudjoe E., Wiederkehr T.B. & Brindle I.D., 2005. Headspace gas chromatography-mass spectrometry: a fast approach to the identification and determination of 2-alkyl-3-methoxypyrazine pheromones in ladybugs. Analyst, 130, 152-155.
Daloze D., Braekman J.C. & Pasteels J.M., 1994. Ladybird defense alkaloids: structural, chemotaxonomic and biosynthetic aspects (Col.: Coccinellidae). Chemoecology, 5, 3/4, 173-183.
Dawson G.W. et al., 1984. Natural inhibition of aphid alarm pheromone. Entomol. Exp. Appl., 36, 197-199.
Dawson G.W. et al., 1989. The aphid sex pheromone. Pure Appl. Chem., 61, 555-558.
Dixon A.F.G. & Agarwala B.K., 1999. Ladybird-induced life-history changes in aphids. Proc. R. Soc. London B, 266, 1549-1553.
Doumbia M., Hemptinne J.-L. & Dixon A.F.G., 1998. Assessment of patch quality by ladybirds: role of larval tracks. Oecologia, 113, 197-202.
Eisner T. et al., 1986. Defensive alkaloid in the blood of Mexican bean beetle (Epilachna varivestis). Experientia, 42, 204-207.
Eisner T. et al., 1994. Defensive use of acquired substance (carminic acid) by predaceous insect larvae. Experientia, 50, 610-615.
Enders D. & Bartzen D., 1991. Enantioselective total synthesis of harmonine, a defense alkaloid of ladybugs (Coleoptera: Coccinellidae). Liebigs Ann. Chem., 6, 659-674.
Evans E.W. & Richards D.R., 1997. Managing the dispersal of ladybird beetles (Col.: Coccinellidae): use of artificial honeydew to manipulate spatial distributions. Entomophaga, 42, 93-102.
Francis F., Haubruge E., Hastir P. & Gaspar C., 2001. Effect of aphid host plant on development and reproduction of the third trophic level, the predator Adalia bipunctata (Coleoptera, Coccinellidae). Environ. Entomol., 30, 947-952.
Francis F., Lognay G. & Haubruge E., 2004. Olfactory responses to aphid and host plant volatile releases: (E)-β-farnesene an effective kairomone for the predator Adalia bipunctata. J. Chem. Ecol., 30(4), 741-755.
Francis F. et al., 2005. Is the (E)-β-farnesene only volatile terpenoid in aphids? J. Appl. Entomol., 129(1), 6-11.
Giles K.L. et al., 2002. Host plants affect predator fitness via the nutritional value of herbivore prey: investigation of a plant-aphid-ladybeetle system. Biocontrol, 47, 1-21.
Girling R.D. & Hassall M., 2008. Behavioural responses of the seven-spot ladybird Coccinella septempunctata to plant headspace chemicals collected from four crop Brassicas and Arabidopsis thaliana, infested with Myzus persicae. Agric. For. Entomol., 10, 297-306.
Greany P.D. & Hagen K.S., 1981. Prey selection. In: Nordlund D.A., Jones R.L. & Lewis W.J., eds. Semiochemicals: their role in pest control. New York, USA: John Wiley & Sons, 121-135.
Guilford T., Nicol C., Rothschild M. & Moore B.P., 1987. The biological roles of pyrazines: evidence for a warning odour function. Biol. J. Linnean Soc., 31, 113-128.
Hagen K.S., 1962. Biology and ecology of predaceous Coccinellidae. Annu. Rev. Entomol., 7, 289-326.
Hamilton R.M., Dogan E.B., Schaalje G.B. & Booth G.M., 1999. Olfactory response of the lady beetle Hippodamia convergens (Coleoptera: Coccinellidae) to prey related odors, including a scanning electron microscopy study of the antennal sensilla. Environ. Entomol., 28(5), 812-822.
Han B.-Y. & Chen Z.-M., 2002a. Composition of the volatiles from intact and tea aphid-damaged tea shoots and their allurement to several natural enemies of the tea aphid. J. Appl. Entomol., 126, 497-500.
Han B.-Y. & Chen Z.-M., 2002b. Behavioral and electrophysiological responses of natural enemies to synomones from tea shoots and kairomones from tea aphids, Toxoptera aurantii. J. Chem. Ecol., 28(11), 2203-2219.
Han B.Y. & Chen Z.M., 2002c. Composition of the volatiles from intact and mechanically pierced tea aphid-tea shoot complexes and their attraction to natural enemies of the tea aphid. J. Agric. Food Chem., 50, 2571-2575.
Heidari M. & Copland M.J.W., 1993. Honeydew: a food resource or arrestant for the mealybug predator Cryptolaemus montrouzieri? Entomophaga, 38(1), 63-68.
Heit G., Castresana J., Puhl L. & Mareggiani G., 2005. Role of volatiles emitted by cultivated and wild Solanaceae in foraging behaviour of coccinellid predators. IDESIA, 23(3), 13-19.
Hemptinne J.-L., Dixon A.F.G. & Coffin J., 1992. Attack strategy of ladybird beetles (Coccinellidae): factors shaping their numerical response. Oecologia, 90, 238-245.
Hemptinne J.-L., Dixon A.F.G. & Lognay G., 1996. Searching behaviour and mate recognition by males of the two-spot ladybird beetle, Adalia bipunctata. Ecol. Entomol., 21, 165-170.
Hemptinne J.-L., Lognay G. & Dixon A.F.G., 1998. Mate recognition in the two-spot ladybird beetle, Adalia bipunctata: role of chemical and behavioural cues. J. Insect Physiol., 44, 1163-1171.
Hemptinne J.-L., Lognay G., Gauthier C. & Dixon A.F.G., 2000a. Role of surface chemical signals in egg cannibalism and intraguild predation in ladybirds (Coleoptera: Coccinellidae). Chemoecology, 10(3), 123-128.
Hemptinne J.-L., Gaudin M., Dixon A.F.G. & Lognay G., 2000b. Social feeding in ladybird beetles: adaptive significance and mechanism. Chemoecology, 10(3), 149-152.
Hemptinne J.-L., Lognay G., Doumbia M. & Dixon A.F.G., 2001. Chemical nature and persistence of the oviposition deterring pheromone in the tracks of the larvae of the two spot ladybird, Adalia bipunctata (Coleoptera: Coccinellidae). Chemoecology, 11(1), 43-47.
Henson R.D. et al., 1975. Identification of precoccinellin in the ladybird beetle, Coleomegilla maculata. Experientia, 31, 145.
Hodek I. & Honek A., 1996. Ecology of Coccinellidae. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Huang Q. et al., 1998. Chilocorine C: a new "dimeric" alkaloid from a coccinellid beetle, Chilocorus cacti. J. Nat. Prod., 61, 598-601.
Ide T., Suzuki N. & Katayama N., 2007. The use of honeydew in foraging for aphids by larvae of the ladybird beetle, Coccinella septempunctata L. (Coleoptera: Coccinellidae). Ecol. Entomol., 32, 455-460.
James D.G., 2005. Further field evaluation of synthetic herbivore-induced plant volatiles as attractants for beneficial insects. J. Chem. Ecol., 31, 481-495.
Jiggins C., Majerus M.E.N. & Gough U., 1993. Ant defense of colonies of Aphis fabae Scopoli (Hemiptera: Aphididae), against predation by ladybirds. Brit. J. Entomol. Nat. Hist., 6, 129-138.
Kazana E. et al., 2007. The cabbage aphid: a walking mustard oil bomb. Proc. R. Soc. London B, 274, 2271-2277.
King G.K. & Meinwald J., 1996. Review of the defensive chemistry of Coccinellids. Chem. Rev., 96, 1105-1122.
Kunert G. & Weisser W.W., 2005. The importance of antennae for pea aphid wing induction in the presence of natural enemies. Bull. Entomol. Res., 95(2), 125-131.
Laubertie E. et al., 2006. The immediate source of the oviposition-deterring pheromone produced by larvae of Adalia bipunctata (L.) (Coleoptera: Coccinellidae). J. Insect Behav., 19(2), 231-240.
Laurent P., Braekman J.-C., Daloze D. & Pasteels J.-M., 2002. Chilocorine D, a novel heptacyclic alkaloid from a coccinellid beetle (Chilocorus renipustulatus). Tetrahedron Lett., 43, 7465-7467.
Laurent P., Braekman J.-C. & Daloze D., 2005. Insect chemical defense. Top. Curr. Chem., 240, 167-229.
Lebrun B., Braekman J.-C., Daloze D. & Pasteels J.-M., 1997. 2-dehydrococcinelline, a new defensive alkaloid from the ladybird beetle Anatis ocellata (Coccinellidae). J. Nat. Prod., 60, 1148-1149.
Lebrun B. et al., 1998. Isopsylloborine A, a new dimeric azaphenalene alkaloid from ladybird beetles (Coleoptera: Coccinellidae). Tetrahedron Lett., 40, 8115-8116.
Lebrun B. et al., 2001. Hyperaspine, a new 3-oxaquinolizidine alkaloid from Hyperaspis campestris (Coleoptera: Coccinellidae). Tetrahedron Lett., 42, 4621-4623.
le Rü B. & Makaya Makosso J.P., 2001. Prey habitat location by the Cassava Mealybug predator Exochomus flaviventris: olfactory responses to odor of plant, mealybug, plant-mealybug complex, and plant-mealybg-natural enemy complex. J. Insect Behav., 14(5), 557-572.
Lognay G. et al., 1996. Adalinine, a new piperidine alkaloid from the ladybird beetles Adalia bipunctata and Adalia decempunctata. J. Nat. Prod., 59, 510-511.
McCormick K.D. et al., 1994. Chilocorine: heptacyclic alkaloid from a coccinellid beetle. Tetrahedron, 50, 2365-2372.
Magro A. et al., 2007. Assessment of patch quality by ladybirds: relative response to conspecific and heterospecific larval tracks a consequence of habitat similarity? Chemoecology, 17, 37-45.
Majerus M.E.N., 1994. Ladybirds. London: Harper Collins Publishers.
Majerus M.E.N., Sloggett J.J., Godeau J.-F. & Hemptinne J.-L., 2007. Interactions between ants and aphidophagous and coccidophagous ladybirds. Popul. Ecol., 49, 15-27.
Malcolm S.B., 1990. Chemical defence in chewing and sucking insect herbivores: plant-derived cardenolides in the monarch butterfly and oleander aphid. Chemoecology, 1, 12-21.
Malcolm S.B., 1992. Prey defence and predator foraging. In: Crawley M.J., ed. The population biology of predators, parasites and diseases. Oxford, UK: Blackwell Scientific Publications, 458-475.
Marples N.M., 1993. Is the alkaloid in 2-spot ladybirds (Adalia bipunctata) a defence against ant predation? Chemoecology, 4(1), 29-32.
Marples N.M., Brakefield P.M. & Cowie R.J., 1989. Differences between the 7-spot and 2-spot ladybird beetles (Coccinellidae) in their toxic effects on a bird predator. Ecol. Entomol., 14, 79-84.
Martos A., Givovich A. & Niemeyer H.M., 1992. Effect of DIMBOA, an aphid resistance factor in wheat, on the aphid predator Eriopis connexa Germar (Coleoptera: Coccinellidae). J. Chem. Ecol., 18, 469-479.
Merlin J., Lemaitre O. & Grégoire J.-C., 1996. Chemical cues produced by conspecific larvae deter oviposition by the coccidophagous ladybird beetle, Cryptolaemus montrouzieri. Entomol. Exp. Appl., 79, 147-151.
Michaud J.P., 2002. Invasion of the Florida citrus ecosystem by Harmonia axyridis (Coleoptera: Coccinellidae) and asymmetric competition with a native species, Cycloneda sanguinea. Environ. Entomol., 31, 827-835.
Moore B.P., Brown W.V. & Rothschild M., 1990. Methylakylpyrazines in aposematic insects, their hostplants and mimics. Chemoecology, 1, 43-51.
Murray R.D.H., Martin A. & Stride G.O., 1972. Identification of the volatile phagostimulants in Solanum campylacanthum for Epilachna fulvosignata. J. Insect Physiol., 18, 2369-2373.
Nakashima Y. et al., 2004. The role of semiochemicals in the avoidance of the seven-spot ladybird, Coccinella septempunctata, by the aphid parasitoid, Aphidius ervi. J. Chem. Ecol., 30, 1103-1116.
Nakashima Y., Birkett M.A., Pye B.J. & Powell W., 2006. Chemically mediated intraguild predator avoidance by aphid parasitoids: interspecific variability in sensitivity to semiochemical trails of ladybird predators. J. Chem. Ecol., 32(9), 1989-1998.
Nalepa C.A., Kidd K.A. & Ahlstrom K.R., 1996. Biology of Harmonia axyridis (Coleoptera: Coccinellidae) in winter aggregations. Ann. Entomol. Soc. Am., 89, 681-685.
Nalepa C.A., Kidd K.A. & Hopkins D.I., 2000. The multicoloured Asian lady beetle: orientation to aggregation sites. J. Entomol. Sci., 35, 150-157.
Ninkovic V., al Abassi S. & Pettersson J., 2001. The influence of aphid-induced plant volatiles on ladybird beetle searching behavior. Biol. Control, 21, 191-195.
Ninkovic V. & Pettersson J., 2003. Searching behaviour of the sevenspotted ladybird, Coccinella septempunctata: effects of plant-plant odour interaction. OIKOS, 100, 65-70.
Obata S., 1986. Mechanisms of prey finding in the aphidophagous ladybird beetle, Harmonia axyridis (Coleoptera: Coccinellidae). Entomophaga, 31, 303-311.
Oliver T.H., Timms J.E.L., Taylor A. & Leather S.R., 2006. Oviposition responses to patch quality in the larch ladybird Aphidecta obliterata (Coleoptera: Coccinellidae): effects of aphid density, and con- and heterospecific tracks. Bull. Entomol. Res., 96(1), 25-34.
Omkar, Pervez A. & Gupta A.K., 2004. Role of surface chemicals in egg cannibalism and intraguild predation by neonates of two aphidophagous ladybirds, Propylea dissecta and Coccinella transversalis. J. Appl. Entomol., 128(9/10), 691-695.
Omkar & Pervez A., 2005. Mating behavior of an aphidophagous ladybird beetle, Propylea dissecta (Mulsant). Insect Sci., 12, 37-44.
Pasteels J.-M. et al., 1973. Distribution et activité des alcaloïdes défensifs des Coccinellidae. J. Insect Physiol., 19, 1771-1784.
Pennacchio F., 1989. The Italian species of the genus Aphidius Nees (Hymenoptera, Braconidae, Aphidiinae). Boll. Lab. Entomol. Agrar. “Fillippo Silvestri.”, 46, 75-106.
Ponsonby D.J. & Copland M.J.W., 1995. Olfactory responses by the scale insect predator Chilocorus nigritus (F.) (Coleoptera: Coccinellidae). Biocontrol Sci. Technol., 5, 83-93.
Radford P. et al., 1997. Pyrrolidinoöxazolidine alkaloids from two species of ladybird beetles. J. Nat. Prod., 60, 755-759.
Richards A.M., 1980. Rapid publication note. Sexual selection, guarding and sexual conflict in a species of coccinellidae (Coleoptera). Aust. J. Entomol., 19, 26.
Riddick E.W., Dively G. & Barbosa P., 1998. Effect of seed-mix deployment of Cry3A-trangenic and nontransgenic potato on the abundance of Lebia grandis (Coleoptera: Carabidae) and Coleomegilla maculata (Coleoptera: Coccinellidae). Ann. Entomol. Soc. Am., 91, 647-653.
Roach S.H. & Thomas W.M., 1991. Overwintering and spring emergence of three coccinellid species in the coastal plain of South Carolina. Environ. Entomol., 20, 540-544.
Rothschild M., von Euw J. & Reichstein T., 1970. Cardiac glycosides in the oleander aphid Aphis nerii. J. Insect Physiol., 16, 1141-1145.
Rothschild M., von Euw J. & Reichstein T., 1973. Cardiac glycosides in a scale insect (Aspidiotus), a ladybird (Coccinella) and a lacewing (Chrysopa). J. Entomol. (A), 48, 89-90.
Ruzicka Z., 1997. Recognition of oviposition-deterring allomones by aphidophagous predators (Neuroptera: Chrysopidae, Coleoptera: Coccinellidae). Eur. J. Entomol., 94, 431-434.
Ruzicka Z., 2001. Oviposition responses of aphidophagous coccinellids to tracks of ladybird (Coleoptera: Coccinellidae) and lacewing (Neuroptera: Chrysopidae) larvae. Eur. J. Entomol., 98(2), 183-188.
Ruzicka Z., 2002. Persistence of deterrent larval tracks in Coccinella septempunctata, Cycloneda limbifer and Semiadalia undecimnotata (Coleoptera: Coccinellidae). Eur. J. Entomol., 99(4), 471-475.
Ruzicka Z., 2003. Perception of oviposition-deterring larval tracks in aphidophagous coccinellids Cycloneda limbifer and Ceratomegilla undecimnotata (Coleoptera: Coccinellidae). Eur. J. Entomol., 100(3), 345-350.
Ruzicka Z., 2006. Oviposition-deterring effects of conspecific and heterospecific larval tracks on Cheilomenes sexmaculata (Coleoptera: Coccinellidae). Eur. J. Entomol., 103(4), 757-763.
Ruzicka Z. & Zemek R., 2008. Deterrent effects of larval tracks on conspecific larvae in Cycloneda limbifer. Biocontrol, 53(5), 763-771.
Schröder F.C. & Tolasch T., 1998. Psylloborine A, a new dimeric alkaloid from a ladybird beetle. Tetrahedron, 54, 12243-12248.
Sengonca C. & Liu B., 1994. Responses of the different instar predator, Coccinella septempunctata L. (Coleoptera: Coccinellidae), to the kairomones produced by the prey and non-prey insects as well as the predator itself. J. Plant Dis. Protect., 101, 173-177.
Shi X. et al., 1995. Spirocyclic defensive alkaloid from a coccinellid beetle. Tetrahedron, 51, 8711-8718.
Smith B.D., 1966. Effects of parasites and predators on a natural population of the aphid Acyrthosiphon spartii (Koch) and broom (Sarothamnus scoparius L.). J. Appl. Ecol., 35, 255-267.
Takizawa T., Yasuda H. & Agarwala B.K., 2000. Effect of three species of predatory ladybirds on oviposition of aphid parasitoids. Entomol. Sci., 3(3), 465-469.
Timmermans M. et al., 1992. Exochomine, a dimeric ladybird alkaloid, isolated from Exochomus quadripustulatus (Coleoptera: Coccinellidae). Tetrahedron Lett., 33, 1281-1284.
Tursch B. et al., 1971. Coccinellin, the defensive alkaloid of the beetle Coccinella septempunctata. Chimia, 25, 307.
Tursch B., Daloze D. & Hootele C., 1972. The alkaloid of Propylaea quatuordecimpunctata L. (Coleoptera: Coccinellidae). Chimia, 26, 74.
Tursch B. et al., 1973. Chemical ecology of arthropods. VI. Adaline, a novel alkaloid from Adalia bipunctata L. (Coleoptera, Coccinellidae). Tetrahedron Lett., 14, 201-202.
Tursch B. et al., 1974. Chemical ecology of arthropods. IX. Structure and absolute configuration of hippodamine and convergine, two novel alkaloids from the American ladybug Hippodamia convergens (Coleoptera - Coccinellidae). Tetrahedron Lett., 15, 409-412.
Tursch B. et al., 1975. Chemical ecology of arthropods. X. The structure of myrrhine and the biosynthesis of coccinelline. Tetrahedron, 31, 1541-1543.
Van den Meiracker R.A.F., Hammond W.N.O. & van Alphen J.M., 1990. The role of kairomones in prey finding by Diomus sp. and Exochomus sp., two coccinellid predators of the cassava mealybug, Phenacoccus manihoti. Entomol. Exp. Appl., 56, 209-217.
Verheggen F.J. et al., 2007. Electrophysiological and behavioral responses of the multicolored Asian lady beetle, Harmonia axyridis Pallas, to sesquiterpenes semiochemicals. J. Chem. Ecol., 33, 2148-2155.
Wang S.F., Braekman J.-C., Daloze D. & Pasteels J.-M., 1996a. Signatipennine: a new alkaloid from the New Guinean ladybird Epilachna signatipennis (Coccinellidae). Bull. Soc. Chim. Belges, 105, 483-487.
Wang S.F. et al., 1996b. N-alpha-quinaldyl-L-arginine-HC1, a new alkaloid from Subcoccinella 24-punctata (Coleoptera: Coccinellidae). Experientia, 52, 628-630.
Ware R.L. et al., 2008. Chemical protection of Calvia quatuordecimguttata eggs against intraguild predation by the invasive ladybird Harmonia axyridis. In: Roy H.E. & Wajnberg E., eds. Biological control to invasion: the ladybird Harmonia axyridis as a model species. Dordrecht, The Netherlands: Springer, 189-200.
Wiedemann R., Woodring J., Volkl W. & Hoffmann K.H., 2004. Amino acid composition of honeydew from aphid species feeding on tansy, Tanacetum vulgare. Mitt. Dtsch. Ges. Allg. Angew. Entomol., 14, 459-462.
Witte L., Ehmke A. & Hartmann Th., 1990. Interspecific flow of pyrrolizidine alkaloids. From plants via aphids to ladybirds. Naturwissenschaften, 77, 540-543.
Yasuda H., Takagi T. & Kogi K., 2000. Effects of conspecific and heterospecific larval tracks on the oviposition behaviour of the predatory ladybird, Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol., 97, 551-553.
Zhu J. et al., 1999. Olfactory reactions of the twelve-spotted lady beetle, Coleomegilla maculata and the green lacewing, Chrysoperla carnea to semiochemicals released from their prey and host plant: electroantennogram and behavioral responses. J. Chem. Ecol., 25(5), 1163-1177.
Zhu J. & Park K.-C., 2005. Methyl salicylate, a soybean aphid-induced plant volatile attractive to the predator Coccinella septempunctata. J. Chem. Ecol., 31(8), 1733-1746.
Para citar este artículo
Acerca de: Delphine Durieux
Univ. Liège - Gembloux Agro-Bio Tech. Unité d’Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : entomologie.gembloux@ulg.ac.be
Acerca de: François J. Verheggen
Univ. Liège - Gembloux Agro-Bio Tech. Unité d’Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
Acerca de: Axel Vandereycken
Univ. Liège - Gembloux Agro-Bio Tech. Unité d’Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
Acerca de: Emilie Joie
Univ. Liège - Gembloux Agro-Bio Tech. Unité d’Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
Acerca de: Éric Haubruge
Univ. Liège - Gembloux Agro-Bio Tech. Unité d’Entomologie fonctionnelle et évolutive. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
