


- Portada
- Volume 15 (2011)
- numéro 3
- The use of semiochemical slow-release devices in integrated pest management strategies


Vista(s): 16091 (110 ULiège)
Descargar(s): 1475 (30 ULiège)

The use of semiochemical slow-release devices in integrated pest management strategies

Notes de la rédaction
Received on July 30, 2010; accepted on January 11, 2011
Résumé
L’utilisation de systèmes à libération lente de sémiochimiques dans les stratégies de lutte intégrée. Le développement des stratégies de lutte intégrée est en croissance depuis que de nombreux problèmes sont apparus suite à l’utilisation abusive et non raisonnée des pesticides de synthèse. Les sémiochimiques (molécules informatives utilisées dans les interactions insecte-insecte ou plante-insecte) sont de plus en plus considérés, au sein des stratégies de lutte intégrée, comme des approches alternatives ou complémentaires aux traitements insecticides. En effet, ces composés, spécifiques à chaque espèce, ne présentent pas d’effets négatifs relatés dans la littérature envers les organismes bénéfiques et n’engendrent aucun risque de résistance chez les insectes ravageurs comme observés avec les insecticides. En raison de leur activité biologique complexe mais aussi de leur risque de dégradation par les rayons ultraviolets ou à l’oxygène de l’air, leur dispersion dans l’environnement nécessite l’élaboration de systèmes garantissant une libération lente et contrôlée des composés volatils actifs. Plusieurs études ont été menées afin d’estimer le taux de libération de systèmes commercialisés ou mis au point en laboratoire. L’influence des paramètres climatiques et du type de diffuseur a été estimée par plusieurs autres auteurs afin de fournir des indications sur la longévité des diffuseurs sur terrain. La présente revue analyse et critique une liste d’études de systèmes à libération lente.
Abstract
The development of integrated pest management (IPM) strategies is increasing since many problems appeared with the use of synthetic pesticides. Semiochemicals – informative molecules used in insect-insect or plant-insect interaction – are more and more considered within IPM strategies as alternative or complementary approach to insecticide treatments. Indeed, these species-specific compounds do not present any related adversely affectation of beneficial organisms and do not generate any risk of pest insect resistance as observed with insecticides. Because of their complex biological activity, their dispersion in the environment to be protected or monitored needs the elaboration of slow-release devices ensuring a controlled release of the biologically active volatile compounds. These sensitive molecules also need to be protected from degradation by UV light and oxygen. Many studies were conducted on estimation of release-rate from commercialized or experimental slow-release devices. The influence of climatic parameters and dispenser type were estimated by previous authors in order to provide indications about the on-field longevity of lures. The present review outlines a list of slow-release studies conducted by many authors followed by a critical analysis of these studies.
Tabla de contenidos
1. Introduction
1During the seventies and the eighties, environmental and social side effects of synthetic pesticides led to the development of integrated pest management (IPM) programs in the USA and Asia. Since then, many IPM strategies have been successful worldwide. Indeed, the overuse of insecticides presents many drawbacks like the appearance of insect resistances, environmental concerns, and risks for human health. Moreover, the action of pesticides is generally non species-specific with the risk of disturbing the natural ecological equilibrium (Witzgall, 2001).
2IPM implies various strategies, which ideally have to be combined at different levels. In 1998, Kogan defined IPM as “a decision support system for the selection and the use of pest control tactics, singly or harmoniously coordinated into a management strategy, based on cost/benefit analyses that take into account the interests of and the impacts on producers, society and the environment”.
3The efficiency of these approaches needs an interdisciplinary collaboration between agronomists, entomologists, chemists having an experience in pest behaviors, technologists, and finally the crop producers. It is particularly true when the IPM tactic implies the use of insect semiochemical slow-release devices as tools to modify the behavior of insect pests. Indeed, the release systems must be economical, effective, environmentally safe without harmful side effects, and field-tested to prove the efficiency towards targeted insects before legal authorization and commercialization. The validation of all these manufacturing steps is not possible without the interaction of multi-disciplinary fields of knowledge.
4On an historical point of view, the role of sexual pheromones in insect mating was demonstrated in the late 19th century. The characterization of the first insect sex pheromone was established in 1959 (Butenandt et al., 1959) and was isolated from female Bombyx mori (Lepidoptera). This technological overhang led, in the mid-seventies, to an increase of commercial activities in synthesis of semiochemicals previously identified as potential agents for controlling pests. This was the first step to replace synthetic insecticides with pheromone products (Cork, 2004). In the same time, the research on insect chemical communication grew up and led to the emergence of a new scientific discipline: the chemical ecology. In 1971, Edward Wilson gave a definition of the chemical communication: “this is the emission of a stimulus by one individual and which induces a reaction in another one, the reaction being beneficial to the emitter, to the receptor or both”. In parallel, the gas chromatography appeared in chemistry and brought simplicity in identification of volatile molecules. Rapidly, the economical interest for using pheromone compounds in pest controls was updated and included in integrated pest management programs (Brossut, 1997).
5The present review deals with the development of the major approaches to control pests by using semiochemical (chemical communication signal) slow-release devices. Furthermore, the authors will focus on the techniques implemented in the study of release rates and on the estimation of on-field longevity of semiochemical devices.
2. Semiochemicals
2.1. Definitions
6Semiochemicals, from semeion (in Greek) or signal, can be defined as chemicals emitted by living organisms (plants, insects, etc.) that induce a behavioral or a physiological response in other individuals. These compounds can be classified in two groups considering whether they act as intraspecific (pheromones) or interspecific (allelochemicals) mediators. Allelochemicals include allomones (emitting species benefits), kairomones (receptor species benefits) and synomones (both species benefit) (Figure 1). However, a single chemical signal may act as both as pheromone and allelochemical.
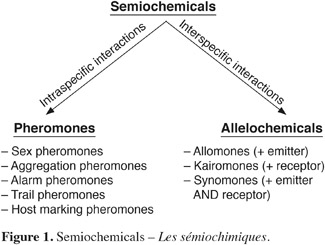
7There are different types of pheromones according to the response they induce on the perceiving individuals. The most common are presented hereafter (Brossut, 1997; Cork, 2004):
8– Sex pheromones are generally produced by females of a species in order to attract males of the same species for mating. Some exceptions exist where male butterflies (e.g. Bicyclus anynana) produce sex pheromones to seduce females during the courtship (Nieberding et al., 2008). Sex pheromones consist in individual molecules or specific blend of compounds in a given ratio. The most studied, and used in IPM, sex pheromones are that emitted by Lepidoptera;
9– Aggregation pheromones are released by one gender of a species to attract individuals (both sexes) of the same species in order to exploit a specific resource (food, appropriate mating site, etc.). They are mainly emitted by Coleopterous species;
10– Alarm pheromones alert conspecifics in case of threats. Generally the response behavior results in dispersion of congeners. These pheromones, characteristic of social or gregarious insects, occur in some important insect pests including Aphididae and Thripidae. This class of pheromones has potential in IPM (Verheggen et al., 2010);
11– Trail pheromones are present in social colonies to indicate the trail to be followed when some scout insects locate food resource. Walking insects, like ants, typically produce these pheromones;
12– Host marking pheromones reduce the competition between members of the same species, like it is observed in parasitoids that mark a host in which they have laid an egg.
2.2. Chemistry and properties of semiochemicals
13Pheromones and semiochemicals in general, consist in a wide range of organic molecules which could be volatile or non-volatile. Non-volatile semiochemicals include cuticular hydrocarbons, acting in mate recognition or in cannibalism regulation of several insect species. Wilson et al. (1963) suggested that the volatile pheromones naturally exploited in insect communication have between 5 and 20 atoms of carbon with molecular weights ranging from 80 to 300. Those having a molecular weight above 300 are not sufficiently volatile to allow a communication at long distance. Cork (2004), in his Pheromone manual, cites the major pheromones identified in moths and butterflies according to their chemical classes.
14The biosynthesis of such semiochemical molecules is supposed to come from the food. They are generally synthesized de novo by excreting cells. The biosynthesis of sexual pheromones is well known in Lepidoptera and Diptera. In both cases, the pheromones consist in long carbon chains (alcohols, aldehydes and acetates for Lepidoptera; hydrocarbons having high molecular weight for Diptera) derived from the metabolism of fatty acids (Brossut, 1997).
15The efficiency of semiochemical substances in chemical communication depends on various physical properties including chemical nature, volatility, solubility and lifetime of the molecules in the environment. An important abiotic factor controlling the effectiveness of the pheromones is the temperature which increases the diffusion of the molecules in the air. The stability of these volatile compounds also affects the efficiency in IPM.
3. IPM strategies using semiochemicals
16There are many benefits to formulate semiochemical substances in integrated pest management outline. These molecules are naturally occurring and are generally environmentally friendly. Additionally, in IPM strategies the compounds are generally used at concentrations close to those found in nature and, due to their high volatility, they can act at long distances and dissipate rapidly. The risk to human health and environment is also reduced compared to pesticides. For all these reasons, semiochemicals are compounds of potentially high interest in IPM.
3.1. IPM strategies
17Various strategies exist depending on the goals and scopes to achieve. Some of them are described hereafter.
18Monitoring. Monitoring of insect populations has generally three purposes: to detect the presence of invasive pests; to estimate the relative density of a pest population at a specific site; to indicate the first emergence or peak flight activity of a pest species in a given area. The appropriate control actions (e.g. local insecticide treatment) can then be carried out (Weinzierl et al., 2005).
19Trapping. Trapping with pheromone lures is a mechanical control action that consists in removing large number of pests in an area after monitoring step. The traps can be used simultaneously with a killing substance (“lure and kill” strategy) which has the benefit of not being in direct contact with the crop. This technique is also useful in stored-product pest control (Phillips, 1997).
20Mating disruption. The technique of mating disruption by using species-specific sex pheromones in large quantity is principally applied to control moth populations in orchards. In moth, females generally release sex pheromones to attract males, at relatively long distances (several kilometers), for reproduction. The females lay their eggs on orchard trees and larvae develop inside fruits which are then no more eatable.
21Mating disruption consists in affecting the behavior of males in their search of a female for mating by releasing high quantities of synthetic female pheromones in the atmosphere. The disruption of males can be achieved by affecting different biological mechanisms which were originally defined by Bartell (1982). These mechanisms have been recently revised by Miller et al. (2006a, 2006b) and were synthesized in a review by Stelinski (2007). To be an efficient technique to control pests, surrounding orchards or fields must ideally also be part of IPM programs. When the population of moth is too large, mating disruption can be associated with targeted pesticides at local and punctual applications.
22Push-pull strategy. Also called stimulo-deterrent diversion, push-pull strategy is a more recent approach than the other described IPM practices. It consists in a combination of repellent and attractive stimuli modifying the behavior of insect pests and/or of their natural enemies. The insects are deterred or repelled away from the crops (push strategy). They are simultaneously attracted by lures (pull strategy) and concentrated in other areas where they are trapped or killed in a controlled manner. This strategy requires a clear understanding of the pest biology, chemical ecology, and of the interactions with hosts, conspecifics and natural enemies (Cook et al., 2007).
23Biological control. Biological control of the insect pests is defined by Stoner (2004) as “the use of living organisms (insects or pathogens) to suppress pest populations, making them less damaging than they would otherwise be”. Insect natural enemies, also called beneficial insects, can be classified in two classes: predators and parasitoids. Beneficial insects, sometimes exotic, can be artificially introduced in infested fields. This practice must be cautiously managed in order to verify that no-indigenous species will not have an adverse environmental and economic impact, like it was the case with the introduction of the Asian ladybeetle Harmonia axyridis Pallas (Coleoptera: Coccinellidae) (Huelsman et al., 2002; Roy et al., 2006; Brown et al., 2008).
24A new concept consists in attracting local beneficial insects on crops by means of kairomonal substances as explained in Heuskin et al. (2009) for the biological control of aphids with their parasitoid wasps (Aphidius ervi Haliday (Hymenoptera: Braconidae)) (Du et al., 1998; Powell et al., 2003) and their hoverflies predators (Episyrphus balteatus De Geer (Diptera: Syrphidae)) (Francis et al., 2005; Verheggen et al., 2008; Verheggen et al., 2009).
4. Slow release of semiochemicals
4.1. Slow release dispensers
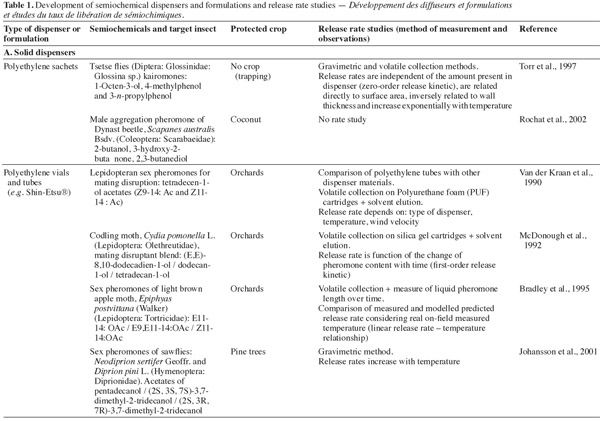
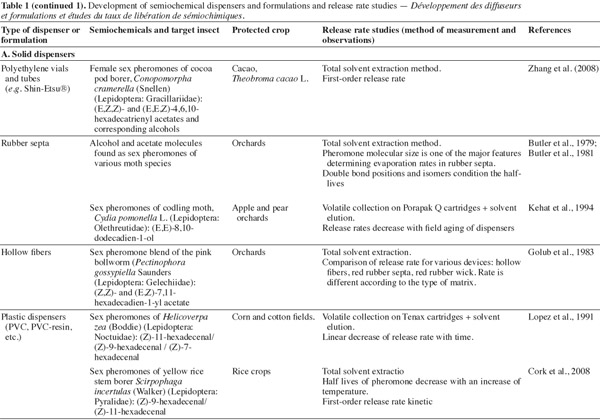
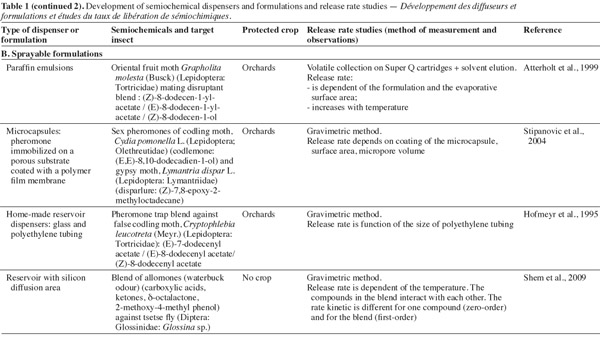
25Major volatile semiochemicals being extremely unstable due to their chemical structure, it is necessary to formulate them so that they are protected from degradation by UV light and oxygen. Moreover, the formulation must ensure a controlled release of semiochemicals. To be efficient in IPM strategies, semiochemical slow-release devices must have particular specifications: the aerial concentration after release must be sufficiently high to be detected by insects; the release of semiochemicals must be effective during all the period of insect occurrence; the production of dispenser must be reproducible. The application of dispensers must be realized early in the season when the pest density is not too high, given that their release rates, for the majority of devices, decrease with time (Witzgall, 2001).
26Several formulations and dispensers have been developed and commercialized with various slow-release capacities. Some examples of dispensers are described hereafter. The majority of them involve mating disruption of moth. Three groups can be distinguished: solid matrix dispensers, liquid formulations to spray and reservoirs of formulations. On an historical point of view, the first related and the most commonly used pheromone dispenser is the natural rubber septum (Roelofs et al., 1972).
27Solid matrix dispensers are hand-applied on crops or in orchards. The semiochemicals are incorporated in a solid matrix. Because of the various materials that can be used to constitute a matrix, the release rates for a single molecule can differ significantly from one device to another, as demonstrated by Golub et al. (1983) for the measurement of release rate of gossyplure ((Z,Z)- and (E,Z)-7,11-hexadecadien-1-yl acetate), the sex pheromone blend of the pink bollworm (Pectinophora gossypiella Saunders, Lepidoptera: Gelechiidae) from different formulations.
28The most common solid matrix used in dispensers are polyethylene tubes (twist tie dispensers like Isomate®), polyethylene sachets (Torr et al., 1997), polyethylene vials (Johansson et al., 2001; Zhang et al., 2008), membrane dispensers (CheckMate CM-XL®), spiral polymer dispensers (NoMate CM®) (Tomaszewska et al., 2005), polymer films, rubber septa (McDonough, 1991; Möttus et al., 1997), rubber wicks, polyvinyl chloride (PVC), hollow fibers (Golub et al., 1983), impregnated ropes, wax formulations, gel-like dispensers matrices (Atterholt et al., 1999).
29Drawbacks encountered with solid matrix dispensers include the difficulty to maintain a zero-order release kinetic (constant release rate) during a long period of time, and the decreasing of aerial semiochemical concentration with the distance from the dispenser. Consequently, these dispensers are only efficient to attract and trap insects at short distance. A way to by-pass this problem is to apply devices in sufficient sites in the crop or in the orchard. The resulting disadvantage is the high manpower needed for application of dispensers in the fields. Another shortcoming is the non biodegradability of the formulated polymers (Stipanovic et al., 2004).
30The effective lifetime of the biggest solid matrix dispensers can range from 60 to 140 days.
31Sprayable slow-release formulations are generally composed of a biodegradable liquid matrix compound in which the semiochemical is dissolved. Regularly, other components can be added to protect the semiochemicals, like UV-stabilizers, antioxidants and surfactants. Frequently, the sprayable formulation consists in a micro-emulsion, resulting in polymeric micro-beads containing the semiochemicals (micro-encapsulated pheromones) dispersed in a liquid matrix (de Vlieger, 2001). In 1999, Atterholt et al. studied the release rates of oriental fruit moth sexual pheromones formulated in aqueous paraffin emulsions as carrier material.
32The time of efficiency of such formulations ranges from days to weeks depending on environmental factors, microbeads size, release capacities, and the pheromones chemical properties (Welter et al., 2005).
33The major advantage of sprayable formulations compared to solid matrix dispensers is that the entire crop can be treated.
34Reservoir dispensers generally consist in two parts, a reservoir and a diffusion area. Hofmeyr et al. (1995) described a dispenser consisting in glass tube acting as a pheromone-impermeable reservoir attached to a short polyethylene tube through which the pheromone can diffuse. Another reservoir was tested by Shem et al. (2009) as repellent allomone device against tsetse flies. The upper part (reservoir) was made of aluminum and the diffusion area was made from Tygon® silicon tubing.
35Aerosol emitters (e.g. Suttera® puffer), consisting in electronically programmed reservoirs of formulation, release large amounts of pheromone by means of a pressurized aerosol. Puffs can be emitted at fixed time intervals. The advantage of this system is the use of fewer dispensers per surface to treat.
36Reservoir systems are the most suitable to approach zero-order release kinetic of semiochemicals (Atterholt et al., 1999).
4.2. Slow release rate studies
37Release rate study does not specify the biological efficiency of a semiochemical delivery dispenser, but gives an idea of the release kinetic over time according to climatic conditions. Many dispensers do not guarantee a release at a steady rate, inducing a decrease of release rate during the season. However, the most important is to know at which moment the quantity of released semiochemical is no more sufficient to influence insect behaviour, and to change the dispenser.
38Techniques to estimate release rates. Given that it is not easy and reliable to measure release rates directly in the field, estimations of semiochemical release rates from formulations were performed in laboratory or semi-controlled conditions. Three different techniques were improved over time: the gravimetric method, the total organic solvent extraction, and the dynamic collection of volatiles. The first procedure, less and less used, consists in weighing dispensers at daily intervals over the season and to determine the percentage of mass loss with time. The major weakness of this technique is the lack of precision and accuracy to set up release rates. Sometimes, the mass increases instead of decreasing due to the presence of humidity and dust deposited on the dispensers.
39The second technique implies the total organic solvent extraction of semiochemicals from dispensers to determine the residual concentration of compound in field-aged devices. The condition to have an optimal pheromone extraction implies the complete dissolution of compound contained in the dispenser (Lopez et al., 1991; Möttus et al., 1997). This technique has the benefit to permit to qualify and quantify the pheromone and its potential volatile degradation products by gas chromatography (GC) analysis. However, it presents a risk of not permitting detection of non-volatile degradation products by GC (Tomaszewska et al., 2005).
40The third method to determine release rate consists in a dynamic sampling and an adsorbent trapping of volatile compounds from field-aged dispensers. The evolution of release rate is estimated according to field-age of devices. It is essential to measure the rate every time in the same conditions of atmospheric pressure, temperature, relative humidity and airflow to obtain analogous analyses over time. The volatile collection system is generally composed of a chamber in which air flows through the dispenser. The carried volatile semiochemicals are trapped on an adsorbent cartridge, followed by solvent extraction or thermal desorption, and GC analysis. Various adsorbents have been tested like Super Q (Mayer et al., 1998; Atterholt et al., 1999; Meagher, 2002), silica gel (McDonough et al., 1992; Pop et al., 1993), Tenax (Cross, 1980), Carbograph, Porapak Q (Cross et al., 1980), activated charcoal, polyurethane foam (PUF) (Van der Kraan et al., 1990; Tomaszewska et al., 2005). The choice of the adsorbent depends on the semiochemical properties, and on the maximum airflow to apply on the cartridge without breakthrough of the compounds.
41Considering the advantages and shortcomings of the three techniques, the last one is the most appropriate and accurate in order to estimate release rate of semiochemicals from dispensers.
42Release rate studies. The release of volatile semiochemicals in the atmosphere is reliant on two major factors: the diffusion speed of the compound through the dispenser matrix and the evaporation speed of the molecule in the air (Krüger et al., 2002). The first factor depends on the characteristics of the dispenser (type of matrix [Golub et al., 1983], size [Hofmeyr at al., 1995], shape, thickness, distribution of the semiochemical in the matrix [Stipanovic et al., 2004]) while the second factor (speed of evaporation) mainly relies on environmental parameters like air temperature, wind speed, relative humidity, and the physical properties of the compound itself (Alfaro-Cid et al., 2009; CBC, n.d.). In the case where the evaporation process of pheromone from the surface of dispenser is slower than the diffusion step, the speed of evaporation is the limiting factor, and the first-order release kinetic equation is considered:
43C0 = Ct e-kt,
44where C0 is the amount of compound in the dispenser at the beginning of evaporation, Ct is the amount of compound at time t, and k is the evaporation rate constant. In case of a first-order kinetic, a half of the amount of the pheromone compound will be evaporated after a time t½, called half-life of the compound (McDonough et al., 1989; Möttus et al., 2001).
45Many studies were conducted to give an estimation of the release rate of pheromone over time from dispensers in definite experimental conditions. However, very few studies dealt with the conception of rate kinetic predictive models according to abiotic parameters (temperature, relative humidity, wind speed, etc.). Moreover, these experiments checked parameters one by one rather than considering their combination regarding an experimental design to finally obtain a realistic rate modeling, close to the kinetic expected on the field.
46Table 1 summarizes studies considering the type of dispenser, the semiochemicals and insects of the research, the targeted crop and the main conclusions of the release rate evaluation.
47Most studies concluded to first-order release kinetics, semiochemical rates decreasing with time and release being dependent on the amount of compound present in the dispenser. Already in 1979 and 1981, Butler et al. showed that alcohol and acetate molecules (sex pheromones of many moth species) were released from rubber septa following a first-order kinetic. Indeed, they concluded that pheromone molecular sizes, double bond positions and isomers conditioned the evaporation rates and the half-life times of the molecules. McDounough et al. (1992) described a modeling of pheromone (codling moth sex pheromones) release rate by determining the half-life times of compounds delivered from field-aged hollow plastic tube dispensers. In 1994, Kehat et al. also found that these codling moth sex pheromones were desorbed from field-aged rubber septa dispensers following a first-order kinetic. Zhang et al. (2008) measured release rate of female sex pheromones of cocoa pod borer, Conopomorpha cramerella, from polyethylene vials placed in a fume hood (20-25°C; 129 ft.min-1 face velocity). They obtained the same kinetic of pheromone delivery. PVC-resin controlled release formulations developed by Cork et al. (2008) for the delivery of yellow rice stem borer sex pheromones were tested at various temperatures (from 22°C to 34°C). Releases followed a first-order kinetic. Moreover, the temperature highly influenced pheromone rates, half-lives decreasing with an increase of the temperature.
48Considering several other studies, temperature is one of the most important climatic parameter that affects volatile release rates. In 1990, Van der Kraan and Ebbers determined the influence of temperature and air velocity on a variety of dispensers delivering moth sex pheromones (tetradecen-1-ol acetate). The authors concluded that the impact of temperature was more important than wind speed on the kinetic of release. Bradley et al. (1995) proposed a linear rate-temperature relationship model to predict release of light brown apple moth pheromones (E11-14:OAc; E9,E11-14: OAc; Z11-14: OAc) from polyethylene tubing dispensers. Two years later, Torr et al. (1997) studied the release of tsetse flies kairomonal substances from polyethylene sachets. Even though release rates were independent of the semiochemical amount present in the dispenser, they increased exponentially with temperature. Atterholt et al. (1999) investigated the release of oriental fruit moth pheromone from paraffin emulsions at three temperatures from 27°C to 49°C. At the lowest temperature, the release rate was constant over time (during 100 days). The release rate was higher at 38°C and 49°C. However, the rate decreased with time at these highest temperatures due to pheromone oxidation and degradation phenomena. Once again, in 2001, Johansson et al. illustrated the increase of sawflies sex pheromone release rate with temperature from polyethylene vial dispensers. More recently, Shem et al. (2009) studied the influence of temperature on the release rate of a blend of allomones derived from waterbuck odor (carboxylic acids, ketones, 2-methoxyphenol, δ-octalactone), in a reservoir type dispenser, to control tsetse flies. As expected, the release rate increased according to the temperature.
49It is not easy to develop and formulate semiochemical delivery systems, which guarantee the diffusion of effective amount of compound along the season. With first-order release kinetics, semiochemical rates decrease quickly and, as a consequence dispenser field-life is often too short to cover the period of pest occurrence.
5. Conclusion
50At the end of this review, two questions remain: what kind of dispenser is the best in IPM programs? What is the lifetime of dispenser in terms of semiochemical diffusion efficiency?
51To answer the first question, the choice of dispenser (solid matrix, formulation, reservoir, puffer) will mainly depend on the needs of the crop farmers, taking into account the labor and the manpower costs to implement IPM strategies. Other important decisional criteria are the targeted pest, the season of occurrence of the insects (with the knowledge of the mean climatic conditions) and the IPM tactic itself. Moreover, environment protection can also be determinative in the dispenser selection. Biodegradable matrix, environmentally safe, could be preferred as slow-release device material for semiochemical delivery. Alfaro-Cid et al. (2009) recently attempted to develop an eco-friendly biodegradable dispenser for codling moth mating disruption. Additionally this experimental system seemed to have small sensitivity to climatic conditions.
52The second question implies the knowledge of the semiochemical release rate kinetic. As demonstrated all along the review, this kinetic relies on the type of molecule, the dispenser, and the climatic conditions. The perspective to develop case by case (semiochemical-dispenser) predictive slow-release models taking into account the climatic parameters is an ideal but difficult approach. Experiments conducted to reproduce the environmental conditions faced the constraint that the fluctuations observed in field are too unpredictable and random to be duplicated in laboratory. The laboratory studies can only predict limitations of use in fixed conditions and give theoretical information on dispenser lifetime. Furthermore, such studies are generally time and money consuming. For these reasons, the best way to estimate diffusion efficiency consists in regularly measuring the residual semiochemical quantity and/or determining release rate from field-aged dispensers. This approach, generally less time consuming, gives a direct indication of the dispenser release effectiveness and the moment to replace pheromone delivery system on field.
53In conclusion, the perspectives of semiochemicals use in IPM programs seem to be promising with the increasing worldwide biological agriculture. Slow-release dispenser and formulation improvement will continue with the contribution of multiple scientific fields of research (entomology, chemistry, ecology, etc.) and the crop farmer skills.
Bibliographie
Alfaro-Cid E. et al., 2009. Modeling pheromone dispensers using genetic programming. In: Giacobini M. et al., eds. Evo Workshops 2009. LNCS 5484. Berlin, Germany ; Heidelberg, Germany: Springer-Verlag, 635-644.
Atterholt C.A., Delwiche M.J., Rice R.E. & Krochta J.M., 1999. Controlled release of insect sex pheromones from paraffin wax and emulsions. J. Controlled Release, 57, 233-247.
Bartell R.J., 1982. Mechanisms of communication disruption by pheromone in the control of Lepidoptera: a review. Physiol. Entomol., 7, 353-364.
Bradley S.J. et al., 1995. A temperature-dependent model for predicting release rates of pheromone from a polyethylene tubing dispenser. J. Chem. Ecol., 21(6), 745-760.
Brossut R., 1997. Phéromones : la communication chimique chez les animaux. Paris, Éditions Belin : Croisée des Sciences.
Brown P.M.J. et al., 2008. Harmonia axyridis in Europe: spread and distribution of a non-native coccinellid. BioControl, 53(1), 5-21.
Butenandt A., Beckmann R., Stamm D. & Hecker E., 1959. Über den Sexuallockstoff desSeidenspinners Bombyx mori. Reindarstellung und Konstitution. Z. Naturforsch., 14b, 283-284.
Butler L.I. & McDonough L.M., 1979. Insect sex pheromones: evaporation rates of acetates from natural rubber septa. J. Chem. Ecol., 5(5), 825-837.
Butler L.I. & McDonough L.M., 1981. Insect sex pheromones: evaporation rates of alcohols and acetates from natural rubber septa. J. Chem. Ecol., 7(3), 627-633.
CBC, n.d. Pheromones and mating disruption, http://www.cbceurope.it/images/stories/file/biocontrol/GuidaBioENG.pdf, (1/7/2010).
Cook S.M., Khan Z.R. & Pickett J.A., 2007. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol., 52, 375-400.
Cork A., 2004. Pheromone manual. Chatham Maritime, UK: Natural Resources Institute.
Cork A. et al., 2008. Development of a PVC-resin-controlled release formulation for pheromones and use in mating disruption of yellow rice stem borer, Scirpophaga incertulas. Crop Prot., 27, 248-255.
Cross J.H., 1980. A vapor collection and thermal desorption method to measure semiochemical release rates from controlled release formulations. J. Chem. Ecol., 6(4), 781-787.
Cross J.H., Tumlinson J.H., Heath R.E. & Burnett D.E., 1980. Apparatus and procedure for measuring release rates from formulations of lepidopteran semiochemicals. J. Chem. Ecol., 6(4), 759-770.
De Vlieger J.J., 2001. Development of a sprayable slow-release formulation for the sex pheromone of the Mediterranean corn borer, Sesamia nonagroides. IOBC wprs Bull., 24(2), 101-106.
Du Y. et al., 1998. Identification of semiochemicals released during aphid feeding that attracts parasitoid Aphidius ervi. J. Chem. Ecol., 24(8), 1355-1368.
Francis F., Martin T., Lognay G. & Haubruge E., 2005. Role of (E)-β-farnesene in systematic aphid prey location by Episyrphus balteatus larvae (Diptera: Syrphidae). Eur. J. Entomol., 102, 431-436.
Golub M., Weatherston J. & Benn M.H., 1983. Measurement of release rates of gossyplure from controlled release formulations by mini-airflow method. J. Chem. Ecol., 9(3), 323-333.
Heuskin S. et al., 2009. Fast gas chromatography characterisation of purified semiochemicals from essential oils of Matricaria chamomilla L. (Asteraceae) and Nepeta cataria L. (Lamiaceae). J. Chromatogr. A, 1216, 2768-2775.
Hofmeyr H. & Burger B.V., 1995. Controlled-release pheromone dispenser for use in traps to monitor flight activity of false codling moth. J. Chem. Ecol., 21(3), 355-363.
Huelsman M.F. et al., 2002. Multicolored Asian lady beetle (Harmonia axyridis) as a nuisance pest in household in Ohio. In: Proceedings of the 4th International Conference on Urban Pests, Integrated Pest Management Program, Ohio State University, www.icup.org.uk/reports%5CICUP226.pdf., (06/07/10).
Johansson B.G. et al., 2001. Release rates for pine sawly pheromones from two types of dispensers and phenology of Neodiprion sertifer. J. Chem. Ecol., 27(4), 733-745.
Kehat M. et al., 1994. Sex pheromone traps for monitoring the codling moth: effect of dispenser type, field aging of dispenser, pheromone dose and type of trap on male captures. Entomol. Exp. Appl., 70, 55-62.
Kogan M., 1998. Integrated pest management: historical perspectives and contemporary developments. Annu. Rev. Entomol., 43, 243-270.
Krüger A.J. & Tolmay A.T., 2002. Prediction of the release characteristics of alcohols from EVA using a model based on Fick’s second law of diffusion. J. Appl. Polym. Sci., 84, 806-813.
Lopez J.D., Leonhardt B.A. & Shaver T.N., 1991. Performance criteria and specifications for laminated plastic sex pheromone dispenser for Helicoverpa zea (Lepidoptera: Noctuidae). J. Chem. Ecol., 17(11), 2293-2305.
Mayer M.S. & Mitchell E.R., 1998. Rapid measure of sex pheromone emission from plastic rope dispensers: exemple of utility in sexual communication disruption of the diamondback moth, Plutella xylostella. Phytoparasitica, 26(2), 1-9.
McDonough L.M., 1991. Controlled release of insect sex pheromones from a natural rubber substrate. In: Hedin P.A., ed. Naturally occurring pest bioregulators. ACS Symposium serie 449. Washington, DC, USA: American Chemical Society, 106-124.
McDonough L.M., Brown D.F. & Aller W.C., 1989. Insect sex pheromones. Effect of temperature on evaporation rates of acetates from rubber septa. J. Chem. Ecol., 15(3), 779-790.
McDonough L.M., Aller W.C. & Knight A.L., 1992. Performance characteristics of a commercial controlled-release dispenser of sex pheromones for control of codling moth (Cydia pomonella) by mating disruption. J. Chem. Ecol., 18(12), 2177-2189.
Meagher R.L. Jr, 2002. Trapping noctuid moths with synthetic floral volatile lures. Entomol. Exp. Appl., 103, 219-226.
Miller J.R., Gut L.J., de Lame F.M. & Stelinski L.L., 2006a. Differentiation of competitive vs non-competitive mechanisms mediating disruption of moth sexual communication by point sources of sex pheromone (Part I): theory. J. Chem. Ecol., 32(10), 2089-2114.
Miller J.R., Gut L.J., de Lame F.M. & Stelinski L.L., 2006b. Differentiation of competitive vs non-competitive mechanisms mediating disruption of moth sexual communication by point sources of sex pheromone (Part II): case studies. J. Chem. Ecol., 32(10), 2115-2143.
Möttus E., Nômm V., Williams I.H. & Liblikas I., 1997. Optimization of pheromone dispensers for diamondback moth Plutella xylostella. J. Chem. Ecol., 23(9), 2145-2159.
Möttus E. et al., 2001. Calculation and using of pheromone communication channel parameters for optimization of pheromone dispensers. In: Metspalu L. & Mitt S., eds. Proceedings of the international workshop, Practice oriented results on the use of plant extracts and pheromones in pest control, 24-25 January 2001, Tartu, Estonia, 101-125.
Nieberding C. et al., 2008. Male sex pheromones in the butterfly Bicyclus anynana: towards an evolutionary analysis. PLoS ONE, 3(7), e2751.
Phillips T.W., 1997. Semiochemicals of stored-product insects: research and applications. J. Stored Prod. Res., 33(1), 17-30.
Pop L., Arn H. & Buser H.-R., 1993. Determination of release rates of pheromone dispensers by air sampling with C-18 bonded silica. J. Chem. Ecol., 19(11), 2513-2519.
Powell W. & Pickett J.A., 2003. Manipulation of parasitoids for aphid pest management: progress and prospects. Pest Manage. Sci., 59(2), 149-155.
Rochat D. et al., 2002. Activity of male pheromone of Melanesian rhinoceros beetle Scapanes australis. J. Chem. Ecol., 28(3), 479-500.
Roelofs W.L. et al., 1972. Codling moth sex attractant-field trials with geometrical isomers. J. Econ. Entomol., 65, 1276-1277.
Roy H., Brown P. & Majerus M., 2006. Harmonia axyridis: a successful biocontrol agent or an invasive threat? In: Eilenberg J. & Hokkanen H.M.T., eds. An ecological and societal approach to biological control. Dordrecht, The Netherlands: Springer, 295-309.
Shem P.M. et al., 2009. Release kinetics of a synthetic tsetse allomone derived from waterbuck odour from a Tygon silicon dispenser under laboratory and semi field conditions. American-Eurasian J. Agric. Environ. Sci., 6(6), 625-636.
Stelinski L.L., 2007. On the physiological and behavioral mechanisms of pheromone-based mating disruption. Pestycydy, 3(4), 27-32.
Stipanovic A.J., Hennessy P.J., Webster F.X. & Takahashi Y., 2004. Microparticle dispensers for the controlled release of insect pheromones. J. Agric. Food Chem., 52, 2301-2308.
Stoner K., 2004. Approaches to the biological control of insects. Cooperative Extension Bulletin 7144. Orono, ME, USA: University of Maine, www.umext.maine.edu/onlinepubs/PDFpubs/7144.pdf, (06/07/10).
Tomaszewska E. et al., 2005. Evaluation of pheromone release from commercial mating disruption dispensers. J. Agric. Food Chem., 53, 2399-2405.
Torr S.J., Hall D.R., Phelps R.J. & Vale G.A., 1997. Methods for dispensing odour attractants for tsetse flies (Diptera: Glossinidae). Bull. Entomol. Res., 87, 299-311.
Van der Kraan C. & Ebbers A., 1990. Release rates of tetradecen-1-ol acetates from polymeric formulations in relation to temperature and air velocity. J. Chem. Ecol., 16(4), 1041-1058.
Verheggen F.J. et al., 2008. Aphid and plant volatiles induce oviposition in an aphidophagous hoverfly. J. Chem. Ecol., 34(3), 301-307.
Verheggen F.J., Haubruge E., De Moraes C.M. & Mescher M.C., 2009. Social environment influences aphid production of alarm pheromone. Behav. Ecol., 20, 283-288.
Verheggen F.J., Haubruge E. & Mescher M.C., 2010. Alarm pheromones. In: Litwack G., ed. Pheromones. Amsterdam, The Netherlands: Elsevier.
Weinzierl R., Henn T., Koehler P.G. & Tucker C.L., 2005. Insect attractants and traps. IFAS Extension. Gainesville, FL, USA: University of Florida, http://edis.ifas.ufl.edu/in080, (05/05/10).
Welter S.C. et al., 2005. Pheromone mating disruption offers selective management options for key pests. California Agric., 59(1), 16-22, http://fruitsandnuts.ucdavis.edu/uops/pheromone_welter_03_05.pdf, (16/05/10).
Wilson E.O., 1971. The insect societies. Cambridge, MA, USA: The Belknap Press of Harvard University Press.
Wilson E.O. & Bossert W.H., 1963. Chemical communication among animals. Recent Progr. Hormone Res., 19, 673-716.
Witzgall P., 2001. Pheromones – future techniques for insect control? Pheromones for insect control in orchards and vineyards. IOBC wprs Bull., 24(2), 114-122.
Zhang A. et al., 2008. Activity evaluation of cocoa pod borer sex pheromone in cacao fields. Environ. Entomol., 37(3), 719-724.
Para citar este artículo
Acerca de: Stéphanie Heuskin
Univ. Liege - Gembloux Agro-Bio Tech. Department of Analytical Chemistry. Passage des Déportés, 2. B-5030 Gembloux (Belgium). E-mail: stephanie.heuskin@ulg.ac.be – Univ. Liege - Gembloux Agro-Bio Tech. Department of General and Organic Chemistry. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Acerca de: François J. Verheggen
Univ. Liege - Gembloux Agro-Bio Tech. Department of Functional and Evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Acerca de: Éric Haubruge
Univ. Liege - Gembloux Agro-Bio Tech. Department of Functional and Evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Acerca de: Jean-Paul Wathelet
Univ. Liege - Gembloux Agro-Bio Tech. Department of General and Organic Chemistry. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Acerca de: Georges Lognay
Univ. Liege - Gembloux Agro-Bio Tech. Department of Analytical Chemistry. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
