


- Home
- Volume 15 (2011)
- numéro 4
- Dimensionnement et extrapolation des bioréacteurs sur base de paramètres physiologiques : cas de la production de lipase par Yarrowia lipolytica


View(s): 11600 (126 ULiège)
Download(s): 1847 (10 ULiège)

Dimensionnement et extrapolation des bioréacteurs sur base de paramètres physiologiques : cas de la production de lipase par Yarrowia lipolytica

Editor's Notes
Reçu le 18 aout 2010, accepté le 8 février 2011
Résumé
L'extrapolation des bioréacteurs est souvent une étape limitante pour le développement des bioprocédés du fait des nombreux paramètres physiques et biologiques à prendre en compte. Dans ce travail, les approches classiques de l'extrapolation des bioprocédés reposant sur les aspects du génie chimique et du génie des procédés seront abordées comme premier niveau d'approche. Ces approches traditionnelles seront comparées à une approche plus récente basée sur les paramètres physiologiques directs mettant en œuvre le comportement de la souche microbienne dans le bioréacteur. Cette approche « physiologique » est innovante car elle fait intervenir des paramètres en relation directe avec la physiologie du micro-organisme, comme par exemple l'induction d'un gène spécifique dans des conditions extracellulaires données, et ouvre donc de nombreuses possibilités au niveau du contrôle des bioprocédés. Ce principe sera développé ici dans le cadre du suivi de la production de lipases par Yarrowia lipolytica à l'échelle de la cellule individuelle.
Abstract
Bioreactor scale-up and design on the basis of physiologically relevant parameters: application to the production of lipase by Yarrowia lipolytica. Bioreactor scale-up often poses a serious issue during the industrial development of a bioprocess considering the numerous physical and biological phenomena occurring in the reacting volume. The basic principles of scale-up coming from the traditional chemical and process engineering approaches will be first reviewed and will be then compared to a new one involving recent development at the level of microbial strain manipulation. This “physiological” approach of scale-up involves directly a biological component of the system (by comparison with the traditional approach for scaling-up involving physical parameters indirectly linked to the physiological phenomena occurring in the bioreactor), i.e. the synthesis of a reporter fluorescent protein when microbial cells are exposed to stress. It will be shown how this principle can be used for a better understanding of the relationship between bioreactor hydrodynamics and microbial stress.
Table of content
1. Introduction
1L'extrapolation d'un bioprocédé est une étape critique qui garantit la viabilité économique du bioproduit visé. Elle consiste à augmenter le volume du bioréacteur afin d'atteindre une productivité suffisante, et ce, en maintenant autant que possible les rendements obtenus lors des tests en laboratoire ou à l'échelle pilote. Pour la plupart des produits biotechnologiques (antibiotiques, alcools, acides organiques, acides aminés, enzymes, probiotiques), un procédé viable n'est atteint que pour des volumes de bioréacteur de quelques centaines de milliers de litres. Ces volumes permettent de satisfaire une demande de marché importante (de 102 à 107 tonnes par an) pour des produits de faible valeur ajoutée (de 0,5 à 100 euros.kg-1). À côté de ces bioproduits de faible valeur ajoutée, les produits biotechnologiques plus fins (vaccins, protéines recombinantes thérapeutiques, etc.) permettent d'atteindre des valeurs ajoutées significativement plus importantes (de l'ordre de 104 à 109 euros.kg-1). Par contre, leur demande mondiale est nettement moindre (de 0,1 à 1 000 kg par an) et leur développement ne nécessite donc pas une étape d'extrapolation importante. Ces procédés à haute valeur ajoutée requièrent des volumes de culture industriels plus modestes, de l'ordre de quelques dizaines de mètres cubes et peuvent mettre en œuvre des cultures microbiennes ou des cultures cellulaires (cellules animales, végétales, d'insecte, etc.) (Palomares et al., 2009). Dans ce travail, nous développerons plus en détail les processus d'extrapolation pour des bioproduits d'origine microbienne (c'est-à-dire issus de bactéries, de levures ou de moisissures). L'extrapolation des bioprocédés peut être abordée de différentes manières comme montré à la figure 1. La première approche est celle du génie chimique (Figure 1A) et consiste à calculer la variation de paramètres de l'opération de mélange (comme la puissance dissipée P par l'agitateur, le nombre de Reynolds Re qui caractérise le type d'écoulement qui règne dans la cuve, etc.) lorsque l'extrapolation du réacteur (de dimension caractéristique D correspondant au diamètre de la cuve) est effectuée en maintenant un ou plusieurs paramètres d'agitation constants. Ce premier niveau d'approche de l'extrapolation est le plus simple et est encore d'application de nos jours. Néanmoins, nous verrons dans la première partie de cet article que cette approche est loin d'être suffisante, d'une part parce qu'elle ne garantit pas un maintien des paramètres de l'opération de mélange lorsque la taille du réacteur augmente (hétérogénéité au sein du volume réactionnel) et d'autre part, parce qu'elle ne tient pas compte de la physiologie des micro-organismes. La seconde approche (Figure 1B) consiste à étudier les relations entre les paramètres de performance (coefficient de transfert d'oxygène KLa, temps de mélange tm, etc.) et les rendements de procédé (quantité de biomasse ou de métabolites produite) ou l'évolution des conditions physico-chimiques au sein du réacteur (pH, oxygène dissous, CO2, température, etc.). Cette approche est complémentaire à la première décrite, puisque les paramètres de performance dépendent directement de l'efficacité de l'opération de mélange, cette dernière étant proportionnelle à la quantité d'énergie investie au niveau des mobiles d'agitation (Nienow, 1998). L'avantage de cette approche est qu'elle peut être directement implémentée au niveau du bioréacteur industriel en mettant en place un système de régulation adapté et permet donc de suivre en ligne les demandes de la population microbienne cultivée. Par exemple, le niveau d'oxygène dissous dans le réacteur (condition physico-chimique) peut être maintenu à un niveau constant en augmentant progressivement le KLa du réacteur par modulation du débit d'air et/ou la vitesse d'agitation en fonction des besoins du micro-organisme. Les problèmes de cette approche sont de trois types :
2– premièrement, les types de sonde disponibles sont limités et leur mesure est indirecte par rapport à la physiologie du micro-organisme ;
3– deuxièmement, au fur et à mesure de l'augmentation de la taille des bioréacteurs, les conditions physico-chimiques vont être de plus en plus hétérogènes du fait de la baisse de l'efficacité de mélange du réacteur. Une mesure réalisée en un point ne sera donc pas représentative du comportement global du bioréacteur ;
4– troisièmement, la population microbienne présente dans le réacteur est considérée comme homogène.
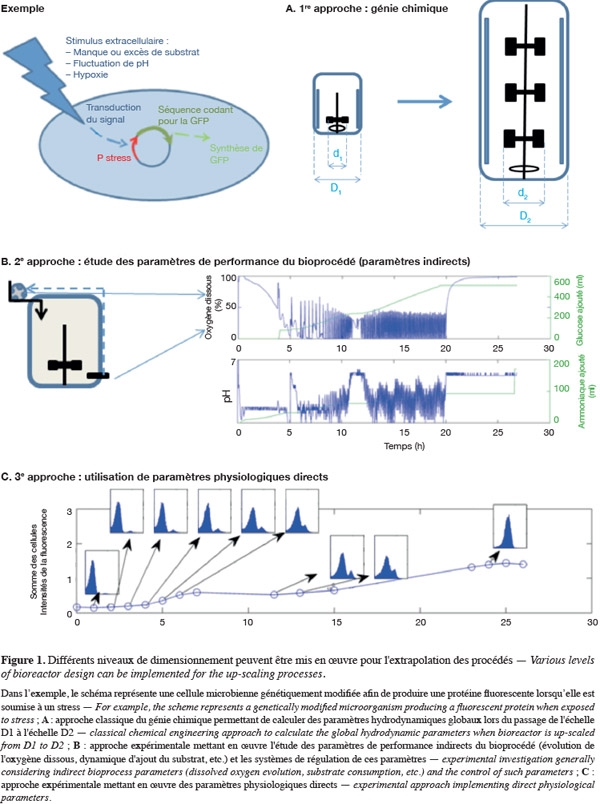
5De nombreuses études réalisées avec des outils microbiologiques adaptés (décrits dans le § 4) ont montré que la population microbienne pouvait être ségrégée au niveau d'une ou plusieurs caractéristiques cellulaires (taille et forme des cellules, viabilité, capacité de synthèse d'une enzyme particulière, etc.) (Müller et al., 2010).
6La troisième approche (Figure 1C) est basée sur l'analyse de l'évolution du bioprocédé à l'échelle de la cellule microbienne et tient compte des limitations relevées lors des deux précédentes approches. L'intérêt de cette méthode est qu'elle permet de prendre en compte à la fois l'hétérogénéité de la population microbienne et l'hétérogénéité spatiale du milieu réactionnel.
7Il est intéressant de noter que l'évolution dans les approches d'extrapolation des bioréacteurs s'effectue en considérant une échelle spatiale de plus en plus petite, de quelques mètres (échelle caractéristique du procédé industriel pouvant être représenté par le diamètre du réacteur D) jusqu'au micron (échelle caractéristique du micro-organisme). Cette multiplicité au niveau des échelles spatiales à considérer est une des clés du succès de l'extrapolation d'un bioprocédé. Nous verrons en effet que différentes échelles spatiales et temporelles caractérisent le comportement du bioréacteur (système physique à extrapoler) et la réaction de la population microbienne (système biologique qui conditionne le rendement du bioprocédé). L'évolution des différentes approches d'extrapolation des bioprocédés sera illustrée par un procédé de production de lipase par Yarrowia lipolytica. Y. lipolytica est une levure non conventionnelle dont la culture entraine une série de contraintes au niveau du dimensionnement du bioréacteur. L'étude de ce cas particulier illustre bien l'importance des caractéristiques du micro-organisme pour l'élaboration du procédé.
2. Paramètres de base à prendre en compte lors de la culture d'une levure non conventionnelle à partir d'un substrat hydrophobe
8Les techniques d'extrapolation des bioréacteurs exposées lors de la section précédente ont été particulièrement bien éprouvées pour toute une série de micro-organismes conventionnels, tels que Escherichia coli (bactérie Gram- utilisée pour l'expression de protéines recombinantes), Bacillus subtilis (bactérie Gram+ utilisée pour la production d'enzymes, de peptides à activités biologiques ou encore pour la production de protéines recombinantes) et Saccharomyces cerevisiae (levure Crabtree positive) (Junker, 2004).
9Nous allons considérer ici les techniques d'extrapolation à mettre en œuvre dans le cas de la levure non conventionnelle Yarrowia lipolytica. Le terme non conventionnel signifie ici que Y. lipolytica ne fait pas partie des micro-organismes les plus couramment utilisés au niveau industriel. Néanmoins, le génome de Y. lipolytica a, comme les micro-organismes de référence cités plus haut, été complètement séquencé (Dujon et al., 2004). La plupart des données génomiques concernant Y. lipolytica ont été compilées sur des bases de données telles que KEGG (Kaneshina et al., 2000). Y. lipolytica fait partie des micro-organismes dits « oléagineux », c'est-à-dire capables d'accumuler des lipides à des niveaux pouvant atteindre 40 % de leur biomasse (Beopoulos et al., 2009a). De plus, Y. lipolytica possède les voies métaboliques nécessaires à l'assimilation des substrats hydrophobes (Beopoulos et al., 2009b). La dégradation des substrats hydrophobes fait notamment intervenir une lipase codée par le gène lip2 qui présente de nombreux intérêts technologiques (Destain et al., 1997 ; Yu et al., 2007). La mise en œuvre d'un procédé de production de lipase par Y. lipolytica doit donc être envisagée sur base d'un substrat hydrophobe (Nicaud et al., 2002). Afin de garantir une viabilité économique au procédé, une source de carbone issue de la valorisation de sous-produits industriels est préférable, comme par exemple le méthyl oléate (Destain et al., 2005). Cette source de carbone hydrophobe a été comparée à d'autres substrats et montre des potentialités importantes pour la production de lipases (Fickers et al., 2004 ; Fickers et al., 2005a). Un paramètre essentiel pour la réussite du procédé est l'excrétion du produit dans le milieu extracellulaire. En effet, les étapes d'extraction et de purification des produits intracellulaires sont couteuses et difficiles à mettre en œuvre industriellement. Le lipase LIP2 est une enzyme extracellulaire, mais les conditions de procédés doivent être optimisées afin de favoriser son excrétion (Pereira-Meirelles et al., 2000 ; Nicaud et al., 2002).
10Le substrat n'étant pas soluble dans la phase aqueuse, les conditions d'agitation doivent être optimisées afin de déterminer sa dispersion. Cette optimisation doit être effectuée en prenant en compte l'effet du stress mécanique sur le micro-organisme. La sensibilité au stress mécanique (stress de cisaillement subi par les micro-organismes lors de leur passage au niveau de l'agitateur) dépend fortement de la taille du micro-organisme par rapport à la plus petite échelle de turbulence, dite échelle de Komogoroff, rencontrée dans le volume réactionnel. Pour des puissances d'agitation classiquement rencontrées dans les bioréacteurs (30 W.kg-1), cette échelle peut être estimée à 10 µm (Hewitt et al., 2007). Cela signifie que toute particule dont la taille est inférieure à 10 µm ne ressent pas le stress de cisaillement. Néanmoins, dans certaines conditions, Y. lipolytica montre un phénomène de dimorphisme cellulaire (Zinjarde et al., 1998), c'est-à-dire un allongement de la taille des cellules vers une morphologie mycélienne (Figure 2). Les conditions environnementales induisant le dimorphisme cellulaire ne sont pas encore bien connues (Herrera et al., 2002), mais plusieurs études ont montré que le pH du milieu avait un impact significatif sur ce phénomène (Szabo et al., 2002 ; Kar et al., 2008). Cet allongement de la taille des cellules doit être pris en compte pour le dimensionnement du procédé, car il augmente la sensibilité du micro-organisme aux contraintes mécaniques.

11Un autre paramètre critique lors de la culture en bioréacteur est l'aération. Dans le cas de Y. lipolytica, micro-organisme aérobie strict, l'apport en oxygène est un paramètre critique (Lopes et al., 2008). La vitesse de transfert de l'oxygène doit donc être maximisée en prenant en compte le phénomène de moussage. En effet, les milieux industriels contiennent des substrats protéiques qui stabilisent la mousse. Lors des processus aérobies intensifs, un compromis doit être trouvé au niveau d'une opération d'agitation/aération assurant un bon transfert d'oxygène tout en limitant la formation de mousse (Delvigne et al., 2010). Généralement, la formation de mousse peut être limitée en considérant un système de régulation permettant l'ajout d'agents antimoussants lors de la culture (Etoc et al., 2006). Néanmoins, l'ajout d'antimousse chimique limite la capacité de transfert d'oxygène du réacteur et entraine de nombreux problèmes lors de la purification de la lipase (Fickers et al., 2006).
12Les caractéristiques de la levure Y. lipolytica soulignées dans ce paragraphe montrent bien la sensibilité de ce micro-organisme face aux conditions environnementales rencontrées dans les bioréacteurs. L'extrapolation du procédé devra donc être effectuée en prenant en compte les caractéristiques physiologiques propres à ce type de micro-organisme. Les paragraphes suivants illustreront comment les trois approches d'extrapolation décrites à la figure 1 peuvent être employées pour la mise au point industrielle d'un procédé mettant en œuvre cette levure non conventionnelle.
3. Premier niveau de dimensionnement des bioréacteurs : paramètres hydrodynamiques
13À l'échelle industrielle, les cultures microbiennes ou cellulaires sont dans la majorité des cas réalisées dans des bioréacteurs (également appelés fermenteurs dans le cas des cultures microbiennes) pouvant être mécaniquement agités (agitation directe par un mobile) ou pneumatique (agitation indirecte effectuée par injection d'air). Notons également à ce niveau le récent intérêt de l'industrie pharmaceutique pour les réacteurs dits « disposables » ou jetables. À l'heure actuelle, ces bioréacteurs en matières plastiques ont des volumes restreints à quelques centaines de litres et sont limités à des applications de type culture cellulaire pour des produits à très haute valeur ajoutée. Les principes de dimensionnement qui seront énoncés dans les paragraphes qui suivent seront donc limités aux bioréacteurs classiques mécaniquement agités qui sont les équipements les plus largement répandus à l'échelle industrielle (Charles et al., 2010).
14D'un point de vue géométrique, l'extrapolation peut être représentée suivant le schéma montré à la figure 1A où le calcul consiste à suivre l'évolution des paramètres de l'opération de mélange lorsque le diamètre du bioréacteur passe de D1 à D2. Les calculs d'extrapolation requièrent la connaissance de plusieurs variables de l'opération de mélange qui seront brièvement présentées au paragraphe suivant.
3.1. Dimensionnement de base et extrapolation des bioréacteurs mécaniquement agités
15La première étape du dimensionnement d'un bioréacteur mécaniquement agité consiste à déterminer le régime d'écoulement du fluide agité : laminaire, transitoire ou turbulent. Il est en effet important, dans la plupart des cas, d'opérer en régime turbulent afin d'intensifier les opérations de transfert (masse, chaleur et quantité de mouvement). Le calcul du régime d'écoulement au sein d'un bioréacteur mécaniquement agité peut être effectué à l'aide d'un nombre de Reynolds adapté :
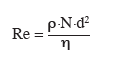
16avec étant la masse volumique du fluide agité (kg.m-³), N le taux d'agitation (s-1), d le diamètre du mobile d'agitation (m), η la viscosité du fluide (Pa.s).
17Dans le cas de l'eau, est de 1 000 kg.m-³ et η est de 0,001 Pa.s. Dans les milieux dont la rhéologie est proche de celle de l'eau, on obtient donc facilement un nombre de Reynolds important. Pour Re > 104, le milieu agité est considéré comme étant en écoulement turbulent.
18Le fait d'opérer en écoulement turbulent permet de simplifier le calcul des paramètres de base de l'opération de mélange, telle que la puissance dissipée au niveau du mobile d'agitation. Cette puissance peut être calculée à l'aide de la relation suivante :
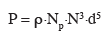
19Np correspond au nombre de puissance ou de Newton et est constant en régime d'écoulement turbulent (il ne dépend que du type de mobile d'agitation utilisé et est généralement fourni par le concepteur).
20Le calcul de la puissance dissipée par le système d'agitation est une étape importante lors du dimensionnement d'un bioréacteur. En effet, plus cette puissance est importante et plus l'intensité des opérations de transfert sera grande au sein du fluide agité. Des corrélations directes sont en effet observées entre la puissance dissipée et le temps d'homogénéisation du réacteur, ainsi que du coefficient de transfert d'oxygène (Palomares et al., 2009). Ces deux paramètres constituent souvent les étapes limitantes de l'extrapolation des bioprocédés. La puissance dissipée est souvent considérée comme un invariant dans les calculs d'extrapolation. Un invariant est un paramètre de l'opération de mélange qui a été identifié comme ayant un impact majeur sur le procédé et qui est maintenu constant lors de l'extrapolation. Il permet de fixer l'évolution de l'échelle temporelle du procédé. Dans les bioprocédés, vu l'impact des phénomènes de transfert, la puissance volumique (en W.m-³) est souvent considérée comme invariant. Les critères d'extrapolation utilisés en pratique au niveau industriel sont les suivants : puissance volumique constante (30 % d'utilisation), coefficient de transfert d'oxygène constant (30 %), vitesse périphérique π.N.d constante afin de limiter les stress liés au cisaillement (20 %), concentration en oxygène dissous constante (20 %) (Garcia-Ochoa et al., 2009).
3.2. Problèmes associés à l'intensification des transferts gaz-liquide
21L'oxygène est très peu soluble dans les milieux aqueux, la concentration à saturation dans le liquide étant d'environ 8 mg.l-1 dans les conditions usuelles de bioprocédé. Le bioréacteur doit donc être dimensionné de manière à maximiser la vitesse de transfert de l'oxygène de la phase gazeuse vers la phase liquide. Cette vitesse de transfert peut être caractérisée par la relation suivante :
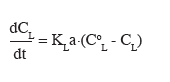
22où CL est la concentration en oxygène dissous (kg.m-³), CoL est la concentration en oxygène dissous à saturation (kg.m-³), KLa est le coefficient volumique de transfert d'oxygène (s-1). Ce coefficient peut être facilement mesuré et est souvent considéré comme critère de performance pour caractériser l'efficacité de transfert d'oxygène des bioréacteurs (Garcia-Ochoa et al., 2009).
23L'intensification du KLa est effectuée en augmentant la vitesse d'agitation et le débit d'air selon des proportions bien définies afin d'éviter le phénomène d'engorgement des mobiles d'agitation par les cavités gazeuses (Nienow, 1998). D'autres solutions peuvent également être envisagées, comme par exemple l'augmentation de la pression interne du réacteur afin d'améliorer la solubilité de l'oxygène dans le milieu aqueux, ou encore l'enrichissement de l'air injecté dans le réacteur par de l'oxygène pur.
24Un problème technologique majeur des processus aérobies intensifs est la formation de mousses (Van't Riet et al., 1992 ; Vardar-Sukan, 1998 ; Delvigne et al., 2010). La présence de protéines ou de biosurfactants dans le milieu réactionnel tend à stabiliser la dispersion gaz-liquide au niveau de l'espace de tête du réacteur (Van't Riet et al., 1992). Le moussage du milieu de culture peut entrainer le débordement du réacteur, ce qui induit des problèmes de perte de produits et de contamination. De nombreux progrès ont été envisagés à ce niveau, comme le réglage des conditions d'agitation et d'aération pour limiter la formation de mousse tout en gardant une capacité de transfert d'oxygène optimale (Hoeks et al., 2003), la conception de nouveaux antimousses chimiques (Etoc et al., 2006) ou encore la conception de capteurs dédiés à la détection de l'intensité du phénomène de moussage (Varley et al., 2004). Malgré ces efforts entrepris, la formation de mousses reste un problème technologique majeur dans la mise en œuvre des bioréacteurs.
4. Deuxième niveau de dimensionnement des bioréacteurs : mesure et contrôle en ligne des paramètres environnementaux
25Les bioréacteurs sont équipés de systèmes de régulation très performants qui permettent de maintenir des conditions de culture à un niveau constant. Ceux-ci comprennent un couple senseur (une sonde qui mesure un paramètre donné, comme le pH ou la température) - actuateur (un système qui permet d'ajuster la valeur du paramètre mesuré, comme par exemple une pompe d'injection de base ou d'acide pour la régulation du pH). Les deux composants sont reliés par une boucle de régulation. Une stratégie d'extrapolation peut donc être de mettre en évidence les paramètres physico-chimiques critiques pour le procédé et de les maintenir à un niveau constant. Le système de régulation peut en effet être facilement transposé à l'échelle industrielle. Les paragraphes qui suivent présentent la mise en place des boucles de régulation classiquement retrouvées au niveau des bioréacteurs ainsi que leur limitation lors de l'extrapolation des bioprocédés.
4.1. Régulation du pH, de la température, de l'oxygène dissous et du niveau de substrat dans le volume réactionnel
26Les systèmes de régulation standards équipant les bioréacteurs font intervenir une boucle de régulation pour le pH et une boucle de régulation pour la température. Ces deux paramètres sont relativement bien maitrisés dans les bioréacteurs actuels. Un autre paramètre crucial lors d'une culture de micro-organismes est le niveau de substrat carboné disponible pour la croissance et/ou la production de métabolites. En effet, celui-ci conditionne directement la vitesse de réaction microbienne. Dans certains cas, les excès de source de carbone sont indésirables car ils entrainent la formation de sous-produits. C'est le cas par exemple de la levure S. cerevisiae, pour laquelle un excès en glucose entraine la formation d'éthanol en condition aérobie par un schéma réactionnel appelé effet glucose. Dans ce cas, le niveau de glucose doit être maintenu à un niveau très bas tout au long de la culture par un apport progressif de type fed-batch. La levure Y. lipolytica n'est pas sensible à l'effet glucose. Néanmoins, des résultats expérimentaux ont montré que la synthèse de lipases est induite lorsque la source de carbone hydrophobe devient limitante dans le milieu de culture (Fickers et al., 2004). Ce phénomène d'induction résulte d'une adaptation de la levure à une disponibilité très faible en substrat. Dans cette optique, une régulation fed-batch de l'apport en substrat peut être envisagée afin de maximiser le rendement en lipase. Une telle approche a été envisagée en séparant une phase de croissance de la biomasse sur glucose suivie par une phase de production de lipase sur méthyloléate (Turki et al., 2009 ; Turki et al., 2010). Un autre paramètre environnemental critique pour la production de lipase par Y. lipolytica est le taux d'oxygène dissous présent dans le volume réactionnel. En effet, Y. lipolytica est un micro-organisme aérobie strict et il a été montré que l'hypoxie a un effet négatif sur le rendement spécifique, mais aussi sur le niveau d'expression du gène lip2 responsable de la synthèse de la lipase extracellulaire (Kar et al., 2008). Malgré les nombreuses possibilités offertes au niveau de la régulation du taux d'oxygène dissous (modulation de la vitesse d'agitation et du débit d'air, enrichissement des gaz d'entrée en oxygène, etc.), aucune de ces stratégies n'a été envisagée de manière concrète pour la régulation du procédé de production de lipase.
4.2. La diminution de l'efficacité de mélange lors de l'extrapolation : cause majeure de perte de rendement des bioprocédés ?
27Les systèmes de régulation actuels sont basés sur des boucles de contrôle mettant en œuvre un senseur et un actuateur. Ces deux systèmes sont physiquement séparés au niveau du volume réactionnel. De plus, la zone de scrutation du senseur est limitée à une portion relativement faible du volume réactionnel. Pour des bioréacteurs de petit volume montrant un comportement dynamique proche du réacteur parfaitement agité, ces limitations n'ont pas de conséquences. Par contre, pour les bioréacteurs industriels, dont les temps de mélange peuvent atteindre plusieurs centaines de secondes (Delvigne et al., 2006), cette méthodologie de régulation pose des problèmes de temps mort entre la mesure du paramètre environnemental par le senseur et sa régulation par l'actuateur. Dans ce cas, une méthodologie de régulation de type « boucle ouverte », c'est-à-dire pour laquelle l'actuateur est actionné sur base d'une équation prédéfinie par l'opérateur, est préférée. Ces problèmes de transposition des systèmes de régulation à l'échelle industrielle ont un impact négatif sur la cinétique globale du procédé avec une réduction du rendement en biomasse et/ou en métabolites. D'un autre côté, plusieurs études ont montré que, outre la perte de rendement globale, l'apparition d'hétérogénéité au sein des réacteurs peut avoir des conséquences bénéfiques au niveau de la physiologie des micro-organismes (Patnaik, 2002). Une amélioration de la viabilité cellulaire a ainsi pu être mise en évidence lors du passage à l'échelle industrielle d'un procédé fed-batch de production de Escherichia coli (Hewitt et al., 2000).
5. Troisième niveau de dimensionnement des bioréacteurs : estimation du stress microbien face aux conditions de procédés
5.1. Stress environnementaux rencontrés dans les bioprocédés : nature, échelles spatiale et temporelle
28La notion de stress n'est pas bien définie au niveau des micro-organismes. Une notion généralement acceptée de stress microbien est une adaptation du métabolisme et de la cinétique de croissance suite à un changement environnemental (choc osmotique, thermique, manque en source de carbone, etc.). La capacité de réponse aux conditions stressantes est très importante à prendre en compte afin de prédire le comportement des souches de levures en bioréacteur. En effet, l'intensité des perturbations, mais aussi leur fréquence affectent la dynamique de la réponse microbienne face au stress (Reijenga et al., 2005). Les fluctuations en oxygène dissous, en substrat et en pH décrites dans le paragraphe précédent sont donc potentiellement impliquées dans la génération du stress microbien. La complexité du phénomène de réponse au stress réside dans la multiplicité des temps caractéristiques mis en jeu. Ceux-ci sont schématisés à la figure 3 par la cascade de réponse ADN, ARN, protéine. Cette figure montre que le promoteur de stress (Pstress) est induit suite à l'interaction avec un facteur de transcription (TA). Dans ce cas, la cascade ou le réseau de régulation qui entraine la synthèse de ce facteur de transcription a été ignorée par souci de simplicité. Le complexe TA_DNA formé est responsable de la synthèse des ARN messagers correspondant (processus de transcription). Ces ARN messagers permettent alors la synthèse d'une protéine (processus de traduction). Cette protéine peut être une enzyme intervenant dans le métabolisme, une chaperone ou encore un facteur de transcription. Plusieurs études ont montré que certains gènes pouvaient de cette manière être considérés comme indicateurs du stress encouru par les micro-organismes lors de la culture en bioréacteur (Schweder et al., 1999 ; Schweder et al., 2002 ; Schweder et al., 2004 ; Pioch et al., 2007).
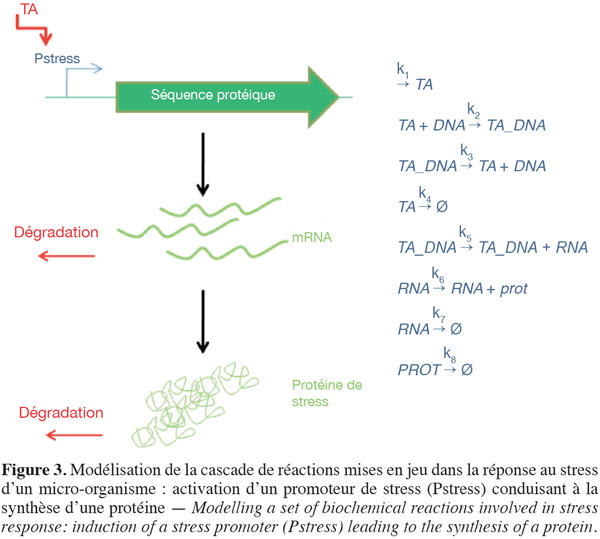
29Selon ce modèle simplifié, la synthèse de la protéine de stress est soumise à un schéma faisant intervenir huit réactions (caractérisées par le taux de réactions allant de k1 à k8) représentant la dynamique des cinq espèces biochimiques mises en jeu (TA, DNA, TA_DNA, RNA et GFP). La valeur de ces différents taux reflète bien la multiplicité des échelles de temps mises en jeu dans les processus physiologiques. À titre d'exemple, le temps de demi-vie d'une protéine très soluble dans le cytoplasme excède les 24 h, alors que celui de certains ARN messagers n'excède pas 1 à 2 min. La dynamique de réaction peut donc être représentée mathématiquement par un modèle comprenant cinq équations différentielles ordinaires :
30Ces équations peuvent être résolues numériquement par un algorithme adapté de manière à donner l'évolution de la concentration des cinq espèces au cours du temps. Néanmoins, il est maintenant évident que les processus biologiques à l'échelle cellulaire sont soumis à des phénomènes aléatoires ou stochastiques. Cette stochasticité peut être attribuée au nombre relativement restreint de molécules mises en jeu dans le volume cellulaire (McAdams et al., 1999 ; Patnaik, 2006). Dans ce cas, il est plus réaliste d'envisager un algorithme de résolution des équations différentielles qui prend en compte la partie aléatoire (ou stochasticité) des processus de synthèse. Nous avons utilisé pour cela une version adaptée de l'algorithme de Gillespie (Gillespie, 2001). Un exemple de simulation stochastique du modèle décrit à la figure 3 est montré à la figure 4.
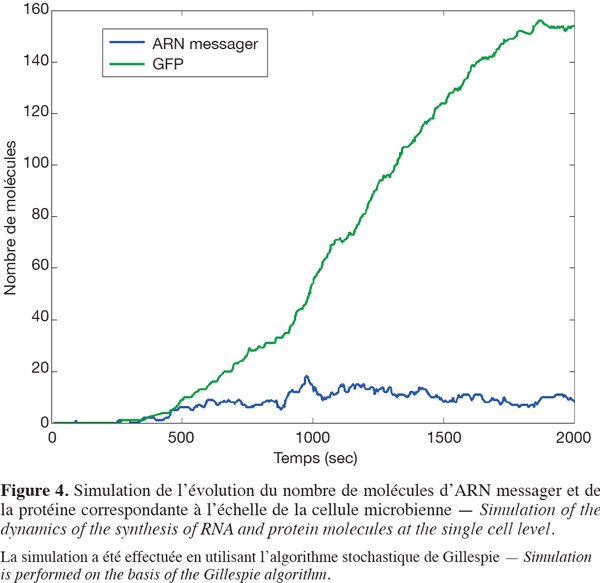
31La difficulté à ce niveau est que le bruit associé à la synthèse de la protéine fluorescente dépend non seulement des fluctuations encourues au sein du bioréacteur, mais également des fluctuations intrinsèques liées à des processus biologiques (phase du cycle cellulaire, mutation, etc.) (Müller et al., 2010). Une alternative est la mise en place d'un protocole expérimental permettant de déterminer ce bruit au niveau de la synthèse protéique. À l'heure actuelle, la technique de cytométrie en flux est de plus en plus utilisée pour l'analyse des systèmes microbiens (Diaz et al., 2010 ; Tracy et al., 2010).
5.2. Application de la cytométrie en flux pour le suivi des bioprocédés
32La cytométrie en flux est de plus en plus considérée comme une méthode permettant d'obtenir des informations sur l'évolution des processus physiologiques dans le cadre des cultures microbiennes (Diaz et al., 2010). Conçue à l'origine pour l'étude des cellules eucaryotes supérieures, cette méthode est à l'heure actuelle de plus en plus considérée pour l'étude des micro-organismes. Cette technique est basée sur la séparation des cellules microbiennes à partir d'un échantillon par un processus de focalisation hydrodynamique. Chaque cellule individuelle est alors orientée vers un laser d'excitation. La séparation des différentes composantes de la lumière réfléchie permet de déterminer de nombreux paramètres cytologiques tels que la taille des cellules, leur forme et leur fluorescence si celles-ci ont été colorées avec un fluorochrome. Néanmoins, peu d'études mettent en œuvre la cytométrie en flux dans le cas de Y. lipolytica. Une des rares études dans ce domaine met en œuvre la cytométrie en flux afin de déterminer l'adhésion de microgouttelettes de substrat hydrophobe à la surface des membranes cellules de Y. lipolytica (Aguedo et al., 2003). Une approche innovante consiste à utiliser des systèmes de gènes rapporteurs sous le contrôle de promoteurs associés à des stress particuliers rencontrés dans les procédés. De cette manière, lorsque la cellule microbienne est soumise à un stress, la synthèse de protéines fluorescentes est induite et celles-ci s'accumulent à l'intérieur de la cellule. Le couplage de cette technique avec la cytométrie en flux permet d'obtenir un signal proportionnel au stress à l'échelle de la cellule microbienne (Delvigne et al., 2009). Cette approche a récemment été appliquée au sein de notre équipe par la mise en œuvre d'un gène rapporteur de la synthèse de la lipase au niveau de Y. lipolytica (Kar et al., 2008).
6. Conclusion
33Nous avons vu au cours de cet article que la réactivité au sein du bioréacteur est contrôlée par les conditions physico-chimiques régnant au sein du volume réactionnel, mais aussi par la réponse physiologique des micro-organismes cultivés. Si les premières conditions sont relativement bien maitrisées à l'heure actuelle (existence de code de calcul de type mécanique des fluides numérique permettant de prédire les caractéristiques des écoulements dans le bioréacteur), il n'en va pas de même pour les secondes. En effet, la réponse microbienne face aux conditions de stress en procédé est complexe et fait intervenir de nombreuses échelles de temps. Afin d'effectuer une opération d'extrapolation, il est donc important de prendre en compte à la fois les paramètres du génie chimique pour le dimensionnement du bioréacteur, mais également de tenir compte des réponses physiologiques potentielles des micro-organismes cultivés. Ces réponses sont complexes et leur dynamique fait intervenir de nombreuses échelles temporelles et spatiales. La complexité des phénomènes de réponse physiologique peut à l'heure actuelle être caractérisée par la biologie systémique, c'est-à-dire l'étude de l'arrangement des gènes, protéines et métabolites en réseaux (génome, protéome et métabolome). À ce jour, de nombreux efforts ont été effectués pour comprendre et maitriser la réponse microbienne face aux conditions de procédé, notamment en intégrant les données de la biologie systémique (Deckwer et al., 2006 ; Wang et al., 2009) et une nouvelle discipline, la fluxomique (la maitrise des flux métaboliques), a même été développée dans ce contexte. Nous en sommes maintenant à considérer des modifications génétiques visant à diminuer la sensibilité des micro-organismes aux conditions de culture. Certains micro-organismes, tels que E. coli, ont même été « reconstruits » avec des simplifications conséquentes de leur génome afin d'éliminer leurs mécanismes de réponse au stress. Dans le cas de Yarrowia lipolytica, seules des délétions ponctuelles ont été envisagées au niveau des systèmes de synthèse de protéases afin d'améliorer le niveau d'activité de la lipase LIP2 retrouvé dans le surnageant de culture (Fickers et al., 2005b). Néanmoins, le séquençage complet du génome de ce micro-organisme devrait conduire sous peu à des remodelages plus conséquents du système de synthèse et d'excrétion de la lipase. Comme soulignés par Neubauer et al. (2010), il est curieux de noter que, malgré les progrès effectués au niveau de la compréhension des mécanismes biologiques, rien n'a été encore envisagé au niveau de la modification de la géométrie globale et des systèmes de régulation des bioréacteurs afin de mieux répondre aux contraintes imposées par la biologie des micro-organismes.
Bibliographie
Aguedo M. et al., 2003. Increased electron donor and electron acceptor characters enhance the adhesion between oil droplets and cells of Yarrowia lipolytica as evaluated by a new cytometric assay. J. Agric. Food Chem., 51, 3007-3011.
Beopoulos A. et al., 2009a. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res., 48, 375-387.
Beopoulos A., Chardot T. & Nicaud J.M., 2009b. Yarrowia lipolytica: a model and tool to understand the mechanisms implicated in lipid accumulation. Biochimie, 91, 692-696.
Charles M. & Wilson J., 2010. Encyclopedia of industrial biotechnology: bioprocess, bioseparation and cell technology. Vol. 7: Fermenter design. New York, USA: Wiley.
Deckwer W.D., Jahn D., Hempel D. & Zeng A.P., 2006. Systems biology approaches to bioprocess development. Eng. Life Sci., 6(5), 455-469.
Delvigne F., Destain J. & Thonart P., 2006. A methodology for the design of scale-down bioreactors by the use of mixing and circulation stochastic models. Biochem. Eng. J., 28(3), 256-268.
Delvigne F., Boxus M., Ingels S. & Thonart P., 2009. Bioreactor mixing efficiency modulates the activity of a prpoS::GFP reporter gene in E. coli. Microb. Cell Fact., 8, 15.
Delvigne F. & Lecomte J.-P., 2010. Encyclopedia on industrial biotechnology: bioprocess, bioseparation and cell technologies. Foam formation and control in bioreactors. New York, USA: Wiley.
Destain J., Roblain D. & Thonart P., 1997. Improvement of lipase production from Yarrowia lipolytica. Biotechnol. Lett., 19(2), 105-107.
Destain J. et al., 2005. Utilization of methyloleate in production of microbial lipase. Appl. Biochem. Biotechnol., 121-124, 269-277.
Diaz M., Herrero M., Garcia L.A. & Quiros C., 2010. Application of flow cytometry to industrial microbial bioprocesses. Biochem. Eng. J., 48, 385-407.
Dujon B. et al., 2004. Genome evolution in yeasts. Nature, 430, 35-44.
Etoc A., Delvigne F., Lecomte J.-P. & Thonart P., 2006. Foam control in fermentation bioprocess: from simple aeration tests to bioreactor. Appl. Biochem. Biotechnol., 129-132, 392-404.
Fickers P. et al., 2004. Carbon and nitrogen sources modulate lipase production in the yeast Yarrowia lipolytica. J. Appl. Microbiol., 96, 742-749.
Fickers P., Destain J. & Thonart P., 2005a. Methyl oleate modulates LIP2 expression in the lipolytic yeast Yarrowia lipolytica. Biotechnol. Lett., 27, 1751-1754.
Fickers P. et al., 2005b. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica and its potential applications. FEMS Yeast Res., 5, 527-543.
Fickers P. et al., 2006. Production and downstream processing of an extracellular lipase from the yeast Yarrowia lipolytica. Enzyme Microb. Technol., 38, 756-759.
Garcia-Ochoa F. & Gomez E., 2009. Bioreactor scale-up and oxygen transfer rate in microbial processes: an overview. Biotechnol. Adv., 27(2), 153-176.
Gillespie D.T., 2001. Approximate accelerated stochastic simulation of chemically reacting systems. J. Chem. Physics, 115(4), 1716-1733.
Herrera J.R. & Sentandreu R., 2002. Different effectors of dimorphism in Yarrowia lipolytica. Arch. Microbiol., 178, 477-483.
Hewitt C.J. et al., 2000. Studies related to the scale-up of high-cell-density E. coli fed-batch fermentations using multiparameter flow cytometry: effect of a changing microenvironment with respect to glucose and dissolved oxygen concentration. Biotechnol. Bioeng., 70(4), 381-390.
Hewitt C.J. & Nienow A.W., 2007. The scale-up of microbial batch and fed-batch fermentation processes. Adv. Appl. Microbiol., 62, 105-135.
Hoeks F. et al., 2003. Scale up of stirring as foam disruption (SAFD) to industrial scale. J. Ind. Microbiol. Biotechnol., 30(2), 118-128
Junker B.E., 2004. Scale-up methodologies for Escherichia coli and yeast fermentation processes. J. Biosci. Bioeng., 97(6), 347-364.
Kaneshina M. & Goto S., 2000. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acid Res., 28, 27-30.
Kar T. et al., 2008. Investigation of the effect of different extracellular factors on the lipase production by Yarrowia lipolityca on the basis of a scale-down approach. J. Ind. Microbiol. Biotechnol., 35, 1053-1059.
Lopes M. et al., 2008. Yarrowia lipolytica lipase production enhanced by increased air pressure. Lett. Appl. Microbiol., 46, 255-260.
McAdams H.H. & Arkin A., 1999. It's a noisy business. Genetic regulation at the nanomolar scale. Trends Genet., 15(2), 65-69.
Müller S., Harms H. & Bley T., 2010. Origin and analysis of microbial population heterogeneity in bioprocesses. Curr. Opin. Biotechnol., 21, 100-113.
Neubauer P. & Junne S., 2010. Scale-down simulators for metabolic analysis of large-scale bioprocesses. Curr. Opin. Biotechnol., 21, 114-121.
Nicaud J.M. et al., 2002. Protein expression and secretion in the yeast Yarrowia lipolytica. FEMS Yeast Res., 2, 371-379.
Nienow A.W., 1998. Hydrodynamics of stirred bioreactors. Appl. Mech. Rev., 51(1), 3-32.
Palomares L.A. & Ramirez O.T., 2009. Encyclopedia of industrial biotechnology: bioprocess, bioseparation and cell technology. Vol. 6. Geometric considerations for bioreactor scale-up. New York, USA: Wiley.
Patnaik P.R., 2002. Can imperfections help to improve bioreactor performance? Trends Biotechnol., 20(4), 135-137.
Patnaik P.R., 2006. External, extrinsic and intrinsic noise in cellular systems: analogies and implications for protein synthesis. Biotechnol. Mol. Biol. Rev., 1(4), 121-127.
Pereira-Meirelles F.V., Rocha-Leao M.H.M. & Sant'Anna G.L., 2000. Lipase location in Yarrowia lipolytica cells. Biotechnol. Lett., 22, 71-75.
Pioch D. et al., 2007. At-line monitoring of bioprocess-relevant marker genes. Eng. Life Sci., 7(4), 373-379.
Reijenga K.A., Baker B.M., van der Weijden C.C. & Westerhoff H.V., 2005. Training of yeast cell dynamics. FEBS J., 272, 1616-1624.
Schweder T. et al., 1999. Monitoring of genes that respond to process related stress in large-scale bioprocesses. Biotechnol. Bioeng., 65(2), 151-159.
Schweder T. et al., 2002. Role of the general stress response during strong overexpression of a heterologous gene in Escherichia coli. Appl. Microbiol. Biotechnol., 58, 330-337.
Schweder T. & Hecker M., 2004. Monitoring of stress responses. Adv. Biochem. Eng./Biotechnol., 89, 47-71.
Szabo R. & Stofanikova V., 2002. Presence of organic sources of nitrogen is critical for filament formation and pH-dependent morphogenesis in Yarrowia lipolytica. FEMS Microbiol. Lett., 206, 45-50.
Tracy B.P., Gaida S.M. & Papoutsakis E.T., 2010. Flow cytometry for bacteria: enabling metabolic engineering, synthetic biology and the elucidation of complex phenotypes. Curr. Opin. Biotechnol., 21, 85-99.
Turki S. et al., 2009. Isolation of bioactive peptides from tryptone that modulates lipase production in Yarrowia lipolytica. Bioresour. Technol., 100(10), 2724-2731.
Turki S. et al., 2010. An enhanced process for the production of a highly purified extracellular lipase in the non-conventional yeast Yarrowia lipolytica. Appl. Biochem. Biotechnol., 160(5), 1371-1385.
Van't Riet K. & van Sonsbeek H.M., 1992. Foaming, mass transfer and mixing: interrelations in large scale fermentors. In: Ladisch M.R. & Bose A., eds. Proceedings of the 9th International Biotechnology Symposium and Exposition, Harnessing biotechnology for the 21st century, August 16-21, 1992, Crystal City, Virginia. Washington, DC, USA: American Chemical Society, 189-192.
Vardar-Sukan F., 1998. Foaming: consequences, prevention and destruction. Biotechnol. Adv., 16(5/6), 913-948.
Varley J. et al., 2004. Dynamic multi-point measurement of foam behaviour for a continuous fermentation over a range of key process variables. Biochem. Eng. J., 20, 61-72.
Wang Y. et al., 2009. Industrial bioprocess control and optimization in the context of systems biotechnology. Biotechnol. Adv., 27, 989-995.
Yu M., Qin S. & Tan T., 2007. Purification and characterization of the extracellular lipase Lip2 from Yarrowia lipolytica. Process Biochem., 42, 384-391.
Zinjarde S.S., Pant A. & Deshpande M.V., 1998. Dimorphic transition in Yarrowia lipolytica isolated from oil-polluted sea water. Mycol. Res., 102(5), 553-558.
To cite this article
About: Tambi Kar
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Bio-Industries. CWBI. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : tambi.kar@student.ulg.ac.be
About: Frank Delvigne
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Bio-Industries. CWBI. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
About: Jacqueline Destain
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Bio-Industries. CWBI. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
About: Philippe Thonart
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Bio-Industries. CWBI. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
