


- Home
- Volume 14 (2010)
- numéro 3
- Influence d'une infection expérimentale à Trypanosoma congolense sur la fonction sexuelle des béliers Djallonké et Sahéliens en zone subhumide


View(s): 1866 (10 ULiège)
Download(s): 82 (1 ULiège)

Influence d'une infection expérimentale à Trypanosoma congolense sur la fonction sexuelle des béliers Djallonké et Sahéliens en zone subhumide

Editor's Notes
Reçu le 6 novembre 2009, accepté le 2 février 2010
Résumé
Pour mesurer l'effet des trypanosomoses animales africaines (TAA) sur la fonction de reproduction mâle, huit béliers Djallonké (DI : 31,2 ± 2,95 kg) et Sahéliens (SI : 41,7 ± 4,64 kg) de 18-24 mois ont été infectés par injection intraveineuse de 104 parasites Trypanosoma congolense. Huit autres béliers Djallonké (DT) et Sahéliens (ST) de poids moyens et d'âges similaires à ceux de leurs congénères infectés ont été utilisés comme témoins. Après quatre semaines, les animaux ont été traités mais maintenus sous observation. L'évolution des paramètres cliniques montre que les Djallonké ont plus d'aptitude que les Sahéliens à contrôler les effets des TAA. Cette différence de sensibilité est confirmée par les pertes de poids plus élevées des SI (- 31,3 g) par rapport aux DI (- 12,8 g) et aux témoins. L'atteinte de la fonction sexuelle par l'infection commence par la baisse de la libido des SI (33 s) par rapport aux DI (22,1 s) (P > 0,05). Les résultats indiquent un effet significatif de la race sur le volume de l'éjaculat (Sahéliens : 1,23 ml vs Djallonké : 0,88 ml ; P < 0,05). Les autres paramètres spermatiques sont peu affectés par la race. Le sperme des DT avait une concentration en spermatozoïdes (3 293,4 x 106.ml-1), des taux de spermatozoïdes anormaux (15,3 %), morts (13,9 %), mobiles (75,5 %), des motilités individuelle (3,76) et massale (3,94) similaires (P > 0,05) à ceux des ST (respectivement, 3 481,7 x 106.ml-1 ; 17,3 % ; 12,4 % ; 74,9 % ; 3,77 ; 3,36). Les ST ont produit 26 % plus de spermatozoïdes totaux (P > 0,05) que les DT. L'infection a réduit le volume de sperme par éjaculat de 15,8 et 14,5 % (P > 0,05), la production de spermatozoïdes totaux de 18,2 et 13,3 % (P > 0,05) et le taux de spermatozoïdes mobiles de 14,2 et 27,9 % (P < 0,05) respectivement chez les Djallonké et les Sahéliens. Ces résultats indiquent un dysfonctionnement testiculaire dû à l'effet néfaste de l'infection, d'où l'augmentation des taux de spermatozoïdes morts de 69,8 et 74,3 % (P < 0,05) et de spermatozoïdes anormaux de 23,6 et 24,1 % (P > 0,05) respectivement chez les DI et les SI. Les effets néfastes, souvent plus prononcés chez les Sahéliens, pourraient être le signe de leur plus grande sensibilité vis-à-vis des TAA par rapport aux Djallonké, d'où la mort d'un SI deux semaines après l'infection, un cas d'azoospermie six semaines après la chimiothérapie et l'absence de parasites dans le sang.
Abstract
Influence of an experimental Trypanosoma congolense infection on the reproductive function of Djallonké and Sahelian rams in subhumid zone. To measure the effect of African animal trypanosomosis (AAT) on male reproductive function, eight 18-24 month old Djallonké (ID; 31.2 ± 2.95 kg) and Sahelian (IS; 41.7 ± 4.64 kg) rams were infected with 104 Trypanosoma congolense. Eight other Djallonké (CD) and Sahelian (CS) rams with similar bodyweight and ages were used as uninfected controls. Four weeks after infection, ID and IS were treated with a trypanocidal drug, while maintained under observation. The evolution of clinical parameters shows a higher aptitude of Djallonké than Sahelian to control the effects of infection. This interbreed difference in susceptibility to AAT was confirmed by the higher weight losses in IS (-31.3 g) when compared to ID (-12.8 g) as to CD and CS rams. The effect of a T. congolense infection consisted in a decreased libido in IS (33 s) when compared to ID (22.1 s) (P > 0.05). The results indicate a significant effect of breed on ejaculate volume (CS: 1.23 ml vs CD: 0.88 ml; P < 0.05). Otherwise, breed did not significantly affect the other spermatic parameters. CD sperm cells concentration (3,293.4 x 106.ml-1), total abnormality rate (15.3%), dead sperm cells rate (13.9%), mobile sperm cells rate (75.5%), individual (3.76) and mass motilities (3.94) did not differ (P > 0.05) from those of CS (respectively 3,481.7 x 106.ml-1; 17.3%; 12.4%; 74.9%; 3.77; 3.36). However, the later produced 26% more total sperm cells than CD (P > 0.05). The infection reduced ejaculate volume by 15.8 and 14.5%, (P > 0.05), production of total sperm cells by 18.2 and 13.3% (P > 0.05) and the rate of mobile sperm cells by 14.2 and 27.9% (P < 0.05) respectively for Djallonké and Sahelian rams. The results show a dysfunctional state of testicles due to the harmful effects of AAT infection, hence the increase in dead sperm cells by 69.8 and 74.3% (P > 0.05) and in the rate of total abnormal sperm cells by 23.6 and 24.1% (P > 0.05) respectively for ID and IS rams when compared respectively to CD and CS animals. Harmful effects often more pronounced in Sahelian breeds during this experiment could indicate a higher susceptibility of this breed to AAT in comparison with Djallonké. One Sahelian ram died two weeks post-infection and azoospermia was observed in another one six weeks after chemotherapy and the disappearance of parasites from the bloodstream.
Table of content
1. Introduction
1Depuis le début des cycles de sècheresse des années 1970-1980 au Sahel, les troupeaux originaires de cette zone effectuent des séjours de plus en plus prolongés et/ou définitifs dans les zones de savane humide infestées de glossines, principaux vecteurs des trypanosomoses animales africaines (TAA). Or, plusieurs études (Murray et al., 1982 ; Osaer et al., 1994 ; 1997) ont montré que seules les races indigènes (ovins et caprins Djallonké, bovins ndama, etc.) réputées trypanotolérantes ont une aptitude à y survivre, se reproduire et maintenir un niveau appréciable de productivité. La présence d'animaux dits trypanosensibles (Touré et al., 1981 ; Bengaly et al., 1993) dans ces zones endémiques de TAA engendre un contexte épidémiologique susceptible d'influencer les caractéristiques physiologiques, telles que la fonction de reproduction et par conséquent, la productivité aussi bien des races introduites que des races autochtones.
2Chez les Djallonké réputés trypanotolérants (Touré et al., 1981 ; Bengaly et al., 1993 ; Osaer et al., 1994 ; Goossens et al., 1997), l'effet des TAA sur la fonction de reproduction peut varier d'une simple baisse du taux plasmatique de testostérone (Adeyemo et al., 1989) entrainant une baisse de la libido (Osaer et al., 1997) à de sévères lésions testiculaires (Ikede, 1979 ; Anosa et al., 1980) pouvant entrainer la baisse de la concentration du sperme en cellules vivantes, des taux élevés d'anomalies spermatiques ou même l'oligo- et l'azoospermie (Agu et al., 1986 ; Sekoni et al., 1990 ; Sekoni, 1993), qui peuvent persister 12 semaines après un traitement au chlorure d'homidium (Sekoni et al., 1991). Ces effets semblent être fonction de la nature du trypanosome et de la durée de l'infection (Osaer et al., 1997).
3Les informations relatives à l'effet des TAA sur la fonction sexuelle des races du Sahel sont limitées, probablement en raison de leur forte sensibilité entrainant une morbidité précocement invalidante (Sekoni et al., 2004) ou une mortalité au cours des stades précoces de l'infection.
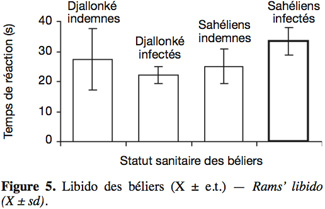
4Cette expérience a pour objectif de mesurer l'effet de l'infection à T. congolense sur la fonction de reproduction des béliers Djallonké et ceux du Sahel et ce, en comparant les animaux indemnes et infectés expérimentalement pendant une courte période.
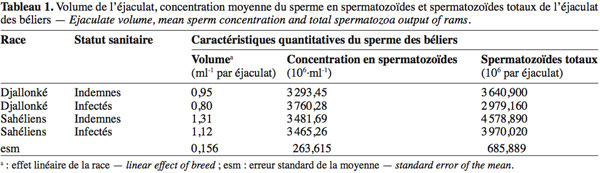
2. Matériel et méthodes
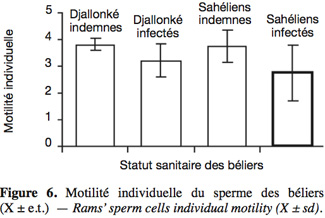
2.1. Site
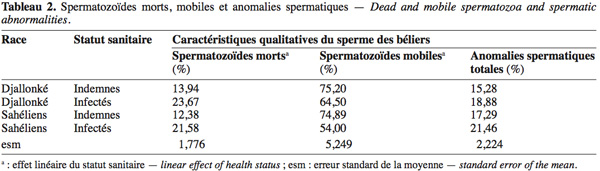
5L'étude a eu lieu entre janvier et avril à l'étable sous-moustiquaire du CIRDES à Bobo-Dioulasso (11°06' N, 04°20' O) situé dans le Sud-Ouest du Burkina Faso en Afrique de l'Ouest. Le climat est de type tropical subhumide, avec environ 1 025 mm de pluie à régime monomodal d'avril à octobre. Les températures minimales et maximales moyennes varient respectivement entre 23 °C en janvier et 31 °C en mai.
2.2. Animaux, traitements et conduite du troupeau
6Les observations ont porté sur seize béliers Djallonké (PV = 31,2 ± 2,95 kg) et Sahéliens (PV = 41,7 ± 4,64 kg) âgés de 18 à 24 mois au début des essais. Ils ont été achetés respectivement sur les marchés de la région de Bobo-Dioulasso (zone subhumide) et autour de Dori et Gorom Gorom (zone sahélienne). Dès leur arrivée à la station, ils ont été bouclés, vaccinés contre la peste des petits ruminants et la pasteurellose, traités avec un complexe antibiotique (tétracycline) + vitamines, traités contre les ectoparasites (fluméthrine), les endoparasites (fenbendazole, ivermectine) et les TAA (acéturate de diminazène). Ensuite, ils ont été mis en observation pendant plus de 15 semaines. Au cours de cette période de quarantaine, les béliers ont été entrainés à la collecte de sperme au vagin artificiel, en présence d'une brebis de même race. Ils étaient pesés toutes les deux semaines avant le début des essais et une fois par semaine en période d'expérience.
7Deux semaines avant le début de l'essai, les animaux ont été adaptés à leur régime alimentaire distribué individuellement, constitué de paille de riz, de sels minéraux et d'eau, offerts ad libitum, de coques de coton et de tourteaux de coton, offerts en quantités limitées. Ce régime leur permettait d'atteindre un niveau d'alimentation de 1,15 à 1,20, avec un gain moyen quotidien (GMQ) de plus de 20 g. Au cours de la semaine (J-7 à J-0) qui a précédé le début de l'essai, le sang prélevé quotidiennement sur chaque animal et analysé (technique du buffy cost selon Murray et al., 1977) a permis de s'assurer qu'ils étaient indemnes de TAA (absence de parasites sanguins circulants) et d'établir les paramètres sanguins de base. La température rectale a été prise quotidiennement à partir d'une semaine avant jusqu'à une semaine après le début de l'essai. Les 16 béliers ont été équitablement repartis entre quatre traitements consistant en une combinaison factorielle de deux races d'ovin (Djallonké et sahélienne) et deux statuts sanitaires (animaux infectés de TAA ou non).
8Le premier jour du début de l'essai (J-0), un lot de béliers Djallonké (DI) et un lot de béliers du Sahel (SI) ont été infectés artificiellement par injection intraveineuse de 104 parasites de l'espèce Trypanosoma congolense à partir de sang provenant de souris infectées (T. congolense souche Karankasso 83/CRTA/57) après immunodépression. Deux autres lots, un de béliers Djallonké (DT) et un de béliers du Sahel (ST), ont été utilisés comme témoins non infectés.
9Après quatre semaines de suivi des paramètres de santé (température rectale, hématocrite, parasitémie), de la croissance pondérale et de mesure de certains paramètres de la fonction sexuelle (paramètres spermatiques, endocriniens et libido), les animaux ont été traités avec un trypanocide curatif (acéturate de diminazène). Certaines observations cliniques (parasitémie et température), la pesée des animaux et le suivi des paramètres spermatiques ont continué pendant dix semaines après le traitement curatif.
2.3. Mesures
10Après l'infection, le sang de tous les animaux a été prélevé quotidiennement jusqu'à la détection des parasites, puis trois fois par semaine pour l'appréciation du degré d'anémie et de la parasitémie. Le degré d'anémie a été estimé par la mesure de l'hématocrite (%), tandis que la parasitémie a été déterminée par la technique d'examen microscopique du buffy cost (Murray et al., 1977). La température rectale a été mesurée quotidiennement avant la détection des parasites dans le sang périphérique, puis tous les trois jours après, à l'aide d'un thermomètre de précision.
11La croissance pondérale des animaux a été déterminée par double pesée (à jeun) 48 h avant l'infection, puis hebdomadairement jusqu'à la fin de l'essai.
12La libido (temps de réaction) de tous les animaux a été évaluée par la mesure du temps écoulé entre la présentation de la brebis et la première monte. Les paramètres spermatiques ont été évalués à partir du sperme récolté à intervalle de 48 h à l'aide d'un vagin artificiel en présence d'une brebis multipare. Le volume de l'éjaculat était lu directement dans le tube de collecte gradué au demi-millilitre près. La motilité massale était déterminée immédiatement après la collecte par l'observation d'une goutte de sperme non diluée déposée sur une lame placée sur la platine (37-38 °C) d'un microscope (x 100). Ce paramètre était évalué selon une échelle allant de zéro (aucune vague) à cinq (vagues déferlantes). La motilité individuelle et le taux de spermatozoïdes mobiles ont été déterminés à partir du sperme dilué au 1/200 (0,05 ml + 9,95 ml) avec du sérum physiologique formolé (NaCl 9 ‰ + formol 1 %). La motilité individuelle a été évaluée par observation de cette préparation au microscope à un grossissement de 200, en utilisant une échelle allant de 0 (spermatozoïdes inertes) à 5 (spermatozoïdes fléchants, à mouvements rectilignes). Le pourcentage de spermatozoïdes mobiles a été déterminé en même temps que la motilité individuelle en examinant au microscope dans plusieurs champs, les spermatozoïdes placides et ceux en mouvement.
13Juste après la récolte, un frottis de sperme dilué coloré à l'éosine-nigrosine était conservé à sec pour la détermination ultérieure du taux de spermatozoïdes morts et des anomalies spermatiques (anomalies spermatiques majeures et mineures de la tête, de la pièce intermédiaire et de la queue) par comptage d'environ 200 cellules dans plusieurs champs. La concentration du sperme en spermatozoïdes a été déterminée à partir du sperme dilué (au 1/401e) avec du sérum physiologique formolé (0,05 ml + 20 ml) par comptage des spermatozoïdes à l'aide d'une cellule de Malassez.
2.4. Analyses statistiques
14La procédure GLM de SAS (1997) a été utilisée pour déterminer les effets de l'infection à T. congolense et de la race sur la croissance pondérale et les paramètres de la fonction sexuelle des béliers dans les conditions environnementales du site de l'étude.
3. Résultats
3.1. Observations cliniques
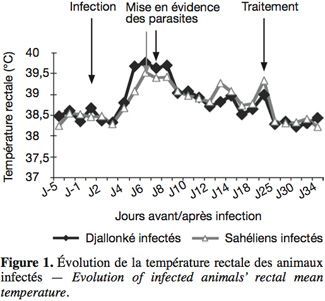
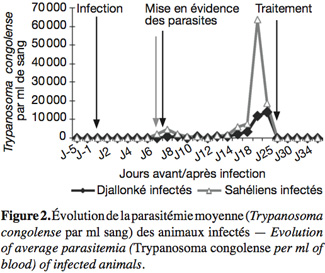
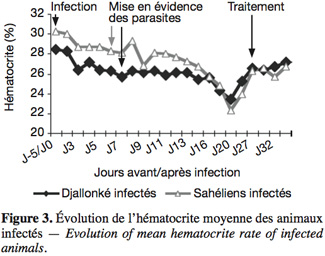
15La température rectale moyenne a culminé chez les béliers infectés des deux races après la mise en évidence des parasites dans le sang périphérique (Figure 1), pour ensuite évoluer en dents de scie avant de devenir normale moins d'une semaine après le traitement. Les parasites ont été mis en évidence chez les béliers du Sahel au 6e jour après l'infection et chez les Djallonké au 7e, soit 24 h plus tard (Figure 2). Les Djallonké ont semblé mieux contrôler la parasitémie qui a culminé à 1,4 x 104 trypanosomes.ml-1 de sang au début de la 3e semaine d'infection. Chez les Sahéliens, elle a augmenté graduellement (2 x 103 à 6 x 103 trypanosomes) jusqu'à la fin de la 2e semaine, puis de façon plus rapide pour atteindre 6,3 x 104 trypanosomes.ml-1 sang au milieu de la 3e semaine. Les parasites ont disparu du sang périphérique de tous les animaux moins d'une semaine après le traitement. La baisse du taux d'hématocrite a débuté chez tous les animaux dès le lendemain de l'infection (Figure 3). Elle a été plus importante chez les Sahéliens (-26 %) que chez les Djallonké (-18 %). L'hématocrite des Djallonké avait presque atteint son niveau normal au bout d'une semaine, tandis qu'au même moment celui des Sahéliens était encore de 12 % inférieur à son taux d'avant infection.
16Un bélier sahélien a succombé à la fin de la deuxième semaine d'infection avec une parasitémie de 2 x 106 trypanosomes.ml-1 de sang, alors que la température rectale était de 40,5 °C et le PCV de 10 points (-32 %) en dessous du taux avant infection.
3.2. Croissance pondérale
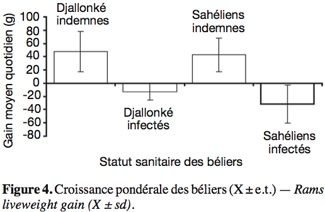
17L'infection a significativement réduit la croissance pondérale des animaux toutes races confondues (P < 0,05). Au cours de l'expérience, les Djallonké infectés perdaient quotidiennement 12,8 g, tandis que les témoins non infectés gagnaient 48,4 g contre respectivement -31,3 g et 43,4 g chez les béliers Sahéliens infectés et indemnes (Figure 4). La race des béliers ou l'interaction entre race et infection trypanosomienne n'ont eu aucune influence significative (P > 0,05) sur le GMQ.
3.3. Libido
18Les temps de réaction à la présence de brebis ont varié entre 22,1 s (Djallonké infectés) et 33,3 s (Sahéliens infectés) (Figure 5). La race n'a eu aucun effet significatif sur ce paramètre (Djallonké 24,7 s vs Sahéliens 29,2 s). Dans l'ensemble, l'infection a significativement réduit la libido des béliers Sahéliens uniquement.
3.4. Caractéristiques spermatiques
19Les béliers Sahéliens, aussi bien infectés qu'indemnes, ont produit significativement plus de sperme par éjaculat que les Djallonké (Tableau 1). L'infection a réduit le volume de sperme produit par éjaculat de 15,8 % et 14,5 % respectivement chez les béliers Djallonké infectés et chez les béliers Sahéliens infectés par rapport à leurs congénères indemnes (P > 0,05).
20Seule l'infection a significativement affecté la motilité individuelle (Figure 6) (P < 0,05) et la motilité massale (Figure 7) des béliers. Bien que l'interaction entre race et infection n'était pas significative, les béliers Sahéliens ont paru plus sensibles à l'infection que les Djallonké pour la motilité individuelle (2,74 vs 3,22 ; P > 0,05). Pour la motilité massale, la différence entre Djallonké infectés et Sahéliens infectés (3,94 vs 3,36) n'était pas significative (P > 0,05).

21Ni la race, ni l'infection n'ont significativement affecté la concentration en spermatozoïdes du sperme des béliers (Tableau 1) au cours de la période expérimentale (avant l'infection et les quatre semaines post-infection). Cependant, bien après l'arrêt de l'infection par l'administration d'un trypanocide curatif et malgré l'absence de parasites dans le sang jugulaire, la concentration en spermatozoïdes d'un des béliers Sahéliens a continué à baisser, jusqu'à une azoospermie totale et définitive à partir de la 6e semaine après le traitement, soit 10 semaines après l'infection. Les données du tableau 1 montrent que les béliers Sahéliens produisent plus de spermatozoïdes totaux que les béliers Djallonké (P > 0,05). L'infection a réduit (P > 0,05) la production de spermatozoïdes totaux de 21,2 % et 15,3 % respectivement chez les béliers Djallonké et Sahéliens infectés par rapport à leurs congénères indemnes. L'infection a significativement augmenté la proportion de spermatozoïdes morts et réduit le taux de spermatozoïdes mobiles chez les deux races (P < 0,05) (Tableau 2). Chez les animaux indemnes, le taux d'anomalies totales a été de 13,1 % plus élevé (P > 0,05) chez les béliers Sahéliens par rapport aux Djallonké. Cette même tendance a été observée entre les animaux infectés des deux races (P > 0,05) (Tableau 2).
4. Discussion
22Les Djallonké ont montré des signes cliniques d'infection trypanosomienne plus modérés que les Sahéliens. En outre, l'évolution des paramètres cliniques et leur temps de retour à la normale (niveau avant infection) montrent que les Djallonké ont une plus grande aptitude que les Sahéliens à contrôler la parasitémie et les effets de l'infection. Cette différence de sensibilité a été confirmée par les cas de mortalité enregistrés chez les Sahéliens infectés ainsi que la perte de poids chez ces derniers par rapport aux Djallonké infectés, aux Sahéliens indemnes et aux Djallonké indemnes (P < 0,05). Ce meilleur comportement des Djallonké par rapport aux Sahéliens vis-à-vis de l'infection trypanosomienne pourrait s'expliquer par une résistance acquise ou innée (Touré et al., 1981 ; Bengaly et al., 1993). La différence non significative entre la croissance pondérale des béliers infectés des deux races s'explique par le comportement hétérogène des Sahéliens face à l'infection.
23L'atteinte de la fonction sexuelle par l'infection à T. congolense commence par la baisse de la libido des Sahéliens infectés par rapport aux béliers des autres groupes (P > 0,05). Ceci pourrait s'expliquer par des facteurs raciaux, notamment la plus grande sensibilité des Sahéliens aux effets de l'infection par T. congolense, pouvant se traduire par une perturbation du profil hormonal dont la baisse du taux de testostérone plasmatique (Adeyemo et al., 1989 ; Boly et al., 1991 ; Mutayoba et al., 1993 ; Al-Qarawi et al., 2004), facteur déterminant du comportement sexuel.
24L'absence d'effet significatif de la TAA sur le volume de l'éjaculat malgré une différence de plus de 15 % entre animaux indemnes et infectés pourrait s'expliquer non seulement par l'hétérogénéité du comportement des Sahéliens, mais surtout par la durée de la période d'infection trop brève pour entrainer l'inhibition de cette fonction. Ceci est confirmé par les résultats de Sekoni (1992 ; 1994) rapportant un arrêt de la production de semence chez les mâles de plusieurs espèces et particulièrement chez des béliers Yankassa au bout de 15 semaines d'infection à Trypanosoma vivax. Les mêmes auteurs (Sekoni et al., 2004) ont rapporté des cas d'oligo- et d'azoospermie chez des métis zébu Bunaji x Friesian au bout de 12 semaines d'infection à T. vivax ou à T. congolense. Cette anomalie pourrait s'expliquer par l'arrêt de la production et l'épuisement de la réserve de sperme dans l'épididyme (Sekoni et al., 1990 ; Adamu et al., 2006), suite à un dysfonctionnement testiculaire et celui d'autres glandes impliquées dans la production de sperme et ceci, en relation avec les effets de l'infection (anémie, hyperthermie, parasitémie, etc.). La baisse significative des motilités massales et individuelles et le faible taux de spermatozoïdes mobiles chez les béliers infectés pourraient être le signe d'une altération de certains stades critiques de la spermatogenèse résultant de l'élévation de la température testiculaire (Baril et al., 1993) suite à l'hyperthermie provoquée par l'infection trypanosomienne. Ces présomptions sont confirmées par les taux élevés de spermatozoïdes anormaux (P > 0,05) et de spermatozoïdes morts (P < 0,05) dans les spermes des béliers infectés par rapport aux témoins et chez les béliers du Sahel infectés par rapport aux Djallonké infectés. Les écarts entre ces paramètres spermatiques enregistrés chez les animaux infectés presque exclusivement en défaveur des races du Sahel, indiquent la plus grande sensibilité de ces derniers par rapport aux Djallonké. L'effet limité de l'infection sur le taux d'anomalies spermatiques totales (18 à 22 %) par rapport aux taux rapportés par Agu et al. (1986) et Sekoni (1992 ; 1993) sur des béliers infectés avec T. vivax (52 % à 100 %) d'une part et Sekoni et al. (1991) sur des zébus infectés avec T. vivax ou T. congolense (100 %) d'autre part, s'explique par la courte durée de l'infection au cours de la présente expérience.
25L'azoospermie observée chez un bélier du Sahel au cours de la présente expérience, en dépit du traitement des animaux, ne semble pas être un cas isolé et indique que les effets de l'infection ou de tout autre facteur sur les paramètres spermatiques peuvent être décalés de quelques jours à quelques semaines. Sekoni et al. (1991) ont également observé une détérioration significative des paramètres spermatiques chez des zébus infectés avec T. vivax ou T. congolense 12 semaines après la chimiothérapie en dépit de la disparition des parasites du courant sanguin. De même, Sekoni (1992) et Sekoni et al. (2004) ont observé une oligospermie suivie d'une azoospermie respectivement au bout de trois semaines d'infection à T. vivax chez des béliers Yankassa et au bout de six semaines d'infection à T. vivax ou à T. congolense chez des taureaux zébu Bunaji x Friesian, qui sont toutes des races originaires des zones exemptes de glossines.
5. Conclusion
26En dépit de la brièveté de la période de suivi, les résultats montrent que les béliers Djallonké ont une meilleure aptitude à contrôler les effets de l'infection à T. congolense sur les fonctions de reproduction par rapport aux béliers Sahéliens et qu'une infection prolongée pourrait affecter au moins temporairement l'aptitude de ces derniers à la reproduction. Un suivi plus prolongé aussi bien en milieu contrôlé qu'en milieu réel durant les différentes saisons de l'année permettrait de mieux caractériser la fonction de reproduction des races locales dans les conditions de leurs différents milieux d'élevage.
Bibliographie
Adamu S. et al., 2006. Sequential testicular and epididymal damage in Zebu bulls experimentally infected with Trypanosoma vivax. Vet. Parasit., 143(1), 29-34.
Adeyemo O., Oyejide A. & Agbedana O., 1989. Plasma testosterone in Trypanosoma brucei-infected West African Dwarf rams. Anim. Reprod. Sci., 22, 21-26.
Agu W.E., Ige K. & Olatunde D.S., 1986. Evaluation of semen quality of rams infected with Trypanosoma vivax. Anim. Reprod. Sci., 11, 123-127.
Al-Qarawi A.A. et al., 2004. Trypanosomiasis-induced infertility in dromedary (Camelus dromedarius) bulls: changes in plasma steroids concentration and semen characteristics. Anim. Reprod. Sci., 84(1-2), 73-82.
Anosa V.O. & Isoun T.T., 1980. Further observations on the testicular pathology in Trypanosoma vivax infection of sheep and goats. Res. Vet. Sci., 28, 151-160.
Baril G. et al., 1993. Manuel de formation pour l'insémination artificielle chez les ovins et les caprins. Études FAO Production et Santé Animales n°83. Rome : FAO.
Bengaly Z. et al., 1993. Comparaison de la trypanosomose expérimentale chez certaines races de petits ruminants au Burkina faso. Rev. Élev. Méd. Vét. Pays Trop., 46(4), 563-570.
Boly H., Thiombiano D., Humblot P. & Thibier M., 1991. Influence de Trypanosoma congolense sur la fonction sexuelle de taurins Baoulé. Rev. Élev. Méd. Vét. Pays Trop., 44(4), 475-480.
Goossens B., Osaer S. & Kora S., 1997. Long-term effects of an experimental infection with T. congolense on reproductive performance of trypanotolerant Djallonké ewes and West African Dwarf does. Res. Vet. Sci., 63, 169-173.
Ikede B.O., 1979. Genital lesions in experimental chronic Trypanosoma brucei infection in rams. Res. Vet. Sci., 26, 145-151.
Murray M., Murray P.K. & McIntyre W.I.M., 1977. An improved parasitological technique for diagnosis of African trypanosomiasis. Trans. R. Soc. Trop. Med. Hyg., 71, 325-326.
Murray M., Morrison W.L. & Whitelaw D.D., 1982. Host susceptibility to African trypanosomiasis: trypanotolerance. Adv. Parasitol., 21, 1-68.
Mutayoba B.M. et al., 1993. Concentrations of plasma luteinizing hormone and testosterone and response to injection of gonadotrophin-releasing hormone in sheep Trypanosome congolense infection. In: Proceedings of 22nd Meeting ISC-TRC Kampala, Uganda.
Osaer S. et al., 1994. A comparison of the susceptibility of Djallonké sheep and West African Dwarf goats to experimental infection with different strains of T. congolense. Vet. Parasitol., 5, 191-204.
Osaer S., Goossens B., Sauveroche B. & Dempfle L., 1997. Evaluation of the semen quality and reproductive performance of trypanotolerant Djallonke rams following an artificial infection with Trypanosoma congolense. Small Ruminant Res., 24, 213-222.
SAS (Statistical Analysis System), 1997. SAS/STAT for personal computers. Cary, NC, USA: SAS Institute.
Sekoni V.O., Saror D.I., Njoku C.O. & Kumi-Diaka J., 1990. Evaluation of morphological abnormalities of spermatozoa in the semen of Zebu bulls consequent to Trypanosoma vivax and Trypanosoma congolense infections. Theriogenology, 33(4), 925-936.
Sekoni V.O., Saror D.I., Njoku C.O. & Kumi-Diaka J., 1991. Effect of novidium (homidium chloride) chemotherapy on elevated spermatozoa morphological abnormalities in the semen of Zebu bulls infected with Trypanosoma vivax and Trypanosoma congolense. Anim. Reprod. Sci., 24(3-4), 249-258.
Sekoni V.O., 1992. Effect of Trypanosoma vivax infection on semen characteristics of Yankasa rams. British Vet. J., 148(6), 501-506.
Sekoni V.O., 1993. Elevated sperm morphological abnormalities of Yankasa rams consequent to Trypanosoma vivax infection. Anim. Reprod. Sci., 31(3-4), 243-248.
Sekoni V.O., 1994. Reproductive disorders caused by animal trypanozomiases: a review. Theriogenology, 42(4), 557-570.
Sekoni V.O., Rekwot P.I. & Bawa E.K., 2004. Effects of Trypanosoma congolense infections on the reaction time and semen characteristics of Zebu (Bunaji) x Friesian crossbred bulls. Theriogenology, 61(1), 55-62.
Touré S.M., Seye M., Mbengue M. & Dieye T., 1981. Trypanotolerance studies of comparative pathology of Djallonké sheep and Sahelian Fulani sheep. Dakar: ISRA/LNERV, 1981.
To cite this article
About: Mamadou Sangaré
Centre International de Recherche-Développement sur l'Élevage en zone Subhumide (CIRDES). 01 BP 454. Bobo-Dioulasso (Burkina Faso). E-mail : mamadousangare@hotmail.com
About: Zakaria Bengaly
Centre International de Recherche-Développement sur l'Élevage en zone Subhumide (CIRDES). 01 BP 454. Bobo-Dioulasso (Burkina Faso).
About: Hamani Marichatou
Université Abdou Moumouni (UAM). BP 10662. Niamey (Niger).
About: Aboubacar Toguyeni
Centre International de Recherche-Développement sur l'Élevage en zone Subhumide (CIRDES). 01 BP 454. Bobo-Dioulasso (Burkina Faso).
About: Hamidou Hamadou Tamboura
Institut de l'Environnement et des Recherches Agricoles (INERA). 04 BP 8645. Ouagadougou 04 (Burkina Faso).
